自閉症に関連した食事の嗜好が自閉症と腸内細菌叢の関連を媒介する

メインコンテンツへスキップ
広告
インスピレーションを与える科学Cell Pressジャーナル
セル
ログイン
検索...
論文|2021年11月24日、184巻、24号、p5916-5931.e17
全号ダウンロード
自閉症に関連した食事の嗜好が自閉症と腸内細菌叢の関連を媒介する
https://www.cell.com/cell/fulltext/S0092-8674(21)01231-9?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867421012319%3Fshowall%3Dtrue
クロエ・X・ヤップ
アンジャリ・K・ヘンダース
ゲイル・A・アルバレス
アンドリュー・J.O.・ホワイトハウス
ナオミ・R・レイ
ジェイコブ・グラッテン20
すべての著者を表示
脚注を表示するオープンアーカイブ発行:2021年11月11日DOI:https://doi.org/10.1016/j.cell.2021.10.015
PlumXメトリクス
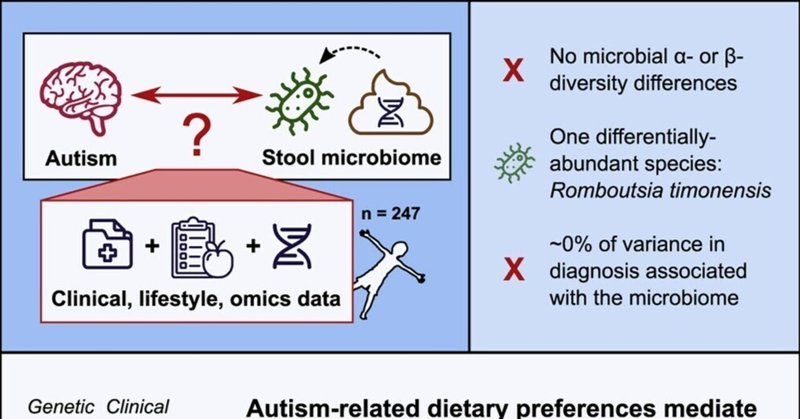
ハイライト
n=247人の小児の便メタゲノミクスから得られた自閉症とマイクロバイオームの限定的な関連性
Romboutsia timonensisは自閉症診断に関連する唯一の分類群であった。
興味の制限などの自閉症特性は、多様性の少ない食事と関連している。
多様性の低い食事は、マイクロバイオームのα多様性の低さと関連している。
まとめ
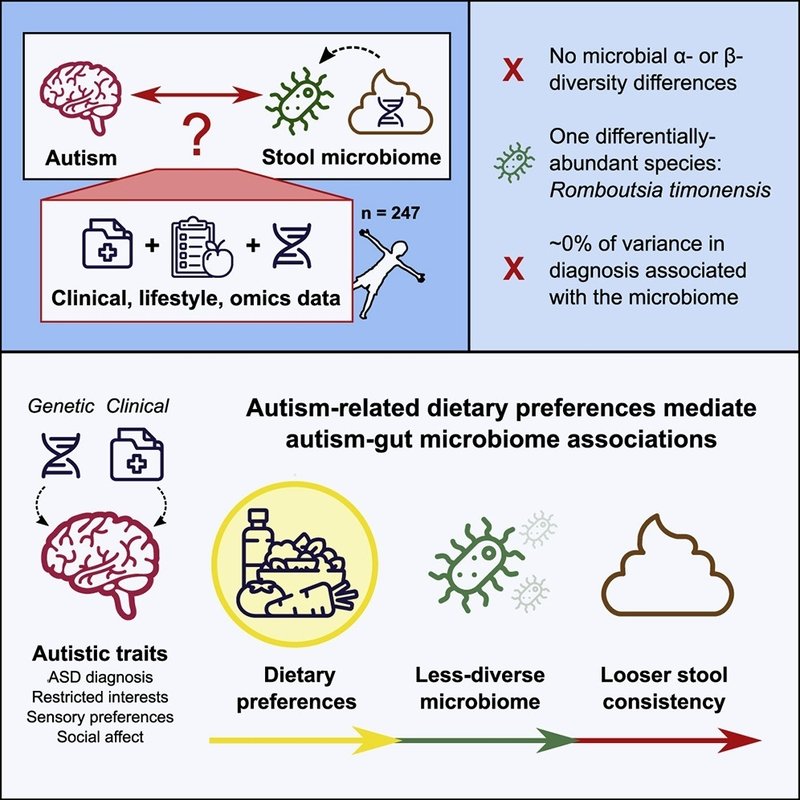
自閉症スペクトラム障害(ASD)に対する腸内細菌叢の寄与の可能性に関心が高まっている。しかし、これまでの研究は検出力不足であり、潜在的な交絡因子を包括的に扱うようにはデザインされていなかった。我々は、Australian Autism BiobankとQueensland Twin Adolescent Brainプロジェクトの参加者をもとに、大規模な自閉症便メタゲノム研究(n=247)を行った。その結果、ASDの診断と腸内細菌叢との直接的な関連はごくわずかであることがわかった。その代わりに、我々のデータは、ASDに関連した制限された関心が、多様性の少ない食事と関連し、その結果、微生物の分類学的多様性が減少し、便の粘度が緩くなるというモデルを支持した。ASD診断とは対照的に、我々のデータセットは、年齢、食事摂取量、便の硬さなどの形質とマイクロバイオームとの関連を検出するのに十分な検出力を有していた。全体として、ASDにおけるマイクロバイオームの相違は、診断上の特徴に関連する食事の嗜好を反映している可能性があり、マイクロバイオームがASDにおいて推進的な役割を担っているという主張には注意が必要である。
図解抄録
図サムネイルfx1
大きな画像を見るダウンロード 高解像度画像
キーワード
自閉症スペクトラム障害
自閉症
腸内細菌叢
制限的・反復的行動と興味
食事
メタゲノム
便の一貫性
脳-腸-マイクロバイオーム軸
はじめに
腸内細菌叢と自閉症スペクトラム障害(ASD)-社会性やコミュニケーションの障害、制限的で反復的な行動、異常な感覚反応などを特徴とする神経発達症-との関係には大きな関心が寄せられている。この関心は、胃腸症状を併発する割合が高いこと(Chaidezら、2014年;Kohaneら、2012年;McElhanonら、2014年;Vargasonら、2019年)や、マイクロバイオームがASD関連の行動特性に因果的に寄与していることを示唆する動物実験(Buffingtonら、2016年;Hsiaoら、2013年;Sharonら、2019年)によってもたらされている。また、さまざまな消化器疾患(過敏性腸症候群、炎症性腸疾患など)および精神神経疾患(うつ病など)に脳-腸-マイクロバイオーム軸が関与していることを示唆する証拠も増えている(Valles-Colomerら、2019)。
このような強い関心と、ASDにおける糞便微生物叢移植研究が第I相臨床試験まで進んでいる最近の状況(Kang et al., 2019; Kang et al., 2017)にもかかわらず、ASDに対するマイクロバイオームの寄与は説得力に欠ける。既存のヒトASDマイクロバイオーム研究では一貫性のない結果が得られているが、これはおそらく、バイアス(Rothschild et al、 2013; Parracho et al., 2005; Williams et al., 2011)、組成を意識した統計解析の一貫性のない使用(De Angelis et al., 2013; Finegold et al., 2010; Williams et al., 2011)、不均一な処理を表す可能性のある大きな部位効果(Fouquier et al., 2021)などがある。実際、参加者が100人を超える大規模研究の多くでは、ASDとマイクロバイオームとの関連は見つかっていない(Gondalia et al.) さらに、少数の例外(Berding and Donovan, 2018; Son et al., 2015)を除き、ほとんどのASDマイクロバイオーム研究は、性別を含む重要なマイクロバイオーム交絡因子(Bokulich et al., 2016; Falony et al., 2016; Rothschild et al., 2018; Vandeputte et al、 2011年;Wangら、2011年)、年齢(Adamsら、2011年)、食事(Adamsら、2011年;De Angelisら、2013年;Finegoldら、2010年;Stratiら、2017年;Wangら、2011年)、Bristol Stool Chart(rBSC)を用いて測定した便の一貫性、または併発する胃腸の愁訴および薬物(Wangら、2011年)を含む。特筆すべきは、ASDは男性に偏りがあり、自閉症スペクトラムの子どもは、(選択的摂食やアレルギーの懸念から)多様性に乏しい食事(Panossian et al., 2020; Schreck and Williams, 2006)や胃腸疾患を併発していることが多く(Chaidez et al., 2014; Kohane et al., 2012; McElhanon et al., 2014)、抗生物質を処方されやすい(Niehus and Lord, 2006)ことである。したがって、ASDにおけるマイクロバイオームとの関連についての既存の報告が、交絡変数によって説明されるかどうかは不明である。また、技術的な限界もある。ほとんどの研究は16S rRNAシーケンス(Ho et al., 2020)を用いており(一部を除く[Dan et al., 2020; Wan et al., 2021; Wang et al., 2019])、分類学的解像度が低く、マイクロバイオームに関する機能的情報が限られている。
これらの研究の限界の中で、最近のメタアナリシスでは、ASD診断状態との関連について一貫したエビデンスを持つ少数の分類群が報告された。すなわち、プレボテラ属、ファーミキューテス門、クロストリジウム目のクラスター、ビフィドバクテリウムの種である(Ho et al.) それにもかかわらず、我々の知る限り、マイクロバイオームとASD診断の全体的な関係は定量化されていない。実際、ASDのマイクロバイオームに対する関心は、主要なエビデンスベースを上回っているようである。2021年4月にPubMedで検索([自閉症[タイトル] AND [マイクロバイオーム[タイトル] OR [マイクロバイオータ[タイトル]])したところ、56の総説が同定されたのに対し、最新のメタアナリシス(Ho et al.) より広範には、ヒトの微生物叢に関連した動物実験のメタアナリシスは、肯定的な知見の範囲が非常に広く、因果推論が時期尚早であり、過度に誇張され、出版バイアスに苦しんでいることを示唆する、あり得ないという懸念を提起している(Walterら、2020)。
ここでは、オーストラリア自閉症バイオバンク(AAB)(Alvares et al、 2018)とQueensland Twin Adolescent Brain(QTAB)プロジェクトで、広範な表現型データが利用可能であった(図1Aおよび1B;表1):人口統計学、Australian Eating Surveyからの食事データ(ここから食事多様性とエネルギー百分率測定からの主成分を導き出し、以下、食事PC1-3と呼ぶ)(STAR Methods;図1B参照)、便の一貫性、詳細な心理測定テスト、およびゲノムワイド一塩基多型(SNP)遺伝子型。その結果、便中マイクロバイオームがASD診断に及ぼす影響はごくわずかであること、一方、食事形質、便の一貫性、年齢には大きな有意な関連が認められた。その代わりに、自閉症スペクトラムの遺伝的・表現型的指標(制限的行動や反復的行動、社会的感情、より高い感覚過敏など)が、多様性の乏しい食事を促進し、それがマイクロバイオームの多様性を低下させ、より緩い便の粘性と関連するという代替モデルを示唆している。
図サムネイルgr1
図1入力データセットと解析の概要
キャプション
大きな画像を見るダウンロード 高解像度画像
表1メタゲノム研究参加者の人口統計学的概要
表現型 AAB+QTABマイクロバイオームコホート UNRサブグループ AAB全体
AAB+QTABマイクロバイオームコホート UNRサブグループ
N 99 51 97a 48 49 1168 262 149
年齢 8.7 (3.8) 8.0 (4.3) 9.2 (3.7) 6.3 (3.0) 12.1 (1.0) 7.7 (3.9) 8.2 (4.2) 6.5 (3.4)
男性 73% 57% 54% 50% 59% 60% 40% 39
BSC 3.56 (1.31) 3.84 (1.25) 3.37 (0.96) 3.35 (1.08) 3.39 (0.83) 3.51 (1.02) 3.56 (0.97) 3.46 (0.92)
rBSC 3.48 (0.96) 3.71 (0.94) 3.40 (0.84) 3.39 (0.91) 3.41 (0.78) 3.58 (1.42) 3.64 (1.23) 3.44 (1.18)
食事性PC1 -0.15 (1.85) 0.13 (1.33) 0.09 (0.93) 0.07 (1.01) 0.10 (0.87) - - - - - - - - - - - - - - - 0.20 (1.85) 0.13 (1.33) 0.09 (0.93) 0.07 (1.01) 0.10 (0.87)
分類学的シャノン指数 3.63 (0.44) 3.59 (0.45) 3.76 (0.38) 3.59 (0.40) 3.92 (0.28)
ゲノムワイドSNPデータ N 91 43 84 48 49 887 218 116
ASD PGS 0.18 (1.10) 0.22 (0.95) 0.028 (0.72) 0.06 (1.00) -1e-6 (1.9e-7) 0.060 (1.00) -0.085 (0.96) 0.033 (1.07)
ASD-ADHD-TS PGS 0.10 (1.10) 0.17 (0.82) 0.04 (0.61) 0.09 (0.87) -1.1e-6 (1.6e-7) 0.047 (0.98) -0.079 (0.90) 0.012 (0.88)
神経質 PGS -0.038 (1.00) -0.086 (0.94) 0.041 (0.68) 0.084 (0.98) -4.1e-5 (4.2e-5) 0.040 (1.00) 0.025 (0.95) 0.145 (0.93)
IQ-DQ複合スコア 85 (24) 100 (17) 100 (15) 100 (15) 100 (15) 79 (24) 101 (13) 103 (13)
CSHQ raw score 43 (11) 37 (6.1) 38 (7.1) 38 (7.1) - 44 (10) 39 (9) 37 (8)
ADOS2/G 比較スコア 6.7 (2.1) - - - - 6.7 (2.0) - - - ADOS2/G RRB Cスコア
ADOS2/G RRB CSSスコア 7.0 (2.3) - - - - 6.8 (2.2) - - - ADOS2/G 社会的影響 CSSスコア
ADOS2/G 社会的影響 CSSスコア 6.8 (2.0) - - - - 6.8 (2.0) - - - - ADOS2/G 社会的影響
SSP 感覚生スコア 41 (12) - - - - 33 (8) - - - - - SRS Tスコア 74 (9)
SRS Tスコア 74 (9.3) 46 (9.1) 51 (9.3) 51 (9.3) - 78 (10) 50 (12) 50 (9)
最初の3列は、本分析で使用したASD/SIB/UNRグループ間の比較を示す。4列目と5列目は、UNRグループをAABとQTABコホートに分けたものである。6~8列目は、より広範なAABコホートの特徴を示している。BSC:Bristol Stool Chart(スケール1-7)、rBSC:再グループ化されたBSC(BSCクラスは1+2、3、4、5+6+7にグループ化)、食事PC:clr変換されたパーセントエネルギー食事データから得られた主成分、ASD PGS: ASDの多遺伝子スコア、ASD-ADHD-TS PGS:ASD-ADHD-TS GWAS要約統計からの多遺伝子スコア、MSEL非言語スコアから集計したIQ-DQ複合スコア、WISC-IV複合スコア、NIH Toolbox(年齢補正IQ)、CSHQ: CSHQ:Children's Sleep Habits Questionnaire raw score;ADOS2/G比較スコア:Autism Diagnostic Observation Schedule version 2(ADOS-2)および-Generic version(ADOS-G)比較スコアの複合スコア;ADOS2/G RRBおよび社会的影響CSSスコア: ADOS-2およびADOS-GのRRBおよび社会的影響領域の較正重症度スコア;SRS Tスコア: SRS Tスコア:Social Responsiveness Tスコア;SSPセンサー生スコア:Short Sensory Profileの「感覚」領域のスコア。
新しいタブで表を開く
結果
研究の特徴
この研究には、ASDと診断された99人(「ASD」)、診断のない51人のペアのきょうだい児(「SIB」)、診断のない97人の無関係な(お互いを含む)未診断児(「UNR」)を含む、合計247人の子ども(2~17歳)が参加した(図1A)。ASD群とSIB群の参加者はAABからのみであったが、UNR群はAABから48人、QTABから49人であった。UNRの2人の参加者は食事データが不完全であり、食事変数が分析されなかった場合に含まれた。AABとQTABの便サンプルは、同一のプロトコルで同時期に採取され、同一のヒトサンプル処理専門家によりレセプトされ処理された。UNR_AAB参加者(平均年齢6.3歳、SD=3.0)はUNR_QTAB参加者(平均年齢12.1歳、SD=1.0)より平均的に若かったが、これらのサブグループを組み合わせると、3つの研究グループ(ASD、SIB、UNR)は年齢で一致した(ANOVA p=0.2)(STAR Methods; Document S1)。ASD研究は一般的に男性に偏りがあり、年齢と性別によるグループのマッチングを試みた結果、最終的にグループごとの男性の割合はASD 73%、SIB 57%、UNR 54%であった。本研究に便サンプルを提供したAAB参加者は、概してAAB全体を代表していた(表1)。ASD群は平均して知能指数(IQ)-発達指数(DQ)複合スコア(IQ-DQ)が低かった(ASD平均=85、SD=24;SIB平均=100、SD=17;UNR平均=100、SD=15)。ASD特性の程度の尺度として、ASD群の自閉症診断観察スケジュール-2/G(ADOS2/G)比較スコアの平均は6.7(SD=2.1)であった。完全な人口統計学的詳細と比較は表1に、参加者の募集とQTAB参加者の更なる詳細はSTAR Methodsに、本研究で用いた表現型データの概要は図1Cに示されている。
ASD診断状態におけるマイクロバイオームとの関連は、年齢、便、食事形質と比較してごくわずかであった。
まず、ASD診断と、神経発達形質(IQ-DQ、Children's Sleep Habits Questionnaire [CHSQ] raw score)、マイクロバイオームと直感的な関係を持つ表現型(食事PCs [図1B]、食事の多様性、便の一貫性;STAR Methods参照)、CD4+ T細胞の割合を含む他の様々な形質(図1C)について、共変量(図2の凡例に記載)で調整した上で、マイクロバイオームと関連する表現型の分散の割合を推定した。この分散推定値-マイクロバイオーム関連指数(または "b2")(Rothschild et al. h2が(直接的または間接的な)因果関係を反映するのに対し、b2は形質変異の原因または結果を反映する。この解析では、個体の各ペア間のマイクロバイオームの特徴(種レベルと微生物遺伝子の様々な階層に焦点を当てた分類群)から「オミックス関連性行列」(ORM)を計算し、線形混合モデルの枠組みで形質に対して回帰する(Zhang et al.)
