
個体発生は系統発生を繰り返す(遺伝子決定論は正しいか? ②)

個体発生は系統発生を繰り返す
皆さんは「個体発生は系統発生を繰り返す」という言葉を聞いた事が有るでしょうか。これはドイツにおいてダーウィン進化論の普及に貢献した、動物学者・哲学者のエルンスト・ヘッケルが述べたもので、「ヘッケルの反復説」「生物発生原則」とも呼ばれています。 ここで個体発生とは動物の発生過程、系統発生は進化の過程を表しています。つまり、この説は生物は系統発生、即ち進化の跡を短縮して繰り返す事によって、個体発生すると言うものです。実際、様々な脊椎動物の初期胚がよく似ており、鰓裂が有るなど聞かれた事も有るでしょう。
しかし、ヘッケルが優生学者として社会ダーウィニズムや生物学的人種差別主義を推進し、後にその主張がナチスのプロパガンダに利用された事などから、彼の反復説は胡散臭い学説として無視され、あるいはタブー視されてきました。しかし、この簡潔で魅惑的な命題の中には、明らかに真理の一片が含まれています。次に、発生過程と遺伝子の関わりについて見て行く事にしましょう。ここでは、幾つもの遺伝子が協調して働くネットワークが、決定的に重要な働きをしているのです。
図10)ヘッケルの胚の図(左から魚・サンショウウオ・亀・鶏・豚・牛・ウサギ・ヒト)

(出典:ウィキメディア・コモンズ)
発生と遺伝子発現
受精卵からどのようにして多細胞生物の体の構造が形成されて行くのか、そのメカニズムが遺伝子レベルで最もよく研究されているのがショウジョウバエです。昆虫など節足動物の体の構造の基本単位となっているのが体節で、成虫は、頭部・3つの異なる胸部体節・8つの腹部体節から構成されています。そして頭部体節からは触覚・眼が、胸部体節からは羽が形成されます。ショウジョウバエでは、この体節の形成から発生過程が始まります。
体節は、受精後約 24時間で見える様になってきますが、体節の形成にはまず前後軸と背腹軸の体軸の決定が必要です。この体軸を決定するのがモルフォゲンと呼ばれる誘導因子で、ある細胞から周囲の細胞に拡散して濃度勾配を形成し、体軸に沿った位置情報を各細胞の遺伝子に与えます。 この誘導因子が細胞表面の受容体に結合すると、シグナル伝達経路を介して1個~複数個の転写因子が活性化され、それが核に移行して遺伝子の発現スイッチ次々とオンにして行く訳です。高濃度の誘導因子に晒された細胞では、シグナル伝達カスケードを介して、特定の遺伝子セットが活性化もしくは不活性化されます。こうして、濃度勾配を持つモルフォゲンの濃度の違いに応じて、体軸上の異なる位置の細胞で、異なった遺伝子セットが発現する事になるのです。
受精前の卵母細胞では、母親の卵巣内で発現した母性効果遺伝子の ”bicoid、”nanos の mRNAが、細胞質間橋を通して卵の前部へ移動し、”bicoid mRNAは前端に留まる一方、”nanos mRNAは細胞骨格により前方から後方へ輸送され、”bicoid mRNAは前端に、”nanos mRNAは後端に集中する事になります(遺伝子の表記はイタリック体ですが表示不能のため(”)を付けます)。そして、受精するとそれぞれ翻訳されて、Bicoid タンパク質は前端から後方に向かって、逆に Nanosタンパク質は後端から前方に向かって拡散し、それぞれ濃度勾配を形成します。
受精卵は急速な細胞分裂を開始しますが、ショウジョウバエでは最初の12サイクルは細胞質分裂を伴わない核分裂のみで、多細胞の胚の代わりに多核の胚が形成されます。この結果、モルフォゲンは容易に胚内に拡散して行きます。また背腹系では、 卵母細胞を囲む濾胞細胞から腹側のシグナル分子が供給され、それが卵の外側の膜貫通型受容体に結合して、卵内に遺伝子調節タンパクのDorsal の背腹軸に沿った核内濃度の勾配が形成されます。
こうして胚の前後軸が決定されると、次は分節(体節形成)遺伝子によって体節の境界と極性が決められて行きます。この分節遺伝子が発現するのは、受精後約3時間で胚に約6000個の核が存在する時期です。次の3群の分節遺伝子が順番に発現して行き、 分節化のパターンを大まかなものから、段階的に詳細なものに作り上げて行きます。
① ギャップ遺伝子: 前後軸に沿って幾つかの広い領域に分ける。
② ペア・ルール遺伝子:胚を 2つずつペアの 7つの領域に分ける。
③ セグメント・ポラリティ遺伝子:体節の境界と前後方向の構造を決定し、14体節に分ける。
前後軸に沿って形成された Bicoid と Nanos タンパク質の濃度勾配は、次のギャップ遺伝子の ”hunchback の発現を調節して、Hunchback タンパク質の濃度勾配を形成します。”hunchback mRNAは、初めは胚内に均一に分布していますが、 前方では転写因子の Bicoid が ”hunchback 遺伝子の転写を促進し、後方では Nanos がその翻訳を阻害するため、前方から後方へ向けて Hunchback タンパク質の濃度勾配が形成されるのです。すると今度は、この濃度勾配の作用で ”krüppel 遺伝子が中央部で発現します。このようにして、9個のギャップ遺伝子によって、胚が大まかな領域に分けられて行くのです。このギャップ遺伝子に変異が生じると、一箇所あるいは数箇所で体節が幾つか連続して欠損するこ事なります。
図11)初期胚の前後軸に沿ったタンパク質の濃度勾配
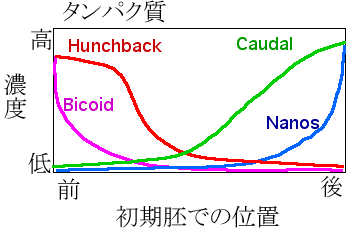
図12)ギャップ遺伝子の発現

