
「種の保存のための進化」はどこが誤りなのか

「種の保存あるいは種族維持のために生物は進化した」という俗説が、未だに一般的に広く語られています。進化生物学のなかでは、この考えは受け入れられていません。本稿は、この俗説のどこが正しくないのか、について解説しました。
補足として、生物学者でも誤解が多い「生物の死の進化」と「有性生殖の進化」についての簡単な解説も載せました。
本記事を修正・加筆した記事が、以下の新書の第3章に収録されています。
レミングの集団自殺?
レミングとは、主にツンドラ地域に生息するネズミの仲間で、3年から4年周期で個体数が急激に増減することが知られている。特に、レミングイヤーと呼ばれる年には、その数は激増し、集団移動をすることがある。この集団移動の時に、多くの個体が海に飛び込み「集団自殺」をするという"迷信"が広まった。この迷信の原因の一つが、1958年に制作された「白い荒野」というディズニーの映画である(ムービー1)。この映画では、レミングが崖から飛び込んで集団自殺する行動が写されている(以下映像のムービーの1分30秒あたりでレミングが海に身をなげる)。
ムービー1. ディズニーの映画「White Wilderness(白い荒野)」のディズニーの映画。
Wood(1) の記事によると、この映画はいわゆる「やらせ」で、レミングが海に飛び込むシーンは、ディズニーの映画制作者たちに投げ入れられて撮影されたものであるという。さらに、「レミングが人口過剰を緩和するために7年から10年ごとに自殺すること」を暗示するナレーションがつけられている(1)。つまり、レミングは個体数が激増すると、食べ物や適した生息地が不足し、集団(あるいは種)が絶滅するのを避けるために「自殺」をするのだ、と多くの人を洗脳させる映像となっている。この映画は「集団自殺」と「集団の維持のための行動」という2つの迷信を広めたのである。
現在でも信じられている「種の保存のための進化」
進化についての情報が配信されるメーリングリスト"EVOLVE"において、新聞の記事に「種の保存のため、生まれつき生物に備わった習性か?」というような表現されていたことに対する意見投稿があった。そこでは「種の維持あるいは種の保存のために生物は進化した」という"間違った考え"が、一般の人たちの間で未だに普及している、という危惧が指摘された。また、一連の投稿の中で、「自分と同じ種族を保とうとする性質がある」と認識を有している大学生が半数以上いる、という調査結果(2)が紹介された。
しかし、これを「当然の正しいこと」と考えているのは、一般の人に限ったことではない。たとえば、脳科学者の中野信子さんは著書の中で以下のように考察している(3)。人のお産は死ぬリスクが高い。しかし、女性がそれを避けてしまうと「種としての人間が絶える」ので、リスクを冒しても産むように進化している。また、生物学者?の福岡伸一さんは、対談の中で「種の保存こそが生命にとって最大の目的なので、個は一種のツールにすぎません」と述べている(3)。「昆虫や魚類では、数千個の卵を生んで、そのうちのわずか数匹が子孫を残す、なんてことがざらにあります。でも、それで種が保存されるなら構わない。それが基本的な[生命の掟]なのです。」のように、福岡さんは、種の保存が生物の最大の目的であるかのような主張をしている(4)。
さらに、生物学の分野では業績も知識もある研究者の中にも、進化学において議論され検証されてきた理論や事実を理解していない人は少なくない。自分の専門分野の知識をもとに進化についてまで踏み込んで考察するときには「種の維持のための進化」を持ち出すことがある。たとえば、細胞老化やがん化の機構を研究している小林武彦さんは『生物はなぜ死ぬのか』という一般向けの本の中で、老化についてわかりやすい解説をしており、一般読者や出版社に好評を得ている(新書大賞2022第2位)。しかし、「なぜ生物が死ぬように進化したのか」という原因として、(i)個体数が増えすぎて、集団が絶滅しないため、(ii)死ぬことで、進化の材料となる多様性を確保する、という理由を挙げている。そして、「親は死んで子どもが生き残ったほうが、種を維持する戦略として正しい」と主張しているのである(5)。生物はなぜ死ぬように進化してきたのかについては、すべてが解決されていないものの幾つかの説が出され議論されている(記事「老化の進化:なぜ老化しない生物がいるのか? 」参照)。小林さんの著書では、これまでの「死の進化」についての議論が全く触れられておらず、一般読者に大きな誤解を与える内容となっている。
個体数が増えすぎて集団が絶滅しないように進化?
