
「利己的遺伝子」の誤解を招かない使い方

一般に使われている「利己的な遺伝子」は、「生物の様々な性質は、遺伝子が自らのコピーを残すのに都合が良いように進化させた結果である」ということを分かり易く表現したものだ。これは、リチャード・ドーキンスが『利己的な遺伝子』という一般向けの本で示した一つの見方である。しかし、進化学では、ドーキンス流の「利己的遺伝子」とは少し意味の異なる「利己的遺伝因子」の意味で使われることが多い。ドーキンス流の「利己的遺伝子」の見方がなぜ進化のプロセスを考える上で、誤解を招きやすいかを解説する。
本記事を修正・加筆した記事が、以下の新書の第3章に収録されています。
「利己的遺伝子」とは
「恋をするのも、争うのもすべては遺伝子の思惑通り?」これは2022年科学道100冊に選ばれたリチャード・ドーキンス「利己的な遺伝子」(1)の紹介のタイトルである。このような表現は、生物のあらゆる性質は遺伝子にとって有利に進化した結果である、ということを示唆したものだ。あまりにも表現が「俗っぽい」ので、うさんくさい、と思う人もいれば、深い理屈は考えずに、そうかもしれないと思う人もいるかもしれない。
ドーキンスのいう「利己的な遺伝子」とは、どのようなものだろうか。ドーキンスによれば、世代を超えて伝わるのは、複製される単位(レプリケーター)としての遺伝子である[実際の遺伝子の定義は Box 1]。これに対して、個体は遺伝子と環境との相互作用でつくられるが、それがそのまま複製されて次世代に伝わることはない。子どもに親の性質が遺伝するのは、遺伝子が複製されるからである。
遺伝子は複製の単位、個体はそれを運ぶ乗り物である。遺伝子は通常、裸では存在できないため、自分のコピーをより多く残すためには、遺伝子を運ぶ乗り物、すなわい個体ができるだけ多くの子どもを残すことが必要になる。そこで、遺伝子は、自らのコピーを残すのに都合のよい乗り物の性質(表現型)をつくり出そうとし続けてきた。その結果、利己的に振る舞う遺伝子が進化したという見方である。ドーキンス自身も、これは「見方」であって、理論でも仮説でもないとしている。
この見方によると、個体の様々な性質は、遺伝子が自らのコピーを残すためにつくられたもの、ということになる。さらに、ドーキンスは、遺伝子がその乗り物である個体の枠を越えて影響を及ぼすことがあるという。たとえば、ある種の病原体や寄生虫は、感染した動物の行動を変化させ、その病原体や寄生虫が次の宿主に移行できるように操作している場合がある。このような場合、遺伝子の影響は、自らを運んでいる個体という乗り物(病原体や寄生虫)だけでなく、その宿主まで影響しているとする 。
一見するところ、納得できるような説明である。実際のところ、「利己的な遺伝子」という用語は、一般の人がなんとなく比喩的な表現で使うのは問題ないかもしれない。しかし、この表現が、厳密に生物進化を適切に理解するための比喩的表現だ考えるのは誤りである。
まず、『利己的な遺伝子』という比喩がだされた背景を理解する必要がある。リチャード・ドーキンスが『Selfish Gene』を出版したのは1976年である。このころ、生物の行動を自然選択によって説明しようとする行動生態学や社会生物学といった分野が急速に浸透してきた。その時期に、これまで生物の様々な性質が「種のため」に進化したという暗黙の考えが誤りであることを示されたこと(「「種の保存のための進化」はどこが誤りなのか」を参照)、そして、W.H.ハミルトンによって、個体が自らの繁殖を犠牲にして、他個体の生存や繁殖を助けるという利他行動が、同じ遺伝子を共有する近縁個体を通して進化するという血縁選択が提唱されたことが、行動生態学などの盛り上がりと関係している。
1960年代から1970年代にかけて、自然選択はどのように何に対して働くのか、という自然選択の単位について議論され、基本的には、自然選択は個体もしくは遺伝子に対して働くという理解になった。そのような時代背景のなかで、ドーキンスは遺伝子の視点から進化を見てみると、自然選択がより分かり易く理解できると主張したわけだ。
Box 1. 遺伝子の定義
遺伝子の定義は大きくわけて、メンデル的遺伝子と分子学的遺伝子とがある(2)。メンデル的遺伝子とは、「異なる対立遺伝子(アレル)で説明される表現型の違いによって認識される遺伝単位である」。たとえば、エンドウ豆の表現型である「丸」と「しわ」には、アレルAとアレルaが関係する。そして、遺伝型AAとAAが丸、aaがしわという表現型を説明できる。この場合、ゲノム配列のどこからどこまでがアレルAあるいはアレルaなのかには言及しない。遺伝子とは、表現型の違いを引き起こすアレル(=遺伝子)であると見なされる。ドーキンスが想定する遺伝子は、このメンデル的遺伝子を指すと思われる。
