
進化における「適応」という言葉をめぐって

進化における「適応」という用語の使い方には、古くから議論があり、その使い方には注意が必要である。最近、G.C. Williamsの『適応と自然選択』の邦訳(1)が出版され、その訳者である辻和希氏の後書き(2)でも「適応」の間違った使い方が指摘された。しかし、適応という概念は、その使い方が難しく、生物学のなかでも、その用法は混乱しているところがある。ここでは、最近の自然選択の研究を踏まえながら、なぜ適応という概念の適用が困難かを解説する。
適応の意味とは
日常生活で、「新しい職場の環境に適応できない」というふうに、その場の状態や条件にうまく対応できることのような意味で「適応 (adaptation)」という言葉を用いることが多い。たとえば、温暖化に対する対応としてもちいられる「適応」は、気候変動による影響に対応して被害を軽減するためのアクションという意見で用いられている。一般的な生物学的意味としては、「ある生物が環境に適合している」ことという意味とみなされることが多い。
しかし、進化学では「生物が環境に適合している」という意味では用いない。そもそも、どのような状態だと適合しているといえるのかが不明確である。たとえば、北海道のような積雪の多い環境では、エゾユキウサギは真っ白な冬毛に換毛する。それに対して、エゾリスの冬毛は、真っ白ではなく、灰色の毛になる。エゾリスの冬毛は環境に適合していないのだろうか。
通常、進化学では、同じエゾユキウサギの集団の中で白く換毛した個体と換毛しなかった個体を比べて生存や繁殖を向上させているとうことが適合(適応)しているかどうかの判断基準となり、エゾユキウサギとエゾリスを比べる意味はない。エゾリスでも灰色の冬毛は、茶色っぽい冬毛のエゾリスと比べると、生存率を向上させているかもしれない。
このように「適応」しているかどうかは、種内の性質の違いの相対的な比較であり、多くの場合、自然選択によって進化したのかどうか、ということと関連づけられている。
現代進化学での適応の定義
進化学でも、様々な適応の定義が主張されている。その中でも、進化の教科書や主要な進化学者が用いている主な定義をみてみると、大きく分類して以下の3つになる。
(定義A) 現在の状態として定義 (非歴史的定義)
(i) 適応とは、その性質を持つ生物の生存や繁殖を、他の性質状態に比べて向上させる性質(3,4)
(ii)適応とは、与えられた環境において、特定の表現型変異の中で最も高い適応度をもたらす表現型 (5)
これらの定義は、生物の性質の現在の状態を観察して「適応」を定義しており、多くの研究者が採用している(16) 。歴史的にも、進化の総合説形成に重要な役割を担った、Mayrは「集団内の他のメンバーに比べて、より生態的・生理的効率の高さ」を適応とし、また、Dobzhanskyも適応を「ある環境において、生存や繁殖を容易にする発達パターンの要因」としている(5)。
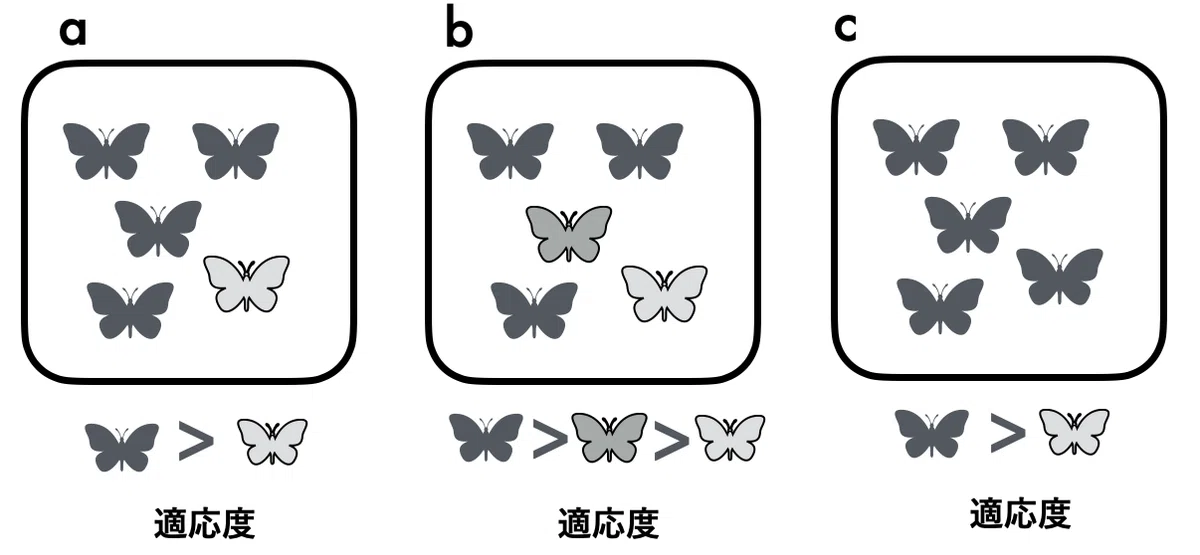
すこしわかりずらいので、補足しよう。たとえば、図1のように、暗い翅色の蝶と明るい翅色の蝶がいる。蝶がとまる樹木の色が黒いという環境では、暗い翅色という性質を持つ蝶は、明るい翅色を持つ蝶に比べて適応度( 一生に残す子どもの数)あるいは生存や繁殖を向上させるので、暗い翅色という性質は適応といえる。また(ii)の定義では、集団中に暗い翅色から明るい翅色まで明暗のバリエーションがあるとき、そのなかで一番暗い翅色の適応度が高いとき、暗い翅色という性質は適応といえる。
それでは図1cのように、集団中にはすべて暗い翅色の蝶しかいなかったとする。このときは、たとえば、実験的に暗い色に影響する遺伝子をノックアウトしたり、翅の色を人工的に変えてやり、適応度が高くなるかを調べて、暗い翅色の蝶が適応度が高いかどうかをみる(20)。
(定義B) プロセスとしての定義
(i)「適応」 とは、生物が世代を重ねるうちに、生存や繁殖に影響を与えるような特徴を持つように変化していく進化の過程 (4)
