
20240630: 睡眠不足・認知能力・反応時間・急速眼球運動・スポーツ外傷

睡眠不足 (SD) は、部分的 SD と完全 SD に分けられます。部分的 SD は、睡眠時間が短くなったり中断されたりした夜を指し、完全 SD は通常の睡眠/覚醒サイクル中に少なくとも 1 晩眠らない状態を指します。完全 SD は頻繁に研究されていますが、これは、SD 期間が長くなると認知機能の低下が進むため、脳の活動のモニタリングが容易になるためと考えられます。研究によると、睡眠を長時間にわたって継続的に制限することは、1 晩の完全 SD と比較して有害です。
睡眠中、人は 3 つの NREM (非急速眼球運動) 睡眠段階と 1 つの REM (急速眼球運動) 睡眠段階を経験します。REM SD は興奮性ニューロンに顕著な影響を及ぼすようです。これは、起こりうる危険を評価し、脅威に関連する刺激に対する反応を処理するために不可欠です。NREM 睡眠不足は、特定の神経伝達物質の正常な放出を減らし、受容体がリフレッシュして感度を回復する能力に影響を与える可能性があります。これらの睡眠段階がないと、結果として認知力が低下します。
睡眠時間に影響する 2 つのプロセスは、概日リズムと恒常性プロセスです。概日リズム理論では、睡眠の開始と終了を引き起こす体内時計の調節を提唱しています。これにより、覚醒と睡眠のサイクル、およびさまざまなホルモンの分泌の制御が容易になります。恒常性プロセスは覚醒期間と睡眠期間に依存し、睡眠時間が不十分な場合は睡眠要件が増加します。SD が増加すると、脳の恒常性機能がますます損なわれます。脳はこれに対抗するために、ホルモンを波のように分泌して覚醒または睡眠を促します。2つのプロセスの相互作用により、睡眠/覚醒サイクルが調整されます。
SDの判断力と意思決定への影響
SD患者は、扁桃体と内側前頭前皮質(mPFC)との間の機能的結合を失っており、この領域は扁桃体への強い抑制投射を示すことで知られている。
さらに、青斑核と扁桃体の自律神経領域でより高度な結合関係が生じる

睡眠不足になると、扁桃体の大脳辺縁系の反応が増加し、否定的な感情的意味合いを持つ刺激が生じます。この扁桃体の活動レベルの変動は、睡眠不足時の mPFC 機能的連結性の喪失と関連しており、前頭葉抑制信号の減少を示唆しています。SD では、mPFC-扁桃体回路の機能不全により、脳の修正リセット反応が失われ、合理的な決定や社会的判断などの不適切な行動反応を引き起こします。SD患者の道徳的判断を調べた研究では、SD により反応潜時が長くなり、適切な行動方針を決定することがより困難になることが示唆されています。これは、SD により認知と感情を統合して道徳的判断を下す能力が損なわれるためです。
しかし、この研究では道徳的判断に関する議論の余地のある単一の評価手順が使用されており、結果の一般化が制限される可能性があります。
脳波スキャンは、倫理的判断と意思決定の発達において内側前頭前野が顕著な役割を果たしていることを示唆している。短期 SD 参加者の mPFC には独特で重要な変化が見られ、SD 中の認知機能の低下を mPFC がどのように補おうとしているかがさらに実証された。この領域の動きも SD 後に大幅に減少する。対照的に、ある研究では、短期 SD は前頭前野の機能に選択的に影響を与えないと主張した。しかし、この研究で実施されたすべてのテストは、主に脳損傷を調べるために臨床目的で作成された神経心理学的バッテリー テストから派生したものである。テストは短期 SD の影響を受けない天井効果を持つ可能性があり、参加者はすべて大学生であった。
スペースシャトル チャレンジャー号事故では、早朝勤務と睡眠不足に関連した判断ミスと人的ミスが認められ、打ち上げの決定は工学的判断に基づくべきであったと述べられています (V、ワシントン DC、1986 年)。これらのコメントにもかかわらず、通信と情報の交換の成功に関連する手続きに影響を与えた可能性のある他の要因も考慮する必要があります。
記憶に対するSDの影響