生物は個体数が増えすぎないように自己調節している、と言われることがある。個体数が増えると餌や生息場所といった資源が減少するために、個体の生存率や繁殖率が下がり、個体数が減る。逆に、個体数が減少すると利用できる資源が増えて、個体数が増える。このような密度依存的な個体数の変動を、自己調節という場合がある。しかし、個体数が増加しすぎると、資源が枯渇し集団が絶滅するのを避けるために、個体は自ら繁殖を抑制するように進化している、という考えは正しくない。
この考えは、まさに冒頭のディズニー映画のレミングの集団自殺の解説でもある。1960年代までは、このような考えは、生物学者の中でも暗黙に受け入れられていたようである。それが、明確な形で解説されたのが、Wynne-Edwards (11) による『社会行動に関する動物の分散』という本である。その本の中で、彼は、動物の分散移動行動は、動物の密度が増えすぎて、集団が絶滅しないための性質であるという説明をした。これが一つの切っ掛けともなり、この考えの妥当性が明示的に議論されるようになった。特に、Williams (12)が、『適応と自然選択』という本を出版し、そのなかで、生物の適応を説明するには個体と遺伝子の選択を考えるだけでよいとした(2022年春、翻訳本が出版予定)。また、自分を犠牲にして他個体を助けるという「利他行動」の進化が、利他行動に関わる遺伝子を血縁者が残すことで進化するという血縁選択説がHamilton (13)によって提唱された。(「適応」という用語についての議論は「進化における「適応」という言葉をめぐって」を参照)
その後、様々な議論がなされ、現在の進化学での一般的な理解は、集団にとってはプラス(集団の維持や保存)に働くが、個体の生存や繁殖にはマイナスに働く性質が、集団にとって有利だということが原因で進化することは少ないと理解されている(後でも述べるように、ここでの集団は種ではない)。ただし、特定の条件が満たせば可能な場合もある。実際に、そのようなケースがどれくらいあるのかについては現在でも議論が続いている。現在、世界で広く用いられている進化学の教科書でも同様の解説がなされている(たとえば14, 15) 。
集団内で変異の増減による進化
進化は、ゲノム上で生じた変異(ゲノム上の変異: 記事「進化的視点からみる人間の「多様性の意味と尊重」」の図2参照)が、集団中で頻度を変化させることで生じる(ここでの集団とは、有性生殖集団では、実際にお互いに遺伝子の交流のある個体の集まり、無性生殖生物ではお互い相互作用している同じ遺伝的系列の個体の集まり)。たとえば、濃い青色の蝶の集団に、突然変異によって明るい青色の蝶が生じたとしよう(図1)。明るい青色の蝶は、濃い青色の蝶に比べて捕食者に見つかりづらくなり、生存率が向上したとする。それにより蝶の色を変える遺伝的変異は集団中に広がり、何世代かかけて、集団の個体は全て明るい色の蝶となる(図1)。結果として、個体の生存に有利なった明るい青色の個体が自然選択を受けて広がったと表現する。そして、明るい青色は個体にとっての適応的な性質であるといえる。
あるいは、蝶の色は、蝶の繁殖や生存には影響を与えない場合もあるだろう。その場合、明るい青色に変化させる遺伝子が、ランダムに次世代に選ばれることで、集団中の個体がすべて明るい青色になることがある(これを遺伝的浮動という;注1)。このような場合、「個体の生存や繁殖に中立な変異が遺伝的浮動で進化した」という。特に集団の個体数が少ないときは、個体の生存や繁殖に有害となる変異が遺伝的浮動によって頻度を増加させ、進化することがある。
図1では、単純化して示したが、生物の新たな性質が獲得される進化は、集団内で生じた遺伝的変異(ゲノム上の変異)が頻度を増加させることで生じるのである。多数の遺伝的変異によって創り出される複雑な生物の性質も、多数の遺伝的変異が、それぞれ別々に集団中で頻度を増加させることで獲得される。

注1; たとえば、血液型AB型の母親と父親から生まれた子どもは、AA型、AB型、BB型のいずれかになる。AあるいはBという遺伝子のどちらが子どもに伝わるかは偶然によるので、BB型だった場合、Aの遺伝子が全く伝わらなかったことになる。このようなランダムに選ばれる効果が、集団全体の遺伝子の頻度を変化させていくのである。
集団存続のための個体の進化
それでは、「個体の生存や繁殖に不利になっても集団の維持や保存に貢献する性質が進化する」という考えのどこが問題なのだろうか? 今度は、濃い青色の個体は、個体数が増えすぎないように自ら繁殖を抑制している性質を持っているとしよう(図2)。繁殖を抑制しない個体(明るい青色)は、個体数を自己抑制することはないので集団は増大し、資源が枯渇すると個体数は激減したり、たまには絶滅するかもしれない。しかし、個体数を自己抑制する濃い青色個体の集団に、移入や突然変異によって、繁殖を抑制しない明るい青色個体が侵入する状況は普通に想定される。そのような状況では、繁殖を自己抑制する濃い青色の個体に比べて、明るい青色の個体はより多くの子どもを残すことができるために、その集団は自己抑制しない個体ばかりになってしまう。つまり、集団維持のために自己抑制する個体は、そうでない個体にすぐに置き換わってしまい、簡単には進化することができないのである。