(ii) 形質が正の自然選択によって進化する過程(6)
ここで「正の自然選択」とは、適応度の高い表現型変異が集団中で頻度を増大させるプロセスである。
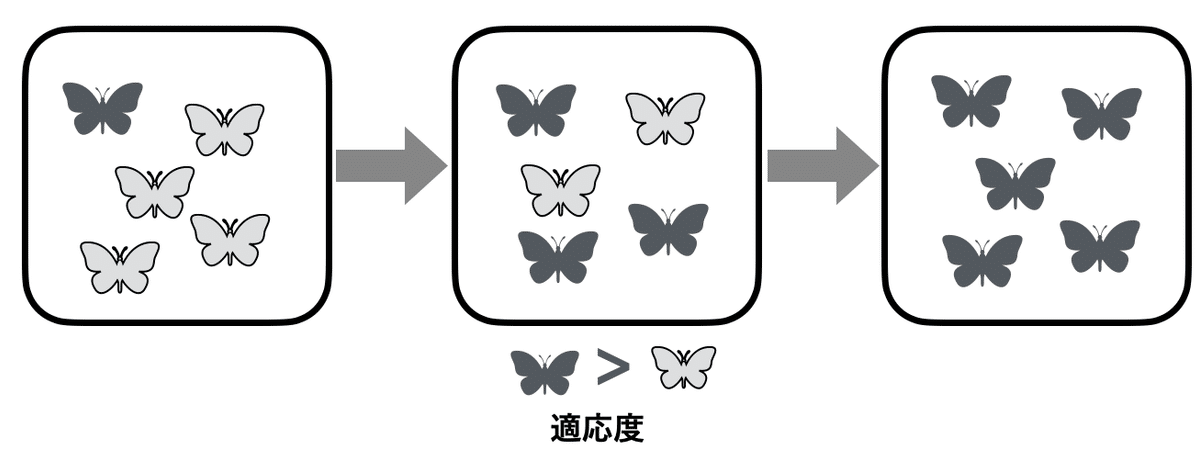
このBの定義では、適応は「高い適応度に貢献する性質が自然選択によって進化していくプロセス」となる(図2)。Endler (11) は、「適応」は、より適応を得るためのプロセスを指し、「適応形質」を「適応」と同義に使うべきではないと主張している。同様に、『アメリカ版大学性物学の教科書.4.進化生物学』 (6)は(ii)を採用し、訳者の斎藤成也氏は、訳注として「日本語では適応は過程(プロセス)のみをさし、形質は適応的形質と呼ぶ」と付記している。
(定義C) 自然選択の結果としての定義
(i)「適応」とは、自然選択によって進化した生物の特性である。(1,2)
(ii) ある性質が集団において適応といえるのは、その性質が自然選択により集団中に広がっているからである(10)、
(iii) 生物に現在の有用性をもたらす性質で、その現在の生物学的役割のために自然選択の作用によって歴史的に形成された性質(15) (歴史的定義)
冒頭でのべたG.C. Williams(1)は適応は自然選択を受けて発達した性質の意味で適応をつかっている(2)。Williamsは、自然選択によって作られた機能的デザイン、あるいは「生物が直面する問題の解決に役立つ」性質やしくみを適応としており、「目的論的定義」ともいわれている(5)。
図3の例でみてみると、暗い翅色の蝶は自然選択で頻度を増大させた結果(赤枠の部分)が適応となる。図3aでは、現在の状況では、暗い翅色の蝶が集団全体を占めていない場合で、図3bはすべて暗い翅色の蝶になった例である

自然選択による適応
生物は、様々な形態的、生理的、行動的性質を持っている。また、それらの性質は、様々な「働き」を持っている。しかし、すべての性質が自然選択で進化した性質ではない。
自然選択によって進化するためには、その性質の個体間の違い(個体変異)が、その個体の生存や繁殖(適応度)に違いをもたらし、そのために性質に影響する遺伝子(アレル)が集団中に頻度を増大させる、というプロセスが必要である。
このとき、適応度の差をもたらす性質の働きが適応的「機能」である。たとえば、前述の蝶の例だと、「捕食者からの隠蔽」という機能の違いが適応度の差をもたらすということになる。
FutuymaとKirkpatrick(4)は、生物の性質には少なくとも4つの説明が可能であるとし、以下のような例をあげている。
物理・化学的結果 たとえば、ヘモグロビンは、酸素を運ぶという機能をもっている。血液を赤くするいう性質もあるが、「赤」という性質は、タンパク質の構造による副産物である。
遺伝的浮動など自然選択以外の進化要因 ヘモグロビンは、アミノ酸が連なってできているが、種間のアミノ酸配列の違いの大部分は、自然選択によるものでなく、ランダムな遺伝的浮動によるものである。性質の違いが遺伝的浮動によって生じる場合は少なくない。
副次的進化 その性質自体が自然選択を受けて進化したわけではなく、別の性質が進化した副次的な結果として進化した可能性がある。たとえば、生物個体の死は、それ自体が適応進化ではなく、生物の早期の繁殖に自然選択が働いたこどによる副産物として老化が進化し、死が進化したと考えられる(記事「老化の進化:なぜ老化しない生物がいるのか?」を参照)。
系統的歴史の結果 ダーウィンが『種の起源』で述べている例として、哺乳類の頭蓋骨の縫合がある。哺乳類では、子どもの頭蓋骨の縫合はゆるく、母親の出産の苦痛を軽減している。しかし、この縫合は、鳥類や爬虫類にもみられることから、この構造は頭蓋骨の成長に重要な性質であり、哺乳類で出産が進化する以前から存在している性質であり、歴史的結果と考えられる。
