
【高校生物】進化④「進化はどのようにして起こるのか?」

~プロローグ~
フランスの偉大な生物学者キュビエは、生物の分類体系を化石にまで広げ、古生物学を樹立した。しかし、信心深い彼は、進化的な変化(種の変化)を受け入れず、周期的な大激変によって生物が全滅・一新されるという説(天変地異説または激変説)を唱えた。そして、聖書の記述は、最後の大激変の後の事象に適用されるとした(しかし、新しい化石が発見されるほど事態は複雑になり、キュビエの後継者の中には、27回の大激変を仮定した者もいた)。キュビエは、フランス革命、ナポレオン帝国、王政復古と、まさに時代の大激変を生き抜き、高い地位を手に入れた。
キュビエは言う。一度進化論を認めてしまえば「ただ時間と環境さえあれば…ポリプがだんだんとカエルや白鳥や象に変わることになる」。(宗教的な議論は抜きにしても)キュビエが感じたであろう違和感と反感に対して、現代の生物学はどのような説明を与えることができるだろうか。
現在、進化をもたらす要因として、突然変異・自然選択・遺伝的浮動の3つが重要であるということがわかっている(突然変異によって生じた新しい遺伝子の頻度は、自然選択や遺伝的浮動によって変化し得る)。しかし、これらの要因が、具体的にどう関係し合い、どう機能して進化が起きているのかについては、完全には解明されていない。
*遺伝的浮動については次の資料で詳しく学ぶ。
以下の図は進化が起こるイメージ。アルファベットは遺伝子を表す。aは突然変異によって生じた新しい遺伝子を表す。
(1)突然変異によって新しい遺伝子が生じる。
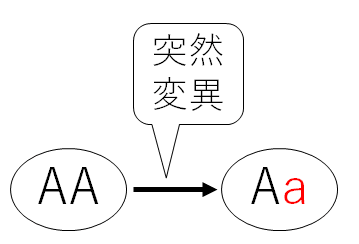
(2)自然選択や遺伝的浮動によって遺伝子頻度が変化する。
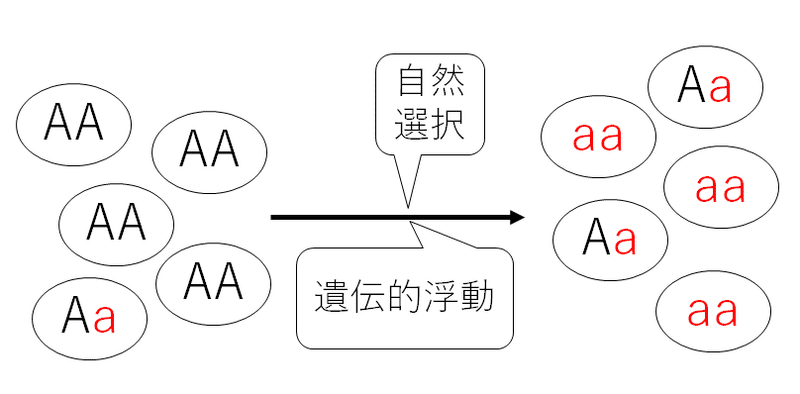
遺伝子頻度の変化こそ進化の定義の一つである。もちろん遺伝子aが後の世代に伝わらず、消える場合も多い(そのような場合がほとんどであろう)。しかし、遺伝子aが固定される(遺伝子aの遺伝子頻度が1になる。すなわち、遺伝子Aがなくなり、全て遺伝子aになる)こともある。
まとめるとこんな感じ。
「人類は常に彼の周りに住む生物の多様性に興味を引かれた。それでこの多様性の意味や原因を知ろうとする試みが多くなされた。この問題は多くの人々にとって制し難い審美的誘惑であった。もともと科学を研究することは一種の審美的努力であるから、この誘惑に応ずることに生物科学の存在理由の一つがあるといえる。」ドブジャンスキー『遺伝学と種の起原』より
★テストに出やすいワード
①突然変異
②自然選択
③収束進化
④共進化
⑤中立説
要点:突然変異によってDNAの塩基配列は変化する。生殖細胞に起きた突然変異は次世代に伝わり得る。
(1)大進化・小進化
