
生命は多様性を目指す(遺伝子決定論は正しいか? ④)

人為淘汰・品種改良の問題点
ダーウィンの自然淘汰説も優生学のどちらも、 作物や家畜の品種改良から発想のアイデアを得ていた訳ですが、人為淘汰による家畜の品種改良には重大な問題があるのです。育種家には良く知られている事ですが、品種改良には限界があり、特定の性質を伸ばそうと代々選択して行っても、最初の間は順調に進んでも直ぐに限界に達して、それ以上の改良は不可能となるのです。さらに選択を続けると、遂にはその系統は不妊となり死滅してしまいます。いわば、種の障壁とも言うべき限界が存在して、人為選択による品種改良は、その限定された許容範囲内でしか有効ではないのです。種には、ある決まった範囲の変異性が存在し、それが人為的に一方向に極端に推し進められると、別の面に歪みが生じると考える事もできます。ですからその限度内では多くの品種改良が行われてきたとはいえ、小麦は依然として小麦であって、それがグレープフルーツになったりはしないわけです。例えば、 人間は 1万年以上にも渡ってイヌの育種を続け、 様々な品種を作り出してきましたが、イヌの品種は全て交雑可能な一つの種に属したままです。作物や家畜の品種改良は、様々な品種を生み出してきましたが、一つとして新しい種が誕生した訳では無いのです。 つまり、人為淘汰によっては進化は起きないのです。
人為淘汰による品種改良には、もう一つ重要な問題が存在します。 ダーウィンは、イギリスの育種家がイヌやウマの新品種を作り出す方法を研究しましたが、このような品評会に出す動物を作る育種には特有の問題が有ります。育種家は、品評会で優勝した親にそっくりな子を作ろうと、動物の母と子、あるいは兄弟同士を掛け合わせる同系(近親)交配を行う結果、非常に深刻な遺伝病や気性の悪さ等の問題が子孫に急増するのです。これに対し、家畜やその他の使役動物では、身体的にも精神的にも融通性が高く、生命力の旺盛な形質を維持するために異系交配、つまり類縁の遠い系統間での交配が利用されます。これは農業用の植物でも同じで、これらの動植物は、多様な環境条件の下でも生命力や持続力・適応性を発揮できる様に育種されるのです。「実際、近代農業の基盤となった雑種強勢は、皆このような異系交配によって作り出されたもの」(『科学の危機』矢沢サイエンスオフィス)なのです。
しかし、こうした問題は、遺伝子の性質を考えればむしろ当然の事と言えます。 遺伝子は、タンパク質のアミノ酸配列をコードしたものでした。 つまり遺伝子とは、一つのタンパク質の設計図に過ぎず、 遺伝子単独では意味をなさないのです。個々の遺伝子は他の遺伝子とネットワークを組み、協調して働く事で、初めて有効に機能する事が出来る訳です。そして、遺伝子と形質とは一対一で対応しているのではなく、一つの形質の決定に多くの遺伝子が関与しています。 それと同時に、一つの遺伝子はそのネットワークを通じて、様々な形質の決定に関与する事にもなります。このように個々の遺伝子ではなく、遺伝子間に張り巡らされた複雑で錯綜した相互作用のネットワークこそが重要な意味を持っており、そこにこそ生命の秘密が隠されているのです。その結果、特定の形質を強化しようと人為選択を繰り返して行くと、遺伝子ネットワークの他の部分で無理が生じ、その歪みが問題を引き起こす事になるのはむしろ当然なのです。
反社会的傾向の家系?と優生結婚
また、品評会用の育種での近親交配の問題は、優生学とも関係しています。優生学では、劣った遺伝形質の持ち主との結婚を避けると言う目的で、結婚相手の家系・血統を重視します。反社会的行動や精神障害などが、遺伝によって家系に伝えられると考え、優れた遺伝的形質を持つ血統を守る為には、劣った家系を避けなけらばならないという訳です。実際、優生運動が隆盛を極めた19世紀末から20世紀初頭のアメリカでは、反社会的傾向の家系の研究という、怪しげな調査が発表され注目を集めています。 ジューク家の調査では、709人の子孫の内、181人が売春に従事し、106人が非嫡出子、142人が乞食、3人が犯罪者となっており、ここから「貧困は遺伝性」だと決め付けています。またカリカック家の調査では、マーチン・カリカックと酒場の女性との間から生まれた 480人の子孫を調査し、そのうち143人が精神障害者、36人が非嫡出子、33人が性的放埓、24人がアルコール依存症だったとしました。一方、後の正しい家柄の女性との結婚からは、社会的な有力な人物が続出したと言うのです。そして「精神薄弱は他の形質と同じように遺伝する」という結論を引き出しています。反社会的行動や精神薄弱が遺伝によるのであれば、そのような劣等な遺伝因子を持つ家系は子供を作らせない様にして、有害な遺伝子は絶滅すべきだと言う事になってしまいます。