この他にも、生物の発生上必然的に生じる形や性質のような発生的制約によって説明される性質や、突然変異のバイアスによって特定の性質の出現率がたかまることで進化した性質など様々な要因が考えられる。様々な要因の中から、自然選択によって進化した性質を区別するという考えは、それを「適応」と呼ぶかどうかは別にしても、生物の性質の特質や働きを理解する上で、進化学だけでなく生物学においても重要である。
Williams(1)は、生物の性質が、何らかの働きがあったとしても、その働きが自然選択されたものを「適応」とよび、そうでない性質を「効果」と呼よんで区別すべきだという(1,2) 。たとえば、人間の耳は、音を集音する適応的「機能」があるが、メガネをかけるという働きは「効果」ということである。適応は必要なときだけにつかわれるべき専門的でわずらわしい概念だとしている(1)。(著書『適応と自然選択』では、Willimans(1)は、特に、「集団のための機能」が進化するという集団選択について批判している。たとえば、集団の個体数の増加を抑える移動分散行動は、集団や集団の絶滅を防ぐ機能のために進化した適応ではなく、単なる個体の進化の結果(効果)であるという点を明確に主張している。「記事「種の保存のための進化」はどこが誤りなのか」を参照)
West-Eberhard(17)は、ある生物の性質を適応とみなすためには、「生物が直面する問題を効果的に遂行する能力が備わっている」ことを示したり、「生物と環境の適合性があることを示す」だけでは不十分であるとしている。ある形質が適応と見なされるためには、自然選択であれ性選択であれ、その形質が自然選択の結果であることが示されなければならないと、多くの研究者は考えている。
現在の表現型から類推する自然選択の証拠
適応を「自然選択の結果」と定義すると、ある性質が自然選択が働いたことを証明しなければいけない。
West-Eberhard(17)は,自然選択が働いたことを確かめる方法として、以下の3つをあげている。
I. 自然界に存在する変異(個体間で異なる性質を示す)を比較し、性質がより高い適応度を示すかどうかを調べる。
II. 実験的に性質を変化させたり除去したりして、それが特定の機能や環境条件における適応度にどのような影響を及ぼすかを調べる
III. 性質と環境、あるいは性質とその用途との相関関係をみる。たとえば、系統的に近いか遠いかを考慮して、同じ環境に生息している種では、同じ性質が進化している傾向があるときは、適応である可能性が高いとみなされる。この手法は、系統樹を用いて、性質の系統関係とその環境との関係を統計的に検定する比較法として使われている(16)。
このIとIIは、適応の定義Aと関連している。Iを確かめるということは、問題となる「性質が高い適応度を示す」という適応の定義A(図1のaとb)と同じである。また、IIの場合は、図1のcで定義された適応を確認することと同じである。つまり、定義Aにあてはまる性質は、自然選択によって進化した性質とみなされ、適応ということになる。
しかし、生物の生存や繁殖を向上させるような性質あるいは個体の適応度を向上させる性質(定義A)は、必ずしも自然選択によって進化したとは限らない。たとえば、自然選択が有効に働くかどうかは、異なる性質を示す個体間の適応度の差と個体数(有効集団サイズ)で決まる。適応度の差が小さかったり、個体数が小さかったりすると、遺伝的浮動による偶然の効果で進化する。たとえば、ヒトの場合(有効集団サイズが12〜14千)、強い選択(10%適応度が高い)が働いても、その遺伝子が集団中に広まり固定する確率は40%に満たない(26)。また、現在の環境で観察される適応度の向上が、その性質が進化した過去でも同じであったかどうかはわからない。
Williams(1)自身が述べている例がある。キツネは、雪の上についた自分の足跡を再びたどることで餌にありつける確率が高まり、生存が向上する。しかし、「足跡をつける」という性質は生存を向上させるが、自然選択によるものではない。
