
カンブリア爆発:捕食者の出現(遺伝子決定論は正しいか? ⑥)

カンブリア爆発
昔から、古生代カンブリア紀(5.41億~4.85億年前)に入ると、 突如として化石記録が急増する事が知られていました。そして、それ以前のほとんど化石が発見されず、生命の痕跡の乏しい地質時代を、先カンブリア時代と一括して呼んでいたのです。ところが、5.41億年前にカンブリア紀が始まると様相は一変します。硬い殻を持つ多種多様な海洋生物が突如出現し、今日地球上に見られる動物の「門」、即ち生物の体の基本構造プランのほとんど総てが、このカンブリア紀のわずか 2000~3000万年間に出揃ってしまったのです。この中には、我々脊椎動物の祖先も含まれています。直前の、エディアカラ紀に見つかる多細胞生物化石が 50種程度であるのに対し、カンブリア紀にはそれらとは全く類縁関係の無い1万種近い多細胞生物が出現し、生態系の多様性が一挙に拡大しているのです。現在、世界中のこの時代の地層からは大量の化石が発見されています。しかも、現在では全くそれに類縁のものが存在しない、完全に絶滅した生物もこの時期には多数出現しています。このカンブリア紀の初めに起こった、空前絶後の進化の爆発現象をカンブリア爆発と呼んでいるのです。
図48)カンブリア爆発(バージェス頁岩動物群)

(出典:ウィキメディア・コモンズ )
このカンブリア爆発の当時の状況を最も良く保存しているのが、スティーブン・J・グールドの『ワンダフル・ライフ』で有名になった、バージェス頁岩動物群です。現生の多細胞生物のグループが化石記録に出現して来るのは、5.41億年前のカンブリア紀の事ですが、その出現の仕方は徐々に数を増して行ったものでは無く、ごく短期間に多様性が爆発的に拡大し、現生の主要動物グループの事実上の総てが、地質学的にはあっという間に出現していたのです。これほど多様な生物の分類群が一時に出現するには、それ以前に長い初期進化の時期が必要と感じられますが、不思議な事に先カンブリア時代の地層からは、これらの動物との類縁関係を示す様な化石は、全くと言っていいほど見つかっていないのです。
図49)バージェス頁岩(カナダ、ブリティッシュコロンビア州)