図サムネイルgr2
図2マイクロバイオーム組成に関連する表現型分散の割合(±SE)(b2)
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
微生物種と微生物遺伝子(Microba Genes [MGENES]データベースのオープンリーディングフレーム、本節では「微生物遺伝子」と呼ぶ;Enzyme Commission classification [この階層のレベル4に着目] [Bairoch, 2000]; Transporter Classification Database [TCDB] [Saier et al、 MetaCyc pathways [Caspi et al., 2020])に分類し、一般的な特徴と稀な特徴に層別化した(表S1;各特徴の中央値カウント>0に基づく閾値)。データの分布を調べ(図S1)、中心対数比(clr)変換が適切であることを確認した(図S2)。
サムネイル図1
図S1様々なマイクロバイオームデータセットのORM対角要素と非対角要素の関係
キャプション
大きな画像を見るダウンロード 高解像度画像
サムネイル図2
図S2OREML推定値における中心対数比変換と2値化0/1符号化の効果
キャプションを表示
View Large ImageDownload 高解像度画像
サムネイル図3
図S3分散成分分析の追加結果
フルキャプションを表示
図を拡大する 高解像度画像
まず、年齢とBMIをベンチマーク形質として分析した。我々の結果(子供の場合)は、それぞれ成人の28%と11%という以前の種レベルのb2推定値と一致していた(Rothschild et al、 2020):共通種(n = 96)のb2推定値は、年齢で35%(SE = 7%、p∼0、偽発見率[FDR]-有意、共変量:性)(図2;表S1)、BMIでb2 = 12%(共変量:年齢、性、SE = 7%、p = 1.8e-2、FDR-有意)であった。注目すべきは、遺伝子レベルのORMは、年齢(b2>99%、SE=14%-17%、p∼0、中央値が0を超えるn=251,617の共通遺伝子または中央値が0を超えるn=1,742,727の希少遺伝子のいずれかに基づく)とBMI(希少遺伝子b2=47%、SE=22%、p=1. 1e-2、FDR有意)、分類学的および機能的マイクロバイオーム測定が表現型分散の異なる要素を捉えていることを示唆している。
年齢に関する結果とは対照的に、ASD診断に関する種および遺伝子レベルのb2推定値は弱く、有意ではなかった(最大:希少遺伝子b2 = 9%, SE = 16%, p = 0.29;共変量:年齢、性別、食事PC1-3)。 20;共変量:年齢、性)、CSHQ生スコアで測定した睡眠問題(共通種b2=11%、SE=9%、p=8.2e-2;共変量:年齢、性)(図2;表S1)、clr変換CD4+T細胞割合(b2∼ 0、SE=0.06、p=0.50;共変量:年齢、性、食事PC1-3)(定量化および統計解析)であった。
これらの神経発達形質や免疫形質とは異なり、便の一貫性(共変量:年齢、性別、群、食事PC1-3;希少種b2=40%、SE=11%、p=2. 8e-5; 希少遺伝子 b2 = 64%, SE = 21%, p = 5.8e-5)および食事PC1(共通遺伝子: b2 = 47%, SE = 15%, p = 4.5e-5; 共変量: 年齢、性別、参加者グループ)(図2; 表S1; 定量化および統計解析)。
感度分析として、関連するマイクロバイオーム指標がb2推定に与える相対的な影響を調べるため、複数のORMのフィッティングを検討した。(1)分類学的データセットと機能的データセットから構築したORMの組み合わせ、(2)特徴の共通サブセットとレアサブセット、(3)複数の階層(例えば、種、属、科のデータ)のフィッティングは、すべてb2推定値を増加させた(定量化と統計解析;図S3;表S1)。
全体的に、我々の結果は、共変量なし(図2、開いた円)、より小さい年齢範囲、MetaPhlAn2(Truongら、2015)分類学的プロファイリングパイプラインの使用、現在抗生物質を使用している者の除外、および兄弟姉妹の除外という複数の感度分析に対して頑健であった(方法S1)。また、マイクロバイオームの予測能力を推定する非加算モデルについても検討し、やはりマイクロバイオームがASD診断ではなく、年齢を予測することがわかった(定量化と統計解析)。
ロンブーツィア・ティモネンシス(Romboutsia timonensis)の関与が示唆された、差次的に豊富な分類群と遺伝子
次に、ASD診断の微生物マーカーを探し、607種、297属、38目、15門の存在量の差を調べた。我々は、多重検定を考慮し、偽陽性率を適切に制御するロバストなノンパラメトリック法であるAnalysis of Composition of Microbiomes (ANCOM)v2.1 (Mandal et al., 2015) (Weiss et al., 2017) を使用した(STAR Methods)。ASD群とSIB群とUNR群の組み合わせ(共変量:年齢、性別、食事PC1-3)を比較すると、Romboutsia timonensisという種のみが従来の検出閾値>0.7で有意に存在量に差があった(ASD群で存在量が少ない)(図3A;表S2)。この結果は広範な感度分析(図3BおよびS4A-S4I;Methods S1)でも一貫しており、減少したErysipelatoclostridium sp003024675は、多くの場合、次に差次的存在量の多い分類群であった(表S2)。差分存在検定(フィッシャーの正確検定、ASD対UNR)でも、同じ2つの分類群(R. timonensis p = 3.9e-4, 56/99 ASD対78/97 UNR; E. sp003024675 p = 1.5e-4, 6/99 ASD対25/97 UNR)が上位にランクされたが、どちらもFDR補正には耐えなかった(表S2)。並べ替え検定(各サンプルの診断ラベルのn = 1000ランダムシャッフル)では、これらの分類群はANCOM検定とフィッシャーの正確検定(表S2)の両方で経験分布と比較した場合、有意に存在量に差があり(p ≤ 0.001)、これらの知見が頑健であることを示すさらなる証拠が加わった。
図サムネイルgr3
図3ANCOMv2.1を用いたアバンダンスの差分検定
キャプション
大きな画像を見るダウンロード 高解像度画像
図サムネイルfigs4
図S4アバンダンスの感度分析
フルキャプションを表示
画像を拡大する 高解像度画像
特筆すべきことに、ASDと腸内細菌叢との関連について、以前に報告されたPrevotella属、ファーミキューテス門、Clostridialesクラスター、およびビフィドバクテリウムの種(Ho et al. しかし、R. timonensis(ペプトストレプトコッカス科、Clostridiales目、Clostridia綱、Firmicutes A門)とE. sp003024675(Erysipelatoclostridiaceae科、Erysipelotrichales目、Bacilli綱、Firmicutes門)はこれらの系統群のメンバーであることに注意されたい。再現性の低さは、(1)先行するマイクロバイオーム研究が力不足でサンプリングバイアスに陥りやすいこと(Rothschild et al、 2016年);そして(3)研究間で統計手法が異なり、多重検定や交絡因子に対する適切な調整の使い方にばらつきがある。
ASDマイクロバイオームの文献を支配してきた)16S配列決定研究とは異なり、メタゲノミクス配列決定では機能的洞察が可能である。そこで、ASD診断に関連する微生物遺伝子を探した。MetaCycグループ、MetaCycパスウェイ、Enzyme Commission(EC)遺伝子ファミリーに対するメタゲノム注釈の相対的な存在量に着目し、(計算および多重検定の負担のため、全遺伝子セットとは対照的に)解像度の高い順に調べたところ、有意な関連は認められなかった(共変量:年齢、性別、食事PC1-3、図S4J-S4L)(表S2)。
次に、ASD関連シグナルの根底にあるR. timonensis由来の特定の遺伝子または経路を同定しようとした。我々は、MetaCycグループ、MetaCycパスウェイ、EC遺伝子ファミリー、および我々のデータセット(共変量:年齢、性別、食事PC1-3)中のR. timonensisに直接マッピングされた特定の遺伝子(サンプル全体で10個以上の非ゼロ値を持つn = 4,950遺伝子)について、ASD対SIB+UNRの存在量の差を検定した(図S4M-S4O)。検出閾値が0.7を超える6つの発現量の異なる遺伝子を同定し(図3Cおよび3D;表S2)、そのうちの1つはEC遺伝子セットと重なったが、MetaCycグループやパスウェイとの関連は見られなかった(図S4M-S4O;表S2)。種レベルでの効果の方向性と一致して、有意に存在量の異なる遺伝子はすべて、ASDグループでclr変換された存在量が減少していた(図3D)。これらの遺伝子の機能には、アミノ酸(L-グルタミン、L-リジン、L-メチオニン、L-スレオニン)、プリンおよびピリミジン、炭水化物(ガラクトース)、ならびに細菌の胞子発芽およびdsDNA消化の代謝が含まれた(定量および統計解析;表S2)。これらの結果は微生物遺伝子の潜在的な転写を表しており、実際の発現を評価するにはメタトランスクリプトミクスのデータが必要であることに留意されたい。
IQ-DQ複合スコアについて種レベルの存在量差分析を行ったところ、ASD対SIB+UNRの比較ではBifidobacterium sp002742445が検出閾値>0.7を下回ったが、ASD対UNRの感度分析では検出閾値>0.6を下回ったのみであった(図3Eおよび3F;表S2;定量および統計解析)。ASD対SIB+UNR解析(n = 200分類群)では、ビロームの関連は見られなかった(表S2)。
食生活の多様性がASDとマイクロバイオームの関連を媒介する
メタゲノミクスデータではASDに関連するシグナルは少なかったが、食事との関連は一貫していた:分散成分分析では、食事のORMはASDと強く関連しており(R2 = 14%、SE = 7%、p = 2.2e-5、FDR-有意)(図2)、年齢と性別で調整した後、ASDではSIBおよびUNRグループと比較して食事のPC3が有意に低い(図1Bに示すように、肉摂取量の減少を示唆)ことが観察された(図S5)。スペクトラムの子どもは多様性の乏しい食事を好むことから(Panossianら、2020;Schreck and Williams、2006)、食事の多様性がマイクロバイオームおよび胃腸症状(便の一貫性を介して)に及ぼす影響を検討した。
サムネイル図5
図S5参加者群と食事および便の変数との関係
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
ASDグループは、123の食品レベル変数(STAR Methodsを参照)から食事のα多様性を測定するためにシャノン指数を用いて推定された、SIBグループとUNRグループ(一元配置分散分析p = 1.3e-7、FDR-有意)よりも、年齢と性別(一元配置分散分析p = 1.5e-6、FDR-有意)およびエネルギー摂取量(分散分析p = 9.0e-7、FDR-有意)で調整した場合を含め、有意に多様性の低い食事をしていた(図4A;表S3)。また、検証された小児および青年用のオーストラリア推奨食品スコア(ARFS)を用いて測定された食事の質も低かった(ANOVA p = 7.9e-4、FDR-有意)(図S6;表S3;方法S1)(Burrowsら、2014;Marshallら、2012)。これらの頑健な食事結果は、ASD診断と分類学的α多様性との間に、測定法や共変量の使用(年齢、性別、食事PC1-3)にかかわらず、あるいはβ多様性(同じ共変量を用いた重み付けUnifrac並べ替え多変量分散分析[PERMANOVA] p = 0.20、PERMDISP2 p = 0.85)にかかわらず、ほとんど直接的な関連がないことと対照的であった(図5;定量化および統計解析)。
図サムネイルgr4
図4食餌および分類学的多様性(シャノン指数を用いて測定)とASD関連表現型との関係
キャプション
大きな画像を見るダウンロード 高解像度画像
図サムネイルfigs6
図S6食事の質および分類学的多様性とASD関連表現型との関係
完全なキャプションを表示
画像を拡大する 高解像度画像
図サムネイルgr5
図5マイクロバイオームの多様性解析
キャプション
画像を拡大する 高解像度画像
ASDと食事の多様性との関連性を追求するため、分類学的多様性はASD診断に直接関連するというよりも、むしろ食事の下流の結果であるかもしれないという仮説を立てた。この仮説と一致して、食事と分類学的多様性の間には有意な正の相関がみられた(Pearson r = 0.25, p = 6. 3e-5、FDR-有意)(図4C;表S3)、相互回帰分析では、食事と分類学的多様性(共変量:年齢、性別、便の一貫性、群)は互いの有意な予測因子であった(それぞれ、b = 0.02、p = 3.7e-2、b = 1.1、p = 3.7e-2)(共変量を追加した図4D、4E、S7A、S7B)。さらに、食事多様性回帰における最大の効果は群によるものであり(UNR:b = 0.035、p = 3.0e-6、SIB:b = 0.021、p = 1.4e-2)、分類学的多様性は群とは関連していなかった(図4Bおよび4E)。このことは、ASDに伴う食事制限(診断そのものではない)がマイクロバイオームの多様性の低下と関連していることを示唆している。これらの効果は、エネルギー摂取量(kJ)を追加の共変量として当てはめた感度分析(図S7)、および食事の多様性をARFSを用いて測定した食事の質に置き換えた場合(図S6;表S3;方法S1)にも頑健であった。
サムネイル図7
図S7共変量を追加した場合の食餌多様性と分類学的多様性の関係
キャプション
大きな画像を見るダウンロード 高解像度画像
次に、食餌多様性、分類学的多様性と便の一貫性の関係を調べた。我々は、便の一貫性と分類学的多様性の間の逆相関(共変量なしでb = -0.41 p = 3.7e-3、FDR-有意)(表S3)を以前に報告された(Hadizadehら、2017;Vandeputteら、2016;Zhernakovaら、2016)を再現し、共変量に頑健であった(図S8C;表S3)。また、便の一貫性と食事の多様性(共変量なしのモデルでb = -2.34、p = 3.7e-2)との名目上有意な関連も確認されたが、これは共変量調整には耐えられなかった(図S8B;表S3)。注目すべきことに、分類学的多様性モデル(共変量なし)は、食餌多様性分析(R2 = 1.5%)よりも便の一貫性においてより大きな分散(R2 = 3.2%)を説明し、分類学的多様性と食餌多様性の両方を説明変数として当てはめた便の一貫性のモデルでは、共変量を含めると、分類学的多様性のみが有意であった(b = -0.36、p = 1.5e-2)(図S8A)(図4F)。全体として、このことは、便の硬さが緩い(rBSCスコアが高い)ことが分類学的多様性の減少と近接的に関連しており、これは食餌多様性の減少の下流にあることを示唆している。このメカニズムにより、胃腸の問題の増加と反復行動の増加との間に報告されている関係が説明できるかもしれない(Chakrabortyら、2020年)。
サムネイル図8
図S8便の一貫性と食事および分類学的多様性との関係(共変数を追加した場合
キャプション
大きな画像を見るダウンロード 高解像度画像
行動と嗜好は食餌と分類学的多様性の減少の上流にある
ASDと診断される行動因子が、食事制限や食事・分類学的多様性の減少の上流にあるかどうかを調べた。後者はAAB(Yapら、2021)とQTAB参加者の血液サンプルから作成したゲノムワイドSNPデータを用いた(STAR Methods参照)。前節では微生物遺伝子の寄与について述べたが、本節ではヒトの多遺伝子スコアと食事および分類学的多様性との関係を探る。
まず、ASDと食事の多様性の関連を、自閉症スペクトラムの連続的な測定値の解析を通して確認した。このコホート(Yapら、2021)におけるASD多遺伝子スコアと食事多様性(b = -1.0e2、p = 1.2e-2、FDR-有意)との間には逆相関が確認されたが、分類学的多様性(b = -4.4e-2、p = 0.17)には認められなかった(図4G)。ASD多遺伝子スコア自体は、このデータセットにおけるASD診断を予測するには検出力が不十分であった(one-way ANOVA p = 0.41。) 表現型的には、食事の多様性とASDの特徴の程度を示す2つの定量的指標との間に負の関連が観察された: ADOS2/G比較スコア(b = -8.6e-3、p = 3.1e-3、FDR有意、n = 99 ASD)(図6Aおよび6B)およびSocial Responsiveness Scale t-スコア(b = -6.4e-4、p = 7.8e-2、わずかに有意、n = 97 AAB児:10ASDおよび87 SIB/UNR)(図6Cおよび6D)。
図のサムネイルgr6
図6他の食事および分類学的多様性測定と他の表現型との関係
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
第二に、反復的で制限的な行動や興味(ASDの2つの診断領域のうちの1つ)が、マイクロバイオームの変化の上流で、制限的な食事の背景にある可能性があるという仮説を立てた。表現型的には、より高い複合(Lord et al、 2012)ADOS-2/G制限行動・反復行動(RRB)キャリブレーション重症度スコアの高さと食事の多様性(共変量なし:b = -7.8e-3、p = 3.8e-3;共変量年齢・性別あり:b = -6.4e-3、p = 1. 8e-2;いずれもFDRで有意)(表S3)、分類学的多様性(共変量なし:b = -4.3e-2、p = 2.4e-2;共変量年齢と性別あり:b = -3.4e-2、p = 7.6e-2)(n = 99 ASD群のみ)とは、名目上有意な負の相関があった(図4H;表S3)。次に、ASD、注意欠陥多動性障害、トゥレット症候群(以下ASD-ADHD-TSと呼ぶ)の交差形質解析(Yangら、2021)から得られたゲノムワイド関連研究(GWAS)の要約統計量を活用し、制限的反復行動の多遺伝子スコア(PGS)を作成した。ASD-ADHD-TSのPGSは、ADOS-2/GのRRBスコアと相関しており(r = 0.09, p = 0.01, n = 867)、遺伝的な代用因子であることが確認された。ASD-ADHD-TS PGSと食事多様性の減少(b = -7.2e-3、p = 0.10)にはわずかな関連が認められたが、分類学的多様性(b = -4.7e-2、p = 0.18)には関連が認められなかった(図6Eと6F)。これらの結果をASD診断のもう一つの主要領域であるADOS2/G社会的影響度スコアと比較したところ、ADOS2/G RRBスコアと社会的影響度スコアは有意に相関していた(r = 0.29, p = 3.7e-3, n = 99参加者)。ADOS2/Gの社会的影響と食事の多様性の間にFDR有意な関連があることを再び発見した(共変量なし:b=-8.2e-3、p=1.0e-2;共変量年齢および性別あり:b=-7.5e-3、p=1. 5e-2 いずれもFDR有意)(図6G;表S3)、分類学的多様性(共変量なし:b=-3.3e-2、p=0.15;共変量年齢・性別あり:b=-2.9e-2、p=0.19)とは関連がなかった(図6H;表S3)。重要なことは、これらの関連はADOS2/G RRBスコアよりも弱かったことである。したがって、ASDに関連する制限行動や反復行動は、社会的情動よりも食事の多様性と強い関係があるようである。
第三に、制限された食事嗜好の根底には感覚過敏も存在する可能性がある(Cermak et al. その結果、食事(b = -9.5e-4、p = 6.9e-2)と分類学的多様性(b = -6.8e-2、p = 8.6e-2)の両方にわずかな関連が認められた(n = 91;図4I)。注目すべきことに、このデータセットでは、Short Sensory Profileの生の感覚スコアはADOS2/G RRB Calibrated Severity Scoresと相関がなかった(r = 0.05, p = 0.64)。
対照的に、食事および分類学的多様性の測定値と、不安の代理としての神経質さ(Kim et al., 2018; Yang et al., 2019)などのASD関連形質との間の仮説的関連性の証拠は見つからなかった(図6Kおよび6L;表S3;定量化および統計解析)。
全体として、これらのデータは、ASDに関連する嗜好や行動が、ASDとマイクロバイオームの弱い関係を媒介する食事の多様性の減少につながることを示唆している(図4J)。注目すべきは、すべての心理測定値が、分類学的多様性よりも食事の多様性と有意な相関を示したことである(図4G-4Iおよび6A-6H)。しかし、このような下流のマイクロバイオーム効果もフィードバックして行動に影響を及ぼす可能性は否定できない。
考察
交絡因子を注意深く考慮した今回の大規模ASD便メタゲノム研究において、便マイクロバイオームとASD診断状態との直接的な関連を示すエビデンスはごくわずかであり、これは他の神経発達特性(IQ-DQ、睡眠問題など)でも同様であった。ASDについては、分類学的多様性やマイクロバイオーム関連指標(b2;図2)との関連を示す証拠は限られており、頑健に同定されたのは、有意に豊富な1種のみであった(図3)。これらの結果は、マイクロバイオーム組成と年齢、食事、便の一貫性との強い関連性(図2)と比較すると顕著であった。重要なことは、ヒトの研究で以前に報告されたASDとマイクロバイオームの関連を再現できなかったことである。その代わりに、自閉症スペクトラムに関連する行動(例えば、反復制限行動や興味、感覚嗜好、社会的影響)と食事の多様性の低下とを関連付ける証拠を見出した。この推定モデルは、マイクロバイオームがASD関連形質と因果関係があるかもしれないという動物実験からの示唆に挑戦するものである(Buffingtonら、2016;Hsiaoら、2013;Sharonら、2019)。我々の知見はまた、マイクロバイオームを標的とすることでASDを「治療」することを提案する実験的介入や初期の臨床試験の急増(Kang et al.)