(出典:ウィキメディア・コモンズ)
次は、8個のペア・ルール遺伝子がさらに詳細な空間パターンを決めて行きます。 この遺伝子に変異が生じると、体節が一つおきに欠失して、通常の半分の体節の胚になってしまいます。最後に、9個のセグメント・ポラリティー遺伝子 が働いて、14の体節に分割されます。この遺伝子が変異すると、体節の数は正常のままで各体節の一部が欠失し、代わりに残りの部分が鏡像となって重複した構造の幼虫が誕生します。この体節化の過程と並行して、ホメオティック遺伝子と呼ばれる一群の遺伝子が、体節の違いを決定し維持する働きをします。
図13)遺伝子カスケードによるショウジョウバエの胚パターン形成

(出典:ウィキメディア・コモンズより改変)
体節形成過程では、母性効果遺伝子の産物が全体的な位置シグナルとなり、特定の領域で特定のギャップ遺伝子を発現させます。次に、ギャップ遺伝子の産物が第 2段階の位置シグナルとなり、さらにペアルール遺伝子など別の遺伝子の発現を調節して、詳細なパターンを段階的に形成して行く訳です。この時、多くの遺伝子の発現によって様々なシグナル分子の濃度勾配の重なりが生まれます。分節遺伝子は、調節領域に結合した多数の調節タンパクの組み合わせを識別して正確に遺伝子をオン・オフする事で、精密な空間パターンの決定が可能となっているのです。
またこの過程は、 母性効果遺伝子から分節遺伝子へ、そしてホメオティック遺伝子(Hox遺伝子)へと続く、遺伝子発現のカスケードとして起こっています。最初の母性効果遺伝子産物がギャップ遺伝子の転写を促進し、 ギャップ遺伝子産物はペアルール遺伝子の転写を促進し、ペアルール遺伝子産物はセグメントポラリティ遺伝子の転写を促進するという具合に、転写因子のカスケードが出来上がっているのです。
全ての動物が共有する Hox 遺伝子群
ホメオティック遺伝子は、胚の長軸に沿って異なる組み合わせで発現する転写因子をコードしており、各体節にどの器官を形成するかの決定に関与しています。この遺伝子に変異が起こると、頭部に触覚の代わりに脚が生えたり(アンテナペディア変異)、 胸部体節にもう一対余分な翅が生えたり(ウルトラバイソラックス変異)というホメオティック変異を引き起こします。
ショウジョウバエのホメオティック遺伝子は、 第 3染色体上でアンテナペディア遺伝子群とバイソラックス遺伝子群の 2つの遺伝子群として並んでいます。 アンテナペディア遺伝子群は頭部から中胸部、バイソラックス遺伝子群は後胸部から腹部を支配し、それぞれ 5個と 3個の遺伝子から構成されますが、驚いた事に染色体上でのこの遺伝子の並びと、体軸に沿って遺伝子が発現する順序とが全く同じ順序で並んでいるのです。これが単なる偶然なのか、染色体上を伝わって行く何らかの活性化機構が存在するのかは良く分かりません。
図14)ショウジョウバエのホメオティック遺伝子群とその発現