● 進化のうち、種の形成(種分化)が起こるような大きな進化を大進化(だいしんか)といい、種分化に至らない進化を小進化(しょうしんか個体群や種内で起こる遺伝的変化など)という。

雑談:大進化には、小進化とは異なる何らかのプロセスが含まれているのか、それとも大進化は小進化の積み重ねなのか、完全には明らかになっていない。
雑談:遺伝学者ドブジャンスキーは、小進化と大進化を連続したものとして見なしている。「小規模の進化は大規模の進化とその原理を異にし、前者は既知の遺伝的原因(突然変異、淘汰、遺伝的浮動 genetic drift)によって理解されるが、大規模の進化は実験的には未知の、あるいは漠然としてわからない動因を含むものと信じている人が少なくない。この種の見解を支持するものは遺伝学者には僅かしかいないが・・・古生物学者や比較解剖学の研究結果に基づいて進化を考える人々の中には相当多い。」「小進化・大進化といっても、まったく比較的のものにすぎないのであって、進化をもたらす動因に関しては、なんら異なることはない。要するにこの区別はただ記載的な意味しかないものである。」ドブジャンスキー『遺伝学と種の起原』
発展:進化の傾向(ウマの進化)
● 化石の記録を調べることによって、進化の傾向を検討することができる。たとえば、5500万年前のヒラコテリウム(別名エオヒップス)の子孫であるウマの進化について調べてみると、一般に、以下のような傾向を見出すことができる。
①体が巨大化した(ヒラコテリウムは犬くらいの大きさだった)。
②指の数が減少した(ヒラコテリウムは前あしには4本の、後あしには3本の指を持っていた。現生のウマには、各足に1本の指しかない[中指だけが発達を続け、残りの指は退化したと考えられる])。下図はイメージ(古今書院『新版地学辞典Ⅲ地質学・古生物学・地形学・土壌学』より 一部改変)。
③臼歯が複雑化した(臼歯のそしゃく面が複雑で大きくなった[ヒラコテリウムは森林で若葉を食べていたが、ウマは草原でかたい草を食べる。臼歯の複雑化にはこのような食性の変化が関係しているとされる])。
*今回は高校の教材に合わせて、あたかも一方向的・一直線的に進化が起こっているように解説している。ただし、現在では、ヒラコテリウムから現生のウマまで一直線的に進化が起こってきたわけではないことが明らかになっている。現生のウマは、多様に分岐した系統樹の中の、唯一生き残った小枝に過ぎないことがわかっている。
*①~③の変化の原因については、完全には明らかになっていない。
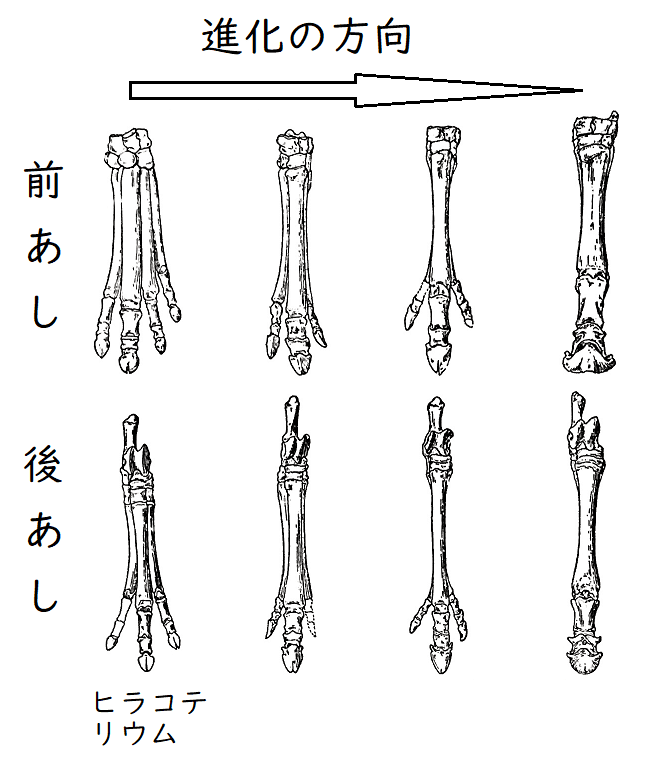
雑談:馬の進化的変遷(たとえば体の巨大化)は定向進化説の好例とされてきたが、アメリカの古生物学者シンプソンは詳細な研究でこれを否定している。
(2)変異(へんい)
● 同種個体間の形質にみられる違い(より正確には「起源を同一にする細胞あるいは個体あるいは集団間にみられる形質の相違」)を変異という。