(出典:ウィキメディア・コモンズ )
しかも注目すべきは、単に短期間で集中的に起こったという事だけではなく、出現した生物の驚くべき多様性です。このカンブリア爆発が終了した直後の、地球の海に棲息していた多様な動物達を今に伝えているのが、カナディアン・ロッキー山中で発見されたバージェス頁岩層(5.3億年前、カンブリア紀中期)の動物群なのです。そこには海綿動物から脊索動物まで、極めて多様な生物が含まれています。グールドは、このブリティッシュ・コロンビア州のたった一ヶ所の化石発掘場から出土した生物の多様性は、現在の海洋に棲息する無脊椎動物すべてを合わせた解剖学的デザインの幅をも凌駕していると主張していました。
実際、バージェス頁岩動物群には、現生種との類縁関係もはっきりしない、奇想天外な形態を持つ奇天烈動物たちが数多く含まれていました。例えば、体長5~6cmのオパビニアは、多数の体節に分かれた胴体の両側に葉状の遊泳肢を持ち、これで海底近くを泳いでいた様です。頭にはドアのノブ状の5つの飛び出した目、頭の先からは胴体の長さの半分近くもある象の鼻の様な器官が伸び、その先端に付いたハサミで食物をつかみ口に運んでいたと考えられています。
図50)オパビニア
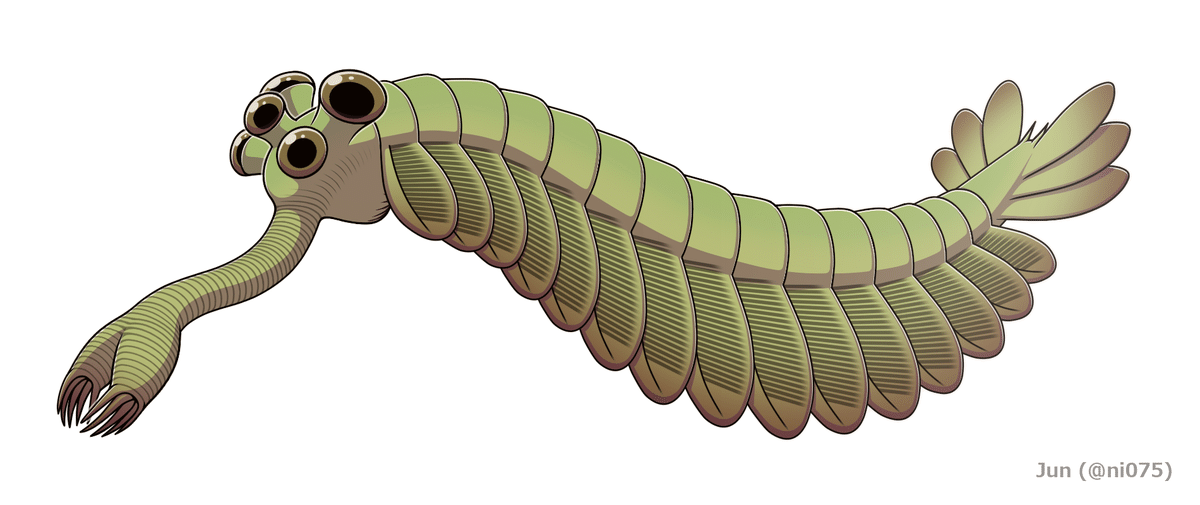
(出典:ウィキメディア・コモンズ )
また、アノマロカリス(「奇妙なエビ」の意)も不思議な動物です。体長は7~50cm、大きな頭部と尻すぼみの胴体から成る水滴を縦割りにした様な形で、胴体の両側にずらりと並んだ三角形のヒレを波打たせて遊泳していました。頭部の下側には、パイナップルを輪切りにした様な丸い口を持ち、頭部先端の2本のエビの尻尾の様な付属肢を使って、獲物を捕らえて食べていたと考えられています。
図51)アノマロカリス類(ラディオドント類、右上:アノマロカリス)

(出典:ウィキメディア・コモンズ )
「幻覚が生んだ動物」と言う意味のハルキゲニアは、体長1~5cmの底生生物で、背中に7対の細長い針の様なトゲを持ち、棒状の胴体から伸びる7~8対の細長い脚で海底を歩いていました。
図52)ハルキゲニア

(出典:ウィキメディア・コモンズ )
オドントグリフス(「歯の生えた謎」の意)も奇妙な生物です。非常に稀な種で体長 3mm~12cm、体全体が薄っぺらい草履の様な形で、発見当初は体全体を波打たせて泳いでいたと考えられていました。下側に有る小さな丸い口の周りには歯の様なものが生えていますが、これは軟体動物が持つ歯舌の初期のものと見られています。また、化石の多くがシアノバクテリアのマット上で見つかる事から、オドントグリフスは微生物マット上を這いながら歯舌でこそぎ落として食べていたと考えられています。
図53)オドントグリフス

(出典:ウィキメディア・コモンズ )
「泳ぐエビ」を意味するネクトカリスは、体長3~10cmで当初はエビの様な形の頭と魚の様な胴体を持つ、節足動物の頭と脊索動物の胴体が合体した様な不思議な動物と見られていました。エビの様に飛び出した目の後ろの、甲羅と見られていた楕円形の構造は、現在では頭足類がジェット推進に使用する漏斗とされ、原始的な頭足類と考えられています。
図54)ネクトカリス

(出典:ウィキメディア・コモンズ )
マルレラは、バージェス頁岩から最も多く産出する化石で、中には脱皮する最中の化石も発見されています。体長は2.4mm~24.5mmで、オール状の付属肢を使って海中を遊泳して小動物や懸濁物を食べていたと考えられています。マルレラは、発見当初は甲殻類と考えられたりしましたが後に否定され、現在も他の節足動物との類縁関係が不明の動物です。
図55)マルレラ