この記事のトップ画像は、Mark Godfrey氏のレミングの集団自殺を風刺した漫画である。一匹だけパラシュートで助かろうとする個体がいる。これは、自殺を避けて助かろうとする個体がいると、他の個体は自殺で減ってしまうので、すぐに集団中に広がっていくということを分かりやすく説明している。(ネズミ類における繁殖抑制の進化についての解説はここを参照)

では、どのような場合に、「個体の繁殖にはマイナスだが集団の維持や保存にはプラスに働く性質が進化する」可能性があるのだろうか?今度は図3で説明してみよう。濃い青色の個体は、自分を犠牲にして「協力的な行動」をとる。それに対して明るい青色の個体は競争的で、他個体を攻撃して餌を確保しようとする「攻撃的な行動」を示す。攻撃的な個体は、協力的な個体が餌を採るのを攻撃するので、同じ集団内では、協力的な個体の頻度は減少する。しかし、協力的な個体の多い集団では、お互い協力して餌を採るので、集団全体として多くの餌を得ることができる。そのため、攻撃的な個体の多い集団に比べて、個体数を増やすことができる。もし攻撃的な個体ばかりになった集団は、その攻撃性のために集団全体としての餌を多く採ることができず、集団のサイズが減少し、絶滅する。絶滅の後に、個体数の増えた協力的な個体の多い集団から、協力的個体が移住して、新たな集団を形成する。このようなプロセスが起こると、協力的な個体が進化する。このようなプロセスを集団選択とよぶ(厳密にはデーム間集団選択とよぶ。デーム内集団選択というプロセスもあるがここでは省略する)(16)。

このように、個体にとっては不利であるが、集団にとって利益となる個体の性質が進化することは理論的には可能である。しかし、このプロセスが働くためには、特定の条件が満たされていなければいけない。一つは、集団内では攻撃的な個体が頻度を増加させて、ほとんどの集団が攻撃的な個体になる前に、協力的な個体の頻度の高い集団が個体数を増加させて、攻撃的な個体の集団が絶滅した後に、新たな協力的な集団を形成させる必要がある。このような現象が実際に生じるのは、頻繁な絶滅や集団形成が起こるコロニーのような小さな集団であろう。また、図3の左の最初の集団をみてほしい。集団選択が生じるためには、集団間で協力的個体の頻度が多い集団と少ない集団が最初から存在する必要がある(集団間の変異) 。この状況が生じるためにも、集団の個体数が小さい必要がある。
実際の例が、アミメアリで示されている(17, 17a)。アミメアリは、雌アリ (女王) をもたず,働きアリが産卵して働きアリを生むことでコロニーが維持される。コロニーの中には、大型の個体と小型の個体がいる。大型個体は繁殖能力は高いが餌をとるのに協力的でないのに対し、小型個体は餌集めをするが、個体としての繁殖は大型に劣る。小型個体は、コロニー間の選択によって維持されている(記事「人はなぜ宗教を信じるように進化したのか」の注2も参照)。
集団選択には複数の異なる様式があるが、どれも限定付き条件のもとで理論的に可能ということである。実際に、アミメアリのように、野外研究で実証データを基に示された集団選択の例は少ない。たとえば、集団で餌を探すクモにおいて、集団選択が働いたとされる研究がNature誌に載るほど現実に実証されていないということである(なお、このクモの論文[リンク]の著者の一人であるPruittは、一連の研究に対してデータねつ造が疑われた。そのため、この論文の信憑性は疑われている)。
ところで、協力行動の進化は、上述した血縁選択説や互恵利他による進化など、集団選択を考慮しない理論によって説明されることが多い。しかし、人の大規模な協力行動がなぜ進化したのかについては未解決な問題である(記事「進化的視点からみる人間の「多様性の意味と尊重」」を参照)
種の保存
ここまで集団の絶滅・再形成について見てきた。ここでの集団が"種"という集団である場合は、集団選択は起こるだろうか。つまり「種にとって有利な性質が進化する」ことが可能かどうか、ということだ。
種とは何だろうか。実は、種の定義は様々である。たとえば、お互いが潜在的に繁殖可能な集団の集まり(生物学的種概念)とか、同じ子孫集団をもつ集団の集まり(系統的種概念)など、20を越える種の定義があり、さらに増える傾向にある(18)。生物の集団のあり方は多様であり、生物集団を単一の概念で定義できないので、生物学者や分類学者は様々な定義を用いるのである。
