
アキノヒメミノガ―身近なミノムシの謎の多い生態―

はじめに
アキノヒメミノガ Bacotia sakabei Seino, 1981という虫がいる。ミノガ科(いわゆるミノムシ類)に属する小型で焦げ茶色の地味な蛾である。ミノムシ類は、幼虫が植物質の材料を綴り合せて作ったポータブルケース(ミノ)を背負って暮らす生態で広く知られている。また、下手に自然度の高い所よりも、人間の居住環境近くの方が多く見られるため、蛾類の中では特に身近な仲間である。その中で、アキノヒメミノガは、ガードレールという人工物に生息環境を見出して、ガードレール上で生活史を完結させているという興味深い生態を持つ。都市環境でも見られる身近な種であるが、その詳細な生態は意外にも未解明のため、私は本種の生態について調査を続けている。これまでに分かった成虫の繁殖行動と、孵化した幼虫の分散方法について、簡単に紹介したい。
基本的な生態
アキノヒメミノガの幼虫は地衣食性で、地衣類が生着した石垣や墓石、石碑等で、幼虫のミノが付いているのがよく見つかるが、後述する通り、陸生藻類が生着したガードレールで特によく見つかる。幼虫は春先に孵化し、秋にミノの中で蛹化、晩秋に羽化する。成虫の出現期間は非常に短い。メスは完全な無翅で、自分が出てきたミノにつかまったままフェロモンを出してオスを呼ぶ。交尾後、メスは自分のミノの内部に産卵し、卵で越冬する。ミノガ科小型種に関する生態情報は乏しく、専門的な図鑑や文献を見ても、生態に関する記述はこの程度である。
ガードレール上での生活
本種はとにかくガードレールでよく見つかる。暗緑色の陸生藻類が生着して汚れたように見えるガードレールが狙い目である。同じように汚れたガードレールでも、場所によっている/いないがはっきり分かれており、いる場所には、下写真のように多数のミノが付いていることが多い。


ミノの長さは7 mm程度。
終齢幼虫のミノの長さは6~7 mm程度であるが、当然ながら、幼齢幼虫のミノは小さい。4月くらいに生息地のガードレールを見て回ると、長さ1 mmにも満たないチビミノを被った幼虫を見つけることができる(下写真)。季節が進むにつれて、ミノが大きくなっていく様子を観察できる。

右の大きい方(たぶんメス親のもの)は空ミノ。
11月下旬から12月上旬くらいの晩秋の一時期に、羽化した無翅のメス成虫が自分のミノにつかまっているのを見つけることができる(下写真)。

オスをフェロモンで呼んでいる。
疑問
このように、本種はガードレール上で生活史を完結させているように見えるのだが、私は以下のような疑問を持った。
①オス成虫はいつ出てくるのか?
晩秋に生息地を見て回ると、上写真のようにコーリングしているメスは昼夜問わず多数見つかるのだが、オスがなかなか見つからない。出現時間帯が限られるのか?
②孵化した新幼虫はどのように分散するのか?
分散とは、生物が生息域を広げることである。本種のようにメスが移動能力を失っている場合、メスは自分が羽化した場所で産卵するため、生まれた新幼虫をいかに適度に分散させるかが、種の存続上重要な問題となる。ミノガ科大型種のオオミノガやチャミノガの場合、新幼虫は、親ミノから脱出後、糸を出してぶら下がって風に吹かれて分散するバルーニングを行うことが知られているが、本種も同様の行動をするだろうか?
③ガードレールに最初に到達した個体はどこから来たのか?
本種は、ガードレールに多くの個体が生息していても、その周辺の環境(樹木とか草むらとか)では全く見つからない。最初の個体はどこからどうやってガードレールに到達したのでしょう?
オス成虫の出現時間帯
蛾の成虫というと夜行性のイメージが強いだろう。昼行性の種もいるが、そのような種は、オスとメスが明るい環境で視覚を用いてお互いを識別するためか、翅に色鮮やかな紋様があることが多い。本種のオスの翅は紋様のない焦げ茶色一色のため、当然夜行性だろうと予想して、2020年11月下旬の夜間に生息地のガードレールを見て回ったが、見つかるのはミノにつかまっているメスばかりで、オスはほとんど見つからなかった。小松貴さんの著書「裏山の奇人」で、キタクロミノガの交尾が日の出前の早朝に行われるのを偶然発見した話があったので、同じように早朝にも見て回ったが、やはりオスは見つからない。
ひょっとして、オスが出てくるのは日中か?
気を取り直して、翌2021年11月下旬の週末に、生息地のガードレールで午前中から張り込み調査を行ったところ、あっけなくオス成虫が見つかった。そこではメスが20頭以上羽化してオスをコーリングしていたが、オスは1時間当たり2,3頭気紛れに飛来してくる感じであった。飛来したオスは、あまり目が見えているようには見えず、メスが出すフェロモンを頼りにメスを探しているように見えた(下写真)。