マイクロバイオーム組成の測定とは対照的に、ASDは共変量にかかわらず、食事変数と頑健かつ有意に関連していた(表S3)。我々は、(1)ASD診断における有意な分散が、b2解析において、マイクロバイオームではなく、食事と関連していたこと(図2)、(2)ASD群における肉摂取量の減少(図S5)、および(3)食事の多様性において有意に高い分散にもかかわらず、ASD群における食事の多様性の減少(図4A)を見出したが、これは、食餌学の文献(Panossianら、2020;Schreck and Williams、2006)および食事データを用いたいくつかの小規模なASDマイクロバイオーム研究(Berding and Donovan、2018)と一致している。
ASDマイクロバイオームへの関心の根拠の1つは、消化器系の愁訴が頻繁に併発することである(Chaidezら、2014;Kohaneら、2012;McElhanonら、2014)。完全な消化器系愁訴の報告がないため、この単一時点のデータが慢性状態をどのように反映しているかは不明であるという注意を払った上で、便の一貫性スコアを分析した。便の一貫性は、食事の多様性よりも分類学的多様性に近接しているようであったが、トップダウン(すなわち、食事および分類学的多様性が下流の便の一貫性に影響を及ぼす)の関係か、ボトムアップ(すなわち、便の一貫性が上流の代用品である)の関係かを区別することは困難であることを認める。前者については、食事制限が腸内生態系と分類学的多様性に影響を与え、それが便の一貫性に影響を与えるというのがもっともらしい。ボトムアップモデルとの関連では、便が緩いということは、根本的な食物アレルギーまたは不耐性を示している可能性があり、これは原因物質を特定するための意図的な(親の)食事制限と関連している可能性がある。さらに、緩い便の粘性は、消化管通過時間の短縮と結腸水再吸収の減少を反映し(Vandeputte et al. 便の硬さに影響する消化管疾患(過敏性腸症候群など)の狭義の遺伝率は小さいため(Wuら、2021年)、遺伝よりも環境の寄与が優勢である可能性が高い(Rothschildら、2018年)。
今回の結果は、ASDやその他の精神科的形質における腸内細菌叢の役割を理解する上で重要な意味を持つ。第一に、医療との関連において、自閉症スペクトラム児の食物選択性は重要な臨床的関心事である。食物選択性は回避性/制限性食物摂取障害(ARFID;自閉症児の20%以上が罹患しているにもかかわらず、過小診断されている可能性が高い[Koomarら、2021])と関連しており、入院や経腸栄養などの侵襲的措置が必要とされるほど、自閉症児の栄養欠乏を引き起こす可能性がある(Zimmerら、2012)(Tangら、2011)。また、我々の結果は、スペクトラム児の食事の質がより低いことを示唆している(Methods S1)。微生物多様性の上昇が健康アウトカムの改善と強固に関連していることを考えると(Valdesら、2018)、ASDと食事の質の低下、食事および分類学的多様性の低下との関連は、この集団における食事および栄養介入の重要性を強調している。第二に、我々の結果は、精神疾患におけるマイクロバイオーム解析における食事との関連における因果関係の解釈に示唆を与えるものである。食事とマイクロバイオームが精神医学的形質(例えば、うつ病[Dash et al., 2015; Molendijk et al., 2018])に寄与することへの関心が高まっているが、我々の結果は、行動がマイクロバイオームに及ぼす(間違いなくより直感的な)影響を考慮する必要性を強調している(Jacka et al.) これらの結果は、食事がマイクロバイオームと健康との関連を促進するという他の報告(Claesson et al.)
今後のマイクロバイオーム研究においては、特にASDやその他の神経精神疾患形質で、食事と診断や治療との共変量がもっともらしいものについては、詳細な食事データを収集することの重要性を強調する(最近の例[Asnicarら、2021;Wangら、2021])。結果がサンプリング効果に対して頑健であることを保証し、より微妙なマイクロバイオームとの関連を同定するために、サンプルサイズを大きくすることを提唱する。また、より詳細な分類学的手法によるマイクロバイオーム組成の測定がより高感度であったこと(表S1)、遺伝子レベルのORMがいくつかの形質についてより多くの分散を説明したこと(表S1)、MetaPhlAn2/NCBIパイプラインでは関連性を検出する力が弱かったこと(方法S1)、分類学的データと機能的データセットがマイクロバイオームの相補的な側面を捉えている可能性があること(図S1およびS3)から、より高分解能のメタゲノミクス技術とデータベースの拡充を推奨する。
結論として、ASDと腸内細菌叢との直接的な関連はごくわずかであり、年齢、食事変数、便の一貫性などの他の表現型との強い関連とは対照的であった。その代わりに、制限的反復行動などのASD特有の特徴と関連する、食事の多様性の制限や質の低下が、分類学的多様性、ひいては便の一貫性の有意な媒介因子であるという証拠が得られた。この結果は、ASDに関連する行動や食事の嗜好が腸内細菌叢に及ぼす上流の役割と一致しており、微生物叢がASDにおいて主要な(あるいは因果的な)役割を担っているという主張とは相反するものである。
研究の限界
第一に、本研究は縦断的デザインではなかったため、ASD診断前のマイクロバイオームの寄与を除外することはできない。第二に、本研究は、ASD便マイクロバイオームに関するメタゲノム研究としては、我々の知る限りこれまでで最大規模であるが、それでもなお、克服するためにはより大規模な研究を必要とするサンプリングバイアスが存在する可能性がある(Rothschild et al.) 第三に、本研究では腸内マイクロバイオームの代理として便サンプルを用いたが、これはよりアクセス困難な粘膜マイクロバイオームを正確に表していない可能性がある(Shanahan et al.) 第4に、このコホートにおける抗生物質摂取に関するデータは体系的に収集されていなかったため、感度分析における除外以外に厳密に説明することができなかった。第5に、ゴールドスタンダードの豊度差解析は、生態学的または代謝的な文脈で生じる相互作用や非依存性を反映しない特徴ごとの検定に依存していた。最後に、これらの結果を再現するために、同等の研究デザイン、交絡因子の考慮、表現型とメタゲノミクスデータの深さを持つデータセットの出現を待ちたい。
STAR★方法
主要リソース表
試薬またはリソースソースの識別子
生物学的サンプル
便サンプル - Australian Autism Biobank Australian Autism Biobank https://www.autismcrc.com.au/biobank
血液 - Australian Autism Biobank オーストラリア自閉症バイオバンク https://www.autismcrc.com.au/biobank
便サンプル - Queensland Twin Adolescent Brain Project Queensland Twin Adolescent Brain Project https://espace.library.uq.edu.au/view/UQ:e803a68
血液 - Queensland Twin Adolescent Brain Project クイーンズランド双生児思春期脳プロジェクト https://espace.library.uq.edu.au/view/UQ:e803a68
重要な市販アッセイ
QIAamp 96 PowerFecal QIAcube HT キット QIAGEN https://www.qiagen.com/us/products/discovery-and-translational-research/dna-rna-purification/dna-purification/microbial-dna/qiaamp-96-powerfecal-qiacube-ht-kit/#orderinginformation
Nextera XT Library Preparation Kit (Illumina #FC -131-1096) イルミナ https://www.illumina.com/products/by-type/sequencing-kits/library-prep-kits/nextera-xt-dna.html
寄託データ
Australian Autism Biobankからの生データおよび解析データ 本論文は、Cooperative Research Centre for Living with Autism(Autism CRC)内のAustralian Autism Biobankへの申請により入手可能: https://www.autismcrc.com.au/biobank
Queensland Twin Adolescent Brain Project の生データおよび解析データ この論文は、仲介者のアクセスにより入手可能: UQ eSpace: https://espace.library.uq.edu.au/view/UQ:e803a68
ヒトリファレンスゲノム NCBI build 38, GRCh38 Genome Reference Consortium https://www.ncbi.nlm.nih.gov/projects/genome/assembly/grc/human/
ヒトリファレンスゲノム NCBI build 37, GRCh37 Genome Reference Consortium https://www.ncbi.nlm.nih.gov/projects/genome/assembly/grc/human/
Microba Genome Database (MGDB) v2.0.0 Microba Life Sciences N/A
Microba Genes (MGNES) v2.0.0 Microba Life Sciences N/A
UniRef90 release 2019/09 UniProt Consortium https://ftp.uniprot.org/pub/databases/uniprot/previous_releases/release-2019_09/uniref/
NCBI RefSeq Viral データベース Brister et al., 2015 https://www.ncbi.nlm.nih.gov/genome/viruses/
Autism GWAS summary statistics Grove et al., 2019 https://www.med.unc.edu/pgc/download-results/
クロス特性(ASD-ADHD-TS)要約統計 Yang et al., 2019 https://www.biorxiv.org/content/10.1101/770222v1
ソフトウェアとアルゴリズム
Burrows-Wheeler Aligner v0.7.17 Li and Durbin, 2009 http://bio-bwa.sourceforge.net/; RRID:SCR_010910
Trimmomatic v0.39 Usadella Lab http://www.usadellab.org/cms/?page=trimmomatic; RRID:SCR_011848
SAMtools v1.7 Samtools http://www.htslib.org/; RRID:SCR_002105
enrichM Boyd and Woodcroft, 2019 https://github.com/geronimp/enrichM
MMseqs2 Steinegger and Söding, 2017 https://github.com/soedinglab/MMseqs2
OSCA Zhang et al., 2019 https://cnsgenomics.com/software/osca/
ANCOMv2.1 Mandal et al., 2015 https://github.com/FrederickHuangLin/ANCOM
Microba Community Profiler v2.0.2 Microba Life Sciences N/A
Microba Gene and Pathway Profiler v0.1.0 Microba Life Sciences N/A
MiCoP LaPierre et al., 2019 https://github.com/smangul1/MiCoP
R v3.6.3 The R Project for Statistical Computing https://www.r-project.org/
ggplot2 Wickham, 2016 https://ggplot2.tidyverse.org/
ggstatsplot Patil, 2021 https://indrajeetpatil.github.io/ggstatsplot/
Plink 1.9 Purcell and Chang, 2015 https://www.cog-genomics.org/plink/
SBayesR Lloyd-Jones ら, 2019 https://cnsgenomics.com/software/gctb/#Overview
原稿に使用したサポートコード 本論文 https://zenodo.org/record/5558047
新しいタブで表を開く
リソースの有無
リードの連絡先
リソースおよび試薬に関する詳細情報およびリクエストは、リードコンタクトであるJacob Gratten (jacob.gratten@mater.uq.edu.au)までご連絡ください。
材料の入手可能性
本研究では新規のユニークな試薬は作成していない。
実験モデルと被験者の詳細
本研究はヒトを対象とした。本研究の247名の参加者は、Australian Autism Biobank (AAB) (Alvares et al., 2018)とQueensland Twin Adolescent Brain (QTAB)プロジェクトという2つのデータセットから得られた。AABの参加者にはASDと診断された子どもたちが含まれる(オーストラリアの4大都市全域の自閉症クリニックや研究センターから募集された): シドニー、メルボルン、ブリスベン、パース;除外基準なし;平均年齢=8.7、SD=3.8、73%男性)、および診断を受けていない兄弟(「SIB」)(平均年齢=8.0、SD=4.3、57%男性)である(Alvares et al.) 診断を受けていない無関係な子どものグループ(「UNR」)は、AAB(地域から募集;除外基準:ASD診断を受けていること)とQTAB(地域から募集した定型発達児)によって募集された(AAB+QTAB UNRグループ平均年齢=9.2、SD=3.7、54%男性)。AABとQTAB参加者の便サンプルは、同じプロトコルで同時期に採取され、Microba Life Sciences社によるDNA単離、ライブラリー調製、DNA塩基配列決定のためにランダム化される前に、同じクイーンズランド大学ヒト研究ユニットで受領・処理された。全体として、サンプルにはn=51の兄弟姉妹ペアが含まれ、n=99の症例とn=97の非血縁対照からなる症例対照デザインと重複していた(表1、図1a)。参加者グループの人口統計学的データの説明と比較は表1に示されている。このメタゲノミクス研究に含まれたAABサブセットは、より広いAABを代表するものであった(表1)。
本研究のサンプル選択は、便サンプルを提供し、食事アンケートに回答したAABおよびQTAB参加者に限定した。この制約の中で、性別と年齢によるマッチングを試みた。具体的には、UNR_QTAB参加者の性別(カイ二乗統計量=8.3、p=1.5e-2)と年齢(ANOVA p=2.0e-3)の格差を改善するために、UNR_QTAB参加者を選択した。 0e-3)、男女のバランスが等しく(50%)若い(平均年齢6.3歳、SD=3.0)UNR_AABサブセット(n=48人)と、男性に偏り(73%)年上(平均年齢8.7歳、SD=3.8)ASD(AAB)グループ(n=99人)との間の性差(カイ二乗統計量=8.3、p=1.5e-2)と年齢(ANOVA p=2.0e-3)を改善した。最終的なサンプルでは、ASDにおける高い男女比と一致するように、(減少したとはいえ)男性の偏りが持続していた(カイ二乗統計量=7.7、p=2.2e-2)が、グループ間の年齢に有意差はなかった(ANOVA p=0.2)(表1)。このため、すべての分析で性別を統計的に調整した。全体として、QTAB参加者を研究に含めることで、検出力が高まり、マイクロバイオームの重要な調節因子である年齢差(そしてより低い程度では性差)を改善し、収集バイアスの可能性を最小限に抑えることができた。
倫理承認と参加同意
本研究に参加することについて、全家族からインフォームドコンセントを得た。
NSW州:シドニー小児病院ネットワークHREC、承認番号HREC/14/SCHN/269。
QLD:Mater Health Services HREC、承認番号 HREC/14/MHS/212、クイーンズランド大学、承認番号 2014001079、QTAB Project: チルドレンズ・ヘルス・クイーンズランドHREC、承認番号HREC/16/QRCH/270、クイーンズランド大学、承認番号2016001784/ HREC/16/QRCH/270