それに対して、分子学的遺伝子は、ゲノム配列から定義する。ゲノム配列には、様々な領域が存在している。RNAに翻訳され、そこからタンパク質が作られるコード領域とよばれる配列、タンパク質の翻訳をコントールする転写因子結合領域や、タンパク質には翻訳されないRNA(ノンコーディングRNA)を転写する領域のほか、機能が不明あるいは機能がない配列、また転移因子(本文での説明参照)とよばれる領域などがある。
遺伝子のより狭い定義として、「ゲノム配列のなかで、mRNAに転写され、タンパク質が作られるDNA領域を遺伝子」と定義される。これは「コード領域」と呼ばれることが多い。前述したように、ヒトゲノムでは、この「コード領域」は30億塩基の1から2%である。現在の分子学的遺伝子の定義としては、「タンパク質に翻訳されるかどうか関係なくRNAに転写されるゲノム領域」を遺伝子とする場合もあれば、それに加えて、「転写因子など遺伝子発現を制御する領域などを含めて遺伝子」とする場合がある。教科書『細胞の分子生物学』では、「ゲノム配列からRNAとして転写される一つの領域」を遺伝子としている。また、「遺伝子は、制御領域、転写領域および/または他の機能配列領域と関連している一つの遺伝する単位としてのゲノム配列領域」とするものもある(3)。どちらにしても、ゲノム配列の中で、タンパク質を直接作ったり、それを制御したりして、何からの生物個体の性質に影響する「機能」をもっている領域を遺伝子としている。
個体に働く自然選択
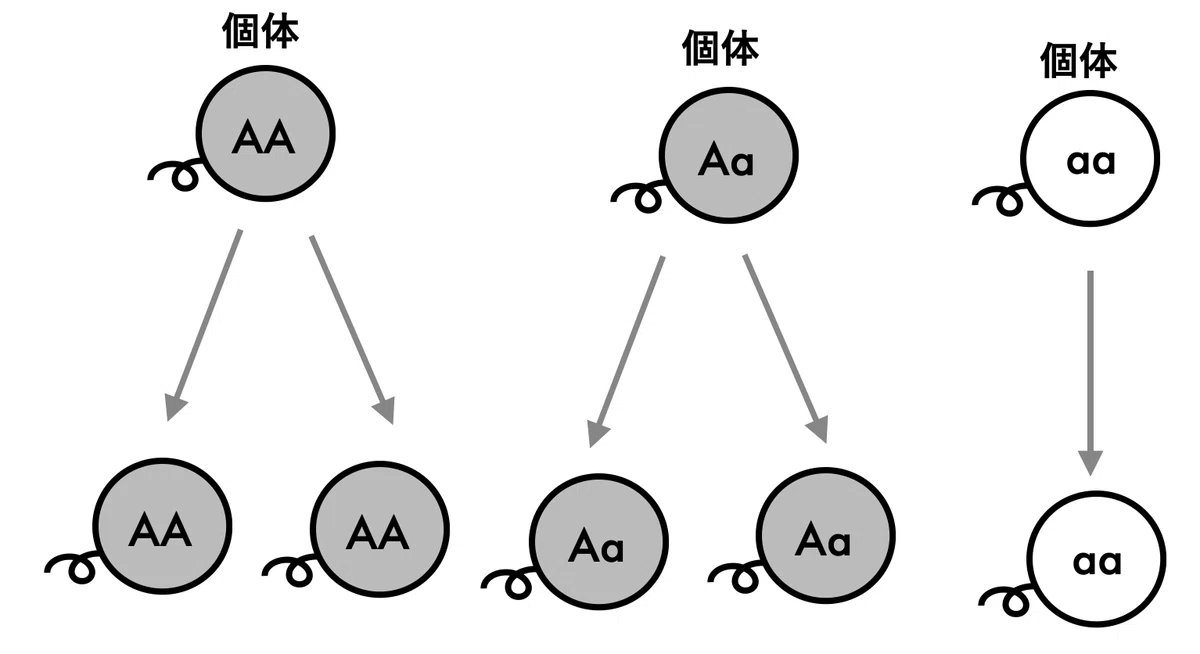
「利己的遺伝子」とは何かを考えるとき、自然選択の単位について理解する必要がある。まず、最も簡単な例でみてみよう(図1)。生物個体は、体色に影響する座位(遺伝子がある位置)に、2種類のアレル(対立遺伝子)Aとaがある。通常、父親から受け継いだ遺伝子と母親から引き継いだ遺伝子の2つのアレル(対立遺伝子)をもつので、個体はAA, Aaあるいはaaの組合わせでアレルをもつ(AA, Aaあるいはaaを遺伝型という)。遺伝型がAAあるいはAaの時に体色は灰色に、aaの時に白になる。個体の表現型(灰色か白か)によって、個体の適応度(一生に何匹の子どもを残すか)に影響し、灰色の場合、白よりも高い適応度を示す。そのために、灰色の個体はより多くの子どもを残し、アレルAが次世代に頻度が増加することで、灰色の個体の頻度が増加する。
この過程が、自然選択によって適応度を向上させる性質(灰色)が進化する(集団中で頻度を増大させる)しくみである。この場合、自然選択が働く単位を考えるとき、2つの自然選択に影響する要因を考えることが重要である。一つは、個体が生存や繁殖を通じて残す子どもの数に違いがでる、という点と、個体の体色(灰色か白色)が適応度の違いに影響する、という点である。そのため、自然選択の原因となる性質は、体色であり、これは個体の性質である。また、その結果、増減するのは個体の子どもを通してであり、個体ということになる。結果としてアレル(Aあるいはa)の頻度が変化する。そのために、このような自然選択は個体に働く、個体選択と呼ぶ。
多くの場合、個体の表現型が進化するのは、個体への選択によるものである。個体選択では、個体は、自らの子どもをより多く残すような性質を進化させるので、「利己的な個体が進化する」と比喩的に表現することがある。このプロセスを「遺伝子に働く選択」といわないのは、遺伝子が頻度を増加させているのは、個体の性質に依存して個体が増えた結果であるからだ。
次のような場合を考えてみよう。アレルAだけでなくBも同じ表現型である灰色の体色を作る。ことのとき、個体はアレルAをもっていてもBを持っていても同じ適応度を示す。そのため、Aが増大するかBが増大するかは偶然に左右される。個体にとっては、個体の性質が灰色であることが重要で、遺伝子がAであるかBであるかは関係がないことになる。
たとえば、アメリカの砂浜に住むネズミは、内陸にすむネズミと違って、白っぽい毛をもっている。砂浜では、白い毛をもつ個体が自然選択で有利に進化した。しかし、南海岸では、Mc1Rという遺伝子の変異が進化して白くなったが、東海岸では別の遺伝子が進化した結果であることがと示されている(4)。つまり、個体の性質である毛の色の進化には、毛を白くする遺伝子であれば、どの遺伝子でもいいことになる。また、身長など量的性質とよばれるものの違いには、多数のゲノム上の変異(アレル)が影響していることが示されている。たとえば、ヒトの身長には2万以上の変異が関わっている(5)。ヨーロッパでは、身長が高くなるように自然選択を受け進化したことが示されているが(6)、身長を高くする効果をもつ多数の変異(遺伝子)の頻度が増加したと推定される。同じ効果をもつ変異ならば、どの変異が頻度を上昇させるかは偶然に左右される。どのような変異であっても個体の性質である身長が高いかどうかが重要なのだ。