その中でも単純化した例でみてみよう(図4)。生物学的種、系統学的種、形態学的種という3つの異なる定義で種を認識すると、それぞれは一致しないことが多い。また、一般的に生物学では、種の定義として生物学的種概念が用いられることが多いが、この種概念ではどこまでが同じ種であるかが曖昧となる生物は多い。別の種として記載されている種の間で交配が生じ、子どもが出来るという現象は一般的である。たとえば、我々、ホモ・サピエンス(Homo sapiens)は一つの種として定義されている。しかし、過去には別種として定義されていたネアンデルタール人(Homo neanderthalensis)と混血して、彼らのゲノムの一部を引き継いでいる(記事「ヒトはいつ出現し、どう進化をたどってきたのか」を参照)。
定義の問題は別にしても、図3のように、実際に絶滅したり、分岐したり、再形成したりしている集団(分集団、地域集団、メンデル集団、デームなど)は、「集団の括り方」によって定義された種ではない。集団にとって利益になる性質が集団選択によって進化することがあったとしても、その集団とは種ではないことがわかる。そのような意味からも「種の保存や維持にとって有利な性質が進化する」ということはないのである。

「種の保存」という言葉は、種の保全活動の現場でも用いられる。しかし、ここでも保全の対象とすべき単位は種ではない。たとえば、西表島に生息するイリオモテヤマネコや対馬に生息するツシマヤマネコは、保全すべき対象と考えるだろう。しかし、イリオモテヤマネコやツシマヤマネコは、大陸に生息するベンガルヤマネコ(Prionailurus bengalensis)の亜種である。同じ種が大陸に生息しているので、イリオモテヤマネコは保全する必要がない、とはならないのではないだろうか。この場合、保全すべき対象は、地域集団であり種ではない。
また、アメリカ合衆国ではアカオオカミ(red wolf)の個体数が減少しているため、保全すべきかどうかが問題となった。しかし、遺伝的な研究の結果、アカオオカミはハイイロオオカミとコヨーテの雑種であった(19) 。純粋な種(生物学的種)ではないので、保全する必要はないという考えと、独自の遺伝的組成をもった集団であるという考えが対立した。保全生物学の分野では、保全すべきかどうかの単位は種ではなく、進化的に重要な単位(独自の区別できる遺伝的組成をもつ単位)とすべきであるとする考えもある(20)。
系統選択
地球上で生じた生物種のほとんどは、現在までに絶滅しているといわれている。「現在まで生き延びることのできた種は絶滅を回避する性質を備えている」と考えるのは間違いではない。どのような性質をもった集団や種が絶滅の危険性が高いのか、という議論は重要である。しかし、「絶滅を回避する性質」が進化したのは、種という集団が絶滅しないことが原因で進化したわけではない、ということを理解する必要がある。
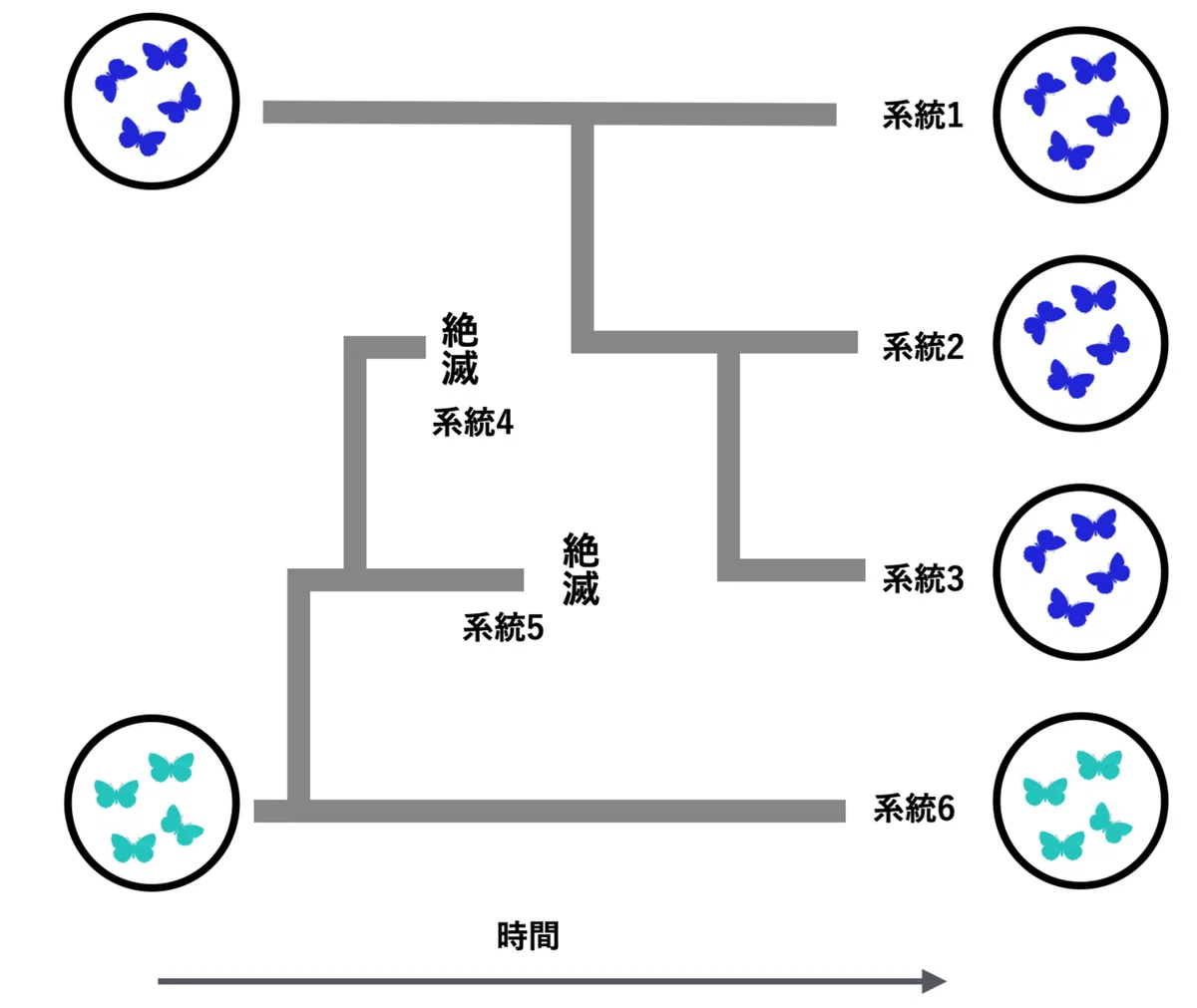
4つの集団(図では系統、つまり祖先子孫関係で連なっている集団)が現在存続している場合を考えてみよう(図5)。明るい青色個体の占める集団は、濃い青色の個体が占める集団に比べて絶滅リスクが高いために、現在、濃い青色の集団が多く生き残っている。これは、濃い青色個体の持つ性質が、絶滅リスク低減に寄与しているからである。
この例のように、異なる系統間(濃い青色個体集団の系統と明るい青色個体集団の系統)を比較すると、一方の系統の方が、他の系統に比べて絶滅率が低く、より多くの系統(集団)が生き延びている。このように、系統間の絶滅率や分岐率の差によって「系統間の性質の頻度の違い」を生じさせるプロセスを系統選択(あるいは種選択)と呼ぶ(21)。
図5では、集団の中の個体はすべて同じ性質を持っている(すべて濃い青色個体かすべてが明るい青色個体)。濃い青色個体が進化したのは、この図で示したより祖先において、集団の中で濃い青色個体の持つ性質が生存や繁殖に有利に働いた(個体への自然選択)か、あるいは、偶然の効果で濃い青色個体が増えた(遺伝的浮動)ことが原因である。その後、突然変異や系統間の交雑によって濃い青色個体と明るい青色個体が混在することがない状況を示している。
両者が混在する状況がしばしば生じるときを考えてみよう。濃い青色個体(絶滅リスクを低下させる性質をもつ)が集団内有利な選択を受け頻度を増加させる場合は、明るい青色個体の集団は濃い青色個体で置き換わってしまう。逆に、明るい青色個体が有利な選択を受け頻度を増加させるときはどうだろう。濃い青色個体集団が、集団内では不利にもかかわらず、集団の絶滅をリスクを減少させることで進化するためには、図3で示したような集団選択と同じプロセスが働かなければいけない。
つまり、図5のような系統選択がみられる場合、集団(系統)の絶滅リスクを低減する性質は、集団内での個体の性質が進化する結果生じたと考えることができる。そして、集団の絶滅率の低下は、多くの集団が「絶滅リスクを低減する性質」を保持している現象(系統間の性質の頻度の違い)の原因とはなっているが、「絶滅リスクを低減する性質」がいかに集団内で獲得され進化したかの原因とはなっていない。言い換えると、「個体の生存を向上させる性質は、それ自身が原因となって集団内で進化する」ことは可能だが、「集団の生存率を向上させる性質は、集団の存続が原因で集団内で進化すること」は限定的である。