実は、日本の保健体育の教科書でも、遺伝的欠陥のある家系は子孫を残すべきでは無いとする、次のような記述が有るのです。(『「現代優生学」の脅威』池田清彦著より引用)
国民の素質を向上させるという優生結婚の立場から、結婚をするにあたって、自らの家系の遺伝病患者の有無を確かめると共に、相手の家系についてもよく確認することが重要である。家系の調査範囲は、両親・兄妹姉妹はもとより、祖父母やいとこまで及ぶことが望ましい。(『保健体育改訂版』一橋出版1971年)
優生上問題になる疾病や異常の遺伝を防ぐために、優生保護や優生結婚が必要となってくる。相互の家系に遺伝的欠陥や疾病がある場合には、なるべく結婚を避け、不健全な子孫が生まれたり、その家系だけでなく社会的にも不幸を招くような疾病のある人は、子孫を残さないようにしなければならない。 (『標準高等保健体育』講談社1975年)
驚いた事に、日本では戦後数十年にもわたって、優生学というエセ科学に基づいて、子供に障害者とその家系に対する差別と偏見を煽るような教育を、 公然と行なって来たのです。上辺だけの綺麗事を並べ立ててきた、戦後民主教育とは一体何であったのか、我々は真摯に検証し直す必要があるでしょう。
近親婚による遺伝的劣化
遺伝的欠陥のある家系との結婚を避けると言う事は、裏返せば、遺伝的に優れた家系との結婚を勧める事に繋がります。実際、優れた血統への憧憬と欲求は、既にプラトンに現れています。プラトンは「優れた血統の交配により、哲人を生み出すべきだ」 と考えていたと言います。そして、優れた家系・血統との結婚への欲望・固執は、必然的に近親婚に行き着きます。これは支配階級の権力者に顕著で、 自らの血統の純潔性を保つ事を優先する結果、近親婚へと傾斜して行くのです。古代エジプトのファラオの近親婚は有名で、エジプト第 19王朝のラムセス 2世は、自分の娘達と結婚したと言われます。また世界三大美女の一人で、カエサルとアントニウスを手玉に取ったクレオパトラ 7世は、弟のプトレマイオス13世と結婚して共同で王位に就いていました。古代の日本においても、 王族・皇族の異母兄弟姉妹婚・叔姪婚・いとこ婚などの近親婚の例が数多く記録されています。天皇家と蘇我氏、平安時代以降の藤原氏との婚姻も、近親婚の連続でした。
図23)ラムセス 2世

(出典:ウィキメディア・コモンズ )
しかし、このような近親婚を繰り返すと、近交弱勢として知られる、遺伝的劣化が生じる事になります。例えば、天武天皇は、兄の天智天皇の皇女で後の持統天皇を皇后としましたが、皇位を継がせようとした子の草壁皇子、孫の文武天皇も、近親婚による虚弱体質の結果、早死にしたと言われます。また、文武天皇の第一皇子の聖武天皇は、亡くなったのは 50代ですが、生涯病弱であったとされます。
中世ヨーロッパにおいても王族・貴族間で近親婚が頻繁に行われ、スペイン・ハプスブルク朝は血族同士の結婚を繰り返しで、17世紀末には虚弱な人物ばかりが誕生し、断絶するに至った事が知られています。ハプスブルク家最後のスペイン国王カルロス 2世は、出生時から病弱であり、当時は「呪いをかけられたため」と噂されていたと言います。またカルロス 2世は、先端巨大症を患っており、「ハプスブルク家の顎」として知られる下顎前突症が著しく、咀嚼にも影響して、常によだれを垂れ流していました。さらに、癲癇や知的障害なども併発して、片足を引きずって歩いていたとされます。幼少期には衣服を身につけた動物のようで、まともな教育を受けることも困難であったと言うのです。また、同母姉でベラスケスの肖像画で知られるマルガリータ王女は、父方の従兄・母方の叔父にあたるレオポルト1世と結婚していますが、夫妻の間に生まれた4人の子の内、3人が1歳未満で夭折しています。
図24)スペイン国王カルロス 2世

(出典:ウィキメディア・コモンズ )
つまり、優生学者が勧める優れた家系・血統との結婚は、遺伝的多様性を低下させ、近親婚を誘発して却って遺伝的形質を劣化させてしまうのです。自分たち優れた家系・血統の者だけが持つ優秀な遺伝子を守る為には、社会階層の低い劣った人間との結婚を避けなければならないと言う、思い上がったエリート意識・選民意識が優生学というエセ科学を生み出し、却って血統の遺伝的形質を劣化させていたと言う訳です。支配階級の思い上がりが生み出した笑い話であり、愚か者の茶番劇と言うべきでしょう。