ガードレールに着地し、翅をパタパタと震わせながら、メスの方に近づいていく。


こうして、ようやく念願の交尾中のペアの写真を撮ることができた。交尾の時間は長く、どのペアも1時間程度続いていた。


この観察で気付いたが、オスはメスとほぼ同時期にほぼ同数羽化しているはずなのに(オスが羽化したミノには蛹の脱皮殻が残るので判別可能)、オスは大変気紛れで、飛来してくる個体数がメスよりも明らかに少ないのである。実際、交尾できずにミノにつかまったまま絶命したメスが多かった。何とも不思議な生態である。
幼虫の分散方法
次は、幼虫の分散方法である。果たして、本種の幼虫は、ミノガ科大型種と同様に親ミノから脱出した直後にバルーニングを行うのだろうか?分散の現場を押さえるべく、2021 年と2022 年の春先に近所にある本種の生息地ガードレールを日中にしつこく見て回ったが、親ミノから脱出直後の新幼虫はまったく見つからず、新幼虫が見つかったとしても、図3のように親ミノのごく近傍に微小なミノが点々と集まっている状態ばかりであった。
飼育下での観察
野外での観察では埒が明かなかったので、飼育下での観察を試みた。2022 年3 月に、ガードレールから採集したメスのミノ(中に卵が入っている)を大型の半透明プラスチック製タッパーの底面に垂直な向きに接着し、底面が垂直になるように壁に立て掛けて保管した(下図)。孵化した幼虫の餌とするために、地衣が生着した樹皮も貼り付けておいた。

1週間ほど待つと、ミノから次々と微小な新幼虫が出てきた。脱出した幼虫は親ミノ表面に留まり、親ミノを盛んに齧って自分のミノを作り始めた(下写真)。


親ミノを材料に自分のミノを作り始めている。
ミノが完成した後は、そのまま親ミノ表面に留まる個体(おそらく親ミノを摂食していた)と、タッパー内を徘徊する個体の両方がいたが、糸を引いて親ミノからぶら下がるような行動は見られなかった。タッパー内を徘徊している幼虫を食餌の樹皮に移すと、幼虫は盛んに地衣を摂食し始め、日に日に成長してミノが少しずつ大きくなっていく様子を観察できた(下写真)。樹皮に100 頭超の幼虫を移して生息密度が高くなってくると、樹皮から糸を引いて落下する個体が頻繁に見られるようになった。

バルーニング誘発実験
上記の観察から、本種の幼虫は、親ミノからの脱出後に一斉にバルーニングを開始するような生態ではなく、生息上不利な状況になったときのみバルーニングでその場から脱出を試みる生態であることが推察された。もはや調べるまでもないかもしれないが、念のため以下のような実験を行い、この仮説の検証を試みた。
竹串の先端に地衣が生着した樹皮片を接着したもの(餌あり)と,先端にボンドのみ付けて乾かしたもの(餌なし)の2つを用意し、大型タッパーの底面に垂直に立てて固着した(下図)。