ただし、集団選択のプロセスが働くような特定の場合は、集団の絶滅や分岐・再形成というプロセスが、「絶滅リスクを低減する性質」を進化させることが理論的には可能である。また、集団内で、有利でも不利でもない中立な性質が偶然進化した場合や個体にとって有利な性質が進化した結果、その性質が集団絶滅リスクを低下させている場合、その性質をもっている集団はより存続を可能しているといえる。この場合は、その性質が進化した原因は集団内の進化現象であるが、同時に系統選択が働いているといえる。
進化の主要なプロセスは集団内で生じるプロセスであり、特に、生物の様々な形質の進化的獲得のほとんどは集団内で生じた変異が頻度を増加させることで生じる。冒頭で述べた遺伝学者の小林さんは、「多様な「種のプール」があって、それらのほとんどが絶滅、つまり死んでくれたおかげで、たまたま生き残った「生き残り」が進化という形で残っているだけです」と述べている(4)。しかし、この記述が正しいとしても、たまたま生き残った種がなぜ個体が様々な性質を進化させたのかは、この記述では説明できない。
ところで、ここで紹介した系統選択のプロセスを種選択と呼ぶことがある。何度も指摘しているように、実質的にプロセスに関与しているのは集団あるいは系統であって、種ではない。種選択と呼ぶことがあるのは、この概念が化石の種の観察から提唱されたからである(21)。しかし、化石の種は形態で認識される形態種であるが、形態だけからは集団とか系統が正確に認識できない。そのため、種が便宜的に使われているのである。
この系統選択の例として、しばしば引用されるのが、有性生殖系統と無性生殖系統の違いである。現在、多くの動植物において、無性生殖をする系統が存在している。しかし、一部を除いて、無性生殖系統は最近生じた若い系統の場合が多いという(22)。大昔に生まれた無性生殖系統がほとんど残っていないことは、無性生殖系統は有性生殖系統よりも絶滅率が高いことを示唆している(22)。つまり、有性生殖系統が生物界で広く見られるのは、無性生殖系統は絶滅率が高いからだというのである。
有性生殖がなぜ進化したかについては、古くから議論がなされてきた。有性生殖する個体は、無性生殖する個体に比べて、2倍のコストがあるといわれている。有性生殖する集団の中に、無性生殖する個体が突然変異で生じたとしよう。有性生殖個体では、一個体の子どもをつくるのにオスとメスの2個体が必要だが、無性生殖では一個体の親で充分である。つまり、一個体あたり残せる子どもの数は、無性生殖個体は有性生殖個体の2倍となる。この2倍のコストを乗り越えて、有性生殖個体は進化しなければいけない。有性生殖するか、無性生殖をするかだけを変えてシミュレーションしてみると、有性生殖個体の集団に無性生殖個体が生じるとすぐに無性生殖個体に置き換わってしまうし、逆に無性生殖個体の集団に有性生殖個体が生じても、集団中に頻度を上げることができずすぐに淘汰されてしまう。
無性生殖集団内で有性生殖個体が生じ、無性生殖個体に勝って、なぜ進化できるのかについてはBox1で簡単に説明した。Box1で紹介した説は、有性生殖をする集団や種がどう有利になるかどうかではなく、有性生殖する個体が無性生殖個体に比べてなぜ有利になるのか、ということを説明している。集団内で、無性生殖に勝って有性生殖が進化した結果として、有性生殖の集団では無性生殖集団に比べて絶滅しづらい性質も進化したといえる。また、無性生殖個体が有性生殖個体に比べて不利となる原因の一つとして、有害遺伝子を効率よく除去できないという要因がある。これは、無性生殖個体にとって不利となるだけでなく、有害遺伝子が集団中にも蓄積していくことから、無性生殖集団にとっても不利となり、絶滅率を上昇させる要因となっている。有性生殖が進化したのは、無性生殖集団よりも絶滅率が低かったからではない。しかし、有性生殖集団が多くの生物でみられる原因には、集団内での有性生殖個体の有利性だけでなく、無性生殖集団の絶滅率の高さが原因となっているのだろう。これは、集団内の自然選択のプロセスと集団間で働く選択のプロセスの両者が働いていると考えられる。
Box 1 有性生殖の進化
有性生殖が個体にもたらす有利性として、よく語られる議論がオスとメスの遺伝子が組み合わさることによって遺伝的な変異が増えるというものである。有性生殖によって、一匹のメスから生まれる個体の間の遺伝的な多様性は、高くなることも低くなることもある。また、子どもの中の多様性が高いほど進化的に有利だと考えるかもしれないが、多様性があるということは、生存率の高い子どももいれば、悪い子どももいるということになる。そのため、メスから生まれる子どもを平均すると、生存率は必ずしも高くはならない。また、オスとメスの遺伝子を組み合わせることで適応的な個体が生じたとしても、その子どもが再び有性生殖をすることで、その組み合わせは崩れてしまう。従って、単純に、オスとメスの遺伝子が組み合わさることによる遺伝的な変異の増加が有性生殖の有利性になるとはいえない(23, 23a)。