「利己的な遺伝子」という比喩からすると、自らのコピーを増やすように表現型を進化させた、ということになる。しかし、表現型の進化は、特定の遺伝子のコピーを増やさなくても、表現型に同じように影響するような遺伝子であれば、どれでもよい。この場合「利己的な遺伝子が個体の表現型を進化させた」という比喩的表現は不適切な使い方のように思える。
利他行動の進化は利己的遺伝子によるものか
生物の中には、自分の生存率を下げても、他個体の適応度を増加するように振る舞う「利他行動」をとる生物がいる。典型的なのが、アリやハチのように、繁殖して子どもを残すのは女王だけであり、同じ巣の他の個体は餌をとってきたり、巣を防衛したりするワーカーとなる。ワーカー個体は、自分の適応度をゼロにしても女王個体の適応度を増加させるように働く。
上記の個体選択の考え方からすると、利他行動を発現する遺伝型をもつ個体は、利他行動を発現しない個体に比べて適応度が低いので、個体選択では進化できないことになる。この問題に、答えを出したのが、W.H. ハミルトンだ(7)。彼は、利他行動を発現する遺伝子が、利他行動を享受した個体がより多くの子どもを残すことで増えると考えた。このとき利他行動を受ける側も利他行動を発現する遺伝子をもっている必要がある。兄弟や姉妹など血縁者の間では、同じ遺伝子を共有する可能性が高いので、利他行動を発現する遺伝子も共有している確率が高い。そこで、利他行動をする個体は、血縁者に対して利他行動をすることで、利他行動を発現する遺伝子を残していくという理論である(図2)。 この血縁選択の理論には、個体の包括適応度という概念がもちいられている。これは、その個体が利他行動をすることによる適応度の低下と血縁者から受ける利他行動による適応度の増加を足して表したものである。

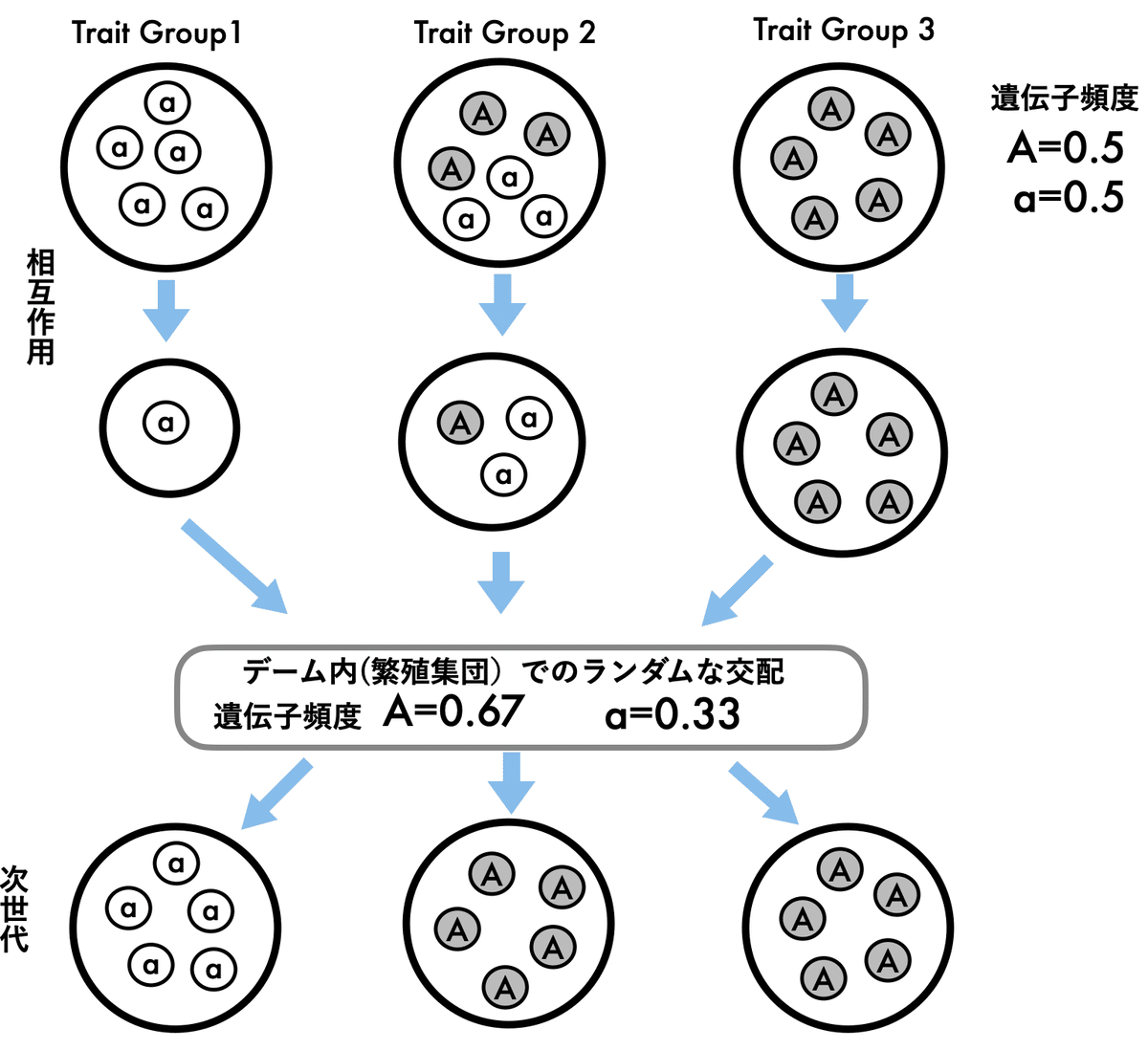
利他行動の進化を、別の見方でみてみよう。ここでも、他個体に対して、自分のコストを払って利他行動をする個体と、利他行動は享受するが利他行動をしない利己的個体を想定してみる(図3)。図3では、簡略化のために利他行動遺伝子をA, 利己行動遺伝子をaとして、利他行動をする個体を灰色、利己的行動をする個体を白としている。
生物個体は、コロニーや群れなどの密接に個体同士が相互作用している小集団(Trait groupという)で生活している。小集団の中で、利他行動をする個体の頻度が高い場合は、お互いに助けあうことで、繁殖するまでに生存する確率が増加する(Trait group 3)。利己的な個体(a)の頻度の高い小集団(Trait group 1)では、競争が激しく、生存率が低下する。利己的な個体と利他的な個体が共存している小集団では、利己的な個体は利他的な個体に比べて相対的に適応度が高いので集団内での頻度は増大するが、集団の個体数は減少する(Trait group 2)。
小集団で生き残った個体は、小集団外の個体と交配して次世代の子どもを残す(交配可能な繁殖集団をデームと呼ぶ) 。次世代では、また小集団に分かれて相互作用をする。このプロセスによって、次世代で利他行動を発現する遺伝子Aの頻度が増加する。
同じ小集団の中では、利他的個体は、利己的個体に比べて適応度が低くなるので、利己的個体が選択される(個体選択)。しかし、集団内での利他的個体の頻度(集団の性質)が高いと、集団サイズ(集団の性質)の低下が減少する。それが、利他行動を発現する遺伝子の頻度の増大に繋がる。つまり、小集団内においては個体にとって不利な利他的性質が進化できるのは、利他的個体の集団内頻度(集団の性質)が高いことによって集団サイズ(集団の性質)の低下の抑制(あるいは増加)
に影響するからである、と考えることができる。この場合の集団とは、繁殖集団(デーム)の中にある小集団(Trait group)であるので、このようなプロセスをデーム内集団選択という(8)。(デーム間集団選択という集団選択もあるがこれは「「種の保存のための進化」はどこが誤りなのか」を参照)