IIの方法はどうだろうか。たとえば、蝶の翅色を人工的に明るい色にして、暗い翅色の蝶は、明るい翅色の蝶に比べて、生存や繁殖を向上させた(適応度が高い)た場合、定義Aでは、暗い翅色は適応となる。しかし、明るい翅色から暗い翅色が進化したのではなく、実際には、明るい翅色から真っ黒な翅色が自然選択で進化し、その後、黒い翅色から暗い翅色(灰色)が遺伝的浮動で偶然に進化したかもしれない。この場合は、暗い翅色は自然選択の結果ではない。IIの方法では、現在の表現型の祖先型が不明なため、自然選択を検出するには限界がある。
Williamsは、複雑な機能と適合している複雑なデザインは現在適合している機能に働く自然選択によって作られるとしているが、現実には、そう単純ではない。たとえば、コウモリの「翼」は、飛翔という機能に対して自然選択が働いたと考えられる。しかし、翼の形成には、中手骨(指の骨の一つ)を選択的に成長を促して重要な指を伸ばし、さらに、指の間に膜が形成されることが必要である(21)。指の骨の伸長は、昆虫などの餌をとるために進化したかもしれないし、指間の膜も、捕獲の際に昆虫などが逃げ出さない機能で進化した可能性もある(22)。コウモリの翼は、もともと「飛翔」以外の機能に対して自然選択が働いたと思われる。
同様に、音を聞く能力、聴覚について考えてみよう。我々、人間にとって聴覚は、コミュニケーションに役立っている。しかし、もともと聴覚が進化的に自然選択によって獲得された理由は、捕食者や餌動物を見つけたりすることで生存を向上させたのかもしれない。しかし、現在の人間の聴覚は、捕食者から逃れる働きをもっているとはいえない。つまり、聴覚は、捕食者回避や餌獲得の機能で進化した適応形質といえるかもしれないが、現在では、聴覚はその機能を果たしていない。GouldとVra(7)は、現在の機能とは別の働きで進化した場合、進化した原因と関係している機能(聴覚なら捕食者回避や餌獲得?)をエグザプテーション(exaptation)と呼んだ。つまり、現在において生存や繁殖に役立っている働きは、その性質が進化した理由とは別かもしれないということである。
現在の生物で観察される性質が生存や繁殖にどのように向上させているかどうかを調べることは可能であるが、そこから、その性質が自然選択が働いた結果として進化したかどうかをみることは難しい。また、自然選択の結果進化した性質が、現在、個体の適応度を向上させているとは限らない。
IIIの比較法では、現在の性質の祖先状態を推測したりできる場合もある。また、現在では、ゲノム配列を用いて、現在あるいは過去の自然選択を検出することが、ある程度できるようになってきた(後述)。しかし、ある性質が、過去のどの段階で、どのような機能(個体の適応度を向上させる機能)のために、自然選択によって進化したのかを、特定するのは、簡単ではない。
非歴史的定義と歴史的定義の関係
「進化の結果として適応(定義C)」が広く受け入れられているのと同様に、「適応とは、その性質を持つ生物の生存や繁殖を、他の性質状態に比べて向上させる性質」という現在の性質の状態を表している定義Aも、多くの研究者が採用している(16) 。
定義Aと定義Cの関係がどのように想定されているかには、複数の暗黙の考えがある。ここでは、定義Aと定義Cとの関係をみてみよう。
(A) 定義Aは定義Cを示唆している。
前項でも説明したように、定義Aで特定できた性質は、その定義から自然選択によって進化したと考える立場である。
Martins (16) は、ほとんどの研究者が認める適応の定義として「同じ環境下で他の表現型よりも進化的に高い適応度を示す表現型である」とし、その上で、その表現型は進化の過程で自然選択をうけ、現在の役割を果たすようになったはずだと主張している。
つまり、定義Aで示された性質は、自然選択で進化した可能性が高い、という認識があるためだと思われる。実際に性質が自然選択で進化した証拠の一つとして、「現在の生物で、その性質を持っている個体と持っていない個体の間の適応度を比較する(前項のI.)」ことがあげらている(17)。また、行動生態学の分野では「1種の生物に注目し、個体の適応度をもっとも高めるような振る舞いを生物が示すならば、環境がおおむね今と変わらない近い過去において、自然選択が作用したことの証拠になるという仮定で研究を進めている」としている(2)。つまり、現在の生物を観察して、適応の定義(A)が満たされれば、それは、自然選択で進化した性質とみなしていい、という仮定を置いていることになる。