ただし、環境が変動する場合、オスとメスの組み合わせで常に作りだされる新しい変異が有利になる場合がある。たとえば、集団中で頻度の高い遺伝型に病原体が感染しやすいとき、新しい希な遺伝型は感染から逃れることができる。有性生殖によって新たな変異つくりだすことで、常に進化する病原体に対抗できるという説である。これは赤の女王仮説といわれている(24)。
有性生殖が有利になる有力な説として、生存や繁殖を向上させる遺伝的変異同士の競争を避ける効果がある。図6で説明しよう。二つの遺伝子座(ゲノム上の異なる場所にある遺伝子)があり、Aはaに比べて、Bはbに比べて生存率を向上させる。しかし、AはBよりも生存率を向上させる効果が高いため、Bは有利な効果があるにもかかわらず、無性生殖では集団中の個体はすべてAAbbとなり、aaBBは淘汰される。しかし、有性生殖による組換えが生じると、AとBを同時にもつ個体が生じるために、AもBも有利になりAABBが集団中に増えていくことが可能になる。この効果をFisher-Muller効果という。この効果により生物は、適応的な進化を促進させることができる(25, 実証研究は26, 26aなど)。
同様に、有利な遺伝子がより有害な遺伝子と組み合わさったとき、無性生殖では、有利な遺伝子は有害な遺伝子といっしょに淘汰されてしまう。これをruby in the rubbish効果(ルビーといっしょにごみ箱へ)という(27)。有性生殖では、組換えによって、この有利な遺伝子を広めることが可能になる。
上の2つの効果は、有利な遺伝子の組み合わせが組換えによって生じることによる有性生殖個体の利益である。同様に, 有害な遺伝子を組換えによって
除去することが可能になる。たとえば図6でbは生存率を低下させる有害な遺伝子だとしよう。無性生殖個体では、有害な変異bは、有利な変異Aといっしょになっているためにbを取り除くことができない 。 有性生殖個体では、有害遺伝子が蓄積して大きな有害効果を個体に対して示さないように、有害遺伝子を取り除くことができる。また、長期的にみたとき無性生殖個体は、取り除けない有害遺伝子が蓄積していくことで、生存できなくなる(28) 。
現在、赤の女王仮説によって性が維持される条件は限られていることが指摘されている(24a)。赤の女王仮説による病原体と宿主の共進化、Fisher-Muller効果、ruby in the rubbish効果、有害遺伝子の蓄積の効果が組み合わさって維持されているのではないかと考えられている(24) .
また、無性生殖集団では、ゲノム中に有害な遺伝子が突然変異で生じると、強い有害効果を示さない限り、その有害遺伝子は、除去されないまま、集団全体の個体に維持される(Mullerのラチェットという)。つまり、無性生殖集団は、個体レベルだけでなく、集団レベルでも有性生殖より不利になるといえる。

個体の進化の副次的な影響としての集団の存続と絶滅
ここまで、述べてきたように、集団の存続や絶滅に影響するような生物の性質は、多くの場合、個体が集団内で進化した結果といえる。集団内では、自然選択によって個体の生存や繁殖を向上させる性質が進化するか、あるいは、遺伝的浮動によって生存や繁殖の違いに関わらない性質あるいは低下させる性質が進化する(生存や繁殖を低下させる進化については記事「人類の進化史と病気の進化」を参照)。その結果として、集団(あるいは種)の存続を高める場合もあれば、低下させる場合もある。 たとえば、有性生殖個体は、無性生殖個体に比べて、有利な効果をもつために進化した。その結果として集団の絶滅率を低下させている。
個体の生存や繁殖を向上させるような性質が進化した結果、それらが集団の増殖率の向上や絶滅率の低下を引き起こしている例は多く見られる。たとえば、集団の中に、個体に働く自然選択によって、集団の中に、行動や色の違いなどが異なる個体が維持される場合がある。例えば、イトトンボでは、メスの間に2つの体色の異なるタイプ(色彩多型)が存在している集団がみられる。この色彩多型は個体に働く自然選択で維持されているが、色彩多型が維持されている集団は、多型を示さない集団に比べて、集団の増殖率が高い(29) 。また、イトトンボだけでなく、色彩多型を示す種は、示さない種に比べて、生息域が広く、絶滅率が低いことが示されている(30) 。
逆に、個体の生存や繁殖の違いに影響する性質が進化することで、集団のサイズが低下する場合もある。例えば、オサムシという甲虫の仲間では、オスは大きな交尾器をもつ。オスにとっては、交尾器が大きいほど繁殖の成功率を高める。しかし、メスにとっては、オスの大きな交尾器で生殖器を傷つけられる可能性が高くなる。そのため、大きな交尾器をもつオスの集団ほど、個体数が減少しているという効果がみられる(31)。