この集団選択が働くために重要な条件の一つが、小集団(Trait group)の間で利他行動を発現する遺伝子(アレル)の頻度が充分に異なっていること、その集団間で集団の適応度(増加率など)に違いがあることである。血縁個体が小集団を形成すると、利他行動を発現する遺伝子を持っている個体が小集団内にいる場合、同じ集団内の他個体も同様に同じ遺伝子を持っている可能性が高い。また、相互作用する相手が血縁者かどうかを識別できる場合は、小集団といったものを形成しなくても、血縁者間で相互作用することになり、血縁集団とみなすことができる。そのため、このような集団選択は、血縁選択と同じプロセスと考えることができる。実際に、血縁選択と集団選択が同じプロセスであるということは数学的にも証明されている(9) 。同じプロセスを違った視点でみているだけの違いであるようだ。「「種の保存のための進化」はどこが誤りなのか」の記事では、集団選択は限られた条件でのみ可能と説明した。血縁集団間の選択は、集団選択が働く条件を満たしているといえる。
個体の利他行動は個体にとって不利だが、利他行動を発現する遺伝子にとって有利なので、遺伝子が自分のコピーを残すために個体に利他行動をとらせている。このような利己的な遺伝子の見方は、利他行動がなぜ進化するのかを理解しやすくするかもしれない。利己的な遺伝子という概念は、利他行動が進化するのは、「種にとって有利」なのではなく、遺伝子にとって有利なため進化したといことで使われることが多い。
しかし、厳密にどのレベルのどの単位で選択が働いてるかを考えると少し違ってくる。利他行動を発現させている遺伝子は、その遺伝子をもつ個体が利他行動をすることで、自分の遺伝子も次世代にコピーを残す点で不利になっている。有利になっているのは、別の個体がもっている利他行動を発現する遺伝子である。血縁選択は、突然変異で利他行動遺伝子が生じ、そのコピーが集団に増えていくプロセスを示している。つまり、利他行動を実際に発現させた遺伝子が選択で有利になったわけではなく、利他行動遺伝子の集団が増加するのに有利になったと考えることができる。
利他行動の進化を、比喩的にどうみるといいのか?と言う問題は見方の問題なので、利己的な遺伝子の見方は概念的な捉え方で異なる。しかし、具体的な進化のプロセスをよくみてみると、利他行動を発現する遺伝子をもつ個体の集団に選択が働いたとみるほうが、より適確に表しているといえるだろう。なぜ適確かというと、以下で述べる遺伝子に働く選択で進化している「利己的遺伝子」と区別する必要があるからだ。
厳密な意味での利己的な遺伝子とは
自然選択のプロセスからみて、より比喩的にも適切な「利己的な遺伝子」とはどのようなものだろうか?自然選択の働き方から考えて見ると、遺伝子自身が、遺伝子のもっている性質によって、その遺伝子のコピーを増やすという過程が遺伝子に働く選択といえる。この遺伝子に働く選択は、個体がより多く子どもを残すことによって遺伝子が集団中に増えるプロセスとは、別のプロセスである。
減数分裂駆動という現象がある。2倍体の生物は、父親から引き継いだゲノムと母親から引き継いだゲノムのペアでもつ。特定の遺伝子座には2つのアレル(対立遺伝子)をもっていることになる。通常、次世代に引き継がれるとき、ペアのうちどちらかのアレルがランダムに選ばれて、一つだけが精子や卵子に受け渡される。しかし、減数分裂駆動という現象は、一方のアレルが、もう一方のアレルが精子や卵子に引き継がれるのを阻止しするのである。
減数分裂駆動を引き起こす遺伝子として、ショウジョウバエのSD遺伝子が有名である。このSD遺伝子とよばれるゲノム領域には複数の遺伝子が含まれている。SD遺伝子は精子が作られるとき、SD遺伝子をもっていない精子の機能不全を誘発するので、ヘテロ接合の雄(SD遺伝子をもっているアレルと持っていないアレルをペア持っているオス)の精子は、ほぼSD遺伝子を持つことになり、子どもはSD遺伝子を持つことになる(10)。これは、SD遺伝子が、ペアとなるもう一方の遺伝子の機能を直接あるいは間接的に阻害することによると考えらている。つまり、SD遺伝子は、その遺伝子がもっている性質のために、他の遺伝子の機能を阻害して、子どもの数を通してではなく、遺伝子の数をとおして増える仕組みである。
SD遺伝子は、個体の適応度を減少させる可能性がある。そのとき、遺伝子レベルでは、自然選択は有利に働き、SD遺伝子の頻度を増大させるが、個体レベルの選択では、頻度を減少させる方向に働いているといえる。つまり、SD遺伝子が増加、維持されている主な原因が遺伝子のもつ性質に起因する遺伝子レベルでの選択によるものなので、利己的遺伝子と呼ばれる。
利己的遺伝因子