餌ありbaitedと餌なしNon-baitedの2条件の竹串の先端に幼虫を移し、バルーニングを誘発する。竹串先端部以外は虫除けスプレーを塗ってあるので、幼虫は竹串を伝って逃げられない。
タッパー底面が垂直になるように壁に立て掛け(竹串は水平になる)、2 本の竹串先端に幼虫を移して、餌あり/餌なしの2 条件で、バルーニングを始めるまでの時間$${T}$$を計測した。竹串の先端以外には虫除けスプレーの液を塗布し、幼虫が竹串先端から歩いて脱出できないようにした。
餌なし群25 頭、餌あり群13 頭の$${T}$$の計測結果を下図に示す。
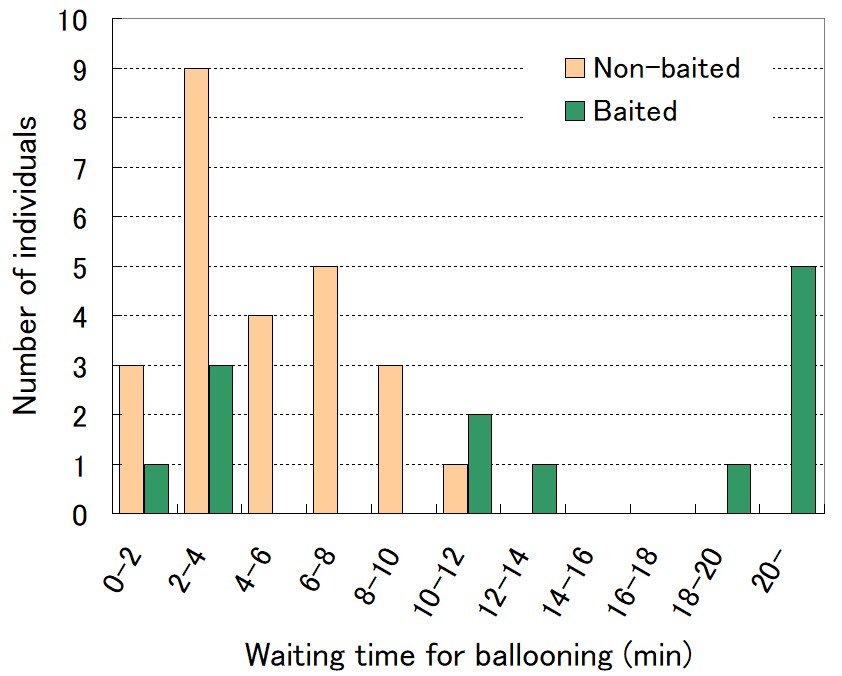
餌なし群では3分程度でバルーニングを始める個体が最も多く、最長でも10分であったが、餌あり群では、地衣表面で静止して20分以上経過してしまう個体がもっとも多かった。$${T}$$ の平均はそれぞれ$${T_\text{non-baited}=4.8\pm 2.8}$$分、 $${T_\text{baited}=12.4\pm7.8}$$分となり、明らかに有意な差が見られた。餌なし群の個体は、竹串に移すと、先端の狭い部分を行き場なく徘徊した後、あきらめたように糸を引いて落下した(下写真)。

餌あり群の個体では、竹串に移すと、すぐに虫除けスプレーが塗布された部分に進入した個体や、樹皮の縁を徘徊中に足場を失った個体が糸を引いて落下したが、その他の個体は樹皮上で落ち着いていた。
考察
今回の野外及び飼育下での観察からの結論として、アキノヒメミノガの幼虫は、バルーニングではなく、親ミノから歩いて周辺に分散していくようである。ミノガ科小型種の幼虫の分散の生態は、世界的にもほとんど知られていないため、けっこう重要な知見が得られたのではないかと自負している。
さて、上述したように、オオミノガやチャミノガのような大型種では、幼虫はバルーニングで分散するのに対し、アキノヒメミノガの幼虫が歩いて分散するというこの違いは何を意味するのか?私は、これは幼虫の食性と密接に関係していると考えている。ミノガ科大型種の幼虫は総じて広食性であり、新幼虫は、風に吹かれて着地した場所に植物さえあれば生存することができる。一方、アキノヒメミノガを含む小型種は総じて地衣食性である。地衣食性の幼虫が不用意にバルーニングで分散すると、食餌が得られない場所に着地してしまう可能性が相当高そうであることは想像に難くない。また、今回の観察から分かったが、アキノヒメミノガでは、1 つの親ミノから出てくる孵化幼虫はせいぜい30 頭程度にすぎないため、バルーニングを主たる方法として分散すると容易に全滅してしまうだろう(バルーニングするオオミノガの産卵数は三千個!)。そもそも、親ミノが固着している場所の周囲には、食餌となる地衣が生着している可能性が高いため、基本的に歩行によって分散するのは合理的な戦略であるといえよう。
おわりに
今回の観察で、上に挙げた3 つの疑問のうち2つは分かったが、ガードレールに生息する個体はそもそもどこから来たのかという3つ目の疑問は未解決である。本種がガードレールに生息している以上、どこか周囲の環境から最初の個体が到達したはずであるが、そもそも本種は自然環境でそれほど普遍的に生息しているのだろうか。フィールドを歩くときはあちこち注視するようにしているが、自然環境では本当に見つからないのである。ミノガ科の謎解きの旅は続く。
※本稿の詳細については、以下を参照されたい:
齋藤孝明, 2022. アキノヒメミノガの成虫期. 誘蛾燈 (247): 22-23.
齋藤孝明, 2022. アキノヒメミノガの幼虫期(I)―新幼虫の分散について―. 誘蛾燈 (249): 137-141.
この記事が気に入ったらサポートをしてみませんか?