睡眠不足は、長期増強 (LTP) を通じて海馬での記憶の定着を阻害するようです。NMDA 受容体は記憶の定着段階に必要であり、記憶を不安定な状態からより永続的な状態へと進める役割を果たしますが、SD ではこれが阻害されます。Gais らは、NDMA 受容体が Ca 2+流入の増加を通じて LTP の発現を可能にすることを発見しました。SD は、Ca 2+流入の減少により、表面および受容体サブユニット構成の発現を変化させることで、グルタミン酸受容体の完全な活性化を阻害する可能性があります。SD ラットは、海馬シナプス刺激後に海馬細胞の NMDA/AMPA 比の低下を示しました。グルタミン酸またはチャネルの感受性に変化はありませんでしたが、架橋アッセイで SD 後に NMDA 受容体の NR2A および NR1 成分が細胞質に残っていることが示された後、SD が NMDAR 表面発現を変化させることが観察されました
海馬におけるタンパク質キナーゼ活性(PKA)および環状アデノシン一リン酸(cAMP)シグナル伝達の活性化は、記憶の強化に重要な役割を果たします。ホスホジエステラーゼ4A(PDE4A)は、海馬におけるcAMPの分解を担う酵素であり、タンパク質キナーゼA(PKA)活性の低下を引き起こします。SDは、PDE4およびPDE4A5タンパク質の活性の増強を伴います。PDE4シグナル伝達を阻害すると、LTPが増加し、海馬依存性の記憶固定におけるSD障害が減少します。PDE4阻害によるこの救済は、SDによるこの分子破壊が可塑性と行動の機能障害を引き起こすことを示しています。14ある研究では、SDがマウスの海馬におけるPDE4レベルを上昇させるかどうかを調査しました。SDマウスではPDE4特異的なcAMP分解が増加しましたが、非PDE4レベルは影響を受けないことがわかりました。SDによるPDE4Aの増加はcAMP細胞内シグナル伝達を阻害する。
SDはまた、記憶の定着に必要な翻訳調節タンパク質である哺乳類ラパマイシン標的(mTOR)シグナル伝達をダウンレギュレーションする。これは、タンパク質の翻訳が可塑性と長期記憶の発達に必要であることを示唆している。しかし、SD後の遺伝子転写とタンパク質合成の変化を調査するには、さらなる研究が必要である。
低レベルの SD は、タンパク質合成の低下につながる未折り畳みタンパク質反応 (UPR) を活性化するのに十分です。覚醒状態中に可塑性修飾が開始されると、タンパク質合成の必要性が高まりますが、SD では覚醒が生理的持続時間を超えると、タンパク質合成が損なわれます。SD でのシナプス伝達によるエネルギー消費が減少することは、徐波睡眠が、膜のリサイクル、グルタミン酸小胞の修復、タンパク質の翻訳、シナプス前貯蔵庫のカルシウムの修復、ミトコンドリアの休息など、脳細胞が多くの役割を果たすための間隔を意味している可能性があることを示しています。
SD の増加はこれらの機能の低下を引き起こし、記憶の定着に悪影響を及ぼす可能性があります。特に、若いマウスで RbAp48 タンパク質を阻害すると、海馬依存性の記憶障害が引き起こされました。fMRI 研究では、海馬体の機能不全によりヒストンのアセチル化が選択的に減少し、記憶喪失につながることが示されました。20 SDは RbAp48 などの特定のタンパク質のタンパク質合成を減少させ、記憶力の低下につながる可能性があると結論付けることができますが、この研究ではマウスにおける SD の影響を観察しなかったため、睡眠が記憶力の低下の主な原因であるかどうかを結論付けることはできませんでした。
側頭葉は、記憶の符号化と言語処理の役割を果たしています。睡眠不足は、脳が神経信号を最適な量で処理できないことを引き起こし、支離滅裂な発話の原因となります。研究によると、SD は、ヒスタミン、セロトニン、ノルエピネフリン神経伝達物質のダウンレギュレーションにおける NREM 睡眠の役割を阻害する可能性があります。これは、受容体が「休息」して感度を回復する能力を欠いていることを意味します。次に、ノルエピネフリン、セロトニン、ヒスタミンが最適に機能できず、自然なレベルで生成されないため、記憶の保存と日常の発話に障害が生じます。 SDは、フリーラジカルによって引き起こされた脳細胞の損傷を修復する酵素の不足にもつながり、これも記憶と発話に影響します。これは、長期の SD 患者の場合、絶え間ない脳活動によりニューロンが変性し始めるため、悪化する可能性があります。