● 変異には環境変異(かんきょうへんい。生育環境の差などによる変異)と遺伝的変異(いでんてきへんい。遺伝する変異)があり、ふつう、進化に関係するのは遺伝的変異である。
*上で説明した「変異( variation )」と、一般に突然変異と同じ意味で用いられる「変異( mutation )」は違う語。どちらも日本語にすると変異になる。
雑談:環境変異(環境効果ともいう)は、正確には「生育環境の差や発育の途上で起こる偶然的な要因などの影響により、遺伝的に均一な集団内の個体間に生ずる量的変異」のこと。変異の大きさはある値を中心に連続的に分布する。この変異は遺伝しない(たとえば、全く遺伝的に同一な[純系の]種子を2つ用意したとする。それらの種子を、片方は良い土壌へ、片方は悪い[栄養に乏しい]土壌にまく。良い土壌には大きな作物が、悪い土壌には小さな作物が育つ。しかし、両者がつくる種には差が出ない。この2つの作物の育ちの良さの違いは、生育環境に由来する違いであり、遺伝しない)。
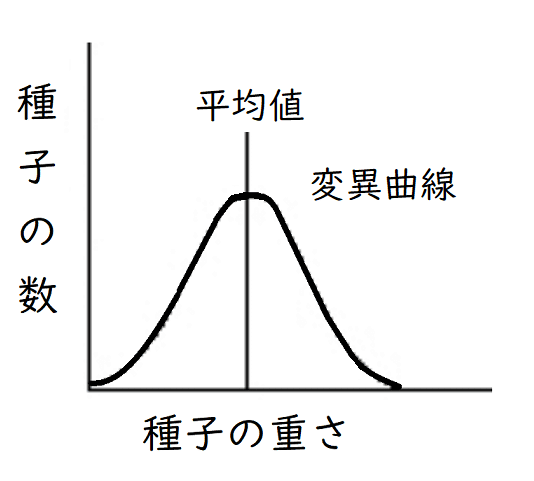
雑談:デンマークのヨハンセンは、自家受精を繰り返し、インゲンマメの純系を作成した。そして、1つの純系の中では、重い種子をまいても軽い種子をまいても、次の代には重い種子~軽い種子まで様々な種子ができ、その平均値は同じになるということを発見した(下図:このようなグラフを変異曲線という)。このように、ヨハンセンは「純系内の個体差は遺伝的変異ではない」ということを証明した。なお、純系のみの集団では選択の効果が無くなり、環境変異の効果だけが残るという説を、純系説という。
*変異曲線:量的に測定できる形質(重さなど)を測り、形質の測定値を横軸に、それに属する個体数を縦軸にして作成したグラフ。
*純系説は、正確には「定量的形質に関わる遺伝子がホモ接合になり純系に至ると、形質の変化は環境因子だけに起因するようになり、選択は無効になるという説」である(純系の集団では、重い種子を選択してまいて育てても、次世代には、また同じように重い種子~軽い種子まで様々な種子が生じる。すなわち、純系の中の「重い」種子、「軽い」種子は、別に遺伝子が異なっているから重さが異なっているのではない。その差は、環境の条件が微妙に異なったことに起因しているのである。もしその差が遺伝子に起因するものだったとしたら、重い種子をまけば重い種子が、軽い種子をまけば軽い種子が多くできるはずである。実際は、両者がつくる種子の平均値は等しかった)。
● 遺伝的変異は突然変異によって生じ、生殖細胞に生じた突然変異だけが次の世代に受け継がれる。
雑談:ワイズマンは「遺伝は、生殖細胞に存在する生殖質(生殖細胞に含まれる要素)が次世代に伝えられる(そして、個体発生では、両親から受け継いだ生殖質が各細胞へ分配されて細胞が分化する)。」とした(このような説を生殖質説[生殖質連続説]という)。