繰り返し述べてきました様に、遺伝子の機能はそのネットワークによって支えられています。そして、遺伝子ネットワークの機能を高く維持する為には、構成要素である遺伝子の多様性が決定的に重要なのです。しかし、優生学の主張する劣った遺伝子の排除は、反対に遺伝的多様性を減退させてしまう訳です。そして、その行き着く先が近親婚による家系の遺伝的劣化であった訳です。
最初に、私たちの遺伝子はゲノムのわずか1.2%に過ぎず、そのほとんどが機能不明のジャンク DNAと見られていたと述べました。 実際、ゲノム中には2万個以上存在すると言われる偽遺伝子や、レトロウイルスが持ち込んだ遺伝子など、明らかに何の役にも立っていない様に見える DNA 配列が大量に存在しています。もし自然界では、劣った遺伝的形質を持つ精神障害者や犯罪者が自然淘汰により排除されると言うなら、ゲノムに於いても役に立たない DNA 配列は当然に自然淘汰により除去されるはずです。細胞分裂時には、このような無駄な DNA 配列も複製される訳で、細胞にとっては負担になっているはずだからです。ところが実際は、役に立つ遺伝子はわずか 1.2%で、反対に何の役に立っているのか、良く分からない DNA 配列の方が圧倒的に多い訳です。つまり、自然は優生学者が言う様に、役に立たないとか劣っているなどの基準で選別して、自然淘汰で多様性を減退させる様な愚かな真似はしていない訳です。自然はむしろ、多くの無駄を抱え込み、多少の負担を負う事になっても、遺伝的多様性を維持する事の方を優先しているのです。
自然は多様性を目指す
生物が遺伝的多様性への強い指向を持っている事を示す、端的な例が有性生殖、つまり性の存在です。ヒトも含めて高等動物では、性は子孫を作る為の唯一の手段であって、性と生殖を切り離して考える事は難しいでしょう。しかし本来、性は生殖と不可分なものとして誕生したのでは有りません。実は、性は生物の増殖にとって必ずしも必要なものでは無いのです。例えば、植物には地下茎やむかごなどで、無性的に増殖できるものが多く存在します。動物界でも、刺胞動物のヒドラは出芽によって子孫を作り、イソギンチャクや海産の蠕形動物は2つに分裂する事で増殖して行きます。ミツバチやアリマキでは、オス無しの単為生殖が見られます。また単細胞生物では、多くの場合、性と生殖は完全に分離しています。バクテリアは普通、無性的に分裂を繰り返す事で急速に増殖します。このように一部の生物では、オスとメスが存在しなくても充分に生殖が可能なのです。
しかも子孫を殖やす、つまり増殖という点に関しては、こうした性を介さない無性生殖の方が、有性生殖に比べて格段に効率的なのです。例えば、分裂によって殖える大腸菌は、条件さえ良ければ 20分に1回の割合で分裂し、その結果、1個の大腸菌が9時間で1億個体にまで増殖します。植物の新芽などにたかるアブラムシも、春から夏にかけては交尾をせずにメス単独で子供を生みますが、その繁殖力は凄まじく、数日で植物がアブラムシで覆われてしまいます。しかし考えて見れば、無性生殖が旺盛な繁殖力を持つのは当然で、反対に我々に身近な有性生殖の方があまりに手間ヒマを掛け過ぎているのです。有性生殖をする為には、まずオスとメスが出会って求愛し、交尾して卵子と精子を受精させなければなりません。被子植物では、受粉を助けてくれる昆虫を引き寄せる為に花や密を作り、裸子植物では風に飛ばす大量の花粉が必要となります。有性生殖は、大変な手間とコストの掛かる生殖方法なのです。単に増殖だけが目的ならば、性は不必要と言うよりも、むしろ邪魔な存在なのです。しかし、大多数の植物や動物は有性生殖で増殖しており、また多くの原核生物や通常は無性的に増殖する生物でも、特定の時期には有性生殖を行っています。では何故、生物はこれほどまでに手間とコストを掛けて、非効率な有性生殖を行っているのでしょうか。それこそ、遺伝的多様性の獲得の為なのです。 有性生殖ではオスとメスの染色体が混ぜ合わされる結果、それだけで驚くべき多様性が生み出されます。 ヒトの場合、23組の相同染色体を持つために、配偶子当たりで 2の 23乗(約840万)通り、子供ではさらに 2乗の約 70兆通りもの遺伝的組み合わせが可能となるのです。
一方、無性生殖で生まれる子は、総て親と全く同じ遺伝情報を持つクローンです。1匹の大腸菌は、分裂を繰り返して急激に個体数を増やしコロニーを形成しますが、そこにひしめく大量の大腸菌は、突然変異を起こしたものを除くと、総てが遺伝的には全く同じコピーに過ぎ無いのです。ところが有性生殖では、生まれて来る子は総て異なった遺伝情報を持っており、全人類の中でも、一卵性双生児を除けば 2人と同じ遺伝情報を持つ個体はいない訳です。有性生殖をする生物は、総て遺伝的に異なる、この世に2つとない唯一無二の存在なのです。