VIC州:ラ・トローブ大学、承認番号HEC16/104
西オーストラリア州 プリンセス・マーガレット・ホスピタル・フォー・チルドレン承認番号2014029EP;西オーストラリア大学承認番号RA/4/1/8184
方法の詳細
表現型データ
食事データ
食事データは、AABおよびQTABの両コホートにおいて、主に両親から報告され、オーストラリア人集団で有効性が確認されているAustralian Eating Survey(AES;幼児および小児版)(Collins et al. 247人の参加者のうちn = 245人について食品レベルの摂取データが利用可能であり、n = 246人についてエネルギー百分率(pe)のデータが利用可能であった。AESには123種類の食品の摂取頻度が記録されており、そこから13の中核食品群(野菜、果物、肉類、代替蛋白質、穀類、乳製品)と非中核食品群(甘い飲み物、包装スナック菓子、菓子類、焼き製品、持ち帰り食品、調味料、脂肪の多い肉類)のそれぞれのpe、大栄養素(各種炭水化物、脂肪、蛋白質)、微量栄養素(各種ビタミン、ミネラル)、オーストラリア推奨食品スコア(ARFS)などの派生変数が生成される。
我々は主に2つの方法で食事データを使用した(図1c)。
1.食品レベルのインプットを用いて、マイクロバイオーム分析で用いたのと同じα多様性の尺度であるシャノン指数を用いて、食事の多様性を測定した(n = 245)。AESは、各食品の摂取頻度を順序変数で記録しており、正確な変数は食品によって異なる(例えば、果物摂取の場合:「全く食べない」、「週に1回」、「週に2~4回」、「週に5~6回」、「1日1回」、「1日2回以上」;一方、ハンバーガーの場合:「全く食べない」、「週に1回未満」、「週に2~4回」、「週に5~6回」、「1日1回」、「1日2回以上」): 一方、ハンバーガーについては、「食べたことがない」、「月に1回未満」、「月に1~3回」、「週に1回」、「週に2~4回」)。これらのN個のカテゴリーを、最も摂取頻度の低いカテゴリーを1、最も摂取頻度の高いカテゴリーをNとする整数の順序尺度で符号化し、この整数データセットからシャノン指数を用いて食事の多様性を算出した。分散成分分析では、整数データセットを入力してOREML分析用のORM(全個体ペア間の相関行列)を生成した。食事多様性の構成要素の妥当性をさらに検証するために、食事の質の検証された尺度であるAustralian Recommended Food Score for children and adolescents (Burrows et al., 2014; Marshall et al., 2012)も検証した(方法S1)。
他の研究者(Bokulich et al., 2016; Rothschild et al., 2018)によって強い関係が確認されていることから、マイクロバイオームに影響を及ぼす可能性のある顕著な食事の特徴を捉えるために、エネルギー割合データ(n = 246参加者;食事PCと呼ぶ;STAR Methods, Methods詳細)から主成分を算出した。
食事項目への負荷量(図1b)に基づいて、最初の3つの食事PCは以下のように解釈できる:
PC1:高い値は、植物性食品(野菜、果物、代替タンパク質)が多く、肉以外の非中核食品(甘い飲み物、パック入りのスナック菓子、菓子、焼き製品、持ち帰り、脂肪分の多い肉)が少ない食事と関連している。
PC2:高値は乳製品が多く、穀物や持ち帰りが少ない食事と関連する。
PC3:高値は、肉類(脂肪の多い肉類を含む)が多く、穀類と乳製品が少ない食事に関連する。
食事データの探索的データ分析を示すRMarkdown文書は、https://zenodo.org/record/5558047。
ブリストル便チャート
ブリストル便チャート(BSC)は、本試験のAABおよびQTAB参加者各自が記入した。BSCは便の硬さを1~7の数字で表したもので、数値が低いほど乾燥便(便秘など)、数値が高いほど水様便(下痢など)を示す。この分布の両極端は代表者が限られていたため、1+2および5+6+7のスコアをまとめて再グループ化した(図1c)。その結果、1グループあたり50サンプル以上の4段階評価となった。この再グループ化されたBSC変数(rBSC)を連続変数として扱った。
神経発達表現型
解析には複数の神経発達表現型を含めた。ASD関連の心理測定尺度には、ADOS-2またはADOS-G反復的・制限的興味サブスコアと社会的影響サブスコア((Lord et al., 2012)に従って組み合わせ、モジュール間の比較を可能にするためにドメインをCalibrated Severity Scoreに変換した(Hus et al., 2014; Hus and Lord, 2014))、Social Responsiveness Scale (Constantino, 2002)のtスコア(未診断AAB児)、およびShort Sensory Profile (McIntosh et al., 1999)の生のSensoryスコアが含まれた。非ASD関連特性のうち、複合IQ(AABの年長児ではWISC-IV(Wechsler, 2003)、QTAB研究ではNIH Toolbox年齢補正フルスケール得点(Akshoomoff et al. これらの質問票は、データセットのより大きな割合を捕捉し、知的発達遅滞の代理を提供するために集計された(以下、IQ-DQと呼ぶ)。検出力を向上させるためにこれらのスコアを組み合わせ、これらの測定値はすべて、母集団で予想される平均値=100、SD=15にほぼ一致するようにノルム化されている点で類似していた。また、AABサブセットにおいて、Children's Sleep Habits Questionnaire (CSHQ) (Owens et al., 2000)を用いて睡眠障害との関連を調べた。
ヒト遺伝子型解析による多遺伝子スコア(PGS)
我々は、ASD(Groveら、2019)、ASD-ADHD-TSの交差障害分析(Yangら、2021)、およびAABおよびQTABデータセットにおける神経質(Nagelら、2018)のPGSを、(Yapら、2021)に記載されたパイプラインを用いて計算した。簡単に言うと、HapMap3リファレンスにあり、両方のデータセットに共通するゲノムワイド関連研究のSNPをフィルターし、これらのフィルターされたSNP要約統計のすべてをSBayesR(Lloyd-Jones et al、 設定:-pi 0.95, 0.02, 0.02, 0.01; --gamma 0, 0.01, 0.1, 1; --chain-length 10000; ---burn-in 2000; --out-freq 10, --exclude-mhcフラグを使用)。次に、PLINK (Chang et al., 2015; Purcell and Chang, 2015) --score関数を用いて、対象サンプル(すなわち、AAB個体とUKB対照)における最良の推測遺伝子型に、再重み付けした効果量を掛け合わせた。
細胞タイプの割合
DNAメチル化データも作成した参加者のサブセットについて、細胞タイプ比率を計算した。このために、好中球、単球、B細胞、CD4+ T細胞、CD8+ T細胞、NK細胞、好酸球の細胞型比率を提供するデフォルトの血液参照(Reinius et al.
検体の収集と調製
AABデータセットとQTABデータセットの両方に同じプロトコルを適用した。
サンプル収集
Alvares et al., 2018)に記載されているように、ティースプーン大の便サンプルを、おむつから掻き取るか便器に吊り下げたライナーから、両親が自宅で採取し、4mLのRNAlaterに懸濁した。サンプルはクリニックに持参され、同日中にクイーンズランド大学分子生命科学研究所に発送された。ほとんどの場合、便サンプルは採取から2-3日以内(発送から1-2日)に届いた。処理には、強力なホモジナイズを行ってから分注し、-80℃で長期保存した。すべてのサンプルは凍結融解サイクルを1回のみ受け、それは配列決定時であった。
DNA抽出
DNA 抽出は、QIAamp 96 PowerFecal QIAcube HT キット(QIAGEN)の製造元の指示に従い、QIACube HT 自動抽出システムで行う前に、最初の機械的溶解ステップを最適化した修正プロトコルを用いて行った。処理に先立ち、サンプルを96ウェルプレートに移し、製造元の指示に従って氷冷PBSで2回洗浄した。
ライブラリー調製
Nextera XT Library Preparation Kit(Illumina #FC -131-1096)を使用し、384ウェルフォーマットで処理できるように反応量を減らして、メーカーのプロトコールに従ってライブラリーを調製した。ライブラリーはNexteraXT v2 384 Index A-D(Illumina FC-131-2001-4)を用いてインデックス付けした。得られたライブラリーを定量し、蛍光定量およびゲル解析を含む適切なQCを評価した。
ライブラリーのプール、QC、ロード、シーケンス
Nextera XTライブラリーを等モル量でプールし、シーケンスプールを作成した。プールを定量し、ゲル分析と蛍光定量でプールQCを行った。このライブラリーを製造元の指示に従ってIllumina NovaSeq6000でシーケンス用に調製し、Microbaラボで2×150bpペアエンドケミストリーでシーケンスした。プールはサンプルあたり3Gbの標的深度まで、最低2GB(約7M~16Mペアエンドリード)でシーケンスした。
定量と統計解析
メタゲノミクスデータセットとQC
品質管理
メタゲノムシーケンスデータのQCと処理はMicroba Life Sciences Limitedが行った。ペアエンドDNAシーケンスデータは、Illumina BaseSpace Bcl2fastq2 (v2.20)を用いて、インデックス配列の1ミスマッチを許容しながら、デマルチプレックスおよびアダプタートリミングを行った。その後、ソフトウェアTrimomatic(v0.39)を用いて、以下のパラメータでリードをクオリティトリミングし、残存アダプターを除去した: -phred33 LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 CROP:100000 HEADCROP:0 MINLEN:100. ヒトDNAは、Burrows-Wheeler Aligner (BWA) v0.7.17 (Li and Durbin, 2009)を用い、最小シードを-k 31に設定した以外はデフォルトのパラメータで、ヒトゲノムリファレンスアセンブリ38 (GRCh38.p12, GCF_000001405)にリードをアライメントして同定・除去した。アラインメントはさらにSAMtools v1.7 (Li and Durbin, 2009)を用い、-ubh -f1 -F2304フラグでフィルターした。少なくとも1つのリードが、リード長の90%以上で95%以上の同一性でヒトゲノムにマップされた残りのペアは、ヒトDNAとしてフラグを立て、削除した。表S4.1および表S4.2にQCプロセスの要約統計量を示す。1サンプルあたり480万~2480万リードがQCに合格した(表S4.2)。その後、全サンプルを700万リードペアの標準深度にダウンサンプリングし、関連解析における感度上昇の影響を緩和した。700万リード未満のn = 19サンプルについては、これらのサンプルを除外するのではなく、レアファクションの必要性について現在進行中の議論(McMurdie and Holmes, 2014)に注意しながら、すべてのQC済みリードを保持した。
微生物量の定量
Microba Genome Database (MGDB) v2.0.0を参照データベースとして、Microba Community Profiler v2.0.2で種プロファイルを取得した(Parks et al., 2021)。簡単に説明すると、リードはMGDBにマッピングされ、信頼できると考えられる十分なエビデンスを持つ種クラスターの相対的な細胞存在量が推定され、報告された。このデータセットでは、このデータセットから新たに採掘された7種を含む、合計1,757種が同定された(MGDBに固有の964種)。このうち、細菌が1,735種、古細菌が12種、真核生物が10種であった。
メタゲノムサンプル中の遺伝子およびパスウェイ存在量の定量は、Microba Genes (MGENES) データベースv2.0.0に対してMicroba Gene and Pathway Profiler (MGPP) v0.1.0を用いて行った。MGPP は 2 段階のプロセスで行われる。ステップ1では、MGDBの全ゲノムからの全ORFを、MMSeqs2 Release 10-6d92c (Steinegger and Söding, 2017)というツールを用いて、リード長80%以上の90%同一性を用いてUniRef90 release 2019/09に対してクラスタリングした。その後、遺伝子クラスターをUniRef90識別子でアノテーションし、UniProt ID Mappingサービスを介してEnzyme Commission(EC)およびTransporter Classification Database(TCDB)のアノテーションにリンクした。ECアノテーションは、enrichM [https://github.com/geronimp/enrichM]を用いて各ゲノム内のMetaCyc(Caspi et al., 2020)パスウェイのエンコーディングを決定するために使用され、完全またはほぼ完全(完全性>80%)であったパスウェイは、エンコーディングされたものとして分類され、さらなる解析のために保存された。ステップ 2 では、MGENES タンパク質クラスター内の遺伝子の塩基配列と 1 塩基以上(ヌクレオチド空間で)アライメントする DNA シーケンスリードペアをすべて合計し、集計した。パスウェイのアバンダンスは、MCP が報告した全生物種の全ゲノム内に存在する各パスウェイの遺伝子数を平均して算出した。このデータセットでは、合計5,165,783遺伝子が同定された。
ウイルス種
この方法はウイルスと真核生物の呼び出しに最適化されているため、MiCoP(LaPierre et al. 参照データセットとして、MiCoPはNCBIのRefSeq Viralデータベース(Brister et al.) このデータセットで合計200種のウイルスを同定した。
変換とフィルター
メタゲノミクスデータ(分類学的および機能的)は構成データの一形態であり、比例によりデータセット内に負の相関が生じるため、独立性の仮定に違反する。したがって、分散成分分析では中心対数比(clr)変換(Aitchison, 1982)を適用し(オフセット=0.001)、一方、存在量の差分分析では、ANCOMv2.1パッケージ(Mandal et al., 2015)内のカウントデータに対して加法対数比変換を行った(オフセット=1)。分散成分および差分存在量分析では、非ゼロ値が10未満の超低有病率特徴を除去した。これにより、607の細菌種(Microbaデータベースに固有の221)、40のウイルス種、1,742,729遺伝子が残った。ただし、Fisherの正確検定と多様性解析のためにすべてのフィーチャーを保持した。
共変量の選択
さまざまな解析に含める共変量を特定するために予備解析を行った。このプロセスについては後述するが、年齢、性別、および様々な食品群からの(clr変換された)エネルギー割合(「食事PC」)の派生変数を用いて計算された上位3主成分を含めることにした。これらの共変数を合計すると、サンプル内の微生物分類学的α多様性(シャノン指数)の分散の13.5%を説明することができた。
人口統計学的変数
ベースライン共変量として年齢と性別が含まれた。月は当初、食事の季節変動を説明するために(因子変数として)含まれていたが、その後、食事摂取量と分類学的α多様性(Shannon index)の両方の測定にほとんど寄与しなかったため、削除された(分散の12.2%を説明した年齢+性モデルと分散の16.0%を説明した年齢+性+月モデルを比較すると、分散の3.8%が追加された)。食餌調査月を余弦曲線に変換した季節性変数は、分類学的α多様性(Shannon index)の分散に寄与しなかった。
食事の主成分
メタゲノム解析の共変量として、Australian Eating Surveyの食事データを含めた。これは、この指標がエネルギー摂取量(年齢と強い相関がある)の交絡効果を最もよく説明し、また完全率が最も高かったためである。
食事データの顕著な特徴を抽出するために、13のエネルギー割合変数について主成分(食事PC)を作成した。この指標は比例データの一形態であるため、特定の組成を考慮した方法が必要であるため、PCを計算する前に中心対数比(clr)変換を行った。clr変換を用いることの重要性は、3つの証拠に基づいて示唆された。第一に、年齢、性別、月齢の共変量は、clr変換を適用するかどうかによって、食事PC1-3の分散の大きな違いを説明することがわかった。第二に、非変換とclr変換された食事PC間の相関は低く(r = 0.32)、組成を考慮した分析は、組成を考慮しない分析とは異なる結果を提供する可能性があることを示唆している。第三に、clr変換されたデータは、非変換データよりも微生物分類学的α多様性の分散の大きな割合を説明した(エネルギー項目の割合は、clr変換されたデータでは分散の3.7%を説明したのに対し、clr変換されていないデータでは分散の2.5%を説明した)。