雑談:ワイズマンは体細胞と生殖細胞を区別し、体細胞の影響は生殖細胞には及ばず、したがって次世代には遺伝しないということを指摘した(この指摘は一般に正しい)。また、ワイズマンは、生殖質は生殖細胞の核にあるとした。(ワイズマンは、現代では用いられない用語を使って、複雑な議論を展開しているが)ワイズマンの考え方は、ラマルクの主張した「獲得形質の遺伝」を否定し、さらに染色体に遺伝子があることを予見していた点で、歴史的に意味があるものであった。
(3)遺伝子突然変異と染色体突然変異
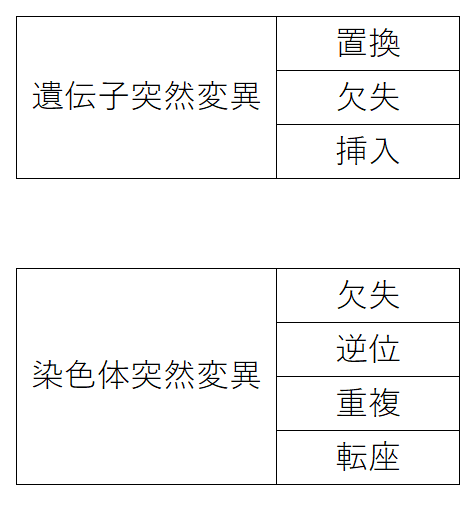
● 遺伝子突然変異(いでんしとつぜんへんい):DNAの塩基配列に置換(ちかん)・欠失(けっしつ)・挿入(そうにゅう)などの変化が生じて起こる突然変異。
例 )鎌状赤血球貧血症(塩基の置換によって、ヘモグロビンβ鎖の6番目のグルタミン酸がバリンに変化している)
雑談:鎌状赤血球貧血症(かまじょうせっけっきゅうひんけつしょう)は、赤血球が三日月状に変形し、毛細血管が詰まったり、溶血したりしてしまう疾患である。致死的な疾患である。しかし、鎌状赤血球貧血症の原因遺伝子と正常遺伝子をもつヒト(ヘテロ接合体のヒト)は、マラリアに抵抗性を持つ(マラリア原虫が赤血球中で繁殖しにくい)ので、アフリカでは、鎌状赤血球貧血症の原因遺伝子の遺伝子頻度が比較的高く保たれている(マラリアは、蚊の吸血によって宿主の血液に入り、まず肝臓に移動するが、やがて赤血球に侵入する)。
*鎌状赤血球貧血症の原因遺伝子と正常遺伝子をもつヘテロ接合体のヒトは、通常は健康であるが、長期間血中の酸素濃度が低下すると鎌状赤血球貧血症の症状が出る。
● 1塩基の挿入や欠失は深刻な影響を及ぼすことが多い。フレームシフト(コドンの読み枠がずれる)が起きるからである。
● 遺伝子突然変異によって終止コドンが生じてしまうと、異常に短いポリペプチドが生じてしまう(よく問題で登場する設定である。問題の途中で、配列に変異が起きたら、必ず終止コドンが生じていないか確認すること)。
● コドンの3文字目が別な塩基に置換しても、同じアミノ酸を指定することが多い(アミノ酸を変化させない置換を同義置換[どうぎちかん]という)。
● 一般に、イントロン領域に置換が起こった場合は、生じるポリペプチドに変化が生じない(イントロン領域はスプライシングで取り除かれるから)。
● タンパク質の重要でない部位に変異が起こった場合、変異が起こってもタンパク質の機能に変化が現れない場合がある。
● 染色体突然変異(せんしょくたいとつぜんへんい):染色体の形や数に異常が生じたために起こる突然変異。欠失(けっしつ。一部が欠落する)・逆位(ぎゃくい。配列順序が逆になる)・重複(じゅうふく。1つの遺伝子座が直列に並んだ二つ以上の遺伝子座に変化する)・転座(てんざ。染色体の一部が他の部分に位置を変える)などがある。