しかし、Cirelli らは、ラットの長期 SD 後に脳細胞の変性の証拠が見つからず、ニューロンの変性の原因を突き止めるにはさらなる研究が必要であるという反対の結果を得ました。
注意力と覚醒度に対するSDの影響
睡眠状態では、タスク関連のデフォルトモードネットワーク(DMN)活動と前頭頭頂ネットワーク(FPN)活動の抑制作用が等しく、これは視床からの覚醒の増加によってサポートされています。この抑制が等しく、脳内で一貫した注意パフォーマンスがあることを意味します。DMNは、個人の外部タスクに応じてアクティブ化および非アクティブ化する脳領域の集合であり、FPNは注意に関連するネットワークと脳領域です。睡眠不足の状態では、タスク関連のDMNとFPMの活動の間に不均衡な抑制があり、視床活動に影響を与える覚醒活動が不規則に増加します。その結果、外部タスク中にDMN活動が不規則に乱れ、FPN活動が減少します。DMNを抑制することは、適切な脳ネットワークがタスクや目標に向かって成功する行動を達成するために不可欠であり、DMNを抑制しないと、個人は特定のタスクに注意を維持することができません。FPN活動とDMNの作用が変化すると、注意と集中のタスクは予測不可能になります。睡眠が得られると、視床からの覚醒入力が安定し、DMNとFPNの同等の活動が再び確立される。
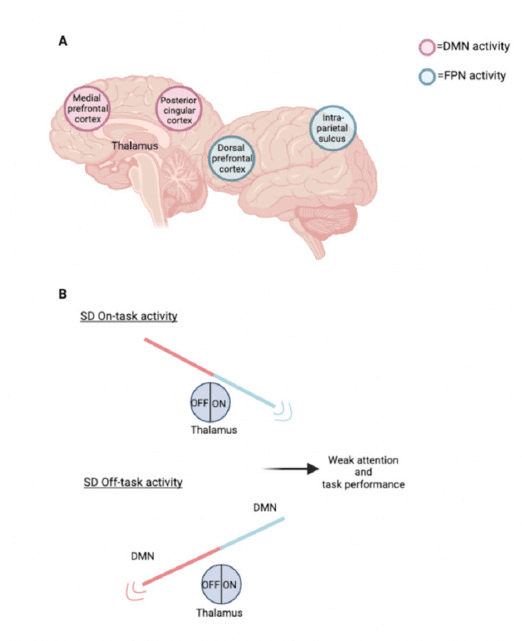
客観的注意に関する研究によると、SD では、画像選別の速度が遅くなり、不正確になるだけでなく、画像に対する記憶検出も低下しました。タスク関連活動に対する海馬傍回 (PPA) と前頭頭頂皮質の活性化は、SD では低下しました。タスクのサブ部分の効果的な統合と機能的結合のこの低下は、タスクを正常に実行する能力に影響します。参加者に与えられた視覚タスクでは、タスクの難易度が頭頂皮質の活性化、および島皮質、視覚皮質、帯状回の不活性化に関連していることが示されました。この活性化と不活性化のパターンは、SD 参加者の方が完全な睡眠のグループよりもかなり低かったです。これらの複合要因により、正確な注意パフォーマンスに不可欠な注意ネットワークに障害が発生し、日常生活でのリスクや事故に対する脆弱性が高まる可能性があります。PETスキャンでは、注意力と覚醒に関連する視床と前頭前野で主として脳活動が低下していることが示されました。
これは、主に視床と前頭前野および後頭頂皮質で観察される相対的な局所的グルコースの大幅な減少によって説明でき、覚醒度と注意力が睡眠不足による疲労の増加によって間接的に影響を受けることを示しています。しかし、対照的に、Kuhn らは SD 患者のグルコースレベルの上昇を発見し、SD 患者の注意力と覚醒度に対するグルコースレベルの影響を確認するにはさらなる研究が必要であることを示しています。SD 状態では、タスク関連の DMN および FPN 活動の抑制が不安定で、視床での覚醒影響活動が一貫して増加しません。これにより、タスク中に DMN 活動の不規則な信号と FPN 活動の低下が発生します。これにより、注意力と作業記憶機能の低下を引き起こす可能性があり、視床活動の増加とともに改善し、視床活動の低下とともに改善しなくなります。
ラットの脳波表示は、SD により、睡眠のゆっくりとした振動に関連する OFF 期間と同じ沈黙期間を示す皮質ニューロンが増えることを示しています。覚醒中のこれらの OFF 期間は、過分極傾向の増加の結果として神経双安定性が高まった結果である可能性があります。この過分極により、ニューロンの膜電位がより負になり、強力な覚醒可塑性によるシナプス余剰の結果である可能性があります。これにより、細胞外アデノシンの増加によって示される供給と需要のエネルギーの不均衡につながる可能性があります。