最初の3つの食事PCは、食事の分散の43%を占めた。共変量としてclr変換した最初の3つのPCを含めると、微生物の分類学的α多様性の分散が、13の割合のエネルギー項目すべて(2.8%対2.5%)よりも大きく説明され、メタゲノミクス解析に共変量として含めることが正当化された。ASD群では、年齢と性で調整した後、食事のPC3が低かった(肉の摂取量が減少したことを示唆)(図S5)。
分散成分分析
オミックス関係マトリックス(ORM)とOREML
ソフトウェアパッケージOSCA(Zhang et al.) 簡単に言うと、この方法は、p個のプローブまたは変数(この分析では、<10個の非ゼロ値を持つ変数を除去した後、メタゲノミクスデータセットからの分類学的または機能的変数のclr変換カウント)に基づいて、n個の個体の各ペア間のn×nのオミックス関係行列(ORM)を推定する;我々は--orm-alg 2設定を使用した。次に、OREMLを用いて、ある表現型(モデルの従属変数)の分散のうち、ORMに関連する割合を推定し、制限最尤法(REML)の枠組みでランダム効果としてあてはめた。共変量の選択は焦点となる表現型によって異なるが、普遍的に性別と年齢(年齢が従属表現型の場合を除く)、場合によっては参加者グループや食事PCが含まれた。
一般的な特徴(中央値が0を超えるもの)とまれな特徴(中央値が0であるが、全標本で10個以上0でないもの)について、別々のORMを作成した(表S1.1)。これは、複数のORMフレームワーク(-multi-orm)でこれら2つのORMをフィッティングする際に、混合モデルを近似することを意図したもので、「コア」分類群(ほぼ正規分布)と「アクセサリ」分類群(あまり普及しておらず、ゼロ膨張分布を持つ)が存在するという観察に動機づけられている。分析ごとの共変量は図2の凡例に記載されている。
共変量なし、10歳未満のAAB参加者に限定、異なる分類学的および機能的プロファイリングパイプライン(MetaPhlAn2(Truong et al.、2015)およびHUMAnN2(Franzosa et al.、2018))の使用、および抗生物質を現在使用している参加者の除外。これらの解析の詳細は、方法S1に記載されている。
ORMのclr変換対0/1コーディング
希少種の分布に大きな広がりがあったことから、ASD診断、便の一貫性(rBSC)、および食事PCについて、分類群数にclr変換を適用した場合と、有無(0/1)をコード化した場合の効果を比較した。結果は非常に類似しており、clr変換したデータを使用することを支持するものであった(図S2)。
CD4+ T細胞の割合と分散分析
血液中のCD4+ T細胞の割合は、meffilパッケージを使用して、血液由来のDNAメチル化データからn = 151人の参加者について予測された。これは、微生物叢とCD4+制御性T細胞との間の相互作用を報告している他の研究者(Zheng et al.、2020)や、免疫状態がASDと一般的に併発する(Atladóttir et al.、2010;Sabourin et al.、2019;Vargason et al.、2019)ことから、興味深いものであった。多重検定の負担を軽減するため、当初は共通遺伝子の影響のみを検定したが(このデータセットでは通常大きな影響があったため)、関係は認められなかった(b2∼0、SE = 0.06、p = 0.50)。
食事形質と分散分析
マイクロバイオーム組成は、食事PC1の分散と関連しており、マイクロバイオーム組成が共通種に基づく場合はb2 = 13%(SE = 7%、p = 0.03)、希少種に基づく場合はb2 = 8%(SE = 5%、p = 0.4)、共通遺伝子に基づく場合はb2 = 48%(SE = 15%、p = 8.1e-5、FDR有意)、希少遺伝子に基づく場合はb2 = 22%(SE = 18%、p = 7.4e-2)であった(図2、表S1.2)。マイクロバイオームORMからの推定値は、PC1と比較して食事PC2-3では低かった(図2、表S1.2)。しかし、食物ORMは一貫して、食物PC1-3(b2~60%)および食物多様性(b2=52%;図2、表S1.2)の分散の最も高い割合と関連していた。食餌性PC1と複数のORMの組み合わせとの関連に関するさらなる結果は、以下の「ORMの組み合わせによって説明される形質の分散」のセクションに記載されている。
年齢関連食餌性PCに対する年齢の影響
食餌性PC1(植物性食餌からのエネルギーの割合が高いことを表す)が年齢と強く関連していることに注目した。そこで、OREML解析で年齢を共変量として除去した場合のb2推定値への影響を検証したところ、これらの共変量を含めると表現型の分散(b2推定値の分母)が減少するため、複数のORMデータセットでb2推定値が増加することがわかった。特に、共変量として年齢を取り除くと、分散推定値は、共通遺伝子データセットでは49%から55%に、希少遺伝子データセットでは21%から38%に、共通種データセットでは13%から18%に増加した。
分散推定値に対する複数のORMの効果
ベンチマーク形質。食餌PC1を内部ベンチマーク形質として使用した理由は、(1)食餌PC1は植物ベースの食餌を反映しており、これは腸内マイクロバイオームと関連していることが知られている(David et al.、2014)(図1b)、(2)食餌PC解析で最も分散を説明した、(3)食餌PC1の分散は種レベルのマイクロバイオーム組成と強く関連していた(図2、表S1.2~S1.3)。
階層内のORMの組み合わせ。我々は、分類群をより高い分類学的レベル(種対属対科)および機能的レベル(酵素委員会データセットでは、レベル4とレベル3を比較)に折りたたみ、「共通」変数(このデータセットでは中央値>0と定義)を用いて、食事PC1で説明される分散への影響を評価した。これらの分類学的(species: b2 = 13%, SE = 8%, p = 0.03; genus: b2 = 14%, SE = 7%, p = 0.01; family: b2 = 4% SE = 4%, p = 0.15)と機能的(Enzyme Commission level 4: b2 = 48% SE = 13%, p = 3. 8e-3; Enzyme Commission level 3: b2 = 7% SE = 7%, p = 0.50)データセットでは、最も粒度の大きいデータ階層が分散の最も大きな割合を説明する傾向があった(図S3、表S1.3)ので、以下ではこれらに焦点を当てる。種、属、科のORMをそれぞれモデルに組み合わせると、食餌PC1のb2 = 24%(SE = 9%)を発見した(図S3、表S1.3)。これらの結果は、分類学的・機能的階層内のORMは直交していないことも示唆している。
すべての特徴に基づくORMの計算。ORMの計算を一般的な特徴と希少な特徴に層別化しない。例えば、年齢については、"species_all "ORMはb2 = 59% SE = 9%を説明し、一方、共通+希少種ORM(2-ORMモデル)はb2 = 60% SE = 9%を説明した。ASD診断では、"species_all "ORMはb2 = 0%(SE = 8%)を説明したが、普通種+希少種ORMはb2 = 1%(SE = 9%)を説明した。このことは、ORM層別化が有効なアプローチであることを示唆している。
共通種と希少種の特徴 コア変数とアクセサリ変数を捕捉し、それらの相対的な貢献についてより詳細な洞察を提供するために、まず普通種と希少種を層別化した。これらのORMの組み合わせ(混合モデルの近似)が、与えられた形質(この場合は食餌PC1)について説明される分散を改善できるかどうかをテストしたところ、推定値は本質的に同等であることがわかった(普通種+希少種を含めるとb2 = 23%、SE = 12%、普通種単独ではb2 = 13%、SE = 8%、希少種単独ではb2 = 8%、SE = 9%)。興味深いことに、普通種と希少種のORMを組み合わせると、出現した分類学的階層から共通の特徴を使用して計算されたORMの組み合わせ(普通種+希少種b2 = 23%、SE = 12%に対して、種+属+科b2 = 24%、SE = 9%)と比較して、食事PC1について同様の分散推定値が得られた(図S3、表S1.3)。これらの結果は、多重ORM解析からのb2が個々のORM解析の合計に近似していることから、これらのデータセットが本質的に直交していることを示唆している(図S3)。
分類学的および機能的データセット さらに、さまざまな分類学的ORMと機能的ORMを組み合わせることで、形質で説明される分散を改善できるかどうかを調べた。分類学的ORMを組み合わせることで、b2が増加した(例えば、食餌PC1-共通の種b2 = 13%, SE = 8%; 共通の属b2 = 14%, SE = 8%; 共通の科b2 = 4%, SE = 4%, これらすべてを組み合わせた複数のORM b2 = 24%, SE = 9% )、 また、分類学的ORMと機能的ORMの組み合わせ(例:rBSC-共通のEClevel4 b2 = 34%, SE = 12%; 共通のTCDB b2 = 3%, SE = 5%; 共通のMetaCycパスウェイ b2 = 16%, SE = 9%, これら全ての組み合わせの複数ORM b2 = 35%, SE = 13%)や、分類学的ORMと機能的ORMの組み合わせ(例:、 また、分類学的ORMと機能的ORMの組み合わせ(例:年齢-普通種 b2 = 33%, SE = 7%; 希少種 b2 = 53%, SE = 9%; EClevel4 b2 = 56%, SE = 10%; TCDB b2 = 42%, SE = 10%; MetaCycパスウェイ b2 = 40%, SE = 9%; これらすべての複合ORM b2 = 79%, SE = 10%) もあった(図S3、表S1.3)。
機能的データセットの有益性 例えば、食事PC1では、1,834の共通特徴(Enzyme Commission Level 4とTCDB変数の組み合わせ)に基づくb2 = 48%(SE = 13%)に対して、n = 251,617の共通遺伝子に基づくb2 = 48%(SE = 15%、p = 8.1e-5)。
非加算モデルを用いた予測
予測可能性の上限としてのb2の推定は、加法モデルを仮定していることに注意。そこで、加法性を仮定しないadaboost (Freund and Schapire, 1997)モデルによって説明される分散も推定した。
adaboostを用いて、1000回の反復で5倍のクロスバリデーションを行い、n = 607の細菌(clr変換された数)に基づいてASD診断を予測する精度を決定した。兄弟姉妹のランダムサンプリングによる血縁関係の交絡を避けるため、このデータセットをn = 99 ASDとn = 96 UNRの参加者に制限した。トレーニングデータセットを生成するために、各反復で同じ数のASDとUNRの子供(n = 76)をサンプリングした。アダブーストには、RのJOUSBoost実装を使用し、maxdepth = 10、n_rounds = 100の設定を使用した。その結果、診断結果は平均して53%(SD=7%)の確率で正しく分類された(Data S2)。これは、非加算モデルを仮定した場合、マイクロバイオームデータがASD診断を予測する能力はごくわずかであることを示唆している。
この分析をベンチマークするために、b2の結果からこの表現型はマイクロバイオームによって予測されるはずであることから、年齢を従属変数として同じ1000回反復の5倍クロスバリデーションモデルも繰り返した。この分析では、再びn=99のASDとn=96のUNR参加者に限定し、n=607の細菌を予測特徴として入力した。このトレーニングデータセットを生成するために、各反復ごとにデータセットの4/5をランダムにサンプリングした。同じJOUSBoost::adaboost設定を使用した。予測能力の評価を簡単にするため、年齢を10歳より若いか古いかを識別するモデルの能力として精度を計算した(つまり二値変数)。その結果、年齢(10歳未満または10歳以上)は平均して62%(SD = 7%)の確率で正しく分類され、OREML解析から予想されるように、年齢がASD診断よりも予測可能であることがわかった(Data S2)。
個体数の差分析
種レベルの分類学的データ
統計的仮定に対してロバストであるため、存在量の差分分析にはANCOMv2.1(Mandal et al., 2015)(R: https://github.com/FrederickHuangLin/ANCOM)を使用した(Weiss et al., 2017)。簡単に説明すると、各特徴(すなわち、種または遺伝子)について、ANCOMv2.1は、加法-対数比変換(オフセット=1)を実行し、選択された予測因子に対して各変換された分類群について統計的検定を実行し、Benjamini-Hochberg偽発見率閾値(0.05)を通過するペアワイズ検定の数をカウントし、その初期特徴についてW統計量(棄却された帰無仮説の数を表す)を生成する。次に、W-統計量の分位値に基づいて、有意に存在量が異なると宣言されます。有意に存在量が異なると宣言するためには、一般的に0.7を超える検出閾値が使用されます。この解析では、すでに10以上の非ゼロ値を持つ変数を除外していたため、ゼロ外れ値カットオフを使用し(zero_cut引数)、すべてのサンプルが480万リード以上であったため、ライブラリカットオフ(lib_cut)は適用しなかった。全体で607種をテストした。
共変量なし、年齢≦10のAAB参加者に限定、異なる分類学的および機能的プロファイリングパイプライン(MetaPhlAn2(Truong et al.、2015)を使用、現在抗生物質を使用している参加者を除外、SIBグループを除外、兄弟ペアのみを考慮、希少化後に7Mリード未満の参加者を除外など、種レベルの差分存在量について広範な感度分析を行った(Methods S1)。
遺伝子レベルの機能データ
我々は、種レベルの結果を活用し、ANCOM解析により、発現量の異なる遺伝子を、発現量の異なる生物種によってコードされる遺伝子群に絞った。これは、各生物種内の各遺伝子の存在量を詳述した個体ごとの表にアクセスできたため可能であった。種レベルの分類学的データと同じフィルターを使用し、最終的に4,950遺伝子をテストした。
Romboutsia timonensis遺伝子アノテーション
Romboutsia timonensis遺伝子のアノテーションは、ASD群で有意に低いアノテーションであった。有意な(検出閾値>0.7)6遺伝子はすべてUniRef90の識別子を持ち、そのすべてがUniProtKBデータベースのRomboutisa属にマップされ、Romboutsia timonensisに焦点を当てたことと一致した(表S2.12)。関連タンパク質は以下の通りであった: アスパラギン酸-セミアアルデヒド脱水素酵素(ASAデヒドロゲナーゼ)(ASADH)(EC 1.2.1.11)(アスパラギン酸-β-セミアアルデヒド脱水素酵素)-アミノ酸生合成(L-リジン、L-メチオニン、L-スレオニン)に関与する;アミドホスホリボシルトランスフェラーゼ(ATase)(EC 2. 4.2.14)(グルタミンホスホリボシルピロホスフェートアミドトランスフェラーゼ)(GPATase)-プリン代謝およびL-グルタミン酸からL-グルタミンへの代謝に関与;チミジル酸キナーゼ-DNA合成(特にピリミジン)経路に関与;ガラクトキナーゼ(EC 2. 7.1.6)(ガラクトースキナーゼ)-ガラクトース代謝に関与;発芽プロテアーゼ(EC 3.4.24.78)(GPRエンドペプチダーゼ)(発芽プロテイナーゼ)(胞子プロテアーゼ)-細菌の胞子発芽に関与;リボヌクレアーゼ3(EC 3.1.26.3)(リボヌクレアーゼIII)(RNアーゼIII)-dsDNA消化に関与。
IQ-DQの存在量差分析
種の存在量とIQ-DQ複合スコアとの関連を検証したところ、SIB群を含めた場合と除外した場合(共変量:年齢、性別、食事PC1-3)とでは、解析間の一致が不十分であった。ASD対SIB+UNR解析では、Bifidobacterium sp002742445のみが検出閾値>0.7を超え、ASD対UNR解析では5番目に豊富であった(図3E-3F、表S2.13-S2.14)。この解析には限界がある: ASDの診断ステータスはより低いIQと関連しており、ASDと診断された人々にIQ測定(言語能力に依存する)を適用することには限界がある。
順序付け
mixOmicsパッケージ(Rohart et al.) これは、10未満の非ゼロ値を持つ変数を除去し、clr変換を行った後の主成分分析を含んでいた。
多様性分析
データセット中の豊かさと均等性の両方を考慮するため、シャノン指数を用いてアルファ多様性を計算した。感度分析として、アルファ多様性の測定に豊かさとシンプソン指数を用いた場合の影響も検証した(Methods S1, Table S3)。β多様性を計算するために、1,731種の細菌(すなわち古細菌を含まない)の系統樹を用いて計算した加重ユニフラック指数行列を作成した。このβ多様性マトリックスを用いて、PERMANOVA(Anderson, 2008)(年齢と性別で調整)とPERMDISP2(Anderson et al.