ゲノム配列の中には、多数の転移因子(transposable elements)とよばれる配列が含まれている。簡単にいうとゲノムの他の場所に移動可能なDNA配列のことである。転移因子には、DNAトランスポゾンとレトロトランスポゾンがある。DNAトランスポゾンは、自分の配列をゲノムから切り出した、他の場所に挿入する。レトロトランスポゾンは、自分のDNA配列をRNAに転写したあと、RNAからDNAに逆転写し、他の場所に挿入する。つまりDNAトランスポゾンは、カット&ペーストで配列がゲノム中を移動し、レトロトランスポゾンはコピー&ペーストで、自分の配列をゲノム中に増やしていく。
これらの転移因子の配列がゲノム上の多くを占めている。たとえば、ヒトでは、ゲノム中の約半分が転移因子の配列だ。それらの転移因子の配列は、転移する能力を維持しているものもあれば、自ら再度転移する能力を失なってしまう場合も多い。このような転移因子は、レトロウィスルやバクテリアのゲノムを起源として進化したものや、生物のゲノム配列から転写されたRNAを起源とするものがある(11)。
転移因子は、自らのもつ配列により、自らの配列を移動したり、コピーして増やしたりするので、DNA配列そのものに選択が働いているとみることができる。そのために、転移因子は利己的遺伝子あるいは利己的DNAと呼ばれている。自らの配列や遺伝要素を増やしていくようなものには、上述した減数分裂駆動を引き起こす遺伝子、転移因子の他にも知られている。たとえば、ホーミングエンドヌクレアーゼ(相同な配列を切断し、自分の配列で置き換える)、B染色体(生物の生存や生殖に必要ではないが、通常の染色体とは別に存在する)、利己的なミトコンドリア、ゲノムインプリンティング(相同なアレルの一方の発現を抑制する)などがある(12)。これらのものは、「遺伝子」とはいえないものもあり、総称して、利己的遺伝因子(selfish genetic elements)と呼ばれている(12)。
遺伝因子と個体のコンフリクト
利己的遺伝因子は、個体の生存や繁殖を低下させことによって、個体レベルでの自然選択では不利になったとしても、それを上回るほど、自らの配列を増やすことで進化してきたと考えられる。遺伝子レベル(あるいはゲノムレベル)で働く自然選択と個体レベルで働く自然選択の利害が対立する状況を、利己的遺伝因子は作り出している。いわば、利己的遺伝因子とそれを持っている個体との関係は、寄生者と宿主の関係のようなものである。
転移因子が移動し、ゲノム上の新たな箇所に挿入されることで、元々あった遺伝子が破壊されたり、発現量が低下したりする可能性がある。また、転移因子が移動しなくても、繰り返し配列などの独自の配列をもっているために、突然変異率を上昇させたり、ゲノムの安定性を低下させたり、ゲノム配列の再構成の原因となる(11,13)。実際に、人では、転移因子は、精神疾患を含めた様々な病気や癌の原因になったりしている(13,14) 。
宿主となる個体は、転移因子の活動が増加すると個体の適応度が低下するために、転移因子の活動を阻止する様々な機構が進化している。たとえば、転移因子の配列をDNAメチル化することで活動を阻止したり、転移因子から転写されたRNAを切断する機構(RNAサイレンシング) が進化している。また、転移因子に特異的に結合するタンパク質などを進化させることで、転移因子のその活動を抑制している(11)。
多くの転移因子の活性化は配偶子形成または初期胚発生の様々な段階に限定されているようである(11)。つまり、転移因子にとって、体細胞で活動を増加させ、自分のコピーを増やしたとしても次世代には伝わらない。次世代に伝えられる生殖系列で、活性化することで、次世代に伝わるチャンスを増やしていると考えられる。一方、体細胞では、転移因子の活性は抑えられることが多い。体細胞系列での活性化は、宿主個体にとっても、利己的遺伝因子にとってもメリットがないからだろう。
利己的遺伝因子と宿主個体の共進化
寄生者と宿主の共進化により、しばしば寄生者が宿主にとって無害になったり、さらには有利になるような共生関係を進化させることがある。同様に、利己的遺伝因子も、ゲノム中の大半を占めるまでに増加しているということからも、個体の生存に中立になるように進化するか、ときには有利に働くように進化している。
利己的遺伝因子が新たに挿入されるゲノム上の位置が個体にとって重要な場所であれば、その遺伝子の機能を壊す可能性が高い。もし利己的遺伝因子が広がる効果よりも個体選択によって減少する効果の方が高ければ、利己的遺伝因子はゲノム上で維持できない。そのために、いくつかの転移因子では、コード領域などの少ないゲノム領域など、ゲノムの「安全な場所」を標的にする移動する高度な分子メカニズムを進化させている(13)。さらに、イネのトランスポゾンでは、タンパク質をコードしているエキソン領域に挿入することをさけ、遺伝子の上流部位(遺伝子の発現を調節する配列が位置する)をターゲットにして移動する転移因子もあるようだ。そのため、トランスポゾンの挿入は多くの場合、宿主個体に影響を及ぼさないか、上流部位に挿入された配列が、遺伝子の発現を増加させることが報告されている(15)。