(B) 定義Aと定義Cを同時にみたすとき適応といえる
適応形質として問題にしている適応形質は、現在の生物で何からの役割を果たしている(適応度を向上させる)性質であり、かつ、それが自然選択によって進化した性質であるという場合である。つまり、適応かどうかを問題にしている性質は現在観察される性質であって、過去の性質については言及しないという立場である。実際に多くの研究は、現在の性質の役割や機能について議論していることが多い。
(C) 定義Aはその原因や生成要因とは独立に定義する
ダーウィンは、現実世界で観察される特徴の一つの説明として自然選択を提唱した(18)。適応あるいは適応形質は、説明される性質であって、説明する原因とセットになっていないと思われる。つまり、適応とは、生物の生存や繁殖を向上させるような性質であるが、その性質が生じた原因の一つが自然選択であるということだ。
しかし、前項でのべたように、生物の生存や繁殖を向上させるような性質あるいは個体の適応度を向上させる性質(定義A)は、必ずしも自然選択によって進化したとは限らない。
Reeveと Sherman (3)は、適応の定義を「それがどのように進化したのか」という進化機構を定義から切り離し、「適応とは、与えられた環境において、特定の表現型変異の中で最も高い適応度をもたらす表現型」(定義Aii)を提案している。
適応が過去の自然選択によって進化した結果とすると、どの程度前に働いた自然選択なら適応といえるのか、生存や繁殖を向上させている現在観察される性質は、過去にその性質の別の側面で自然選択を受けた可能性はないのか。このように、自然選択の過去の歴史を証明するのは困難であることから、適応の定義に歴史的な自然選択の要因を考慮するのは、実用的ではないとする。
Reeveと Sherman (3)は、適応を自然選択とは別に定義することで、検証すべき仮説として扱えるというのである。つまり、適応的性質を定義した上で、その進化的理由を探っていくことができる、という主張である。自然選択理論は、ある特定の選択肢の中で適応度を向上させる表現型が想定された環境で優勢になると予想できる。この予想が棄却された場合は、その表現型の頻度には自然選択が関係していないということになる。
ゲノム解析による自然選択の検出
2000年代以降、生物のゲノム配列が解読され、様々な生物のゲノム配列情報が容易に用いることができるようになったきた。また、数万年前の古代人ゲノムなど、過去の生物のゲノム情報も得られるようになった。これらゲノム配列情報から、自然選択を受けた遺伝子を検出する研究が増加してきた。また、量的形質(体長のように連続的に変化する性質)やポリジーン形質(多数のゲノム変異が絵影響している性質)に影響するゲノム上の変異を検出することで、それらの性質が自然選択を受けたかどうかも検出することが可能になった(たとえば参照「古代人・現代人ゲノムから探る人の「こころ」の性質の進化」参照)。
近年のゲノム解析による進化研究から、現在の生物の現状から自然選択を類推することの難しさを別の観点から浮き彫りになってきた。その例の一つを紹介する。
最近、ヒトの乳糖消化酵素(ラクターゼ)の進化の興味深い研究が報告された(8)。日本人では、ミルクに含まれる乳糖を、赤ちゃんのときは消化できるが、成長すると消化できなくなる。しかし、ヨーロッパ人は、ヒトが牧畜を開始し、ヤギや牛の乳を飲むようになって、大人になっても乳糖を分解することが可能な持続性ラクターゼ遺伝子(=持続性アレル)を進化させた。この持続性アレルは自然選択を受け頻度を上昇させたことは、ゲノム解析からも示されている。持続性アレルは、ミルクをよく飲む現在の環境において適応といえるだろうか?