講義動画【遺伝子突然変異と染色体突然変異】
<Q.遺伝子突然変異と染色体突然変異って何が違うの?…注目する視点・規模が異なると考えてよい。突然変異が遺伝子に起きたか(DNAの塩基の変化に注目している)、染色体に起きたか(染色体の構造の変化に注目している)の違いで、本質的な違いはない。>
雑談:「ちょっと間違いを犯すという受容力こそがDNAのまさに驚嘆すべき性質である。この特別な能力が欠けていたら、われわれは今もって嫌気性細菌のままだったろうし、この世に音楽も存在しなかっただろう。」ルイス・トーマス(生物学者)
(4)倍数性
● 一般に、真核生物の個体は、体細胞の染色体数が2nの二倍体であるが、次のように変化することがある。
・異数性(いすうせい):染色体数が2n±αになる。
異数体(いすうたい)の例 :ヒトのダウン症候群(21番染色体のトリソミー:2n+1=47)
・倍数性(ばいすうせい):染色体数が3n(三倍体)や4n(四倍体)などになる。
倍数体(ばいすうたい)の例:パンコムギ(六倍体)
*パンコムギのゲノムについては後述する。
発展:コルヒチン
コルヒチンは、ユリ科イヌサフランの種子や鱗茎に含まれるアルカロイドの一種。コルヒチンには、染色体数を増加させる効果がある。これは、細胞分裂時の紡錘体の形成を阻害することによることが明らかになっている(暗記する必要はないが、コルヒチンは有名なので、たまに問題文に出てくる)。
雑談:コルヒチンの水溶液で成長の盛んな細胞を処理すると、人為的に倍数体を作り出すことができる。生物の染色体を倍加させると、一般に、体のサイズが大きくなる傾向がある。植物の染色体を倍加させることによって、器官の肥大を行おうとする研究が行われている(なお、今日の主要な作物の多くが倍数体である。大型の植物は、食用として望ましい)。しかし、不均一な肥大化など、植物体に様々な障害があらわれてしまい、成功例は少ない。
発展:突然変異説
ド・フリースは、突然変異によって進化が起こるとした(突然変異説)。
発展:人為突然変異(じんいとつぜんへんい)
マラーは、X線を照射することで、人為的に突然変異を起こして見せた。人為的処理により誘発される突然変異を人為突然変異という。
雑談:19世紀後半、物理学の進展により、地球の歴史に対して知見が深まりつつあった。困ったことに、その歴史は生物学者の予想より浅すぎた。生物学者は(進化がダーウィンの言うとおりに進むとしたら)進化の速度があまりに速すぎると考えた。
また、ヨハンセンは、純系内の淘汰は無効であることを示した。純系になると、選択の効果はなくなり、このときに見られる変異は環境変異(生育環境の違いによる変異。遺伝しない)だけになる。このようなヨハンセンの説を純系説という。これは、「淘汰の限界」を示したものとされ、ダーウィンの自然選択説への批判となった。
そこで、何か「突然の、急激な変化」が生物に起こるのではないか?と言う考えをもつ生物学者が現れ始めた。