分類学的多様性とASD診断
複数のアルファ多様性指標を用いた種レベルでのアルファ多様性には、無視できるほどの群間差があった(ANOVA検定の統計量: シャノン指数 p = 0.13、豊かさ p = 0.13、シンプソン指数 p = 0.08)、年齢、性別、食事PC1-3(ANOVA検定の統計量: シャノン指数 p = 0.31、豊かさ p = 0.34、シンプソン指数 p = 0.37)(図S5B)、および広範な感度分析(方法S1、表S3)。種の多様性を年齢、性、食事PC1-3に回帰すると、α-多様性が高いほど高齢(b = 0.037、p = 1.3e-7)となり、食事PC2は高乳製品食(b = 0.042、p = 0.022)となった。β多様性(加重ユニフラック)(PERMANOVA p = 0.20;図S5A)には群間差はなく、ASD群ではSIB群およびUNR群と比べて分散性に差はなかった(PERMDISP2 p = 0.85;図S5A)。
共変量を加えた多様性解析
共変量として食事性PCおよびrBSCを含めた場合に、食事性と分類学的多様性の関係が変化するかどうかを検証した(データ欠落のためn = 233)。食餌性PC2-3は食餌性多様性と有意に関連し、モデルのR2を改善したが、食餌性-分類学的多様性の関係はまだわずかに有意であった(図S7A-S7B)。
また、エネルギー摂取量を共変量として含めた場合の影響も検証したが、食事と分類学的多様性の関係に有意な影響は認められなかった(図S7C)。
臨床的およびPGSと多様性の関係
線形モデルを用いて、多様性指標(食事および分類学的)と潜在的に関連する表現型指標(rBSC、ADOS2/G比較スコア、ADOS-2/G反復・制限行動(RRB)校正重症度スコア、ADOS-2/G社会的影響校正重症度スコア、社会的反応性尺度tスコア(SRS)、短感覚プロファイル生センサースコア(SSP))、および生物学的指標(ASDの多遺伝子スコア(PGS)(Grove et al、 2019)、ADHD-ASD-TS交叉特性(Yang et al.、2021)および神経症(Nagel et al.、2018)、CD4+ T細胞割合)。多重検定を考慮するために、Benjamini-Hochberg FDR補正を行った(表S3)。これらのモデルを検証するために広範な感度分析を行った(方法S1)。これらの仮説は表S3に明示的に列挙されており、これらの仮説に対する感度分析もそこに説明されている。
多様性の他の上流媒介因子
神経症。不安、神経質、微生物叢の関連を提唱している者もおり(Kimら、2018;Yangら、2019)、機能性胃腸障害(過敏性腸症候群など)と不安およびASDの両方(名目上有意)との遺伝的相関を示す証拠がある(Wuら、2021)。さらに、不安(Simonoffら、2008;Sukhodolskyら、2008)と胃腸の問題(Chaidezら、2014;McElhanonら、2014)の両方がASDと併発することが多い。AABとQTABに包括的な不安特性測定がないため、不安GWASはパワー不足であり、ASDと神経質さには遺伝的相関があるため(Grove et al. 我々は、食事(p = 0.94)または分類学的多様性(p = 0.43)のいずれとも関連を見いださなかった(図6K-6L)。それにもかかわらず、不安の役割についてさらなる調査が必要である。
CD4+ T細胞の割合。我々はさらに、免疫細胞の割合と食事または分類学的多様性との関連を評価し、CD4+T細胞の割合に注目した。この分析の根拠は、自閉症では食物アレルギーや不耐症の割合が増加することが報告されており(Bresnahan et al. 生物学的には、食物アレルギーや不耐症は腹痛、下痢、嘔吐、摂食障害などの消化器症状を引き起こす可能性がある。社会的には、この可能性を認識している親が、子どもの食事を意図的に制限し、それによって行動が改善するかどうかを確認することがある。n=150個体におけるclr変換CD4+T細胞割合のデータを用いたところ、食事(b = 1.3e-2、p = 0.44)または分類学的多様性(t = -0.20、p = 0.10)のいずれとも関連を示す証拠は見つからなかった(図6Iおよび6J)。
代謝産物生産ポテンシャルの推定
機能的MetaCycパスウェイデータセットを用いて短鎖脂肪酸代謝産生能を推定した。ある代謝物について、寄与するパスウェイを同定し(表 S5)、それらのリードカウントを合計した。
プロット
プロットにはRパッケージのggplot2(Wickham, 2016)とggstatsplot(Patil, 2021)を使用した。
データとコードの利用可能性
本論文の結論を裏付けるAABデータセットは、Cooperative Research Centre for Living with Autism(Autism CRC)内のAustralian Autism Biobankに申請することで入手可能である:https://www.autismcrc.com.au/biobank。本解析で使用したQTABデータセットは、仲介アクセスにより入手可能である: UQ eSpace: https://espace.library.uq.edu.au/view/UQ:e803a68 。コードは https://zenodo.org/record/5558047 で公開されている。本論文で報告されたデータの再分析に必要な追加情報は、要請があれば主任連絡先から入手可能である。
謝辞
Australian Autism Biobankの参加者と家族に感謝する。このプロジェクトで使用されたデータは、適切な倫理承認を受けたCo-operative Research Centre for Living with Autism(Autism CRC)から提供されたものである。Autism CRCはオーストラリア政府のCooperative Research Centre Programのもとに設立され、支援を受けている。シドニー小児病院ウェストミードのナタリー・シローブ博士と小児発達病棟チーム、KUマーシャ・バージェス自閉症特異的早期学習・ケアセンターとKUチルドレンズサービスのスタッフ、ビクトリア州でのリクルートにおいて重要な役割を果たしたラ・トローブ大学オルガ・テニソン自閉症研究センターのナンシー・サドカ博士、パースでの血液サンプルの収集と処理に貢献したパスウェストのアニャ・ラヴィーン博士と病理学サービスに感謝する。AABを支援してくれたFelicity Roseに感謝する。QTABプロジェクトは、双子とその家族が快く研究に協力し、時間を惜しみなく提供してくれたことに感謝する。QTABチームの多くの研究アシスタント、およびQTABサンプルの処理とクイーンズランド双生児登録研究にご協力いただいたUQのIMB、複雑形質遺伝学プログラムのHuman Studies Unitに特に感謝する。ADHD-ASD-TS PGS解析のGWAS要約統計を提供してくれたZhiyu Yang、メタゲノミクス解析を手伝ってくれたKylie Ellisに感謝する。Autism CRCの財政的支援に感謝する。QTABは、Australian National Health and Medical Research CouncilのCentre of Research Excellence Grant (1078102)の支援を受けた全国的なリソースであるTwins Research Australiaへのアクセスを通じて促進された。また、Australian National Health and Medical Research Councilからの資金援助(J.G.に1103418と1127440、N.R.W.に1078901と1173790、N.R.W.とP.M.V.に1113400、A.J.O.W.に1077966と1173896、 およびM.J.W.に10787561)、オーストラリア研究評議会(A.F.M.にFT200100837、P.M.V.にFL180100072)、クイーンズランド大学(RTP奨学金および授業料相殺、ならびにC.X.Y.にSam and Marion Frazer HDR Top-up Scholarship in Neurological Disease)。本研究は、メーテル・リサーチとメーテル財団の助成を受け、オーストラリア政府からの助成金を受けたトランスレーショナル・リサーチ・インスティチュート(TRI)で一部実施された。
著者貢献
Australian Autism Biobankは、A.J.O.W.、C.D.、V.E.、H.S.H.、G.A.A.、P.A.D.、J.G.、R.G.、A.K.H.、L.P.L,が発案・設計した。オーストラリア自閉症バイオバンクの患者募集、評価、データ収集は、A.J.O.W.、V.E.、C.D.、M.L.F.、H.S.H.、P.A.D.が主導した、 R.G.、C.H.、A.H.、H.H.、R.J.、F.K.、L.P.L.、J.L.、M.L.F.、A.M.、N.E.M.、M.M.、M.N.は、4つのサイトを通じて調整、データ収集、管理に貢献した。クイーンズランド双生児思春期脳プロジェクトは、M.J.W.、G.I.d.Z.、P.M.T.、K.L.M.が発案・設計し、N.K.H.とL.T.S.が継続的なプロジェクト設計と管理に貢献し、A.K.H.が生物学的サンプルの管理を監督し、J.L.M.とL.N.がサンプルの処理を行った。J.G.、C.X.Y.、A.K.H.はメタゲノム研究をデザインした。C.X.Y.はメタゲノミクスのQCとプロファイリングをD.L.A.W.とL.K.が行い、R.R.、G.A.A.、A.F.M.、G.W.T.、G.H.の助言を得て解析を行った。C.X.Y.、J.G.、N.R.W.は、全著者からの重要な意見を得て原稿を執筆した。
利益申告
David L.A. WoodとLutz KrauseはMicroba Life Sciencesの社員である。Gene W. TysonはMicroba Life Sciencesの共同設立者であり、取締役である。Gerald HoltmannはServatus Biopharmaceuticalsの顧問。他の著者は競合する利害関係はないと宣言している。
インクルージョンと多様性
本試験の質問票が包括的な方法で作成されるよう努めた。被験者のリクルートにおいて、民族やその他の多様性の確保に努めた。被験者のリクルートにおいてジェンダーバランスの確保に努めた。本論文の著者の1人または複数が、科学分野で十分に代表されていない少数民族であることを自認している。本論文の著者の1人または複数が、科学におけるマイノリティ代表を増やすことを目的としたプログラムから支援を受けている。
補足情報
pdfをダウンロード(.33 MB)
pdfファイルのヘルプ
資料S1. STAR Methodsに関連するデータS1とS2
ダウンロード .xlsx (.03 MB)
xlsxファイルのヘルプ
表S1. 分散成分分析結果、図2、S2、S3関連
表S1.1. 分散成分分析に用いた「共通」特徴と「まれ」特徴の数(マイクロバイオームデータセットごと)。表S1.2. 本文の図2に示した解析のOREML解析によるb2推定値。表S1.3. 本文中の図2に示した解析および補足図に示した解析におけるOREML解析からのb2推定値。
.xlsxのダウンロード (.81 MB)
xlsxファイルのヘルプ
表S2. 図3およびS4に関連する存在量差解析結果
表S2.1. ASD診断状態に関する種レベルの細菌分析(非ゼロ値が10未満の分類群を除去した後のn = 607)のANCOM統計量。表S2.2. ASD診断ステータスに関する折りたたみ分類学細菌分析(属、目、門)のANCOM統計量。表S2.3. ASD診断状態に関する種レベルの細菌分析(非ゼロ値10未満の分類群を除去した後のn=607)および感度分析としてSIB群(n=196)を除去した場合のANCOM統計量。表S2.4. ASD診断状態に関する細菌(n = 1754)の種レベルでの差分存在に関するフィッシャーの検定。表S2.5 ASDとSIB+UNRを比較した、MetaCycグループレベルでの全生物種にわたる機能データの存在量の差に関するANCOM統計量。表S2.6 ASDとSIB+UNRを比較したMetaCycパスウェイレベルの全生物種における機能データの差分存在量のANCOM統計量。表S2.7 Enzyme Commissionレベルにおける全生物種間の機能データの差分存在量のANCOM統計量、ASDとSIB+UNRの比較。表S2.8 Romboutsia timonensisに直接マッピングされた遺伝子に着目した遺伝子レベル解析のANCOM統計量。表S2.9 MetaCycグループレベルでRomboutsia timonensisに直接マッピングされた機能データの差分存在量に関するANCOM統計(ASDとSIB+UNRの比較)。表S2.10 MetaCycパスウェイレベルでRomboutsia timonensisに直接マッピングされた機能データの存在量の差に関するANCOM統計量(ASDとSIB+UNRの比較)。表S2.11 Enzyme Commission レベル4データベースからRomboutsia timonensisに直接マッピングされた機能データの存在量の差に関するANCOM統計量、ASDとSIB+UNRの比較。表S2.12 Romboutsia timonensisをターゲットとした遺伝子レベル解析(表S2.8)で検出閾値>0.7となったUniRef90 IDのUniProtKBアノテーション。表S2.13 IQ-DQ複合スコア(n=218)に対する種レベルの細菌解析(非ゼロ値が10未満の分類群を除去した後のn=603)のANCOM統計量。表S2.14 IQ-DQ複合スコアと感度分析としてSIB群(n = 172)を除去した場合の種レベルの細菌分析(非ゼロ値10未満の分類群を除去した後のn = 603)のANCOM統計量。表S2.15 種レベルのウイルス分析(n = 200種)のANCOM統計量。
.xlsxのダウンロード (.02 MB)
xlsxファイルのヘルプ
表S3. 図3、S6、S7、S8に関連する分類学的および食餌多様性線形モデルからの多重検定補正のための統計量の集計。
効果の大きさは、関心のある変数(下線)について提供される。P値は生のP値を示す。P_FDRは、FDR補正されたp値を示す。赤は有意な検定を示し、青はわずかに有意な検定を示す。「参照 "は、この統計量が使用されている箇所を示す。仮説と感度分析は、"N_hypothesis "と "Hypothesis "に明示されている。
ダウンロード .xlsx (.41 MB)
xlsxファイルのヘルプ
表S4. STAR Methodsに関連するメタゲノミクスプロファイリングQC統計量
表S4.1. Microba Genome Database v2 (MGDBv2) の分類学的プロファイリングに関するサマリー統計(本データセットから直接マイニングしたものを含む)。注:本解析のn = 246サンプルはQTAB研究からの追加個体で処理されたため、n = 298サンプルを含む。表S4.2. MicrobaのサンプルごとのQC統計(入力ペアリード数、マッピング統計)。
ダウンロード.xlsx (.01 MB)
xlsxファイルのヘルプ
表S5. STAR Methodsに関連する推定短鎖脂肪酸(SCFA)代謝物量解析結果
a) SCFA量を推定するためにカウントを使用したMetaCycパスウェイ識別子のリスト、b) 食事PC1 ∼SCFAsと共変量の線形モデル、c) 食事PC2 ∼SCFAsと共変量の線形モデル、d) 食事PC3 ∼SCFAsと共変量の線形モデル、e) Bristol Stool Chart(再グループ化) ∼SCFAsと共変量の線形モデル。
ダウンロード.docx (.61 MB)
docxファイルのヘルプ
方法S1. 感度分析、STAR Methods関連
参考文献
Adams J.B.
Johansen L.J.
Powell L.D.
Quig D.
ルービン R.A.
自閉症児の消化管細菌叢と消化管状態--定型児との比較および自閉症重症度との相関。
BMC Gastroenterol. 2011; 11: 22
論文で見る
スコープス (698)
PubMed
クロス
グーグル奨学生
Aitchison J.
組成データの統計分析
J. R. Stat. Soc. B. 1982; 44: 139-160
記事で見る
Google Scholar
アクショモフ N.
ボーモント J.L.
バウアーP.J.
ディクメン S.S.
ガーション R.C.
ムンガスD.
スロトキンJ.
タルスキー D.
ワイントローブ S.
ゼラゾ P.D.
ヒートン R.K.
VIII. NIHツールボックス認知バッテリー(CB):結晶化認知、流動性認知、総合認知の複合スコア。
Monogr. 子どもの発達。2013; 78: 119-132
論文で見る
スコープス (156)
パブコメ
クロス
グーグル奨学生
アルバレスG.A.
ドーソンP.A.
ディッサナーヤケC.
イーペンV.
グラッテンJ.
グローブ R.
ヘンダーズ A.
ホイスラー H.
ローソン L.
マシ A.
他
オーストラリア自閉症バイオバンクチーム
オーストラリア自閉症バイオバンクの研究プロトコール:自閉症発見研究を進めるための国際的リソース。
BMC Pediatr.
論文で見る
スコープス (17)
PubMed
クロス
グーグル奨学生
アンダーソン M.J.
ノンパラメトリック多変量分散分析のための新しい方法。
Austral Ecol. 2008; 26: 32-46
論文で見る
クロスリファレンス
Google Scholar
アンダーソン M.J.
エリングセン K.E.
McArdle B.H.
ベータ多様性の尺度としての多変量分散。
Ecology Letters. 2006; 9: 683-693
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
アスニカーF.
ベリー S.E.
バルデスA.M.
グエン L.H.
ピッキーノG.
ドリュー D.A.
リーミング E.
ギブソン R.
ル・ロイ C.
カティブ H.A.
他。
1,098人の深く表現型決定された個体から得られた、宿主の代謝および習慣的食事とマイクロバイオームとの関連。
Nat. Med. 2021; 27: 321-332
論文で見る
スコープス (356)
PubMed
クロス
グーグル奨学生
アトラドッティル H.O.
ソーセンP.
シェンデルD.E.
Østergaard L.
レムケ S.
パーナー E.T.
小児期の感染症による入院と自閉症スペクトラム障害の診断との関連:デンマークのコホート研究。
Arch. Arch. Pediatr. 思春期。Med. 2010; 164: 470-477
論文で見る
スコープス (79)
PubMed
クロス
グーグル奨学生
Bairoch A.
2000年のENZYMEデータベース。
核酸研究 2000; 28: 304-305
論文で見る
PubMed
クロスフィルム
グーグル奨学生
ベルディング K.
ドノバンS.M.
食事は自閉症スペクトラム障害を持つ子供の微生物叢組成に影響を与えることができます。
Front. Neurosci. 2018; 12: 515
論文で見る
PubMed
クロス
グーグル奨学生
ボクリッチ N.A.
チョンJ.
バタリアT.
ヘンダーソン N.
ジェイ・M.
リー H.
D リーバー A.
ウー F.
ペレス・ペレス G.I.
チェン Y.
et al.
抗生物質、出生様式、および食事は、幼少期のマイクロバイオームの成熟を形成する。
Sci. Transl. Med. 2016; 8: 343ra82
論文で見る
スコープス (861)
PubMed
クロス
グーグル奨学生
ボイドJ.A.
ウッドクロフトB.J.
EnrichMを用いた比較ゲノム解析。
GitHub. 2019;
https://github.com/geronimp/enrichM
記事で見る
グーグル・スカラー
ブレスナハン M.
ホーニッヒ M.
シュルツ A.F.
グネスN.
ヒルツ D.
リー K.K.
マグヌス P.
ライヒボーン・イェンネルド T.
ロート C.
Schjølberg S.
他
母親が報告する乳幼児の胃腸症状と自閉症との関連:前向き出生コホートからのエビデンス。
JAMA Psychiatry. 2015; 72: 466-474
論文で見る
スコープス (90)
PubMed
クロス
グーグル奨学生
ブリスター J.R.
アコ・アデイD.
バオ Y.
ブリンコバ O.
NCBIウイルスゲノムリソース。
Nucleic Acids Res. 2015; 43: D571-D577
論文で見る
スコープス (371)
PubMed
Crossref
グーグル奨学生
バフィントン S.A.
ディ・プリスコG.V.
オーヒトゥンT.A.
アジャミ N.J.
ペトロシーノ・J.F.
コスタ-マティオリM.
微生物の再構成は、母親の食事が誘発する子孫の社会的およびシナプス欠損を逆転させる。
Cell. 2016; 165: 1762-1775
論文で見る
スコパス (732)
PubMed
概要
全文
全文PDF
グーグル・スカラー
バローズT.L.
コリンズ K.
ワトソン J.
ゲスト M.
ボーゲス M.M.
ニーブ M.
ロロ M.
ダンカンソン K.
コリンズC.E.
就学前の子供のための食事の質指数としてのオーストラリア推奨食品スコアの妥当性。
Nutr. J. 2014; 13: 87
記事で見る
スコープ(27)
PubMed
クロス
グーグル奨学生
カスピR.
ビリントンR.
ケセラーI.M.
コタリ A.
クルムメナッカーM.
ミッドフォード P.E.
オング W.K.
ペイリー S.
スブラベティ P.
カープ P.D.
代謝パスウェイと酵素のMetaCycデータベース - 2019年アップデート。
Nucleic Acids Res.
論文で見る
スコープス (436)
PubMed
クロスリファレンス
グーグル奨学生
サーマック S.A.
カーティンC.
バンディーニL.G.
自閉症スペクトラム障害児における食物選択性と感覚過敏。
J. Am. Diet. 2010; 110: 238-246
論文で見る
スコープス (406)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
チャイデスV.
ハンセン R.L.
ハーツ-ピチョットI.
自閉症、発達遅滞、定型発達児における胃腸障害。
J. 自閉症Dev。Disord. 2014; 44: 1117-1127
論文で見る
スコープス (329)
PubMed
クロス
グーグル奨学生
チャクラボルティ P.
カーペンター K.L.H.
メジャー S.
ディーヴァー M.
フェルメール S.
ヘロルド B.
フランツ L.
ハワード J.
ドーソンG.
胃腸の問題は、自閉症スペクトラム障害の幼児における反復行動の増加と関連しているが、社会的コミュニケーションの困難とは関連していない。
自閉症。2020; 25: 405-415
論文で見る
スコープス (19)
パブコメ
クロス
グーグル奨学生
チャン C.C.
チャウ C.C.
テリエL.C.
ヴァティクティ S.
パーセル S.M.
リー J.J.
第2世代PLINK:より大規模でリッチなデータセットへの挑戦。
Gigascience. 2015; 4: 7
論文で見る
スコープス (5487)
PubMed
クロス
グーグル奨学生
クレッソン M.J.
ジェフェリー I.B.
コンデS.
パワーS.E.
オコナー E.M.
キューザック S.
ハリス H.M.B.
コークリー M.
ラクシュミナラヤナン B.
オサリバンO.