異質を求める性
また有性生殖をする生物は、遺伝的多様性の拡大をより確実なものにする為に、できるだけ遺伝的に異なる生殖相手を求める性向を持っています。「性では常に異質を求める」という原則があるのです。
例えば、大腸菌の接合による交配は異なる株間でしか起こりません。また、真核生物のゾウリムシや酵母の性では、同じ形・大きさの細胞同士で接合が行われ、性の違いは接合型とか交配型と呼ばれる番号や記号で表わされますが(ゾウリムシではⅠとⅡ、酵母ではaとα、藻類では+と-)、こうした仲間でも自分と異なる接合型のものとしか接合できません。つまり、自分と同じ接合型のものとは接合できないのです。しかも、ゾウリムシの属する繊毛虫の仲間には、接合型をたくさん持つ種が少なく有りません。単細胞の緑藻が共生して、美しい緑色をしたミドリゾウリムシでは、性すなわち接合型が4種類とか8種類も有るものもいます。特に多いのはキノコの仲間で、例えばスエヒロタケでは、遺伝的には4000以上の接合型がある計算になると言います。
我々に馴染みの深い種子植物が、多量の花粉を作って風に飛ばしたり、美しい花と密で昆虫を誘って花粉を運んでもらうのも、自分と遺伝的に異なる相手と有性生殖する為です。多くの植物は1つの花に雌しべと雄しべを持ち、自分の花粉で自家受粉している様にも見えます。しかし良く調べて見ると、こうした植物でも同じ花の中ではできるだけ受粉・受精をしない様に工夫している事が分かります。多くの花で、雌しべは雄しべよりも長く、雄しべの先のやくが破れて花粉がこぼれ出ても、雌しべの先には付きにくい構造になっています。これらの花の雌しべは、風や昆虫によって運ばれて来る外来の花の花粉を、優先的に受け取る様に作られているのです。また、雌しべと雄しべの熟す時期が異なる為に、同じ花の中では自家受粉できない花も多いのです。モクレンやオオバコの雌しべは、雄しべより先に熟して、花粉が出る頃にはしおれてしまいます。反対にユキノシタやホウセンカでは、雄しべの方が先に熟して花粉を放出し、雌しべが熟す時には花粉は無くなっています。小さな花がたくさん集まって1つの花の様になったキク科の花も、雌の時期と雄の時期を分け自家受粉を防いでいます。例えば、ヒマワリは周囲の1枚の花弁を持つ舌状花(雄しべと雌しべは退化)と、中央に集合した筒状花から出来ていますが、筒状花は外側のものから内側へと咲いて行き、その間に雄期から雌期へと変化して行きます。つまり、筒状花の中心部はつぼみ、その周囲に花粉を出している雄期の花、一番外側を雌しべの柱頭を出した雌期の花が取り巻いているのです。
もっと手のこんだ植物も有ります。マツバボタンやオシロイバナは、開花当初には雌しべと雄しべが離れていて自家受粉し難く、昆虫などが運んで来る他の花の花粉を受けやすい状態になっていますが、花がしおれる直前になると、雌しべと雄しべが接近し自家受粉してしまいます。これらの花は、できるだけ他家受粉に努めるが、それに失敗した時には自分の花粉で自家受粉する様にできているのです。植物の中には、もっと徹底して自家受粉を避ける仕組みを持つものも有ります。それは、リンゴ・ナシ・ブドウなどの果樹の多くで知られる自家不和合性と呼ばれるもので、同じ株の花粉では受精がうまく行かず実を結ばないのです。一般に、花粉が雌しべの先に付くと発芽して花粉管を出し、これが雌しべの中を伸びてその付け根にある小室に到達すると、花粉管から精核が放出され小室内の卵細胞と合体して受精します。ところが自家不和合性の植物では、雌しべが自分の花粉と他株の花粉とを区別し、自分の花粉では花粉の不発芽、花粉管の花柱への侵入不能、花粉管の成長速度の低下や停止などによって、受精が正常に行われない仕組みになっているのです。また、こうした不和合性が花の形状と結びついている場合も有ります。サクラソウでは雌しべが雄しべより短い花と、逆に雌しべの方が長い花とがあり、同じ者同士では株が違っても、蜜を吸いに来た昆虫に付着する花粉の位置関係からうまく受粉できないのです。
胞子で殖えるシダ植物では、多くの場合、胞子が発芽してできるハート型の配偶体(前葉体)は雌雄同株で、同じ配偶体に造精器と造卵器を持ち自家受精の可能性が高いのですが、ワラビ属などでは両者の成熟の時期をずらす様に工夫しています。他にも、造卵器と造精器の位置を離したり、先に大きくなった前葉体が一種のホルモンを放出して周囲の小さい前葉体を雄にしてしまうといった仕組みを持つものも有ります。また色々なシダ植物で、自家受精の回避を目的に、配偶体の雌雄の分化が進化したと言われます。