(出典:ウィキメディア・コモンズ )
ウィワクシアは、多数の鱗状の骨片で覆われた背面に、前後に走る2列の10本前後の鋭い棘を持つ、体長約2.5~5cmのお椀を伏せた様な形の動物です。鱗と棘で身を守りながら、海底を這い回って餌を取っていたと思われます。ウィワクシアも分類について長らく議論が有った動物で、顎らしい構造の歯舌との類似性から、現在では軟体動物と見なされている様です。
図56)ウィワクシア

(出典:ウィキメディア・コモンズ )
図57)バージェス動物群(1:ヴァヌシア (海綿)、2:チョイア (海綿)、3:ピラニア (海綿)、4:ニスシア (腕足動物)、5:バージェスソキータ (環形動物多毛類)、6:オットイア (鰓曳動物)、7:ルイゼラ (鰓曳動物)、8:オレノイデス (節足動物三葉虫類)、9:シドネイア (節足動物)、10:レアンコイリア (節足動物メガケイラ類)、11:マルレラ (節足動物マーレロモルフ類)、12:カナダスピス (節足動物 Hymenocarina類)、13:モラリア (節足動物)、14:バージェシア (節足動物)、15:ヨホイア (節足動物メガケイラ類)、16:ワプティア (節足動物 Hymenocarina類)、17:アイシェアイア (葉足動物)、18:スケネラ、19:エクマトクリヌス、20:ピカイア(脊索動物)、21:ハプロフレンティス(ヒオリテス類)、22:オパビニア (節足動物オパビニア類)、23:ディノミスクス、24:ウィワクシア、25:ペイトイア (旧復元、節足動物ラディオドンタ類))
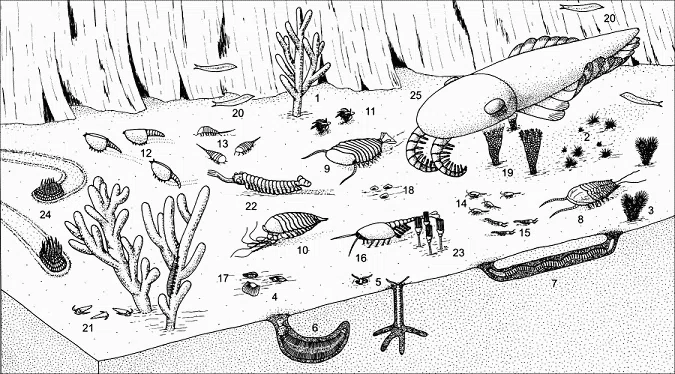
(出典:ウィキメディア・コモンズ )
バージェス頁岩からは、今日の節足動物門の主要な4グループを代表する、初期の種も見つかっています。今日約110万種あまり知られている節足動物の総ては、鋏角類(クモ・サソリ・カブトガニなど)・多足類(ムカデ・ヤスデなど)・甲殻類(カニ・エビ・フジツボなど)・六脚類(昆虫・トビムシなど)の主要4グループに収まっています。ところがバージェス頁岩からは、このいずれにも入らない節足動物が 20~30種類も見つかっているのです。バージェスの時代にはユニークな解剖学的デザインが極めて多数存在し、また現在の地球でお馴染みのグループも、今の枠組みを越える多様なデザインを実験中だったのです。今日では、極端な意見として批判的に見られる事が多いのですが、J・グールドは『ワンダフル・ライフ』の中で、次のように述べていました。
「解剖学上の多様さの幅が最大に達したのは、多細胞生物が最初の多様化を終えた直後の事だったのである。………なるほど現在の地球は、かってなかったほど多数の種を擁しているかもしれない。しかしその大半は、解剖学的に見ると少数の基本的デザインの繰返しである。分類学者は50万種以上の甲虫を記載しているが、そのほとんどすべてはただ1つの基本設計図に最小限の変更を加えたコピーにすぎない。………バージェスの海にくらべると、今日の海洋に生息する動物は、種数こそ多いものの、それらの土台となっている解剖学的な設計プランの種類ははるかに少ないのだ」(『ワンダフル・ライフ』スティーブン・J・グールド著)
カンブリア爆発で一気に出現し、カンブリア紀だけで絶滅し消えて行った奇妙な動物たちと同時に、カンブリア紀には現在の動物界につながるほとんど総ての門、つまり生物の基本型も出現しています。これらの生物の子孫たちは、その後の進化により外見上の形は変わって行きますが、基になる基本構造は驚くほど変わっていません。そういう意味では、このカンブリア紀以後の進化は、マイナーチェンジでしかなかったとも言えるのです。
酸素濃度の急増と多細胞生物の適応放散
古生代カンブリア紀に入り、突然の様に大量の多細胞動物の化石が出現する事になったカンブリア爆発ですが、この急激な多細胞動物の多様性の拡大は、クライオジェニアン時代(8.5~6.35億年前)の2度目のスノーボールアース(全球凍結)が終わった直後の、6.35億年前頃の大気中の酸素濃度が急上昇が原因であった可能性が高いと思われます。図58)「過去10億年の大気中の酸素濃度の変化」を見ると、エディアカラ紀(6.35億~5.41億年前)に、それまでは3%程度で安定していた大気中の酸素濃度が、一挙に12~13%まで急上昇していた事がわかります。現在の酸素濃度が 21%ですので、カンブリア紀の始めには今日の約 60%程度にまで急激に高まっていたのです。
図58)過去10億年の大気中の酸素濃度の変化(赤破線:現在値、横軸:100万年前)