また、集団内の個体の間での競争が激しいとき、競争に強い個体が進化する結果として、集団のサイズや増加率は減少する場合がある。しかし、その結果として異なる種が共存することが促進される可能性も示されている(32)。この例も、種がお互いに共存するように進化が生じたわけでなく、適応的な個体が集団内に出現した結果、種の共存が促進されるといえる。同様に、生物多様性の高くなるほど、群集が安定する場合がある。これも群集全体が安定するように進化したわけではなく、群集を構成する生物集団内で進化が生じた結果である。理論的に個々の要素が適応的に進化することで、全体の頑健性や安定性が創り出される条件が示されている(35)。
もちろん集団の間でみられる絶滅率の差が原因となって、生物群集内での種構成を変化させたり、単系統(同じ祖先をもつ系統の集まり)の中で、特定の性質をもつ集団や種が多く観察されたりする。何度も述べたが、種の絶滅や生存は、その種が示す性質を進化させた原因とはならない。
ところで、リチャード・ドーキンスの『利己的遺伝子』(33)を読んだことのある人は、個体の生存や繁殖を向上させる性質が進化するのではなく、遺伝子のコピーが増え、引き継がれるような性質が進化するのではないか?と思うかもしれない。「利己的遺伝子」は理論ではなく、どのように進化をみるとわかりやすか、という見方である。そのような見方が招く誤解については別の記事にまとめた(「利己的遺伝子」の誤解を招かない使い方)。
なぜ種の利益が想起されるのか?
進化学についての基礎知識を持っていない人は、「生物進化が、種の保存のために重要」となぜ考えてしまうのだろうか?人間は、様々な特徴に基づいて、生物を含めた様々な個物をカテゴリーに分類し、一般化した手がかりをもとにステレオタイプ化してものごとを分類しようとする傾向がある(34)。つまり、人には、生物学的な実態の不明確なものを意味のある集団として認識する認知バイアスがあるのだろう。また、人は、他者を「集団内」または「集団外」に分類し、集団内のメンバーにとって有利になるような行動をとる傾向にある(34)。このような人のもつ認知バイアスにより、「生物も種や集団のために行動する」と、つい考えてしまうのかもしれない。
「生物の個体は死ぬことで、新たな個体を生み出し、常に入れ替わっており、それにより種という調和した存在を保っている(動的平衡?)」という一見響きの良い哲学的言説に人々は惹かれることがある。また、「生物学者が専門家以外の聴衆に話しかけるときよくあるように、主として意識的に哲学的になる」とWilliams は指摘している(12)。そのため、進化学の一般教養のない生物学者が、進化について語るときは妄想的な哲学議論に陥りやすいのかもしれない。
一般の人々に誤解が多いのは、我々進化学を専門とする研究者の情報発信不足が原因の一つかもしれない。しかし、少なくとも生物学を専門とする研究者は、正しい情報を一般の人に伝えてほしい。
謝辞
公開前の原稿について、琉球大学の辻和希さん、東京大学の石川麻乃さん 、東北大学の香川幸太郎さんに有益なコメントを頂いた。
以下の記事もご一読ください
「利己的遺伝子」の誤解を招かない使い方
進化における「適応」という言葉をめぐって
古代人・現代人ゲノムから探る人の「こころ」の性質の進化
ビール酵母の進化とビールの多様化
さらに詳しく知りたい方へ
自然選択の単位とレベルについて、『講座 進化 1 進化とは』東京大学出版会の第2章で解説しています。より専門的な解説を知りたい方は、以下をダウンロードしてください。(専門的解説)
やや専門的な解説書。行動生態学を学ぼうとする人向け
本文でも紹介した、G.C.Williamsの古典的名著。集団選択を明確に批判した本。辻和希さんによって翻訳されました。
ジンマー,K ・D.J. エムレン [更科功・石川牧子・国友良樹訳] 『進化の教科書 第3巻』 講談社ブルーバックス (第13章の行動の進化を参照)
河田雅圭『はじめての進化論』講談社現代新書. (集団選択の理論の章でも簡単にレミングのことを触れています)。以下からダウンロードできます
シリーズ進化学 6 行動・生態の進化. 岩波書店
河田 雅圭・斎藤 隆 「エゾヤチネズミの社会構造と繁殖抑制」『サイエンス』1987年10月号. 繁殖抑制のメカニズムについての実証的解説。集団選択についても解説しています。『サイエンス』は1990年から『日経サイエンス』に誌名を変更しています。(日経サイエンス社の許可の上掲載しています)[このリンク先からダウンロード]
「生物はなぜ死ぬのか」についての良質な解説 第2章「生・老・死の生物学」を参照。
引用文献
1. Woodford, R. (2003) Lemming Suicide Myth Disney Film Faked Bogus Behavior. Alaska Fish & Wildlife News. http://www.adfg.alaska.gov/index.cfm?adfg=wildlifenews.view_article&articles_id=56
2. 山野井貴浩, 佐藤綾, 古屋康則 (2018) 大学生対象の「種族維持」概念の保有状況調査: 高等学校生物および大学での進化に関する講義の履修の影響に注目して. 理科教育学研究, 59: 285-290.