(出典:ウィキメディア・コモンズ)
この 8個のホメオティック遺伝子は、母性効果遺伝子・分節遺伝子産物のタンパク質の結合部位が存在する、65万塩基対に及ぶ非常に長い調節 DNA 領域の中に散在しています。そして、この調節領域に結合した複数の誘導因子がもたらす位置情報を多元的に解釈する事で、体節ごとに特定のホメオティック遺伝子セットが発現するのです。こうして発現したホメオティック遺伝子は、さらにその次の遺伝子群に発現の連鎖を引き起こし、体節ごとに固有の器官を作り出して行く訳です。
これらのホメオティック遺伝子は、ホメオボックスと呼ばれる 180塩基対の共通の配列を持っています。それがコードする 60アミノ酸配列はホメオドメインと呼ばれ、そのヘリックスターンヘリックスの 3次元構造によって標的遺伝子の DNA の溝に結合します。ホメオティック遺伝子産物の転写因子は、ホメオドメインによって標的遺伝子のエンハンサー配列に結合し、転写を調節しているのです。このホメオボックス配列を持つ遺伝子を、Homeobox から Hox 遺伝子群と呼んでいるのです。
図15)ホメオドメインの α ヘリックスが DNA の溝に結合

(出典:ウィキメディア・コモンズ)
実は、このホメオドメイン、ホメオボックスは、動物の進化の過程で高度に保存されている事が知られています。ショウジョウバエのホメオティック遺伝子と相同性のある遺伝子が、ヒドラ・線虫・ミミズ・甲虫・軟体動物・ウニ・魚・カエル・鳥・哺乳類と、あらゆる動物で見つかっているのです。そしてこれらの遺伝子は、ショウジョウバエのアンテナペディア遺伝子群とバイソラックス遺伝子群を合わせた、Hox 複合遺伝子とよく似た複合遺伝子としてまとまっています。哺乳類では、ショウジョウバエの Hox複合遺伝子とよく似たものが、別々の染色体上に 4つ存在し、HoxA・HoxB・HoxC・HoxD 複合遺伝子と呼ばれています。そして哺乳類の 4つの Hox複合遺伝子のそれぞれが、昆虫の Hox複合遺伝子に相当しています。しかも、各 Hox複合遺伝子内での遺伝子の並び方が、基本的に昆虫のそれと同じなのです。そして発現パターンも、ショウジョウバエと同様に、各複合遺伝子の構成遺伝子が体軸に沿って頭部から尾部へと順に並んで発現しています。これらの事から、脊椎動物の 4個の Hox複合遺伝子は、原型となる祖先の一個の複合遺伝子から重複によって生じたと考えられます。
この遺伝子重複と言うのは、いわば遺伝子のコピー&ペーストで、コピーされた遺伝子に変異を蓄積させる事で新しい機能を獲得させる、遺伝子の進化実験を行っていると考えられています。先に紹介した偽遺伝子は、この進化実験の成れの果てだったのです。また遺伝子重複では、遺伝子の重複だけでなく、ゲノム全体が重複する事も珍しく有りません。Hox複合遺伝子では複合遺伝子全体が重複し、その後それぞれに変異が蓄積して、異なる働きをする様に進化して行ったと言う訳です。
図16)Hox複合遺伝子の進化(上から線虫・ショウジョウバエ・ナメクジウオ・マウス/ヒト)