オフ段階が睡眠の治癒的役割、すなわちシナプス恒常性維持のいくつかを担うことができるかどうかは不明である。しかし、覚醒期間中のオフ期間の存在は興味深い説明を提供する。例えば、SD 中の注意力と特定の認知機能の分離は、覚醒皮質下促進システムにおける局所的な睡眠の発生と、特定の皮質領域における追加期間によって説明できる。さらに、覚醒を制御する脳幹および視床下部神経細胞で局所的な睡眠中の覚醒が発生した場合、SD 後の注意力の全体的な障害と眠気の増大の原因を説明できる可能性がある。
特に、それほど要求が厳しくなく、興味のない活動の場合である。SD 被験者の注意力と覚醒度の全体的なパフォーマンスのテストは非常に不安定で、正常レベルから危険なエラーまでの範囲である。
シナプス恒常性仮説
シナプス恒常性仮説 (SHY) は、NREM 睡眠の徐波段階では、脳全体のシナプスが減少することで、学習などの覚醒時のネットワーク シナプスの純強化が打ち消されると提唱しています。覚醒中は、周囲の雰囲気を吸収するために、脳内のネットワークを固める必要があります。これには、エネルギー、学習の強化、信号対雑音比の低下のための細胞プロセスの増加が必要です。タスクによって駆動されない神経活動である自発活動は、睡眠中のシナプス強度を正常化し、細胞の恒常性を回復することができます。

シナプスの強度低下は、シナプス興奮のネットワーク効果と覚醒期間後のニューロン活動の増加を打ち消す際にエネルギーが節約されるため、睡眠が記憶の獲得と強化に及ぼす利点を説明できます。シナプス強度の増幅に続いて、異なる結果が生じる可能性があり、これにはエネルギー利用の高速化、細胞にストレスを与えるシナプスへの細胞成分の分配の必要性の増加、およびグリア支持細胞の変化が含まれます。このシナプス力の増加は、ニューロン反応の選択性を弱め、情報を吸収するスキルを強化することもできます。シナプスの力を正常化しないと、SD は神経細胞の可塑性の重みを増大させ、ニューロンの選択性と学習能力を再構築できず、信号対雑音比の向上ができなくなります。SD が続くと、睡眠不足により、通常の覚醒中に起こる可塑性変化によって細胞とシナプスの恒常性を再構築する脳の能力が低下し、記憶の強化と統合が起こります。
シナプス再正規化は、シナプスネットワークの脱飽和、エネルギー供給の強化、細胞圧の低下を通じて、学習体験と記憶の保持を改善するなど、睡眠の多くの利点を示します。たとえば、軽い SD の夜の後には、新しい画像のコーディングが損なわれ、全体的な SD の減少を除いて、徐波活動が減少します。ただし、Gais らは fMRI 研究を通じて、宣言的記憶は長期 SD の影響を受けないと結論付け、Voderholzer らは、長期 SD は青年の長期宣言的記憶に影響を与えないと結論付けました。
同様に、訓練中に反応時間に変化がないにもかかわらず、訓練課題をSD後に行うとエピソード記憶の保持が大幅に低下する可能性があり、これはSDによるコーディング能力の低下を示唆している。これとは対照的に、経頭蓋刺激と模擬刺激によって低速振動が促進された短い睡眠の後には、絵、単語ペア、リストの暗号化能力の向上が観察された。これは、前の覚醒期間中に飽和に向かって増強された海馬のシナプスネットワークの縮小によるものと考えられる。
脳のグリンパティックシステム
グリンパティック システムは、睡眠中に中枢神経系から老廃物を除去する血管の複合体で構成されています。これは、脳脊髄液 (CSF) が動脈傍領域に移動し、アクアポリン 4 (APQ4) 水チャネル ルートを横切って間質領域に移動するときに発生します。睡眠段階中のリズミカルな脈動により、間質液 (ISF) と CSF の間で交換が発生します。未使用の産物は動脈から静脈に移動し、静脈傍領域に移動します。
水チャネル経路である AQP4 の発現が低下すると、ベータアミロイドと呼ばれるペプチドがさらに分泌される可能性が高まります。睡眠状態と覚醒状態では、グリンパティックシステムの役割が異なります。睡眠状態の間、脳脊髄液の流量が増加し、間質腔が 60% 増加し、毒性のある細胞分子を効果的に除去できます。睡眠状態の後、脳脊髄液によってベータアミロイドのレベルが低減します。ベータアミロイドは、認知症やアルツハイマー病など、多くの神経変性疾患の主な原因です。これは、睡眠不足がグリンパティックシステムの活動プロセスを低下させ、毒素の蓄積につながり、認知能力、運動機能、行動パターンに悪影響を与える可能性があることを示しています。

