
ニホンイシガメ

ニホンイシガメ Mauremys japonica は日本固有種である。今回はこのカメを紹介する。

分布
本州、四国、九州及び周辺島嶼(佐渡島、壱岐、対馬、淡路島、五島列島など)に広く分布する。Yasukawa et al(2008)によると、東限は千葉県・茨城県、北限は新潟県、南限は鹿児島県となるようだ。東北地方からも分布が確認されているが、これらは人為的な移入によるものと考えられている。
生息環境

平地から丘陵地にかけての河川、溜池や湿地に生息する。灌漑期には河川や溜池に隣接する水田を餌場として利用する。ただし、現在の分布は丘陵地に集中し、平地で見られるのことは稀である。
形態
背甲は橙色、腹甲は黒色をしている。背甲には一本の隆起があり、後縁は鋸歯状になっている。四肢や尾には黄色から薄い橙色の線が入り、虹彩は真っ黒に見える。体サイズは雌雄で異なり、オスは背甲長が145mmほど、メスは200mmほどまで成長する。

孵化幼体(0歳)
孵化幼体は35mmほどの大きさで、背甲の後縁部は鋸歯が発達する。

オス(成体)
オスは尾の根本が太く、メス(成体)と比べて体高が低いのが特徴である。

メス(成体)
オス(成体)に比べ、体高が非常に厚い。

雌雄判別は総排出腔の位置から行う。総排出腔が背甲よりも大きく離れていればオス、ほぼ背甲の端と同じくらいの位置にあればメスと判断する。また、オスの尾には生殖器が入っているため、尾の根本は太くなっている。
*メスの総排出腔付近が白くなっているのは皮膚病のためです。

腹甲は雌雄ともに真っ黒。
食性
動物から植物まで幅広く捕食する雑食性を示すが、水中由来よりも陸生由来の餌を捕食する傾向にある。具体的には、甲虫目,トンボ目,セミ目,ハエ目,バッタ目,鱗翅目の幼虫などの昆虫,ニナ目等の淡水生貝類,マイマイ目等の陸生貝類、サワガニやエビ目など甲殻類、魚類、イネ目、ガマ目、フトモモ目、マメ目や藻類などを捕食することが知られている。
また、哺乳類、鳥類、爬虫類や昆虫などの死体も摂食することが知られている。具体的には、フナ、オイカワ、カラス、ニホンイシガメなどの水生生物、ヤマカガシなどの陸生生物、トンボ目やセミ目の成虫などの飛翔昆虫の死体を摂食することが知られている。
捕食者
卵から幼体にかけた若齢期には多くの在来捕食者に襲われる一方で、成体になると主要な在来捕食者はいなくなる。具体的には、若齢期にはイタチ、カラス、シマヘビなどの在来種、ウシガエルやアメリカザリガニなどの外来種に捕食される。一方で、成体の在来捕食者はほとんど知られていないが、近年になり、外来種のアライグマが新たな主要捕食者となってしまった[関連記事]。
繁殖(準備中)
繁殖期、繁殖年齢、クラッチサイズ等については後日書き足します。
行動(準備中)