多くの無脊椎動物・魚類・両生類にも、精子と卵子が同一個体で作られる雌雄同体種が見られますが、精子と卵子の発達時期を分離していたり、生殖器官の配列や交尾方法によって、自家受精しない様な工夫がされています。ミミズでは雌雄の外部器官が、自家受精できない様な位置になっています。また軟体動物のウミウシも、雌雄同体で繁殖にパートナーは必要ない様に見えますが、繁殖期には這い跡をたどって必死に配偶相手を探し回り、相手を見つけられないと自分一人では繁殖でき無いのが普通だと言います。ただ、相手は同種でありさえすれば良く、雌雄異体の我々の様に異性である必要は無い訳です。
雌雄同体のホヤ類にも、同様に自家受精を妨げる機構を持つものが有ります。アメリカ西海岸に生息するウスイタボヤは、受精して発生した個体(大きさ約1mm)が群体を形成します。ゼラチン状の膜で覆われた群体には血管が走り、成長につれて広がって行き、隣の群体と接触すると合体して1つの大きな群れとなる事が有ります。この時には、まずゼラチン状の膜が癒合し、続いて血管が繋がります。しかし、反対に臓器移植の際に似た拒絶反応が起こる場合も多く、その時には血管が後退して両者は分離してしまいます。拒絶反応は、移植細胞表面に存在する非自己抗原に対する免疫反応で、この抗原となるのが組織適合性分子という細胞表面タンパクです。その中で最も重要なのが、高等脊椎動物の細胞で発現する、主要組織適合性遺伝子複合体(MHC)と呼ばれる遺伝子群にコードされたMHCファミリータンパクです。つまり、MHC分子によって自己と非自己とを区別している訳で、MHCが一致すれば拒絶反応は起きない訳です。ホヤもそれに似た前MHCの様なものを持ち、両親からもらった前MHC遺伝子の1つでも異なると拒絶反応が起きると考えられます。そして癒合できるのは、完全に一致する前MHCを持つ場合だけなのです。ところが、ウスイタボヤが海中に精子を放出して有性生殖を行う受精時には、癒合の時とは全く反対の事が起こります。つまり、卵子と異なる遺伝子を持つ精子でなければ受精できないのです。癒合する時には同じ遺伝子でなくては拒絶されたのに、受精では反対に自分とは異なるものでなければならない。生命は、自らの体内に侵入して来る異物に対しては、頑ななにこれを拒絶しますが、性に於いては逆に自分とは異なるものを進んで受け入れようとしているのです。
また生殖の相手として、自分とは異なるものを求めるという行動は、哺乳類でも観察されています。遺伝的にMHCのタイプのみが違う純系のマウスを使った性行動の実験によると、オスは交尾の相手として自分とは異なるMHCを持つメスを選ぶ傾向があると言います。その確率は70%にのぼり、マウスはこのMHCの違いを尿の匂いでかぎ分けているらしいのです。ヒトでも、自分と最もかけ離れたMHCの遺伝子型を持つ異性の体臭に、最も強く惹かれるという実験結果が出ていると言います。また、子供時代を兄妹の様に共に過ごした男女は、成人しても互いに性的興味を示さない事が知られています。イスラエルのキブツでは、子供は誕生とともに乳児院に預けられ、以後、小人数のグループで育てられていましたが、このグループ内での結婚は全く無く、それどころか恋愛事件さえ起こらなかったと言います。
このように、性に於いては自分とは異なる相手を求めるという現象が広く存在しているのです。しかし、これは考えて見れば当前の事で、性が遺伝的多様性を拡大する為のシステムだとするならば、遺伝的に同じもの同士が結びついてもそれは無性生殖と同じで効果が無いからです。生物は高いコストを払ってまで有性生殖をしている訳で、その効果を高めようとすれば、出来るだけ自分と遺伝的に異質なものを生殖相手として求めるのはむしろ当然と言うべきでしょう。こうして遺伝的多様性を広げる為に、「性では異質を求める」という原則が存在しているのです。
有性生殖の複雑なシステム
有性生殖は、遺伝的多様性を生み出す事を目的として生まれた訳ですが、生物はその為に極めて複雑なシステムを進化させています。性は真核生物だけではなく、原核生物に於いても見られるのですが、その様式は両者では全く異なっています。原核生物の細菌は、普段は無性生殖で増殖していますが、ごく低頻度(約百万分の1)ですが性による遺伝的組換えも起こしています。ただ真核生物の受精の様に雌雄の細胞が完全に融合し、2つのゲノムを混合して2倍体ゲノムを形成するのではなく、細菌同士の接合により、オスからメスへ一方通行にほんの数個の遺伝子が伝達されるだけです。つまり、交配は異なる細菌の株間で接合して、一方から他方に伸びる小さな中空の管の性線毛を介して、小さな遺伝因子のプラスミドを伝達する事によって行われているのです。