転移因子は宿主のゲノムとは独立に複製や移動を可能にする遺伝子をコードしている。しかし、その遺伝子の発現は、宿主個体の生成物に依存しいている。たとえば、転移因子は自らの配列で移動などに必要な酵素を作るが、その遺伝子の発現を開始するためのプロモーター配列をもっている。プロモーターに転写因子(transcriptional factor)と呼ばれるタンパク質が結合することで、mRNAの転写が開始される。そのために、転移因子のプロモーターは、宿主個体のゲノムが使っている転写因子を利用するように進化している。そのため、宿主個体のゲノムの遺伝子の上流の遺伝子調節領域に、転移因子が挿入されると、挿入された転移因子のもつプロモーターが作用して、宿主個体の遺伝子が発現するようになることがある(13)。
通常は、宿主個体の遺伝子の調節領域に転移因子が挿入されると、遺伝子が機能しなくなったり、本来の宿主個体がコントロールしている遺伝子発現とは、違った発現をしめすようになったりして、宿主個体には不利にはたらく。しかし、挿入箇所によっては、宿主個体の適応度が増加するように遺伝子がコントロールされる場合が生じる可能性もある。たとえば、有名な適応進化の例として、イギリスの産業革命のときの大気汚染がもとで、暗い色の蛾が進化した工業暗化とよぼれる進化現象がある。蛾の翅色が暗い色に変化した突然変異個体が出現したのは、蝶の色素沈着や、鱗粉の発生速度を制御する遺伝子であるcortex遺伝子にトランスポゾン転移因子が挿入されたためである(16)。
利己的遺伝因子と宿主個体との長期的な共進化
ゲノムに挿入さた転移因子は、多くの場合、宿主個体に抑制され、転移因子の活性に必要な配列が失われ、ゲノム上に残骸として残る。しかし、それらの配列が宿主個体の機能に関与し、適応度が増加するように進化していく場合がある。上述したように、転移因子は、宿主個体の転写因子を利用するプロモータ配列をもっているので、その配列を基盤に、配列が進化することで、宿主個体にとって有利になるような遺伝子制御を進化させることができる。全くランダムな配列から機能的なプロモーターやエンハンサー(タンパク質をコードする領域から離れた場所にもある転写制御領域。転写因子が結合する)が作られるよりも、比較的簡単に進化が可能であると思われる。実際に、宿主個体は、転移因子の配列を利用して、遺伝子制御機構を進化させている。たとえば、哺乳類では、特定の転写因子が結合する部位の5〜40%、平均約20%が転移因子の配列と関係していると推定されている(13)。このような、もとの機能(転移因子のプロモーター)を別の機能(宿主個体の遺伝子の転写制御)に利用するようになる進化を、コオプション(cooption)とよんでいる(13)。
また、宿主ゲノムの様々な場所に挿入された転移因子の類似した配列が、プロモーターやエンハンサーとして働くことで、多数の遺伝子を制御する遺伝子ネットワークの構築が進化することもある(17)。たとえば、哺乳類の新皮質の進化において、前脳特異的に発現する遺伝子を調節するエンハンサーが転移因子によって進化した可能性が指摘されている(18)。また、マウスに特異的な転移因子の一種は、、過去1500万年から2500万年の間に何百もの胎盤特異的エンハンサーをゲノム中に増加させ、哺乳類の胎盤の急速な形態の多様化を反映していると考えられている(19)。
さらに。転移因子はプロモーターやエンハンサーなどの遺伝子制御の配列を提供するだけでなく、遺伝子発現を転写時や転写後に調節するノンコーディングRNA(タンパク質に翻訳されないRNAの総称、RNAマイクロRNA[miRNA]、ロングノンコーディングRNA[lncRNA]、サーキュラーRNA[circRNA]などがある)の起源にも関与している(13)。ヒト、マウス、ゼブラフィッシュにおいて、lncRNAの3分の2以上に転移因子由来の配列が含まれているという。これらのlncRNAは単にゲノムDNAをもとにRNAに転写されているだけで、宿主個体にとっては何の役にも立っていない可能性もある。しかし、少なくとも一部は、宿主の遺伝子発現を制御するように転移因子の配列が進化したと考えられる。
ゲノム全体に占める利己的遺伝子の進化