オランダの植物学者ド・フリースは、アムステルダムの郊外の荒れた草地に、アメリカ原産のオオマツヨイグサを見つけた。ド・フリースは、あるオオマツヨイグサが、他のものと大きく外観が異なる「変わりもの」であることに気付いた。彼はその「変わりもの」を自分の庭に持ち帰って研究し、「変わりもの」に突然変異体( mutant )の名を与え、突然変異説( mutation theory )を提唱した。ド・フリースは突然変異( mutation )という概念を明確にした。mutation はラテン語の「変化」に由来する(ただし、彼がオオマツヨイグサで観察したのは、遺伝子突然変異ではなかった。しかし、突然変異説は、彼の名前を不朽のものにした)。
雑談:突然変異は非常にまれにしか起こらないため、ショウジョウバエの突然変異を研究するためには、何百万匹ものショウジョウバエを飼育する必要があった。アメリカの遺伝学者マラーは、人為的に突然変異を誘発することはできないかと考えた。そして、ついにその方法を発見した。マラーは、X線の照射がショウジョウバエの遺伝子突然変異を150倍も高める作用をもち、人為的に突然変異を誘発できるということを発見した。マラーは次のように語っている。「まさに、ここに素晴らしい約束の地があった。これらの実験の結果は全く驚異的かつ明瞭なものであった。生命の本質ー遺伝子ーが実際に攻撃され、降伏していた。」
雑談:第二次世界大戦の間に、マスタードガスの研究者が、この毒ガスによって起こされた皮膚の障害とX線によるやけどが似ていることを見出した。そしてその後、マスタードガスも突然変異を誘発することが明らかになった(ただし軍事機密のため、すぐには公表されなかった。マスタードガスは最初の化学的突然変異原である)。
要点:自然選択によって遺伝子頻度が変化する。
(1)自然選択(しぜんせんたく)
● ダーウィンは『種の起源』を書き、自然選択説を唱えた。
● 遺伝的多様性をもつ同一種内の個体間に繁殖力や生存力の違いが存在することにより、それぞれの遺伝子型によって子孫を残す割合が異なる現象をを自然選択(自然淘汰[しぜんとうた])という(遺伝子型によって、有利な個体や不利な個体[やどちらでもない個体]が出てくる。有利な個体が増えていく可能性が高くなる)。
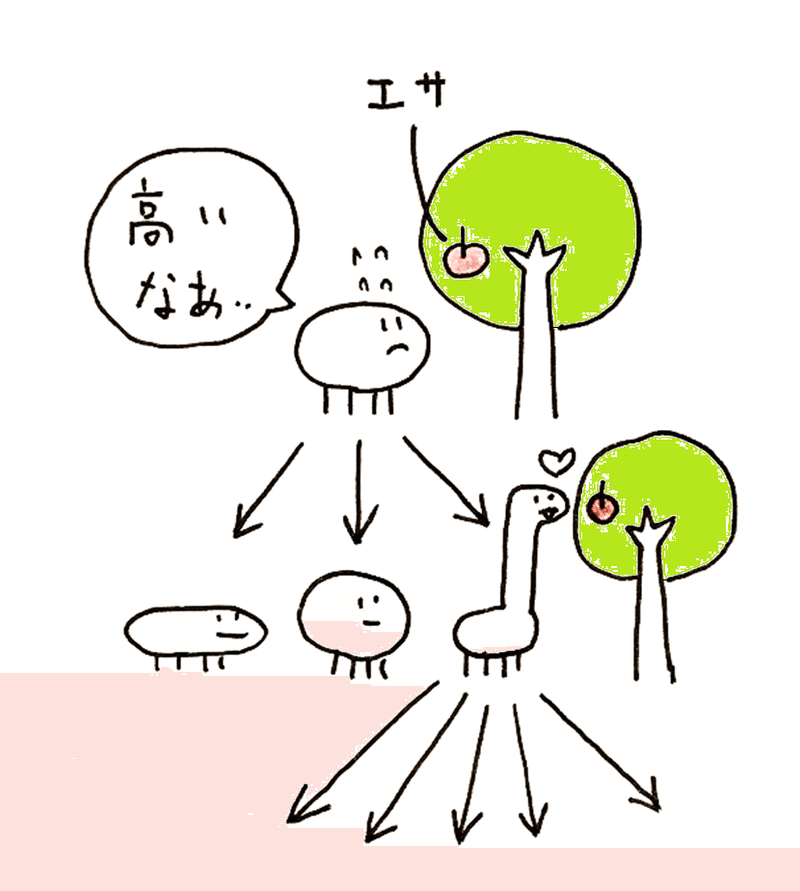
下図は自然選択の大雑把なイメージ。突然変異などが原因となり多様な個体が生じ得るが、その中で、生存に有利な個体(下図では、首が長くなって、エサを簡単にとれる個体)は、より多く子孫を残していく(やがてこの動物は首の長いものだけになるかもしれない)。
雑談:自然選択の少しスマートな定義は「ある遺伝的特性を持つ個体が、それを持たない個体よりも、その特性のために高確率で生き残ったり、繁殖しやすくなったりするプロセス」である。