(出典:ウィキメディア・コモンズ )
今日の世界各地の酸素欠乏海域を調査すると、酸素濃度が表面海水の0.5%未満と極めて低い海底領域でも、微小な蠕虫が生息していると言います。しかし、こうした酸素欠乏環境では食物網が単純で、動物は微生物を直接食べているだけです。ところが、酸素濃度が表面海水の3~10%程度にまで高まると、他の動物を食べる捕食者が出現し始めると言うのです。つまり、海中の酸素濃度のわずかな上昇が、捕食者の出現を後押しして、生態系の多様性を急激に拡大させる可能性が有るのです。
また、エディアカラ生物のように海底でじっとしているのに比べ、バージェス動物群の様に獲物を求めて動き回るには、多量のエネルギーが必要となるのは当然です。大量の有機物の供給と同時に、そこから効率的にエネルギーを生産するのに不可欠な酸素濃度の上昇が、捕食者の進化を促し、カンブリア紀での生態系の多様性の爆発的拡大の鍵になっていたのです。エディアカラ紀に入り突然出現した奇妙な多細胞生物のエディアカラ生物群と、引き続き起こったカンブリア爆発での多細胞動物の急激な進化は、エディアカラ紀の酸素濃度の急上昇が原因だった可能性が高いのです。
その後の地球生態系の景観を一変させてしまう事になる、原生代と顕生代の境目で起こった多細胞生物の急激な進化は、スノーボールアース直後の酸素濃度の急上昇と言う、地球環境の急変が引き起こしていた訳です。進化論者の言う「生存闘争」や「自然淘汰」は、ここでは何の役割も果たしていない事に注意して下さい。46億年前の誕生以来、繰り返されてきた地球環境の大変動こそが進化の原動力であって、それが生物の急激で不連続な進化を生み出してきたのです。
捕食動物の出現と多様性に富む生態系の誕生
原生代末のエディアカラ生物群と、バージェス動物群に代表されるカンブリア紀の動物では、その種類の多さだけではなく行動や形態にも大きな違いが存在します。前回にも触れた様に、エディアカラでは、はっきりした捕食動物の化石が発見されていません。また、エディアカラ生物のほとんどが、柔らかい組織の軟体性の生物だったにも拘わらず、生存中に傷つけられた様な化石も存在しません。これらの事から、原生代末のエディアカラの時代には、まだ多細胞生物を狩る肉食の捕食動物が出現していなかった可能性が高いのです。
また、エディアカラ生物群が繁栄していた当時、海底はシアノバクテリアの微生物マットで広く覆われていたと言われます。というのも、これを食べる動物が出現して、微生物マットが破壊された形跡が無いのです。エディアカラ生物群の様な軟体性の生物の印象化石が残ったのも、微妙な堅さを持つ微生物マットで海底が覆われていた為と考えられるのです。