(出典:ウィキメディア・コモンズ)
Hox複合遺伝子は全ての左右相称動物と、イソギンチャクなどの刺胞動物にも存在する事から、左右相称動物と刺胞動物が分岐した 5億 5000万年以上前に進化したと考えられます。これは、動物の発生過程で非常に重要な役割を果たしているHox複合遺伝子が、5億 5000万年もの長きに渡って大切に保存されて来た事を示しています。進化によって体の外見が全く異なってしまった動物の間にも、発生の基本機構は高度に保存されていたのです。しかも、単に並び方が同じというだけではなく、遺伝子の機能自体も変わっていない場合が有るのです。
例えば、昆虫の複眼などは、ヒトの眼とは構造上全く異なっています。ところが、眼の胚における形成では、両者で共通の発生経路が関与していると言うのです。 ”eyeless(眼なし)と呼ばれる、眼の無いショウジョウバエの変異体が有るのですが、これは眼の発生に必要な遺伝子の発現を制御する転写因子の ”eyeless 遺伝子の突然変異によって起こります。この ”eyeless 遺伝子を異なる胚組織で強制的に発現させると、脚・触角・翅の下など、様々な体の部位に余分な眼を作る事が出来ます。この ”eyeless 遺伝子によく似た配列の遺伝子(”Pax6)が、マウスにも見つかっているのです。しかも驚いた事に、マウスの ”Pax6 遺伝子をショウジョウバエの胚で発現させると、”eyeless 遺伝子と同じ様に、様々な部位に眼を発生させる事ができるのです。哺乳類の眼を発生させる遺伝子によって、系統上大きく異なる昆虫の全く異なる種類の複眼が作られた訳です。
”eyeless 遺伝子と ”Pax6 遺伝子は、この 2種だけではなく、他の種においても高度に保存された配列を含んでおり、この様な遺伝子を相同遺伝子と呼んでいます。近年、非常に多くの相同遺伝子が、縁遠い動物種で発生を制御している事が明らかになって来ています。アンテナペディアやバイソラックスなどの、ショウジョウバエのホメオティック遺伝子も、マウスやヒトの発生において同様の役割を果たしている遺伝子と似通っているのです。これは、発生を制御する特定の配列から構成される遺伝子ツールキットが、幅広い動物種の中で高度に保存されると同時に、進化の中で繰り返し利用されてきた事を示唆しています。そして、この遺伝子ツールキットを様々に組替え、あるいは遺伝子重複によって新たな機能を生み出しながら多様化させて、現在の我々が見る驚くべき多様性に富んだ、地球の生態系を進化させて来たと考えられるのです。
多細胞生物の進化
ややこしい説明が長々と続いて、辟易した人も多いかも知れません。しかし、いよいよ結論に近づいてきました。ここで多細胞生物の進化について考えてみましょう。 多細胞生物はすべて、一個の受精卵から発生過程を経て誕生します。従って多細胞生物の進化とは、見方を変えればその発生過程の変更と捉える事ができます。しかも、多細胞生物は 5億 5000万年以上もの長きにわたって、器官形成を制御するホメオティック遺伝子群を高度に保存し、使い続けてきた訳です。勘のいい人は、もうお分かりですね。 胚の発生過程で「個体発生は系統発生を繰り返す」様に見えたのは、多細胞生物が祖先の発生メカニズムを繰り返し利用しながら、その最後の部分を改変し、あるいは改善・拡充して行く事で進化して来た結果だったのです。多細胞生物の進化とは、祖先の発生プログラムを利用しながら一部を書き換え、あるいは新たなプログラムを付け加える事で、発生メカニズムをより高度化して行く過程だったのです。そのために、祖先の発生過程の一部をなぞる様にして、個体発生が進められている訳なのです。
そして、この多細胞生物の進化を規定している発生プログラムは、遺伝子発現のカスケード、遺伝子のネットワークとして書かれている訳です。実際、遺伝子発現を変化させる事で、個体の形や機能の進化が起こっている事が知られています。以下に。幾つか例を挙げましょう。
例えば、ショウジョウバエの翅の発生を見てみましょう。ほとんどの昆虫は 2対の翅(4枚の翅)を持っていますが、ショウジョウバエは 1対の翅(2枚の翅)で双翅目と呼ばれています。双翅目昆虫の 1対の羽は、Hox遺伝子のアンテナペディア(”Antp)が発現して第 2胸部体節に発生します。実は、第 3胸部体節でも ”Antp 遺伝子は発現しているのですが、同じ Hox遺伝子のウルトラバイソラックス(”Ubx)も発現して ”Antp を抑制する結果、平均棍と呼ばれる 1対の小さな突起が翅の代わりに発生するのです。従って、”Ubx を不活性化すると、第 3胸部体節にも 2番目の翅が発生する事になります。このように遺伝子発現の比較的小さな変化が、 種間の大きな形態の違いを生み出しているのです。
図17)ショウジョウバエの翅(7)と平均棍(3)