雑談:「実のところ、淘汰の本質は、異なる遺伝子型を持つ個体が次代の集団の遺伝的構成に、それぞれ異なる程度に寄与するということである。」ドブジャンスキー『遺伝学と種の起原』より
雑談:以下の条件がそろえば必ず自然選択がはたらくと考えられている。①ある形質が個体間で異なる。②その形質が遺伝する。③その形質が原因となって繁殖や生存に個体差が生じる。
雑談:1831年、大学を出たばかりのあるひ弱な青年が、ビーグル号へ乗り込んだ。ビーグル号がガラパゴス諸島にたどり着いた時、その青年は特に魅力を感じなかったようである。地表の熱気は「息苦し」く、「灌木林までが不快な香りがするような気がした」と『ビーグル号航海記』に記した。しかし、どの島も似たような気候であるにも関わらず、動物の各々の変種が別々の島から産出するという事実は、その青年を刺激した。その青年、ダーウィンは、考えを進め、ある仮説にたどり着くことになる。
「わたしは50~60マイルくらいしかへだたっていない島々—―しかも寸分たがわず同じ岩で形成され、まったく同様の気候の下におかれ、ほぼ等しい標高をもつ島々が、ちがう棲み手をもっているとは、夢にも想像しなかったが、まもなくそれが事実であることを見せられたのである。」ダーウィン『ビーグル号航海記』より
*自然選択の例
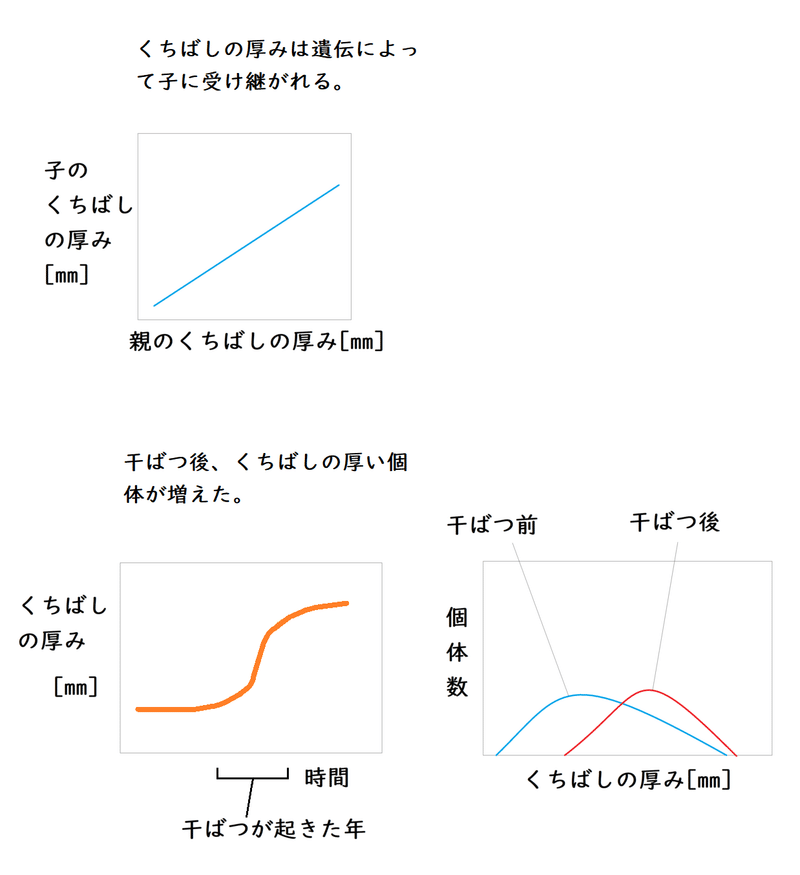
ガラパゴスフィンチにおいて、島に干ばつが起きた後、くちばしが厚くなった(覚える必要はない。そんな例があるのか、くらいに理解しておけばよい)。
1977年の干ばつによって島の多くの植物が枯れてしまったので、ガラパゴスフィンチが食べることができるのは、大きくて堅い果実だけになってしまった。結果、多くのガラパゴスフィンチが死に絶えた。
相対的によく生存したのは、『厚いくちばし』をもつガラパゴスフィンチだった(厚いくちばしにより堅くて大きい物を食べることができる)。
生き残ったくちばしの厚い親から次世代にくちばしの形質が遺伝した(親と子のくちばしの厚みには相関があり、くちばしの厚みは遺伝によって子に受け継がれることがわかっている)。
下の図はイメージ。くちばしの厚みという形質に対して、短期間に強い自然選択が働いたと考えられている。
雑談:ダーウィンは、『種の起源』の中で、品種改良の研究の歴史について論じている。
「すべての品種が、今われわれが目にしているような完全なもの、有用なものとして突然に生じたとは思えないのだ。・・・自然は変異を継起させるだけで、人間がそれを自分の都合のよい方向に積み上げるのだ。」ダーウィン『種の起源』より
我々は、昔から、残したい形質を選んで(つまり、選択を行い)、その形質をもつ個体同士を交配させることなどにより、品種改良を行ってきた(現在の品種の多くは同じ親の子孫であると考えられる)。それと同様のことが、自然や生存闘争によって行われない保証はあるだろうか?