プラスミドと言うのは、細菌や酵母の細胞質内に存在する、核様体のDNAとは独立して自律的に複製する環状2本鎖DNAです。現在では、遺伝子組み換え操作に於ける、遺伝子の運び役のベクターとして、多くの研究に利用されています。プラスミドには、抗生物質に対する薬剤耐性遺伝子を持つものも有りますが、Fプラスミドと呼ばれる細菌の雌雄を決めるプラスミドは、稀にバクテリアの主たる環状染色体の中に挿入されてその一部となる事があります。この挿入後に接合が起こると、宿主染色体の一部も一緒にF-の受容体細胞に伝達される事になります。そして、受容体細胞の染色体と移動して来たDNAとの間で遺伝的組換えが起こるのです。これが細菌における遺伝的組換えの基本的なメカニズムです。
図25)細菌の接合とプラスミドの伝達(青いリングがプラスミド)

(出典:ウィキメディア・コモンズ )
他方、我々真核生物では、減数分裂と受精という極めて複雑なシステムを進化させる事によって、遺伝的組換えを行なっています。まず、真核生物の細胞分裂(有糸分裂)自体が、極めて複雑な過程です。原核生物では、遺伝情報は1本のリング状のDNAに収められ、1ヶ所で細胞膜に固定されているだけです。その為、細胞分裂(無糸分裂)時には、DNAが複製された後に2本のDNAが細胞膜に固定されたまま両側に移動し、その間に隔壁が形成されて分裂は完了します。ところが、真核生物では遺伝情報が複数の糸状のDNAに分散して収められ、それが核の中に閉じ込められている訳です。その為、細胞分裂時に正確にゲノムの1組を子孫の細胞に分配する必要から、真核細胞のDNAは染色体という非常にコンパクトに凝縮された構造を作り上げたのです。
まず真核生物のDNAは、DNA結合タンパクのヒストンと結び付く事でコンパクトに畳み込まれます。ヒストンは真核細胞にしかない、しかも非常に大量に存在するタンパク質で、その総質量はDNAのそれに匹敵します。このヒストンと核DNAの複合体がクロマチン(染色質)です。ヒストンには5種類あり、その内の4種類2分子ずつから8量体のヒストンコアが作られ、これにDNAの二重らせんが約 1.65回左巻きに巻き付く事で、クロマチンの基本単位であるヌクレオソームを形成します。そして、糸に通したビーズ状に並んだヌクレオソームは、もう1種のヒストンH1を介してさらに密に詰まったらせんを形成して、直径約30ナノメートル(nm:1/100万mm)の繊維(30nmクロマチン繊維)となります。このクロマチン繊維は、さらにループを作りながら足場となるタンパク質に結合し、それがさらにらせんを形成して凝集クロマチンを作ります。分裂していない間期の細胞の染色体は、クロマチンに凝縮しているとはいえ極めて細い糸状で核全体に広がっている為に、染色体全体をはっきり見る事はできません。しかし細胞が分裂を始めると、凝集クロマチンのループはさらに絡みつき圧縮されて、光学顕微鏡でもはっきりと見える様になります。これが良く知られている染色体で、この凝縮によって5cmのDNAが約5μmにまで縮む事になるのです。各染色体は1本のDNAからできており、真核細胞ではこうした染色体が複数個存在してゲノムを形成しています。この染色体の形成によって、細胞分裂時に複数の染色体を正確に2つの娘細胞に分配する事が可能となったのです。この時の染色体が糸の様に見える事から、真核細胞の分裂は有糸分裂と呼ばれています。そして、分裂が終わると染色体は再び脱凝縮してはっきり見えなくなります。これは染色体に凝縮した状態では、DNAの転写活性が失われてRNAの合成ができない、つまり遺伝情報を引き出す事ができない為です。こうして細胞分裂をしていない間期の細胞では、染色質は拡散して染色質網を形成します。クロマチンが緩んで拡散している場所では、DNAの転写が活性化しており、子猫がじゃれた後の毛糸玉の様に見えます。
図26)染色体とクロマチン

(出典:ウィキメディア・コモンズ より改変)
有性生殖と2倍体の進化
真核生物の中には、わずかですが決して有性生殖を行わず、有糸分裂だけで増殖するものも存在します。原生生物のアメーバ類やトリパノゾーマ、単細胞性の紅藻類や菌類などで、こうした生物は染色体を1組しか持っていません(1倍体:haploido)。しかし、ほとんどの真核生物は一生のどこかで有性生殖を行い、染色体が2組ある2倍体(diploido)の細胞が出現します。ゾウリムシの接合などの特殊な例もありますが、普通は真核生物の有性生殖では雌雄2つの性が分化し、両性の個体から生じた1倍体の配偶子(卵と精子)が合体、受精する事で行われます。この配偶子の合体によって、染色体を2組持つ2倍体が生まれる訳です。