(出典:ウィキメディア・コモンズ)
また、チャールズ・ダーウィンが進化論を思いつく切っ掛けとなったと言われる、ガラパゴス諸島のダーウィンフィンチ類は、食性の違いに応じてクチバシの形状が大きく異なっています。オオガラパゴスフィンチは硬く大きな種子を砕く為に大きくがっしりしたクチバシを持ち、昆虫食やサボテンの花・果実を 食べる種は、細く長いクチバシをしています。このクチバシの形状は発生過程で決定されます。 クチバシが発生する時に、顔面骨を作る胚の前部にある組織の細胞分裂をコントロールする事によって、クチバシの形状を変化させているのです。それには、2種類のシグナルタンパク質が関与しています。一つは骨形成タンパク質の BMP4 で、これが早期に大量に存在するとクチバシは広く厚くなり、もう一つのカルモジュリン(CaM)が早期に大量に存在すると、クチバシは細く長くなるのです。つまり、この 2種類のタンパク質の遺伝子の発現調節によって、クチバシの形状が決定されている訳です。
図17)ダーウィンフィンチのクチバシ(1.オオガラパゴスフィンチ、2.ガラパゴスフィンチ、3.コダーウィンフィンチ、4.ムシクイフィンチ)

(出典:ウィキメディア・コモンズ)
進化論でよく引き合いに出されるキリンの長い首も、遺伝子の発現調節によって説明できます。キリンの頸椎の数は、ほとんどの哺乳類と同じ 7個で、首が長いのは一つ一つの頚椎が長くなっている為です。哺乳類の骨成長は、軟骨細胞と呼ばれる軟骨生成細胞の増殖によって起こり、軟骨細胞のアポトーシス(細胞死)と骨基質の石灰化に至る遺伝子シグナルによって成長は停止します。キリンでは、骨成長停止のシグナル過程が遅れて、頚椎が長く成長しているのです。つまり、キリンの長い首の進化は、骨成長を制御する遺伝子発現のタイミングの変化によって起こったのです。
鳥の足の水かきの有無も遺伝子発現が関わっています。カモなどの水鳥の足には水かきが付いていますが、ニワトリには有りません。実は、全ての鳥類の胚で、足指間をつなぐ水かきが存在しています。ところが、ニワトリなどの陸鳥では、発生過程で水かきの細胞がアポトーシスを起こして消失してしまうのです。これには、クチバシの発生にも関与していた、BMP4 タンパク質がアポトーシスを誘導しています。カモ・ニワトリとも ”BMP4 遺伝子は発現するのですが、カモでは ”Gremlin 遺伝子も水かき細胞で発現して ”BMP4 の発現を抑制するする結果、アポトーシスを促進する BMP4 タンパク質が産生されず、水かきの残った足が発生するのです。
体長10 cm ほどのトゲウオ科のイトヨ(糸魚)は、川で生まれた稚魚が海に下って成長し、産卵前に再び川を遡行する回遊を行いますが、中には淡水域に留まって成長する個体も有ります。海洋性のイトヨは、捕食性の海洋魚から身を守るために、背中に 3本の背側突起、腹に 2本の骨盤突起を発達させています。一方、こうした捕食者の危険のない淡水性のイトヨでは、これらの突起はずっと短いか全く欠けています。この差は、骨板と突起の産生を促進する転写因子をコードする、”Pitx1 遺伝子の発現の違いによって生まれます。海洋性のイトヨでは、頭部・体幹・ 尾部・ 骨盤で ”Pitx1 が発現しますが、 淡水性のイトヨでは不活性化されて突起が生えてこないのです。
図18)イトヨの背側突起と骨盤突起