海底に這い跡を残す多細胞動物が現れたのは、原生代後期(新原生代)の約10億年前頃です。エディアカラでも多くの生痕化石が残されていますが、そのほとんどは堆積物の表面を這い回った動物のもので、現在の様な堆積物中を深く掘り進むタイプの動物は、先カンブリア時代にはほとんど知られていません。ところが顕生代のカンブリア紀に入ると、海底面から10cm以上深く掘り下げられた生痕化石が、世界中から頻出する様になるのです。恐らく、ミミズの様に泥の中を掘り進みながら海底の堆積物を食べて有機物を得る動物が、古生代に入って急激に現われたものと思われます。この様な生痕化石の出現をもって、顕生代の始まりとされる事も多いのです。しかも、こうした生痕化石は、古生代に入った直後の同時期に世界的規模で、掘り込む深さを一気に海底面下10cm以上にまで増大させていると言います。これは大型の肉食動物の出現で、その攻撃を逃れる為に泥の中深く潜りこむ必要が出てきたとも考えられます。
エディアカラの砂岩層に軟体部をもつ印象化石が保存されたのは、1つには当時の酸素濃度がまだ低く死体の分解がゆっくり進んだ事と、海底の砂地に潜って生活する動物が存在せず、化石を含む地層が乱される事が少なかった為と思われます。またカンブリア紀には、三葉虫や貝の様に硬い殻で身を包んだ動物が多数出現しますが、これも捕食者から身を守る為の進化と考えられます。実際、エディアカラ紀とカンブリア紀の境界付近から産出する、1cmにも満たない小型の有殻生物化石の硬い殻の表面には、しばしば他の生物によってあけられたと思われる孔が認められると言います。また、カンブリア紀の三葉虫化石の中には、捕食動物によって食いちぎられた様な傷跡を持ち、しかも傷の周囲が盛り上がった治癒の跡が見られるものも発見されているのです。
こうして原生代の終わりとともに、微生物マットの上に静かに体を横たえていたエディアカラ生物の時代は終わり、顕生代に入ると酸素濃度の急上昇を背景に、硬い殻を持った肉食の捕食者が進化してくる事になるのです。カンブリア紀には、アノマロカリスなど体長50cmもある巨大な捕食動物が、餌を求めて海中を自由に泳ぎ回る様になります。そして、この肉食の捕食動物の進化によって、今日に通じる複雑な生態系が完成する事になるのです。実際、バージェス頁岩の奇妙な動物達は、豊富な生態的地位を棲み分け、複雑な群集構造を形成していた事が分かっています。
バージェス動物群は、その生息場所から海底と海中に棲む、2つの大きなグループに分けられます。圧倒的多数を占めているのが底生種と海底近くで生活する種で、彼等は水深 90m未満の浅海底で生活していたと考えられています。節足動物のマルレラの様に砂や泥の上を歩き回ったり、鰓曳虫類のオットイアの様に海底に穴を掘って隠れ棲むものもいました。もう1つのグループは、分類不明のアミスクウィアや脊索動物のピカイアの様に、淀んだ海底から離れて海中を遊泳しながら生活していた少数の生物です。
図59)オットイア