真核生物に特徴的な2倍体の存在は、その独特な有性生殖の結果なのです。こうして出来た2倍体細胞が、再び有性生殖の為の配偶子を作るには、染色体を1倍体に戻さなければなりません。この為に、真核生物が発明した特殊な分裂方法が減数分裂です。こうして有性生殖をする真核生物では、受精と減数分裂のサイクルの間に、1倍体の世代と2倍体の世代が交代で現れる事になったのです。そして、2個の1倍体細胞が融合して2倍体細胞を形成する事で、両者のゲノムは完全に混合される訳です。実は、この2倍体細胞が減数分裂して再び新しい1倍体細胞が生じる過程で、遺伝子の組換えが起こり、新しい組み合わせのゲノムの1組ずつが1倍体細胞に分配される仕組みになっています。そして、この組み合わせを変えられた1倍体細胞同士が融合して、次の新しい2倍体世代が誕生する訳です。こうして1倍体世代・受精・2倍体世代・減数分裂というサイクルにより、次々と新しい遺伝子の組み合わせを持つ子孫が生み出されて行く事になるのです。
分裂酵母の様な原始的な真核生物の中には、1倍体世代で有糸分裂をして増殖し、接合で2倍体細胞(接合子)になると直に減数分裂をして1倍体に戻ってしまうものも存在します。しかし、有性生殖をする真核生物のほとんどは、2倍体世代で有糸分裂を行い増殖しています。植物の中には、1倍体世代でも2倍体世代でも細胞分裂するものも有りますが、最も原始的なものを除くと植物でも1倍体世代はごく短く単純であり、反対に2倍体世代の方は発生や増殖を行い期間も長くなっています。脊椎動物を含むほぼ総ての多細胞動物は、実質的にその生活環の全体を2倍体で過ごしています。そして、1倍体細胞は全く分裂せず短期間のみ存在して、生殖時の細胞融合の為だけに特殊化しています。この有性生殖の為だけに特殊化した1倍体細胞を、配偶子と呼んでいる訳です。通常は、形態の異なる2種類の配偶子、即ち大きくて運動性のない卵子と、小さくて運動性のある精子が形成されます。そして、この配偶子の融合によってできた2倍体細胞が、分裂・増殖して分化し、複雑な多細胞生物を作り上げているのです。
減数分裂での遺伝的組換え
減数分裂では、どの様にして遺伝的組換えが行われているのでしょうか。通常の体細胞の有糸分裂では、まずDNAの複製が行われ、その後、倍化した染色体が細胞分裂によって2個の娘細胞分けられます。その結果、分裂の前後で染色体数に変化は有りません。しかし、有性生殖では2つの1倍体の配偶子が融合する事により、染色体数は倍加して2倍体細胞が出来上がる訳で、2倍体細胞から再び配偶子を作るには、染色体数を半分にしなければなりません。この生殖細胞が作られる時に起こる、染色体数を正確に半分に減少させる特殊な細胞分裂が減数分裂です。1倍体の配偶子2個が融合する事で生まれる2倍体細胞には、父方の染色体と母方の染色体が各1組ずつ入っています。そのため、性を決定する性染色体を別にすると、2倍体の核にはオスとメスに由来する非常に良く似た1対の染色体(相同染色体)が存在する事になります。そして減数分裂では、各相同染色体の片方のみを受け継ぐ事によって、染色体数が半分の1倍体の配偶子が生まれるのです。つまり配偶子は、各遺伝子について父方か母方どちらか一方のコピーのみを持っている訳です。
この減数分裂の機構として考えられる最も簡単な方法は、染色体の複製をせずに相同染色体同士が対合して、そのまま紡錘体上で分離する事でしょう。そうすれば、減数分裂は染色体の複製(S期)を欠いた有糸分裂の変形と見なす事ができます。ところが、実際の減数分裂はもっと複雑な機構を採用しているのです。減数分裂でもまずDNAが複製され、それに引き続いて2回の細胞分裂が立て続けに起こり、これによって染色体数が半減する仕組みになっています。つまり減数分裂では、1個の2倍体細胞から4個の1倍体細胞が生じるのです。通常の有糸分裂では、1個の2倍体細胞が分裂して2個の2倍体細胞ができるに過ぎません。減数分裂の1回目の分裂に於いては、DNAの複製により倍化した染色体(姉妹染色分体)が分離せず、そのまま相同染色体同士の対合が起こり、4本の姉妹染色分体が結合した2価染色体と呼ばれる構造が出来上がります。この時、母方の染色体の一部と、これと相同な父方の染色体の一部が交差して入れ替り、遺伝的組換えが起こるのです。そして紡錘体上に並んだ2価染色体は、2本の姉妹染色分体が接着したまま相同染色体が分離し、反対の極へ移動して2つの娘細胞に分裂する。こうして減数分裂の第1分裂の結果、DNA量は2倍体と同じだが、相同染色体の片方のみを持つ娘細胞が2つ出来上がる事になります。この第1分裂に続いて、染色体の複製のないまま第2分裂が起こります。染色体は再び紡錘体上に並び、今度は通常の有糸分裂の場合と同様に姉妹染色分体が分離して、4個の1倍体細胞(配偶子)が形成されるのです。