ヒトがミルクを消費しはじめたのは、紀元前7千年紀にアナトリアであるといわれている。古代人ゲノム配列をつかって、持続性アレルの頻度を調べたところ、イギリスでは、3000年前ころから持続性アレルが出現し、2000年前ころに急速に増加したことが示された(8)。ところが、現在のイギリス人を対象に、持続性アレルを持つ人とそうでない人とを比べたところ、生存率には差がないことが示された。持続性アレルを持たない人は牛乳を飲んだ後、腹部膨満感、痙攣、下痢などの軽度から重度の症状を経験することがあるが、大量に牛乳を飲まないかぎり、生存に影響するほどではないという結果だ。
ヨーロッパの鉄器時代(約2,000年前)には、人口が増加して農畜産物に依存するようになり、気候変動による不作で飢餓の影響が大きくなったり、感染症リスクが増大したりした。そのような飢餓などの状況では、持続性アレルをもっているかどうかが生存に影響し、自然選択が働いた可能性が指摘された。一方で、現代のイギリスでは、持続性アレルを持っているかどうかは生存率に差はなく、自然選択は働いていないようだ。
この例は、乳糖分解持続性という性質は、自然選択で進化した結果であるが、現在の人間の適応度の差には影響していない。つまり、最近(約2000年前)に自然選択で進化した性質であるが、定義A(持続性アレルともつ人は持たないヒトに比べて適応度が高い)は満たされないことになる。
おそらくこのような例は、例外ではなく、むしろ一般的だと推測される。過去に環境が一次的に激変したときに、強く自然選択が働き、それ以外の安定した環境下では自然選択が働いていないという場合は多いと考えられる。
また、強い自然選択は、急速に遺伝子頻度を増大させ固定(集団中にすべて占める)するので、現在の遺伝的な変異からは自然選択が検出しづらいということもある。
たとえば、ショウジョウバエの野外集団のゲノム配列を調べた研究では、現在の集団中で維持されている遺伝的変異のほとんどは、中立か有害である。つまり、現在、強い自然選択が働いているような遺伝的変異はごくわずかしかないということになる(9) 。しかし、近縁種と種が分岐してから、自然選択を受けて進化したと推定される遺伝子は5から10%ある(9)。つまり、過去に自然選択が働いた遺伝子は相当数あるが、現在の集団で自然選択が働いている遺伝子は少ないということだ。
近年、ゲノム解析によって自然選択の検出が行われるようになってきた。それらの研究が示していることは、現在あるいはそれほど古くない過去で強い自然選択を受け頻度を増大させている遺伝子は、全遺伝子の数%にすぎないということである。例えば、日本人の集団において、過去3000年前に強い正の自然選択を受けたと推定された遺伝子は3つ(12)、約2万年前に自然選択をうけたと推定された遺伝子は29に過ぎない(13)。
もっとも、現状のゲノム解析手法だけでは検出できない弱い選択を受けている変異は多数ある。たとえば、日本人の身長は低くなる方向に自然選択が働いていることが検出されているが(14)、身長には多数の変異(SNP)が少しずつ影響しており、単独の変異では自然選択を検出することは難しい。いずれにしても、現在の生物で表現型変異に適応度の違いがないことは、近い過去に自然選択が作用していなことを意味していない。
自然選択、特に強い自然選択が働くと、自然選択に有利な変異は、比較的早く頻度を増大させ、ほぼ固定(すべてがその変異だけになる)する。従って、現在観察されている性質をみても、適応度に違いのある遺伝的変異はそれほど多くないという可能性はある。
自然選択のプロセスか結果か
適応を自然選択が働いているプロセスとして定義する場合も少なくない(定義B) 。元来、適応とは、ad + aptusで「適応する」という意味で、生物の集団が有利な形質が優勢になるにつれて、より環境に適合するように進化していくプロセスのことをいう(25)。つまり、ある性質が自然選択によって集団中に広がっている正の自然選択の過程を適応と呼ぶ。英語では、プロセスとしての適応を「Adaptation」として、結果としての適応を「an adaptation (加算名詞)」あるいは「adaptive trait」(適応的形質)として区別している。
『野外での自然選択』という有名な著書の中で、Endlerは、適応をプロセスとして定義している(11)。また、いくつかの進化学の教科書でもこの定義を採用している(6)。
進化には、正の自然選択による適応進化、遺伝的浮動による中立進化および有害進化が区別できる。この適応進化という言葉は、自然選択というプロセスで生じる進化という意味であり、定義Bを意味している。
自然選択が働いているプロセスを適応とするという定義は、問題としている性質が過去に起こった自然選択によるものか、現在のものかに関わらずに適用できるという意味では、曖昧さはない。また、多くの性質は、自然選択のみによって形成されるわけではないので、自然選択の結果としての性質を認識することの困難さを回避できる。
たとえば、コウモリの翼の例でみてみよう。自然選択が働くことで、翼の形態や運動機能に影響する遺伝子が進化する。このプロセスが翼における適応進化である。しかし、現在の翼のどの部分に自然選択が働いたかは、結果としての翼をみただけではわからない。この場合、翼に関わる遺伝子の適応プロセスは明確に定義できるが、翼を自然選択の結果といえるかどうかは曖昧になる。
この定義Bを使う場合は、進化のプロセスと進化の結果を同時に用いて混同しないことが重要である。「生物は環境に適応した性質をもっている」– それは「その性質が環境に適応しているからだ」という説明が、トートロジー(循環論法)になっているという批判が古くからある。この場合と同様に、適応をプロセスの意味と結果としての意味に同時に用いてしまうと「生物のある性質は適応によって適応した」ということになり、循環論法として矛盾した表現になる。このようなことからも、Endler (11) は、選択の結果を「適応形質」とし、プロセスを「適応」として、同義に使うべきではないと主張している。
前述したように、「生物の適応性質が生物の適応度を上昇させている」ことは適応的な変異が自然選択によって広がった結果とは限らない。そのため、自然選択による適応の説明が循環論法になるというのは、表現上の問題にすぎない。しかし、適応という用語を使うときは、そのような表現にならないような注意が必要である。
結局、「適応」をどう定義すればよいのか?