グリンパティック活性の低下は、脳内のアポリポタンパク質 E 循環の調節不全、グルコース代謝の調節不全、インスリン抵抗性にもつながり、認知機能の低下につながります。時間の経過とともに、これらの代謝物の蓄積と調節不全は、重篤な脳合併症を引き起こす可能性があります。トレーサーを使用した PET スキャンを使用して、マウスの通常の睡眠と睡眠不足によるアミロイドベータの量を調べました。 1 晩の比較では、マウスの視床と海馬のベータアミロイドレベルが大幅に増加していることが示され、睡眠不足が既知の神経変性プロセスに与える影響の生体内証拠が示されました。
これらの結果にもかかわらず、PET スキャン技術では可溶性ベータアミロイドと不溶性ベータアミロイドを区別できなかったため、結果に影響が出た可能性がある。不溶性ベータアミロイドよりも可溶性ベータアミロイドの方がアルツハイマー病などの神経変性疾患の予測因子としてより有効だからである。高齢の参加者を対象とした別の研究では、ベータアミロイドといくつかの認知領域のパフォーマンスとの間に逆相関関係があり、ベータアミロイドが増加することが判明した。これは SD 患者のグリンパティック システムの欠陥を示している可能性があり、これが全般的認知、言語流暢性、対決呼称、言語学習のパフォーマンス低下の原因となっている。
しかし、参加者の SD は研究されていないため、ベータアミロイドの増加が本当にグリンパティックシステムの障害、特に睡眠不足によるものなのかどうかを結論付けるには、さらなる研究が必要です。メルツァーらの研究の反対の証拠では、ベータアミロイドの沈着と認知能力の低下との関連が見つからず、認知障害の主な原因となる可能性のある他の要因があることを示唆しています。
睡眠は、健康的なライフスタイルに欠かせない要素である
ことは間違いありません。私たちの体を一貫した覚醒と睡眠のサイクルに保つために、脳は絶えず神経ホルモンを分泌しています。SD の悪影響は、全体的な行動と認知能力に明らかです。神経経路が遅くなり、反応時間と精神状態が低下します。脳が再び刺激されるまで、体内のシステムは対処手段として生命維持状態に入ります。視床活動、シナプス再正常化、グリンパティック システムの役割、DMN 活動、扁桃体活動、海馬活動の変動により、脳に不均等な刺激が生じ、SD を管理するために脳が不規則な活動を起こす可能性があります。その結果、注意力、作業記憶、記憶の統合、警戒、判断、意思決定、その他多くの認知能力の低下が起こります。特定の人に必要な量の睡眠をとった後、脳は反応時間と知的活動が向上し、有利になります。 SD の正確なメカニズムとその後の影響の妥当性を証明するには、さらなる研究が必要ですが、これはより多くのリソース、研究、時間があれば達成できます。それにもかかわらず、現在の証拠は、健康状態に関係なく、毎日または数日にわたって睡眠不足になると、体のシステムが徐々に衰退することを示しています。人間の心と体は、十分な睡眠をとり、認知能力を最適な状態に保つことで脳機能を最大限に高めるために、十分な NREM 睡眠段階と REM 睡眠段階を必要とします。
まとめ
睡眠は完全には理解されていないものの、人体が最高のパフォーマンスを発揮できるように毎晩規則的な周期プログラムに従う、重要かつ組織化された一連のイベントとして認められています。睡眠不足、つまり睡眠不足 (SD) は、認知能力に悪影響を与える可能性のある広く見られる現象です。このレビューでは、SD が認知能力に及ぼす可能性のあるさまざまな影響を調査する研究だけでなく、生物学的な説明にも重点を置いています。睡眠不足は、脳の記憶、注意、覚醒、判断、意思決定、および全体的な認知能力への影響とは無関係に起こるわけではなく、機能の低下と認知能力の低下をもたらします。
この記事が気に入ったらサポートをしてみませんか?