(出典:ウィキメディア・コモンズ )
図60)アミスクウィア

(出典:ウィキメディア・コモンズ )
図61)ピカイア

(出典:ウィキメディア・コモンズ )
また、バージェス動物群を摂食様式で分けると、次の4つのグループに分類する事ができます。
① 堆積物を摘まみ取って食べる動物
(大半が節足動物、全個体数の60%、全属の25~30%)
② 堆積物を濾過して食べる動物
(大半は硬組織を持つ軟体動物、全個体数の1%、全属の5%)
③ 懸濁物食者
(大半が海綿類で水層から食物を直接取込む、全個体数の30%、全属の45%)
④ 肉食者と死肉食者
(大半が節足動物、全個体数の10%、全属の20%)
このように、バージェス動物は様々な生態的地位を巧みに棲み分け、その後の地質時代の世界とも良く似た、複雑な生態系を形成していたのです。
多細胞生物は、約10億年前の海の中で登場しますが、植物と動物では異なる道を歩む事になります。植物の場合、多細胞化は海の中ではあまり進まず、本格的な細胞分化を伴った多細胞植物が進化して来るのは、4.5億年前に陸上に進出した後の事です。 一方、動物は海中で多細胞化を急速に押し進めて行きます。まず、海底の微生物マットや堆積した有機物を摘み取り、あるいは濾過摂食する多細胞動物が最初に誕生します。彼等は原生代後期に出現し、海底に様々な這い跡の生痕化石を残す事になります。また原生代末には、不思議なシート状のエディアカラ生物群も出現します。そして5.41億年前の顕生代に入ると、今度はこれらの多細胞動物を捕食する肉食の大型動物が進化して来る事になるのです。つまり、食物連鎖の頂点に立つ捕食者は古生代に入って初めて登場するのであり、これによって海の中に光合成で生み出された有機物を基礎とする食物連鎖のピラミッドが完成し、複雑で多様性に富む生態系が誕生する事になるのです。こうして、古生代には今日的な生態系が形成されて生物の多様性が一気に拡大し、肉眼で見える大型の多細胞動物が大量に出現する事になったのです。これが、多細胞動物が一斉に適応放散したカンブリア爆発だったのです。
カンブリア爆発では、1万種にも及ぶ大量の種が一度に出現し、生物界の多様性が一挙に拡大した訳ですが、この原因の1つは捕食者の出現にあったと考えられます。捕食者の存在が、特定の種による生態系の独占を制限し、他の種のために空間を開放する事により、種の多様性を増大させる事が知られています。捕食動物の出現によって、様々な種が相互作用し合う複雑な生態系が作り出され、多様な多細胞生物が共存出来る様になったのです。
ところで、有能な捕食者となるには2つの条件を満たす必要があります。1つは獲物の位置を正確に捉える為の優れた感覚器と、そこからの情報を素早く処理する脳、つまり精巧な神経系の発達です。もう1つは、獲物を捕える為の俊敏な運動能力で、この為には高性能な筋肉組織とそれを支える呼吸・循環系の充実が不可欠となります。さらに硬い殻を持つ獲物に対しては、強力なアゴや殻を溶かす化学物質の分泌器官などが必要です。そして、これらの機能には大量のエネルギー生産が必要な訳で、それを可能にする酸素呼吸能力に大きく依存しています。古生代に入ると同時に、複雑な機能を持つ大型動物が急激に出現した背景には、原生代末期に於ける海水中の溶存酸素量の急増が有った事は明らかなのです。
エディアカラ紀からカンブリア紀にかけて、多彩な多細胞動物の急激な進化を可能にしたのが、エディアカラ紀の酸素濃度の急増にあった事は間違い有りませんが、もう一つ重要な点が有ります。 それは当時、多細胞生物が進化を始めたばかりで、生態系が誰も居ない全くのがら空き状態だった事です。進化し始めた多細胞生物は、この競争相手の全く居ない空き地だらけの生態系に、一気に適応放散して急激に多様性を拡大して行ったのです。つまり、進化論者の大好きな生存競争が存在しなかったからこそ、多細胞生物は制限される事なく伸び伸びと新たな生態的地位に進出し、急速に適応放散して進化する事が可能だったのです。カンブリア爆発は、生態系をゼロから作り出す、1回限りの多細胞生物の最初の適応放散として起こっていたのです。以後は、繰り返す大量絶滅後の崩壊した生態系への適応放散として、多細胞生物は進化して行く事になります。
(つづく)
① 遺伝子とは何者か?
② 個体発生は系統発生を繰り返す
③ 生存闘争は進化を推進するか?
④ 生命は多様性を目指す
⑤ 激変する地球環境の下で生命は誕生し、進化してきた
⑦ 気候変動と大量絶滅

この記事が気に入ったらサポートをしてみませんか?