ここで紹介した3つの適応の定義は、進化生物学の研究者によって現在も用いられているものである。どの定義も、想定した環境下で自然選択によって生物が生存や繁殖を向上させる性質と関係づけられている。しかし、その関係の付け方が、研究者によって異なるということだろう。
ここで、この3つの定義のどれか1つだけにすべきだと主張しても、おそらく統一されることはないだろう。重要な点は、適応という言葉を用いるときに、どのような意味で用いているかを明確にすることである。特に、原因と結果、プロセスと結果を混同しないことが重要である。
進化学にとって重要な点は、生物の様々な性質がどのような機構で進化したのか、ということである。ダーウィンの時代から、総合説形成、行動生態学・社会性物学の全盛期まで(1990年代頃まで)は、「生物が特定の環境で生存や繁殖を向上するのに寄与する表現型は自然選択でのみ進化可能である」という考えが強かった。進化要因の中で、自然選択の重要性が強調されすぎた時代でもあった。そのような時代には「自然選択によって進化した性質だけを適応をする」という主張は意味があったのだろう。
当時は、生物の表現型に関与する遺伝子やゲノム上の変異についてはほとんどわかっていなかったこともあり、表現型に働く進化要因の複雑さや過去に働いた進化プロセスの推定などがまだ困難であった。しかし、集団内の多数の個体や古代生物のゲノムを解析したり、近縁種とのゲノム配列の比較することで、過去から現在にまでの間で働いた正の自然選択が検出が可能になってきた。
現在の環境下で、現在の生物を対象に、性質の適応度を調べることで、自然選択を検出しようとする試みは限界がある。同様に、そこから適応を特定しようとすることにも限界があると思われる。
また、生物の複雑な形質は、複数の形質(モジュール)の組合わせでできている。それぞれのモジュールでは、別々のゲノム変異が関連しており、それぞれ異なった要因で獲得されたり、維持されている(23)。
たとえば、前述したコウモリの翼は、飛ぶという機能を果たすためには、運動制御や翼の形態の変化などに関与する遺伝子が自然選択を受けたと思われる。しかし、飛翔を可能にした翼を構成する複数の形態は、別の自然選択を受けたり、遺伝的浮動が関与している可能性が高い。たとえば、指の伸長には、TGF-β活性、BMP2遺伝子やその制御が関連し、指の間の物質を除去するアポトーシスを抑制する遺伝子が、膜の進化に関与している(24)、それらは餌獲得に対する自然選択が働いたのかもしれない。
これからの進化学では、具体的にどのような性質に関わる遺伝子がどのような自然選択や遺伝的浮動を受け、進化したのか、を調べていくことが必要である。「コウモリの翼は飛ぶ機能のために自然選択を受けた結果の適応である」という表現は、一般向けにはいいかもしれないが、進化学上では曖昧な表現といえる。
科学において現象をどう定義するのかという問題は、(1)定義することで、現象の理解を高める、(2)研究者によって異なる意味で同じ言葉が使われるなどの混乱を避ける、ということだろう。その点からすると、適応という用語は、「理解を高める」というよりも混乱を招いているといえる。混乱を避けるためには、進化の原因や結果と関連づけて「適応」という言葉を用いるときは、どの定義を採用しているかを、明確にすることが重要である。
私自身としては、適応を定義Aの「適応とは、その性質を持つ生物の生存や繁殖を、他の性質状態に比べて向上させる性質」の意味で使うことが多い。ただし、そのような適応的性質は、自然選択の結果進化したとは限らず、また、自然選択が関与していたとしても、それ以外の様々な複雑な要因も絡んで進化したと考える。
本記事は、「適応」という言葉の定義について明確にするというよりも、読者には、かえって分かりづらくさせてしまったかもしれない。結局のところ、適応という用語は、正確な意味として定義することは難しく、やっかいな言葉である、ということだ。
謝辞
本記事の最初の原稿に対して、琉球大学の辻和希さんに有益なコメントを頂きました。
進化のしくみについては以下の新書を参照してください。
以下関連記事です
「種の保存のための進化」はどこが誤りなのか
老化の進化:なぜ老化しない生物がいるのか?