他。
腸内細菌叢組成は高齢者の食事と健康に相関する。
Nature. 2012; 488: 178-184
論文で見る
スコープス(2327)
PubMed
クロス
グーグル奨学生
コリンズ C.E.
バローズT.L.
トゥルビーH.
モーガン P.J.
ライト I.M.R.
デイヴィス P.S.W.
キャリスターR.
食物摂取頻度質問票で評価した幼児のエネルギー摂取量と二重標識水法で測定した総エネルギー消費量の比較。
J. Acad。栄養。ダイエット。2013; 113: 459-463
論文で見る
スコープス (35)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
コンスタンチノJ.N.
社会的反応性尺度。
ウェスタン・サイコロジカル・サービス、ロサンゼルス2002
記事で見る
Google Scholar
Dan Z.
毛X.
Liu Q.
Guo M.
Zhuang Y.
Liu Z.
Chen K.
Chen J.
Xu R.
Tang J.
et al.
腸内細菌プロファイルの変化は、自閉症スペクトラム障害の代謝活性異常と関連している。
Gut Microbes. 2020; 11: 1246-1267
論文で見る
スコープス (143)
PubMed
クロス
グーグル奨学生
ダッシュ S.
クラークG.
ベルク M.
ジャッカF.N.
精神医学における腸内細菌叢と食事:うつ病を中心に。
Curr. 意見。Psychiatry. 2015; 28: 1-6
論文で見る
スコープス (261)
PubMed
クロス
グーグル奨学生
デビッド L.A.
モーリスC.F.
カーモディ R.N.
グーテンバーグ D.B.
バトンJ.E.
ウルフ B.E.
リン A.V.
デブリン A.S.
ヴァルマ Y.
フィッシュバッハM.A.
ほか
食事はヒトの腸内細菌叢を迅速かつ再現性よく変化させる。
Nature. 2014; 505: 559-563
論文で見る
スコープス (6360)
PubMed
クロス
グーグル奨学生
デ・アンジェリス M.
ピッコロM.
ヴァニーニ L.
シラグーザ S.
デ・ジャコモ A.
セラザネッティ D.I.
クリストフォリ F.
グエルゾーニ M.E.
ゴベッティ M.
フランカヴィラR.
自閉症と特定不能の広汎性発達障害を持つ子供の糞便微生物叢とメタボローム。
PLoS ONE。2013; 8: e76993
論文で見る
スコパス (565)
PubMed
クロス
グーグル奨学生
ファロニーG.
ヨーセンスM.
ヴィエイラ・シルヴァ S.
Wang J.
ダージ Y.
ファウスト K.
クリルシコフ A.
ボンダー M.J.
バジェス・コロマーM.
ヴァンデプッテD.
他。
腸内細菌叢変異の集団レベル解析。
Science. 2016; 352: 560-564
論文で見る
スコープス (1417)
PubMed
クロス
グーグル奨学生
ファインゴールド S.M.
ダウド S.E.
ゴンチャロワV.
リュー C.
ヘンリー K.E.
ウォルコット R.D.
ユン E.
Summanen P.H.
グランピーシェD.
ディクソン D.
他。
自閉症児と対照児の糞便微生物叢のパイロシークエンス研究。
Anaerobe. 2010; 16: 444-453
論文で見る
(712件)
PubMed
クロス
グーグル奨学生
フーキエJ.
モレノ・ワイザーN.
ドネリーJ.
グリックマン C.
カン D.-W.
マルドナド J.
ジョーンズ R.A.
ジョンソン K.
アダムス J.B.
クラジュマルニク・ブラウン R.
ロズポーンC.
自閉症における腸内細菌叢: 行動変容の研究現場効果と縦断的分析。
mSystems. 2021; 6 (e00820): e00848
論文で見る
パブコメ
クロス
グーグル奨学生
フランゾーザ E.A.
マカイバーL.J.
ラーナバードG.
トンプソン L.R.
シルマー M.
ワインガートG.
リプソン K.S.
ナイト R.
カポラソ J.G.
セガタ N.
ハッテンハワー C.
メタゲノムとメタトランスクリプトームの種レベルでの機能プロファイリング。
Nat. Methods. 2018; 15: 962-968
論文で見る
スコープス (843)
PubMed
クロス
グーグル奨学生
フロイント Y.
Schapire R.E.
オンライン学習の決定論的一般化とブースティングへの応用。
J. Comput. Syst. サイエンス 1997; 55: 119-139
論文で見る
クロス
グーグル奨学生
ゴンダリア S.V.
パロンボ E.A.
ノウルズ S.R.
コックス S.B.
マイヤー D.
オースティンD.W.
自閉症児(消化管機能障害の有無にかかわらず)とその神経型きょうだい児の消化管微生物叢の分子的特徴。
自閉症研究 2012; 5: 419-427
論文で見る
スコープス (147)
PubMed
クロス
グーグル奨学生
グローブJ.
リプケ S.
アルスT.D.
マットハイゼン M.
ウォルターズ R.K.
ウォン H.
パレセン J.
アゲルボ E.
アンドレアセン O.A.
アニーR.
他
精神医学ゲノミクスコンソーシアム自閉症スペクトラム障害ワーキンググループ
BUPGEN
精神ゲノミクスコンソーシアムの大うつ病性障害ワーキンググループ
23andMe研究チーム
自閉症スペクトラム障害の一般的な遺伝子リスク変異の同定。
Nat. Genet. 2019; 51: 431-444
論文で見る
スコープス(1023)
PubMed
クロス
グーグル奨学生
ハディザデ F.
ウォルター S.
ベルフアンM.
ボンフィリオF.
ハインセン F.-A.
アンドレアソン A.
アグレウス L.
エングストランド L.
ベインズ J.F.
ラフターJ.
他。
便の回数は腸内細菌叢の構成と関連している。
Gut. 2017; 66: 559-560
論文で見る
スコープス (43)
PubMed
クロス
グーグル奨学生
ホー L.K.H.
トンV.J.W.
シン N.
ナガラジャン N.
タム E.H.
テイ S.K.
ショーリー・S.
タムビー P.A.
ローE.C.N.
自閉症スペクトラム障害児における腸内細菌叢の変化:系統的レビュー。
Gut Pathog. 2020; 12: 6
論文で見る
スコープス (72)
PubMed
クロス
グーグル奨学生
シャオ E.Y.
マクブライド S.W.
Hsien S.
シャロンG.
ハイド E.R.
マキュー T.
コデリ J.A.
チャウ J.
ライズマン S.E.
ペトロシーノ・J.F.
他
微生物叢は神経発達障害に伴う行動的・生理的異常を調節する。
Cell. 2013; 155: 1451-1463
論文で見る
筑波大学
パブコメ
概要
全文
全文PDF
グーグル奨学生
フスV.
ロードC.
自閉症診断観察スケジュール、モジュール4:改訂されたアルゴリズムと標準化された重症度スコア。
J. 自閉症Dev. Disord. 2014; 44: 1996-2012
論文で見る
スコープス (355)
パブコメ
クロス
グーグル奨学生
フス V.
ゴッサムK.
ロードC.
ADOS領域スコアの標準化:社会的情動の重症度と制限・反復行動の分離。
J. Autism Dev. Disord. 2014; 44: 2400-2412
論文で見る
スコープス (383)
パブコメ
クロス
グーグル奨学生
ジャッカ F.N.
チェルビンN.
アンスティK.J.
バターワースP.
逆因果は食事とうつ病の関係を説明するか?
J. 感情。Disord. 2015; 175: 248-250
論文で見る
パブコメ
クロスレフ
グーグル奨学生
カン D.-W.
アダムスJ.B.
グレゴリー A.C.
ボロディT.
チティック L.
ファザーノ A.
コルッツ A.
ガイス E.
マルドナード J.
マクドノー=ミーンズS.
他。
微生物叢移植療法は腸内生態系を変化させ、胃腸症状と自閉症症状を改善する:非盲検試験。
Microbiome. 2017; 5: 10
論文で見る
スコープス (822)
PubMed
クロス
グーグル奨学生
カン D.W.
イルハン Z.E.
イサーン N.G.
ホイト D.W.
ハウズモン D.P.
シェイファー M.
ロズポーン C.A.
ハーン J.
アダムス J.B.
クラジュマルニク-ブラウンR.
自閉症スペクトラム障害児の糞便微生物代謝産物および微生物叢の相違。
Anaerobe. 2018; 49: 121-131
論文で見る
スコープス (215)
PubMed
クロス
グーグル奨学生
カン D.-W.
アダムスJ.B.
コールマン D.M.
ポラード E.L.
マルドナードJ.
マクドノー=ミーンズ S.
カポラソ J.G.
クラジュマルニク-ブラウンR.
自閉症症状と腸内細菌叢に対する微生物叢移植療法の長期的効果。
Sci. Rep. 2019; 9: 5821
論文で見る
スコープス (371)
PubMed
クロス
グーグル奨学生
Kim H.-N.
Yun Y.
Ryu S.
チャン Y.
Kwon M.-J.
Cho J.
シン H.
Kim H.-L.
成人における腸内細菌叢と性格の相関: 横断的研究。
脳行動。Immun. 2018; 69: 374-385
論文で見る
スコープス (64)
PubMed
クロス
グーグル奨学生
コハネ I.S.
マクマリーA.
ウェーバーG.
マックファデンD.
ラパポートL.
クンケル L.
ビッケル J.
ワッタナシンN.
スペンス S.
マーフィー S.
チャーチルS.
自閉症スペクトラムの小児および若年成人の併存疾患負担。
PLoS ONE。2012; 7: e33224
記事で見る
PubMed
クロスフィルム
グーグル奨学生
クーマー T.
トーマスT.R.
ポットシュミットN.R.
ルター M.
マイケルソンJ.J.
大規模自閉症コホートにおけるARFIDの有病率と遺伝的リスク機序の推定。
フロント。Psychiatry. 2021; 12: 668297
論文で見る
スコープス (23)
PubMed
クロス
グーグル奨学生
ラピエール N.
マングルS.
アルザー M.
マンドリック I.
ウー N.C.
コスリッキ D.
エスキン E.
MiCoP:メタゲノムサンプル中のウイルスおよび真菌を検出するための微生物群集プロファイリング法。
BMC Genomics. 2019; 20: 423
論文で見る
スコープス (22)
PubMed
クロス
グーグル奨学生
Li H.
ダービン R.
Burrows-Wheeler変換を用いた高速で正確なショートリードのアライメント。
Bioinformatics. 2009; 25: 1754-1760
論文で見る
筑波大学
PubMed
クロスリファレンス
グーグル奨学生
ロイド-ジョーンズL.R.
ゼン J.
シドレンコJ.
イェンゴ L.
モーザー G.
ケンパー K.E.
ワン・エイチ
鄭 Z.
マギ R.
エスコ T.
他。
要約統計量を用いたベイズ重回帰による多遺伝子予測の改善。
Nat. Commun. 2019; 10: 5086
論文で見る
スコープス (186)
PubMed
クロス
グーグル奨学生
ロードC.
ラッターM.
ディラヴォアP.C.
リジ S.
ゴッサム K.
ビショップ S.L.
自閉症診断観察スケジュール。
第2版。西部心理サービス、2012年(ADOS-2)
記事で見る
Google Scholar
マンダル S.
Van Treuren W.
ホワイトR.A.
エッゲスボM.
ナイト R.
ペッダダ S.D.
マイクロバイオームの組成分析:微生物組成研究のための新しい方法。
Microb. Ecol. Health Dis. 2015; 26: 27663
論文で見る
パブコメ
クロスフィルム
グーグル奨学生
マーシャル S.
ワトソンJ.
バローズT.
ゲストM.
コリンズC.E.
オーストラリアの子供と思春期の推奨食品スコアの開発と評価:横断的研究。
栄養学。J. 2012; 11: 96
記事で見る
スコープ(57)
PubMed
クロス
グーグル奨学生
マケルハノンB.O.
マクラッケンC.
カーペンS.
シャープW.G.。
自閉症スペクトラム障害における胃腸症状:メタアナリシス。
小児科。2014; 133: 872-883
論文で見る
スコープス (480)
パブコメ
クロス
グーグル奨学生
マッキントッシュ D.N.
ミラーL.J.
シュウ V.
ダン W.
短感覚プロフィールの開発と検証。
Psychological Corporation, San Antonio, TX1999.
記事で見る
グーグル・スカラー
マクマーディ P.J.
ホームズ S.
Waste not, want not: Why rarefying microbiome data is notadmissible.
PLoS Comput. Biol.
論文で見る
スコープス (1782)
PubMed
クロス
グーグル奨学生
ミン J.L.
ヘマニ G.
デイビー・スミスG.
レルトン C.
スダーマン M.
Meffil:非常に大規模なDNAメチル化データセットの効率的な正規化と解析。
Bioinformatics. 2018; 34: 3983-3989
論文で見る
スコープス (132)
PubMed
クロスフィルム
グーグル奨学生
モレンダイク M.
モレロ P.
オルトゥニョ・サンチェス=ペドレーニョF.
ヴァン・デル・ドス W.
アンヘル・マルティネス-ゴンサレスM.
食事の質とうつ病リスク: 前向き研究のシステマティックレビューと用量反応メタアナリシス。
J. 感情。Disord. 2018; 226: 346-354
論文で見る
スコープス (316)
PubMed
クロス
グーグル奨学生
ミューレンE.M.
マレン・スケールズ・オブ・アーリー・ラーニング.
AGS版。アメリカン・ガイダンス・サービス, 1995
記事で見る
グーグル・スカラー
ナーゲル M.
ヤンセン P.R.
ストリンガー S.
ワタナベ・K.
デ・リュー C.A.
ブリオワ J.
サベージ J.E.
ハマシュラグ A.R.
スキーン N.G.
ムニョス・マンチャドA.B.
他
23andMe研究チーム
449,484人を対象とした神経症に関するゲノムワイド関連研究のメタアナリシスにより、新規の遺伝子座と経路が同定された。
Nat. Genet. 2018; 50: 920-927
論文で見る
スコープス (340)
PubMed
クロス
グーグル奨学生
ニーフス R.
ロードC.
自閉症スペクトラム障害児の早期病歴。
J. Dev. Behav. 2006; 27: S120-S127
論文で見る
パブコメ
クロスフィルム
グーグル奨学生
オーエンスJ.A.
スピリートA.
マクギンM.
子どもの睡眠習慣調査票(CSHQ):学齢児童を対象とした調査票の心理測定学的特性。
睡眠。2000; 23: 1043-1051
論文で見る
パブコメ
クロスフィルム
グーグル奨学生
パノシアン C.
リオンズウォールP.
ホワイトハウスA.
オディ W.H.
ロー J.
スコット J.
オサリバンT.A.
自閉傾向が強い若年成人は、小児期において食品の種類と食事の質が低い。
J. 自閉症Dev。Disord. 2020; 51: 685-696
論文で見る
スコープス (8)
クロスリファレンス
グーグル奨学生
パークス D.H.
リガートF.
ヴェラ=ウルフP.
クラウスL.
ヒューゲンホルツP.
タイソンG.W.
ウッド D.L.A.
ヒト腸内細菌叢のメタゲノミクスデータセットの分類学的プロファイリングにおけるMicroba Community Profilerの評価。
Front. Microbiol. 2021; 12: 643682
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
パラチョ H.M.R.T.
ビンガム M.O.
ギブソンG.R.
マッカートニーA.L.
自閉症スペクトラム障害児と健常児の腸内細菌叢の違い。
J. Med. Microbiol. 2005; 54: 987-991
論文で見る
スコープス (596)
PubMed
クロス
グーグル奨学生
パティル I.
統計の詳細を視覚化する: ggstatsplot'アプローチ。
オープンソースソフトウェアのジャーナル。2021; 6: 3167
https://doi.org/10.21105/joss.03167
記事で見る
クロス
グーグル・スカラー
パーセル S.M.
チャン C.C.
PLINK 1.9.
2015 (https://www.cog-genomics.org/plink/1.9)
記事で見る
グーグル・スカラー
レイニアス L.E.
アセベド N.
ヨーリンク M.
パーシャゲンG.
ダーレン S.-E.
グレコ D.
セーデルヘル C.
シェイニウス A.
ケレ J.
精製ヒト血液細胞におけるDNAメチル化の差異:細胞系譜と疾患感受性研究への示唆。
PLoS ONE. 2012; 7: e41361
論文で見る
スコープス (752)
パブコメ
クロス
グーグル奨学生
リカボーニ D.
マイリェM.
ケライフィア S.
ラウール D.
ミリオン M.
ヒト腸管から分離された新種Romboutsia timonensis。
New Microbes New Infect. 2016; 12: 6-7
論文で見る
PubMed
クロスフィルム
グーグル奨学生
ロアート F.
ゴーティエB.
シン A.
レー・カオ K.-A.
mixOmics: オミックス特徴選択と複数データ統合のためのRパッケージ。
PLoS Comput. Biol. 2017; 13: e1005752
論文で見る
スコープス (1665)
PubMed
クロス
グーグル奨学生
ロスチャイルドD.
ワイスブロッドO.
バーカンE.
クリルシコフ A.
コレム T.
ゼビ D.
コステア P.I.
ゴドネバ A.
カルカ I.N.
バーN.
他
ヒトの腸内細菌叢の形成において、宿主の遺伝学よりも環境が優勢である。
Nature. 2018; 555: 210-215
論文で見る
スコープス (1594)
PubMed
クロス
グーグル奨学生
ロスチャイルドD.
レヴィアタンS.
ハネマンA.
コーエンY.
ワイスブロッドO.
シーガル E.
2大陸の大規模コホートに基づく、表現型形質との頑健なマイクロバイオーム関連性のアトラス。
bioRxiv;
https://doi.org/10.1101/2020.05.28.122325
論文で見る
スコープス (0)
クロスリファレンス
グーグル奨学生
Sabourin K.R.
レイノルズA.
シェンデルD.