1980年に、DNAの構造を解明したクリックらを著者に含む、2つの論文がNatureに出版された(20,21)。そこでは、ゲノムの多くは利己的DNAあるいは利己的遺伝子であり、個体の生存には、有益でないジャンク(がらくた)なDNAであるという主張がなされた。そこでは、ゲノムの大きさと種の複雑さの相関関係がないこと(C値パラドクス)が、生物個体の適応度とは関係のないジャンクDNAがゲノムの大きさに影響しているからだという説明がされた。しかし、2012年にエンコードプロジェクトによって、ヒトゲノムの詳しい解析がだされ、直接タンパク質をコードするコーディング領域はゲノム中の1〜2%に過ぎないが、ゲノムのDNAの約76%が何らかの形でRNAに転写されていることが示された(22)。遺伝子調節領域を含め、ゲノムの80%が生化学的な機能に関連していると主張し、ヒトゲノムの大部分が「ジャンクDNA」であるという見解を否定した。
その見解に対しては、反論がだされ(たとえば23) 、RNAとして転写されていることと、何らかの「機能」をもっていることとは同じではないということが指摘された。本稿の転移因子の解説でもみてきたように、転移因子を起源とするノンコーディングRNAは、宿主個体の適応度を上げるような遺伝子調節に関連しているが、すべてではない。転写されているRNAの多くは、宿主個体にとっては役に立っていないかもしれない。今後は、転写されているRNAのどの程度が実際に宿主個体の生存や繁殖に有利に働いているのか、という点が問題になるだろう。
遺伝子レベルの選択と個体レベルの選択を区別することの重要性
「遺伝子は、自らのコピーを残すのに都合のよい個体の性質(表現型)を進化させた」というドーキンス流の利己的遺伝子の見方によると、多くの個体の性質は遺伝子レベルでの利益によって進化したことになる。本稿で述べたように、この見方には、遺伝子自体が自分のコピーを増やす場合だけでなく、個体に働く自然選択によって進化している場合も含まれてしまう。しかし、遺伝子にとっては有利な性質が個体にとっては不利なために進化を抑制される場合とか、個体によって有利なために、遺伝子の性質が利用され進化したというような場合を区別できないことになる。遺伝子レベルの選択と個体レベルの選択を区別して、相対的にそれぞれ異なる選択がどう働いてるかを理解する必要がある。
特に、上述した転移因子などの利己的遺伝因子などの進化を説明するときは、利己的遺伝子と宿主個体とのコンフリクトあるいは共進化がどのように生じたかを理解することが重要である。ゲノム中に存在する多数の転移因子やノンコーディングRNAなどが、なぜ進化し、維持されているのかを解明することで、ゲノム中に多数存在している遺伝子をコードしないゲノム配列の進化を理解することできると思われる。
生物進化のメカニズムを適切に理解するためには、遺伝子レベルで働く選択によって進化した利己的遺伝因子を限定して利己的遺伝子とよび、ドーキンス流の利己的遺伝子の見方は避けた方がよいだろう。
以下は、関連記事です。
- 進化における「適応」という言葉をめぐって
- 老化の進化:なぜ老化しない生物がいるのか?
-「種の保存のための進化」はどこが誤りなのか
引用文献
リチャード・ドーキンス (2018) 利己的な遺伝子 40周年記念版。 紀伊國屋書店. 日高敏隆他訳
Orgogozo, V., et al. (2016) The “Mendelian Gene” and the “Molecular Gene” Two Relevant Concepts of Genetic Units. Current Topics in Developmental Biology 119, 1–26.
Pearson, H. (2006) What is a gene? Nature 441, 398–401.
Barghi, N., et al. (2020). Polygenic adaptation: a unifying framework to understand positive selection. Nature Reviews Genetics 11, 1–13 .
Yengo, L. et al. (2022) A saturated map of common genetic variants associated with human height. Nature 610, 704–712.
Field, Y. et al. (2016) Detection of human adaptation during the past 2000 years. Science 354, 760–764.
Hamilton, W. D. (1964),The genetical evolution of social behaviour. I & II Journal of Theoretical Biology, 1-52.