犬はなぜ多様に進化できたのか:祖先の遺伝的多様性の役割
引用文献
Williams G.C. (1992) 『適応と自然選択』共立出版 (辻和希訳、2022)
辻和希 (2022) 訳者後書 『適応と自然選択』共立出版
Freeman, S. and J.C. Herron (2004) Evolutionary Analysis. 3rd ed. Pearson Prentice Hall.
Futuyma, D. & Kirkpatrick, M. (2018) Evolution. 4th ed. (Oxford University Press, 2018).
Reeve, H. K. & Sherman, P. W. (1993) Adaptation and the goals of evolutionary research. Quarterly Review of Biology 68, 1–32 .
Hillis, D. M., et al. (2012) Principles of Life. Sinauer Accociate.斎藤成也訳『アメリカ版大学性物学の教科書.4.進化生物学』
Gould, S. J. and E. S. Vrba. (1982). Exaptation- a missing term in the science of form. Paleobiology, 8: 4-15.
Evershed, R. P. et al. (2022) Dairying, diseases and the evolution of lactase persistence in Europe. Nature 608, 336–345.
Mackay, T. F. C. et al. (2013)The Drosophila melanogaster Genetic Reference Panel. Nature 482, 173–178 .
Sober, E. (1984) The Nature of Selection. MIT. Press, Cambridge.
Endler, J. A. (1986) Natural Selection in the Wild. Princeton University Press, Princeton.
Okada, Y. et al. (2018) Deep whole-genome sequencing reveals recent selection signatures linked to evolution and disease risk of Japanese. Nature Communications 9, 1–10.
Yasumizu, Y. et al. (2020) Genome-Wide Natural Selection Signatures Are Linked to Genetic Risk of Modern Phenotypes in the Japanese Population. Molecular Biology and Evolution 37, 1306–1316.
Akiyama, M. et al. (2017) Genome-wide association study identifies 112 new loci for body mass index in the Japanese population. Nature Genetics 49, 1458–1467.
Lauder, G. V., Leroi, A. M. & Rose, M. R. (1993) Adaptations and history. Trends in Ecology & Evolution 8, 294–297.
Martins, E. P. & Martins, E. P. (2000) Adaptation and the comparative method. Trends in Ecology & Evolution 15, 296–299.
West-Eberhard, M. J. (1992) Adaptation: current usages. In E. F. Keller and E. A. Lloyd (eds.), Keywords in Evolutionary Biology, pp. 13-18. Har- vard University Press, Cambridge.
Fisher, R. A. (1958) The Genetical Theory of Natural Selection, 2nd ed. Dover, New York.
Futuyma, D. J. (2010)Evolutionary constraint and ecological consequences. Evolution 64, 1865–1884.
Grafen, A. (1988) On the uses of data on lifetime re- productive success. In T. H. Clutton-Brock (ed.), Reproductive Success, pp. 454-471. The University of Chicago Press, Chicago.
Weatherbee, S. D., et al. (2006) Interdigital webbing retention in bat wings illustrates genetic changes underlying amniote limb diversification. PNAS 103, 15103–15107.
Anderson, S. C. & Ruxton, G. D. (2020)The evolution of flight in bats: a novel hypothesis. Mammal Review 50, 426–439.
Sella, G. & Barton,N. H. (2015) Thinking About the Evolution of Complex Traits in the Era of Genome-Wide Association Studies. Annual Review of Genomics and Human Genetics 20, 1–33 .
Wilkins, A. S. (2007) Between “design” and “bricolage”: Genetic networks, levels of selection, and adaptive evolution. PNAS 104, 8590–8596.
Gregory, T. R. (2009 )Understanding Natural Selection: Essential Concepts and Common Misconceptions. Evolution: Education and Outreach 2, 156–175).
Olson-Manning, C. F., et al. (2012). Adaptive evolution: evaluating empirical support for theoretical predictions. Nature Review Genetics 13, 867–877.
この記事が参加している募集
この記事が気に入ったらサポートをしてみませんか?