図27)減数分裂(二価染色体で染色体交差が発生)
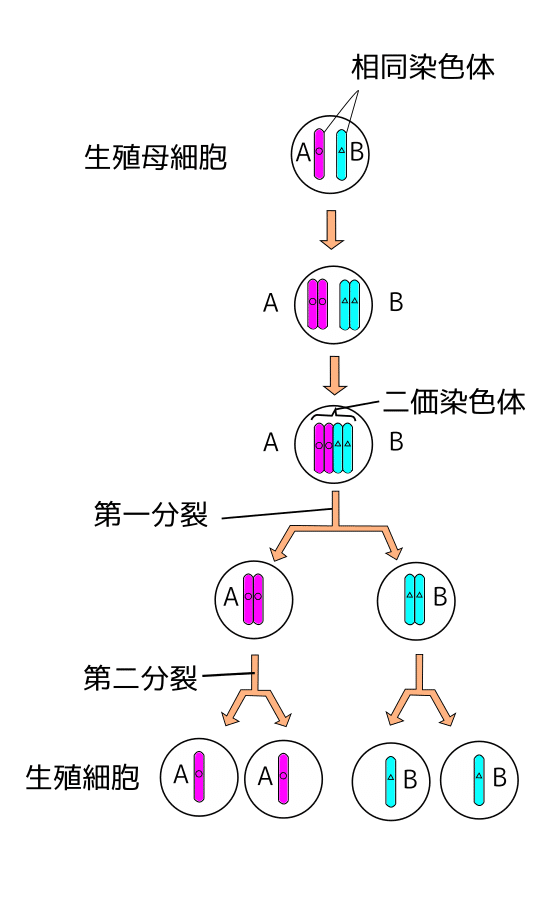
図28)染色体交差(乗換え)

(出典:ウィキメディア・コモンズ )
ここで注意しなければならないのは、第1分裂で起こる染色体交差による遺伝的組換え(乗換え)は、偶然に起こるものでも、他の機構の副産物として生じるものでも決してないという事です。交差の分子的機構が不明だった時代には、細胞学のイメージから、減数分裂で対合して捻じれた染色体に物理的な張力が働いて切断し、生じた切断端が交差して再結合して、2本の組換え染色分体ができると考えられていました。しかし、その分子的機構が明らかになって来ると、この遺伝的組換えは、はるかに複雑で組織化された過程である事が分かって来たのです。遺伝的組換えは、少なくとも50の遺伝子とその作り出す酵素が必要とされる、極めて複雑でシステム化された過程だったのです。しかも、染色体の交差が起こる第1分裂の前期(染色体の凝集から2価染色体が紡錘体の中央部に並ぶまで)は、減数分裂の全期間の90%以上という異常に長い時間を占めているのです。これらの事は、減数分裂という極めて複雑なシステムが、遺伝的組換えを目的として生み出されたものである事を強く示唆しています。
第1分裂前期に入ると染色体の凝集が始まりますが、減数分裂では凝集はもがきながらゆっくりと進行します。凝集の初期には細長い糸の様に見える染色体が、まるで何かを探し求めるかの様にうごめき、細胞核はゆっくりとうねりもまれ始めます。これは各染色体が、その対合相手の相同染色体を見つけようと蠢動している状態です。そして、2つの相同染色体がうまくぶつかると、染色体の相同領域が並んで結合し、両者はファスナーを閉める様に融合して行きます。このファスナー様の対合構造はシナプトネマ構造(対合複合体)と呼ばれ、長い梯子状のタンパク質コアの両側に2本の相同染色体が並び、長い直線状の染色体対を形成したもので、電子顕微鏡では文字どおりファスナーの様に見えます。染色体の交差の為には、2つの相同染色体が密着して並んでいる事が必要で、遺伝子の組換えはこの対合複合体の内部で起こります。シナプトネマ構造の所々には、組換え小節という直径約90nmのタンパク質を含む大きな会合体があり、これが100nm幅のファスナー部を横断して、父方と母方の染色体上のDNAを局所的に引き合わせ、組換えを行うと考えられています。組換えが終わると事態の進行は少し速まり、染色体の短縮化と凝集はまだ続くが、ある時点でシナプトネマ構造は壊れ相同染色体は分離し始めます。この時、2本の相同染色体は幾つかの個所で固く結合し、この結合点のキアズマで染色体の交差が起きている訳です。2価染色体は少なくとも1個、中には2個以上のキアズマを持つものも多く、相同染色体間には複数の交差が生じる事が可能です。ヒトでは、この染色体の交差が各対ごとに平均2~3ヶ所で起きており、これにより各配偶子に、遺伝的組成の組換えられた染色体が分配される事になるのです。
このように真核生物は遺伝子の組換え、つまり遺伝子の多様性を増大させる為に、有性生殖という極めて複雑で精緻なシステムを進化させ、また多くの時間とコストを掛けて実行しているのです。この事実は、遺伝的多様性を増大させるという事が、生物にとって如何に重要で必要不可欠なものであるかを如実に物語っています。減数分裂における、遺伝子の組換えによる多様性の増大は2段階に分けて起こっています。まず第1分裂の際に、父方と母方の染色体がランダムに組み合わされて、娘細胞に分配される事による遺伝子の混合です。これだけでも、2n個の染色体を持つ1つの個体から、遺伝的に異なる配偶子が2のn乗種類も生じるはずです。例えば、ヒトではこの数は2の23乗≒8.4×10の6乗種類にもなります。さらにその上、染色体の交差によって各相同染色体単位で遺伝子の組換えが起きる結果、配偶子の遺伝的多様性は無限と言っていいほどになってしまいます。この無限に近い多様性を持つ配偶子同士が結合する事により、親とは異なる新しい遺伝子組成を持つ子が生まれる。こうして生物の遺伝的多様性は、飛躍的に拡大する事になるのです。
(つづく)
① 遺伝子とは一体何者か?
② 個体発生は系統発生を繰り返す
③ 生存闘争は進化を推進するか?
⑤ 激変する地球環境の下で生命は誕生し、進化してきた
⑥ カンブリア爆発:捕食者の出現
⑦ 気候変動と大量絶滅

この記事が気に入ったらサポートをしてみませんか?