Wilson, D. S. (1980) The Natural selection of Populations and Communities. Benjamin/Cummings.
Marshall, J. A. R.(2011) Group selection and kin selection: formally equivalent approaches. Trends in Ecology and Evolution 26, 325–332.
1.Larracuente, A. M. & Presgraves, D. C. (2012)The Selfish Segregation Distorter Gene Complex of Drosophila melanogaster. Genetics 192, 33–53.
Senft, A. D. & Macfarlan, T. S. (2021)Transposable elements shape the evolution of mammalian development. Nature Review Genetics 22, 691–711.
Ågren, J. A. & Clark, A. G. (2018) Selfish genetic elements. Plos Genetics 14, e1007700 .
Chuong, E. B., Elde, N. C. & Feschotte, C. Regulatory activities of transposable elements: from conflicts to benefits. Nature Reviews Genetics 18, 71–86 (2017).
1.Kazazian, H. H. & Moran, J. V. (2017) Mobile DNA in Health and Disease. New Engl J Med 377, 361–370 .
Naito, K. et al. (2009)Unexpected consequences of a sudden and massive transposon amplification on rice gene expression. Nature 461, 1130–1134.
Hof, A. E. et al. (2016) The industrial melanism mutation in British peppered moths is a transposable element. Nature 534, 102–105.
Sundaram, V. et al. (2014)Widespread contribution of transposable elements to the innovation of gene regulatory networks. Genome Research 24, 1963–1976 .
Notwell, J. H. et al. (2015). A family of transposable elements co-opted into developmental enhancers in the mouse neocortex. Nature Communication 6, 6644 .
Chuong, E. B. (2013) Retroviruses facilitate the rapid evolution of the mammalian placenta. Bioessays 35, 853–861.
Orgel LE, & Crick FH(1980 ) Selfish DNA: the ultimate parasite. Nature 284: 604–607.
Doolittle WF, & Sapienza C. (1980) Selfish genes, the phenotype paradigm and genome evolution. Nature 284: 601–603.
The ENCODE Project Consortium (2012) An integrated encyclopedia of DNA elements in the human genome. Nature 489, 57–74.
Doolittle, W. F. (2013) Is junk DNA bunk? A critique of ENCODE. PNAS 110, 5294–5300.
この記事が参加している募集
この記事が気に入ったらサポートをしてみませんか?