ローゼンバーグ S.
クロエンL.A.
ピント-マーティンJ.A.
シーブ L.A.
ニューシャファー C.
リー L.-C.
ディギセッピC.
自閉症スペクトラム障害児における感染症: 早期発達を探る研究(SEED)。
自閉症研究 2019; 12: 136-146
論文で見る
スコープス (20)
PubMed
クロス
グーグル奨学生
サイアー・ジュニア、M.H.
レディ V.S.
津B.V.
アーメド M.S.
リー C.
Moreno-Hagelsieb G.
トランスポーター分類データベース(TCDB):最近の進歩。
Nucleic Acids Res. 2016; 44: D372-D379
論文で見る
スコープス (468)
PubMed
Crossref
グーグル奨学生
シュレック K.A.
ウィリアムズK.
自閉症スペクトラム障害児の食物嗜好と食物選択性に影響を与える要因。
Res. 開発。Disabil. 2006; 27: 353-363
論文で見る
スコープス (211)
PubMed
クロス
グーグル奨学生
シャナハン E.R.
チョン L.
タリー N.J.
モリソン M.
ホルトマンG.
消化管粘膜関連微生物叢の特性解析:内視鏡手技中の交差汚染を防止するための新規技術。
Aliment. Pharmacol. Ther. 2016; 43: 1186-1196
論文で見る
スコープス (58)
PubMed
クロス
グーグル奨学生
シャロン G.
クルーズ N.J.
カン D.-W.
Gandal M.J.
ワン・ビー
キム Y.-M.
ジンク E.M.
ケイシー C.P.
テイラー B.C.
レーン C.J.
他
自閉症スペクトラム障害のヒト腸内細菌叢はマウスの行動症状を促進する。
Cell. 2019; 177 (e17): 1600-1618
論文で見る
スコープス (568)
PubMed
要旨
全文
全文PDF
グーグル奨学生
シモノフ E.
ピクルス A.
チャーマンT.
チャンドラー S.
ルーカスT.
ベアードG.
自閉症スペクトラム障害児の精神疾患:集団由来サンプルにおける有病率、併存率、関連因子。
J. J. Am. J. Am. Child Adolesc. Psychiatry. 2008; 47: 921-929
論文で見る
スコパス(2305)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ソン J.S.
鄭 L.J.
ロウエル L.M.
ティエン X.
Zhang Y.
Zhu W.
リッチャー・ケリー L.
ガドー K.D.
ガトゥング G.
ロバートソン C.E.
他
サイモンズ・シンプレックス・コレクションにおける自閉症スペクトラム障害児と神経発達障害児の兄弟における糞便微生物叢の比較。
PLoS ONE. 2015; 10: e0137725
論文で見る
スコープス (152)
PubMed
クロス
グーグル奨学生
シュタイネガーM.
Söding J.
MMseqs2により、膨大なデータセットの解析のための高感度なタンパク質配列検索が可能になった。
Nat. Biotechnol. 2017; 35: 1026-1028
論文で見る
スコープス (948)
PubMed
クロス
グーグル奨学生
ストラティ F.
カヴァリエリD.
アルバネーゼD.
デ・フェリーチェ C.
ドナティ C.
ハイエク J.
ジョウソン O.
レオンチーニ S.
レンツィ D.
カラブロ A.
デ・フィリッポC.
自閉症スペクトラム障害における腸内細菌叢の変化に関する新たな証拠。
Microbiome. 2017; 5: 24
論文で見る
スコープス (590)
PubMed
クロス
グーグル奨学生
スホドルスキー D.G.
スカヒルL.
ガドーK.D.
アーノルドL.E.
アマン M.G.
マクドゥーグル C.J.
マクラッケン J.T.
ティアニー E.
ウィリアムズ・ホワイト S.
ルカバリエ L.
ヴィティエロB.
広汎性発達障害児における親が評価した不安症状:頻度と自閉症の中核症状および認知機能との関連。
J. 異常。Child Psychol. 2008; 36: 117-128
論文で見る
スコープス (369)
PubMed
クロス
グーグル奨学生
タン B.
ピアッツァC.C.
ドレザルD.
スタインM.T.
自閉症児2名における重度の摂食障害と栄養不良。
J. Dev。Behav. 小児科 2011; 32: 264-267
論文で見る
スコープス (15)
PubMed
クロス
グーグル奨学生
チュオン D.T.
フランゾーザ E.A.
ティックル T.L.
ショルツ M.
ワインガートG.
パゾッリ E.
テット A.
ハッテンハワー C.
セガタ N.
メタゲノム分類学的プロファイリングを強化するMetaPhlAn2。
Nat. Methods. 2015; 12: 902-903
論文で見る
スコープス (1272)
PubMed
クロス
グーグル奨学生
バルデス A.M.
ウォルターJ.
セガール E.
スペクターT.D.
栄養と健康における腸内細菌叢の役割。
BMJ. 2018; 361: k2179
記事で見る
スコープス (1178)
PubMed
クロス
グーグル奨学生
バジェス-コロマーM.
ファロニーG.
ダルジ Y.
Tigchelaar E.F.
ワン J.
ティト R.Y.
シヴェック C.
クリルシコフ A.
ヨッセンス M.
ワイメンガC.
他
生活の質とうつ病におけるヒト腸内細菌叢の神経活性の可能性。
Nat. Microbiol. 2019; 4: 623-632
論文で見る
スコープス (995)
PubMed
クロス
グーグル奨学生
ヴァンデプッテD.
ファロニーG.
ヴィエイラ-シルヴァS.
ティト R.Y.
ヨッセンス M.
レーズ J.
便の一貫性は、腸内細菌叢の豊かさと組成、腸型および細菌増殖速度と強く関連している。
Gut. 2016; 65: 57-62
論文で見る
スコープス (618)
PubMed
クロス
グーグル奨学生
ヴァルガソンT.
フライ R.E.
マクギネスD.L.
ハーンJ.
幼児期における自閉症スペクトラム障害の併発疾患のクラスタリング: 医療費請求データのレトロスペクティブ分析。
自閉症研究 2019; 12: 1272-1285
論文で見る
スコープス (33)
PubMed
Crossref
グーグル奨学生
ウォルターJ.
アーメットA.M.
フィンレイ B.B.
シャナハン F.
腸内マイクロバイオームの因果関係の確立または誇張: ヒト微生物叢関連げっ歯類からの教訓。
Cell. 2020; 180: 221-232
論文で見る
スコープス (262)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ワン Y.
Zuo T.
Xu Z.
Zhang F.
Zhan H.
チャン D.
Leung T.-F.
Yeoh Y.K.
チャン F.K.L.
チャン R.
他
自閉症スペクトラム障害児における非侵襲的予測マーカーとしての腸内細菌叢の未発達と細菌種。
Gut. 2021;
https://doi.org/10.1136/gutjnl-2020-324015
論文で見る
スコパス (47)
クロスリファレンス
グーグル奨学生
Wang L.
クリストファーセン C.T.
ソリッヒ M.J.
ガーバー J.P.
アングリーM.T.
コンロン M.A.
自閉症児の糞便中の粘液溶解菌Akkermansia muciniphilaおよびビフィドバクテリウム属菌の相対存在量の低さ。
Appl. Microbiol. 2011; 77: 6718-6721
論文で見る
スコープス (309)
PubMed
クロス
グーグル奨学生
Wang M.
Wan J.
Rong H.
He F.
Wang H.
Zhou J.
Cai C.
Wang Y.
Xu R.
Yin Z.
Zhou W.
自閉症スペクトラム障害児における腸内細菌叢組成の変化に伴う腸グルタミン酸代謝の変化。
mSystems. 2019; 4 (e00318): e00321
論文で見る
パブコメ
クロスフィルム
グーグル奨学生
ワン D.D.
グエン L.H.
Li Y.
Yan Y.
Ma W.
リノット E.
アイヴィー K.L.
シャイ I.
ウィレット W.C.
フー・F.B.
他
腸内細菌叢は、地中海食と心代謝疾患リスクとの間の保護的関連を調節する。
Nat. Med. 2021; 27: 333-343
論文で見る
スコープス (145)
パブコメ
クロス
グーグル奨学生
ワトソンJ.F.
コリンズ C.E.
シブリットD.W.
ディブリー M.J.
ガーグ M.L.
オーストラリアの小児および青年を対象とした食物摂取頻度調査票の再現性および比較妥当性。
Int. J. Behav. Nutr. Phys. Act. 2009; 6: 62
論文で見る
スコープス (166)
PubMed
クロス
グーグル奨学生
ウェクスラーD.
ウェクスラー児童用知能尺度。
第4版。PsychCorp, San Antonia, TX2003
記事で見る
グーグル・スカラー
ワイス S.
シューZ.Z.
ペッダダ S.
アミールA.
ビッティンガーK.
ゴンザレスA.
ロズポーン C.
ザネベルト J.R.
バスケス=バエサ Y.
バーミンガムA.
他。
正規化と微生物存在量の差分戦略はデータ特性に依存する。
Microbiome. 2017; 5: 27
論文で見る
スコープス (1097)
PubMed
クロス
グーグル奨学生
ウィッカム H.
ggplot2: データ分析のためのエレガントなグラフィックス.
Springer-Verlag New York, 2016 (ISBN 978-3-319-24277-4. https://ggplot2.tidyverse.org)
記事で見る
クロスリファレンス
グーグル・スカラー
ウィリアムズ B.L.
ホーニッヒ M.
ブイエT.
バウマン M.L.
チョー・ペク M.
ウィック I.
ベネット A.
ジャバド O.
ハーシュバーグ D.L.
リプキンW.I.
自閉症と胃腸障害を有する小児の腸における炭水化物の消化・輸送障害と粘膜異種生殖障害。
PLoS ONE. 2011; 6: e24585
論文で見る
スコープス (330)
PubMed
クロス
グーグル奨学生
ウー Y.
マレーG.K.
バーンE.M.
シドレンコ J.
ヴィッシャー P.M.
Wray N.R.
消化性潰瘍疾患のGWASは、ヘリコバクター・ピロリ感染、他の消化器疾患、うつ病に関与している。
Nat. Commun. 2021; 12: 1146
論文で見る
スコープス (73)
パブコメ
クロス
グーグル奨学生
ヤン B.
ウェイ J.
Ju P.
Chen J.
腸内細菌叢の調節が不安症状に及ぼす影響: 系統的レビュー。
General psychiatry. 2019; 32
https://doi.org/10.1136/gpsych-2019-100056
論文で見る
スコープス (76)
Crossref
グーグル奨学生
ヤン・Z.
ウー・H.
リー P.H.
Tsetsos F.
Davis L.K.
ユー D.
リー S.H.
ダルスガード S.
Haavik J.
バルタ C.
他
衝動性-衝動性スペクトラムに沿ったトゥレット症候群と併存する神経発達障害に共通する遺伝的基盤の調査。
Biol. Biol. 2021; 90: 317-327
論文で見る
スコパス (41)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
ヤップ C.X.
アルバレス G.A.
ヘンダース A.K.
リン T.
ウォレス L.
ファレリー A.
マクラーレンT.
ベリー J.
ヴィンクフイゼン A.A.E.
Trzaskowski M.
他。
オーストラリア自閉症バイオバンクにおける共通遺伝子変異と希少CNVの解析。
Mol. Autism. 2021; 12: 12
論文で見る
スコープス (9)
PubMed
クロス
グーグル奨学生
Zhang F.
Chen W.
Zhu Z.
Zhang Q.
Nabais M.F.
Qi T.
ディアリー I.J.
レイ N.R.
ヴィッシャー P.M.
マクレー A.F.
Yang J.
OSCA:オミックデータに基づく複合形質解析のためのツール。
Genome Biol.
論文で見る
スコープス (63)
PubMed
クロスフィルム
グーグル奨学生
鄭D.
リウィンスキーT.
エリナブE.
健康と病気における微生物叢と免疫の相互作用。
細胞研究 2020; 30: 492-506
論文で見る
スコープス(1359)
PubMed
クロス
グーグル奨学生
ツェルナコワ A.
クリルシコフA.
ボンダーM.J.
Tigchelaar E.F.
Schirmer M.
ヴァタネン T.
ムジャギッチ Z.
ビラ A.V.
ファロニー G.
ヴィエイラ・シルヴァS.
他
LifeLinesコホート研究
集団ベースのメタゲノム解析により、腸内細菌叢の組成と多様性のマーカーが明らかになった。
Science. 2016; 352: 565-569
論文で見る
スコープス (1169)
PubMed
クロス
グーグル奨学生
ジマー M.H.
ハートL.C.
マニング-コートニーP.
マレー D.S.
ビン N.M.
夏S。
自閉症児の栄養状態の予測因子としての食品の多様性。
J. 自閉症Dev。Disord. 2012; 42: 549-556
論文で見る
スコープス (155)
パブコメ
クロス
グーグル奨学生
論文情報
出版履歴
発行日:2021年11月11日 2021年11月11日、オンライン版で訂正: 2023年12月15日
受理済み 2021年10月13日
修正版受理:2021年10月13日 2021年6月14日
受理 2020年12月16日
識別
DOI: https://doi.org/10.1016/j.cell.2021.10.015
著作権
© 2021 Elsevier Inc.
ユーザーライセンス
Elsevierユーザーライセンス|情報アイコンの再利用方法
サイエンスダイレクト
ScienceDirectでこの記事にアクセスする
図
図サムネイルfx1
グラフィカルアブストラクト
サムネイルgr1
図1入力データセットと解析の概要
図のサムネイルgr2
図2マイクロバイオーム組成に関連する表現型分散(±SE)の割合(b2)
図サムネイルfigs1
図S1様々なマイクロバイオームデータセットのORM対角要素と非対角要素の関係
サムネイル図2
図S2OREML推定値における中心対数比変換と2値化0/1符号化の効果
サムネイル図3
図S3分散成分分析の追加結果
図サムネイルgr3
図3ANCOMv2.1を用いたアバンダンスの差分検定
図サムネイルfigs4
図S4アバンダンスの感度分析
図のサムネイルfigs5
図S5参加者グループと食事および便の変数との関係
図のサムネイルgr4
図4食事および分類学的多様性(Shannon Indexを用いて測定)とASD関連表現型との関係
図サムネイルfigs6
図S6食事の質および分類学的多様性とASD関連表現型との関係
図サムネイルgr5
図5マイクロバイオームの多様性解析
図5マイクロバイオームの多様性解析
図S7共変量を追加した食事の多様性と分類学的多様性の関係
図5マイクロバイオームの多様性解析
図S8便の一貫性と食事および分類学的多様性との関係(共変量を追加した場合
図のサムネイルgr6
図6他の食事および分類学的多様性の指標と他の表現型との関係
表
表1メタゲノミクス研究参加者の人口統計学的概要
リンク記事
自閉症に関連した食事の嗜好が自閉症と腸内細菌叢の関連を媒介する
Yap et al.
Cell2024年1月18日
インブリーフ全文PDF
関連記事
広告
ライフ&メディカルサイエンスジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノム学
細胞宿主と微生物
細胞代謝
細胞レポート
セルレポーツ医学
細胞幹細胞
細胞システム
カレントバイオロジー
発生細胞
免疫
医学
分子細胞
ニューロン
構造
アメリカン・ジャーナル・オブ・ヒューマン・ジェネティクス (パートナー)
生物物理学雑誌 (パートナー)
生物物理レポート (パートナー)
Human Genetics and Genomics Advances (パートナー)
分子植物 (パートナー)
分子療法 (パートナー)
分子治療法と臨床開発 (パートナー)
分子療法核酸 (パートナー)
分子療法腫瘍学 (パートナー)
Plant Communications (パートナー)
幹細胞レポート (パートナー)
生化学の動向
癌の動向
細胞生物学の動向
生態学と進化の動向
内分泌学・代謝学の動向
遺伝学の動向
免疫学の動向
微生物学の動向
分子医学の動向
神経科学の動向
寄生虫学の動向
薬理学の動向
植物科学の動向
物理科学・工学ジャーナル
セルレポート 物理科学
化学
化学触媒
デバイス
ジュール
物質
化学の動向
学際ジャーナル
セルリポーツ・メソッド
セルリポーツ・サステイナビリティ
Heliyon
アイサイエンス
ワン・アース
パターン
STARプロトコル
ネクサス(パートナー)
イノベーション(パートナー)
バイオテクノロジーの動向
認知科学の動向
著者
論文投稿
複数ジャーナル投稿
STARメソッド
プレビュー - プレプリント
査読者
査読者向け情報
ニュース&イベント
ニュースルーム
細胞シンポジウム
コンソーシアムハブ
ウェビナー
ラボリンク
マルチメディア
セルプレスポッドキャスト
セルプレスビデオ
カラーリングとコミック
フィギュア360
セル画ショー
研究篇
セルプレスについて
セルプレスについて
オープンアクセス
COVIDハブ
持続可能性
インクルージョンと多様性
コンタクト
お問い合わせ
ヘルプ&サポート
採用情報
セルプレス採用情報
サイエンティフィックジョブボード
アクセス
登録する
請求
今すぐ読む
司書に推薦する
出版アラート
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
Nucleusコレクション
スナップショット・アーカイブ
インフォメーション
広告主の皆様へ
リクルーターの方へ
図書館員の方へ
プライバシーポリシー
ご利用条件
アクセシビリティ
本サイトのコンテンツは、あらゆる分野の医療従事者および研究者を対象としています。
当サイトでは、サービスの提供・向上およびコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を更新するには、このサイトのクッキー設定をご覧ください。
このサイトのすべてのコンテンツ 著作権 © 2024 Elsevier Inc.
テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビア社に帰属します。
すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条件が適用されます。
プライバシーポリシー 利用規約 アクセシビリティ ヘルプ&サポート お問い合わせ
RELX
この記事が気に入ったらサポートをしてみませんか?