3. 中野信子 (2020) 人は、なぜ他人を許せないのか?アスコム
4. 生物学者・福岡伸一×為末大対談 (AGUAR I-PACEINNOVATORS’ TALK「遊びやゆらぎ」を許容する生物は淘汰されない.https://www.jaguar.co.jp/jaguar-range/i-pace/special-contents/hukuoka-tamesue.html
5. 小林武彦 (2021) 生物はなぜ死ぬのか。講談社
6. Walker, D. W., et al. (2000) Evolution of lifespan in C. elegans. Nature 405, 296–297 .
7. Kirkwood, T. B. L. (1977) Evolution of ageing. Nature 270, 301–304.
8. Medawar, P. B. (1952) An Unsolved Problem of Biology. H. K. Lewis, London.
9. Williams, G. C. (1957). Pleiotropy, natural selection, and the evolution of senescence. Evolution 11: 398-411.
10. Jones, O. R. et al. (2014) Diversity of ageing across the tree of life. Nature 505, 169–173 .
11. Wynne-Edwards, V.C, (1962) Animal Dispersion in Relation to Socail Behavior. Oilver & Boyd, Edinburgh.
12. Williams, G.C. (1966) Adaptation and Natural Selection, Princeton University Press, Princeton. (辻和希訳)『適応と自然選択』共立出版
13. Hamilton, W. D. (1964),The genetical evolution of social behaviour. I & II Journal of Theoretical Biology, 1-52.
14. Futuyma, D. and M. Kirkpatrick (2018) Evolution. International 4th Edition. Sinauer.
15. Zimmer C. and D. J. Emlen (2016) Evolution:Making Sense of Life. W.H.Freeman. Second ed. (最初の版は、翻訳本が出版されている。ジンマー,K ・D.J. エムレン [更科功・石川牧子・国友良樹訳] 進化の教科書 第1巻から第3巻. 講談社ブルーバックス)
16. Wilson, D. S. (1980) The Natural selection of Populations and Communities. Benjamin/Cummings.
17. Tsuji. K. (1995) Reproductive conflicts and levels of selection in the ant Pristomyrmex pungens: contextual analysis and partitioning of covariance. American Naturalist 146,586-607.
17a. Dobata S. and Tsuji K. (2013) Public goods dilemma in asexual ant societies. Proceeding of the National Academy of Sciences of the USA 110: 16056–16060.
18. Mayden, R.L. (1997)A hierarchy of species concepts: the denouement in the saga of the species problem. in M.F. Claridge et al. (eds.) Species: the Units of Biodiversity. Chapman & Hall, pp. 381-424.
19. Morell, V. (2016) How do you save a wolf that's not really a wolf?https://www.science.org/content/article/how-do-you-save-wolf-s-not-really-wolf
20. Conner, J and , D. Hartl. (2004) A Primer of Ecological Genetics. Sinauer
21. Jablonski, D. (2008) Species selection: Theory and data. Annual Review of Ecology, Evolution and Systematics, 39: 501-524.
22. Normark, B. B., et al. (2003) Genomic signatures of ancient asexual lineages. Biological Journal of Linnean Society, 79: 69-84.
23. Otto, S. P. (2009) The evolutionary enigma of sex. American Naturalist 174: S1- S14.
23a. Otto, S. P. (2008) Sexual Reproduction and the Evolution of Sex. Nature Education 1:182
24. Meirmans, S. & Meirmans, P. G. (2019) The Queen of Problems in Evolutionary Biolog eLS. 1–8 (2019)
24a. Otto, S. P. & Nuismer, S. L. (2004) Species Interactions and the Evolution of Sex. Science 304, 1018–1020 .
25. Felsenstein, J. (1974) Evolutionary advantage of recombination. Genetics 78: 737-756.
26. Matthew R. Goddard (2005) Sex increases the efficacy of natural selection in experimental yeast populations. Nature 434,636-640
26a. McDonald, M. J., et al. (2016) Sex speeds adaptation by altering the dynamics of molecular evolution. Nature 531, 233–236 (2016).
27. Peck, J. R. (1994) A ruby in the rubbish-beneficial mutations, deleterious mutations, and the evolution of sex. Genetics 137: 597- 606.
28. Kondrashov, A.S.(1988) Deleterious mutations and the evolution of sexual reproduction. Nature 336, 435–440 .
29. Takahasi, Y. , K. Kagawa, E. I. Svensson and M. Kawata (2014) Evolution of increased phenotypic diversity enhances population performance by reducing sexual harassment in damselflies. Nature Communications 5: 4468
30. Takahashi, Y. and Noriyuki, S. (2019) Colour polymorphism influences species’ range and extinction risk. Biology Letters 15, 20190228 (2019).
31. Takami, Y.,T. Fukuhara, J. Yokoyama, and M. Kawata. (2018) Impact of sexually antagonistic genital morphologies on female reproduction and wild population demography. Evolution, 72: 2449–2461
32. Yamamichi, M. et al. (2020) Intraspecific Adaptation Load: A Mechanism for Species Coexistence. Trends in Ecology & Evolution, 35, 897-907
33. リチャード・ドーキンス (2018), 日高敏隆ほか訳 利己的な遺伝子. (40周年記念版) 紀伊國屋書店
34. Dreu, C. K. W., et al. (2020) Group Cooperation, Carrying-Capacity Stress, and Intergroup Conflict. Trends in Cognitive Sciences, 24 :760–776.
35. Watson et al. (2011) Global adaptation in networks of selfish associative memory at the system scale. Artifical Life 17: 147–166