年間の行動と性差、移動性と回帰性については後日書き足します。
最大移動距離は30-210mで(平均100m)、行動圏の面積は100㎡-12,640㎡(平均は4,000㎡)。
成長
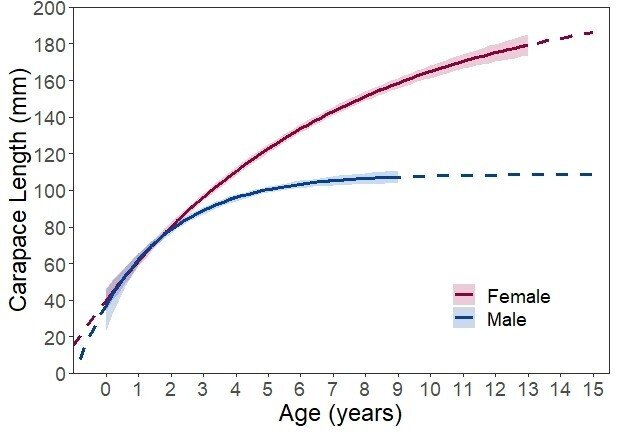
丘陵地に生息する個体群の成長曲線(von Bertalanffy growth curve)
雌雄で異なる成長パターンを示す。オスは背甲長が100mmを超えるあたりから成長が鈍化する一方で、メスは鈍化することなく成長を続ける。
また、環境によって体サイズや成長速度が異なり、これは水温・気温(活動期間)や餌生物(種類や量)、同種の生息密度など様々な要因によるものと考えられている。
性決定様式
ニホンイシガメは孵卵時に曝露される温度によって性別が決まる性決定様式(温度依存性決定)を示します。孵卵時の温度が低温(28℃以下)だとオス、高温(30℃以上)だとメスに分化し、性比(オス:メス)が1:1となる温度(臨界温度)は28.8℃と算出されています(Okada et al 2010)。
この性決定様式を反映してか、環境の異なる溜池では性比が著しく異なることが報告されています。周囲に林が多く、産卵場所が低温となる溜池ではオスが多く生じ、周囲が開け産卵場所が高温となる溜池ではメスが多く生じると指摘されています(Okada et al 2011)。
越冬(準備中)
越冬時期、越冬環境、雌雄の低温耐性の違いについては後日書き足します。
参考文献
Haramura, T., Yamane, M., and Mori, A. 2010. Radiotelemetric study of movement patterns of lotic freshwater turtles during breeding and hibernation seasons. Journal of Freshwater Ecology 25: 251-259.
加賀山翔一. 2020. 湿地生態系におけるニホンイシガメの役割と保全すべき理由の整理. 亀楽 19: 8-12.
Kagayama, S. 2020. Geographic Variation in the Growth of Japanese Pond Turtles, Mauremys japonica, in the Flatland and Mountain Regions of Chiba Prefecture, Japan. Current Herpetology 39: 87-97.
Kagayama, S., Ogano, D., Taniguchi, M., Mine, K., Ueno, S., Takahashi, H., Kamezaki, N. and Hasegawa, M. 2020. Species Distribution Modeling Provides New Insights into Different Spatial Distribution Patterns among Native and Alien Freshwater Turtles in Japan. Current Herpetology 39: 147-159.
加賀山翔一・小賀野大一. 2021. 日本に生息する淡水性カメ類の捕食者に関する文献調査. 爬虫両棲類学会報 2021: 36-43.
Okada, Y., Yabe, T., and Oda, S. 2010. Temperature-dependent sex determination in the Japanese pond turtle, Mauremys japonica (Reptilia: Geoemydidae). Current Herpetology 29: 1-10.
Okada, Y., Yabe, T., and Oda, S. 2011. Interpopulation variation in sex ratio of the Japanese pond turtle Mauremys japonica (Reptilia: Geoemydidae). Current herpetology, 30(1), 53-61.
竹原康史. 1996. 鈴鹿・亀山のヌマガメ類の生態. 三重自然誌 3: 17-21.
谷口真理・佐藤由佳・角道弘文. 2021. ため池及びその周辺における日本固有種ニホンイシガメの生息に影響を及ぼす環境要因の推定. 農業農村工学会論文集 89: I_19-I_27.
Yabe, T. 1989. Population structure and growth of the Japanese pond turtle, Mauremys japonica. Jpn. J. Herpetol 13: 7-9.
Yabe, T. 1992. Sexual difference in annual activity and home range of the Japanese pond turtle, Mauremys japonica, assessed by mark-recapture and radio-tracking methods. Jpn. J. Herpetol 14: 191-197.
カメは売らずにカメでお金を稼ぎたい。そんなお年頃です。