(出典:ウィキメディア・コモンズ)
このように形態の進化は、革新的な新しい遺伝子の誕生によってではなく、既存の遺伝子の発現パターンを変化させる事で起こされています。生物の進化は、祖先の胚発生プログラムを利用しながら、遺伝子発現のカスケード、そのネットワークパターンを変更し、あるいは新たな遺伝子発現のネットワークを構築して行く事で進められて来たのです。重要なのは個々の遺伝子ではなく、遺伝子間の相互作用のネットワークであり、その組み換え・新規創造・重層化だったのです。生命活動の本質が、生体高分子間の相互作用で有った様に、進化に於いても重要なのは遺伝子間の相互作用、その絡まり合ったネットワークシステムの構築に有るのです。
新しい遺伝子を進化させる場合でも、遺伝子重複を利用して組織的に実施していました。古い遺伝子は保持したままで、そのコピーに様々な変異を蓄積させる事で、安全に遺伝子の進化実験を行なっている訳です。また突然変異自体も、生物自身がコントロールしている可能性が有ります。ヒトゲノムの 40%以上を占めるトランスポゾンが、遺伝子の突然変異に関与している可能性が有るのです。
また最初に、ヒトゲノムにはレトロウイルスに由来する DNA が、5~ 8%も組み込まれていると述べましたが、このレトロウイルスは異種間で遺伝子を水平移動させる事で、生物の進化に重要な役割を果たして来たと考えられています。レトロウイルスは RNA を遺伝物質として持つ RNA ウイルスの一種で、宿主細胞に感染すると逆転写酵素で RNA から DNA のコピーを作成し、それを宿主ゲノム内に挿入します。そして宿主細胞の転写機構を利用して、子ウイルスの RNA ゲノムを合成させているのです。このように宿主ゲノムに組み込まれた、レトロウイルス由来の DNA をプロウイルスと呼んでいます。ところが、このプロウイルスは再び宿主から飛び出す事が有り、その時に宿主の遺伝子の一部を一緒に持ち出す場合が有るのです。このレトロ ウイルスが、他の生物に感染してゲノム内に入り込むと、前の宿主の遺伝子を別の生物に運び込む事になる訳です。もし、この感染相手が生殖細胞の場合は、持ち込まれた遺伝子はその子孫にまで受け継がれて行く事になります。
このようなレトロウイルスによる遺伝子の水平移動は、自然界の遺伝子操作とも呼べるもので、バイオテクノロジーではこの性質を利用して、レトロウイルスをベクター(遺伝子の運び手)として使っているのです。昨今では、ウイルスを目の敵にする風潮も有る様ですが、生命誕生以来 40億年の長きにわたって生物とウイルスは共存してきた訳で、実は生物にとってなくてはならない存在でも有るのです。
このように生物は新しい遺伝子の進化に於いても、偶然に起こる突然変異などに頼るのでは無く、 極めて組織的・合理的な進化のシステム自体を進化させて来た訳です。ここには、生存競争や自然淘汰と言った進化論者の世迷言が、入り込む余地など全く無い事が分かると思います。
(つづく)
① 遺伝子とは何者か?
③ 生存闘争は進化を推進するか?
④ 生命は多様性を目指す
⑤ 激変する地球環境の下で生命は誕生し、進化してきた
⑥ カンブリア爆発:捕食者の出現
⑦ 気候変動と大量絶滅

この記事が気に入ったらサポートをしてみませんか?