「選抜というこの原理が秘める威力は、仮説の話ではないのだ。」「選抜の原理は人間の手で行う場合にはきわめて有効であるが、この原理は自然界にも適用可能なのだろうか。それは自然界でもきわめて効果的にはたらきうるというのが、私の考えである。」「このように、有利な変異は保存され、不利な変異は排除される過程を、私は自然淘汰と呼んでいる。」ダーウィン『種の起源』より
雑談:南アメリカには、数種類のフィンチが生息している。およそ300万年前に、1種類のフィンチのうちの小さなグループが、大陸の太平洋岸から飛び立った。小鳥たちは1000kmもの旅をして、ある火山群島の島にたどり着いた。この群島は、後にガラパゴスとして知られるようになる。鳥たちは、島の新しい資源を食べながら徐々に特殊化し、新しい種へと分かれていった。ダーウィンがこの島に上陸するまでに、多くの種類のフィンチが群島内の島々に見られるようになった。これらの中には、種子をたべるもの、果実を食べるもの、昆虫を食べるもの、そして、花粉、さらには、血を食べるものまでいた。
講義動画【進化と自然選択】
発展:赤の女王仮説
植物の生産する防衛物質に対して、昆虫が解毒作用をもつように進化する可能性がある。このように、敵対関係にある種の間では、常に進化(変化)をし続けなければ絶滅してしまうというような場合がある。「ある生物種をとりまく生物的環境は、その環境にいる他種の進化的変化などによって、平均的に絶えず悪化している。したがって、自らも持続的に進化していかなければ(すなわち走り続けなければ)、絶滅に至る」という仮説を、赤の女王仮説という。これは、キャロルの童話『鏡の国のアリス』に登場する赤の女王の「同じ場所に留まるためには、力の限り走らねばならぬ」という言葉にちなむ。
発展:性選択(性淘汰)
成体の獲得する配偶者の数の違い(獲得する異性の数の違い[たとえばあるオスの個体が獲得するメスの数の違い])に起因する選択を、性選択(性淘汰)という。鹿の大きな角や、鳥の美しい羽などは、一見、生存に不利な形質であるように思える(邪魔そうだし、目立って敵に見つかりやすくなりそうである)。ダーウィンはこれを、通常の自然選択では説明できない考え、これらは『異性に対する魅力』、すなわち『異性が配偶者を選択するにあたって有効な形質』として発達したものであるとして、性選択の概念を提唱した(ただし、性選択は自然選択の一種とする立場もある)。派手な装飾をもった生物は、確かに外敵に狙われやすくなる。しかし、そのデメリットを上回って、異性に選ばれやすく(つまり、モテるように)なれば、その派手な装飾をつくる遺伝子は広まり得るだろう。
*そもそも、どうして一見生存に不利に見えるような形質を持つ個体が異性にモテる例が多く見られるのかについては、わかっていない(多くの動物において、雄は、とても派手な体をもっている。派手であるほどモテる場合が多い。しかし、本来、派手であることは生存に不利である。敵から見つかりやすくなるからである。そんな不利な形質をもつ個体をメスが選びがちなのは、たしかに一見おかしい。現在も研究が続いている(「生存に不利な派手な形質が進化したのは、そうしたハンディキャップにもかかわらず生存できるほど、その個体の生存力が優れていることを示すのだ。だから異性は、そのようなハンディキャップを持った個体を選ぶようになったのだ。」という、ハンディキャップ説などが唱えられている)。
(2)工業暗化
● 工業暗化(こうぎょうあんか):オオシモフリエダシャクとよばれるガの野生型は白地で明るい色をしている(『明色型』)。そのため、白色の地衣類で覆われた木の幹の上では目立ちにくい。
対して、『暗色型』は暗色であり、煙で黒くなった木の幹では目立ちにくい。
イギリスの工業地帯では、樹が大気汚染によって黒ずんで明色型の個体がよく目立つようになり、明色型の個体が鳥に捕食されやすくなった。その結果、体色の黒い『暗色型』が増加した。これは単に煙に汚れて黒くなったガが増えたのではない。自然選択の結果、体色を暗くする遺伝子の遺伝子頻度が増えたのである(遺伝子頻度の変化は、すなわち小進化である)。以上のような工業暗化の話は、進化(遺伝子頻度の変化)が実際観察された稀な例として有名である。
下図は模型を使ったイメージ。周囲の環境の色によって、有利・不利はコロコロ変わり得る。


雑談:上記の工業暗化について、詳細な研究が進むにつれて、暗色型のガの増加は、鳥の捕食による自然選択だけでは説明できないということがわかってきた。鳥の捕食以外による要因(たとえば紫外線に対する抵抗性など)についても、詳しく調べる必要があるかもしれない。
雑談:生物集団中に遺伝子型の異なった個体が存在し、その間で生存率や繁殖率に「差」がある場合、そのような「差」を引き起こす作用や操作を「選択(淘汰)」といい、この「選択(淘汰)」の作用を物理的な圧力にたとえて「選択圧(淘汰圧)」と言うことがある。たとえば、「木の幹が黒くなったことは、黒色のガの生存率を上昇させる選択圧として働いた」などと言う。
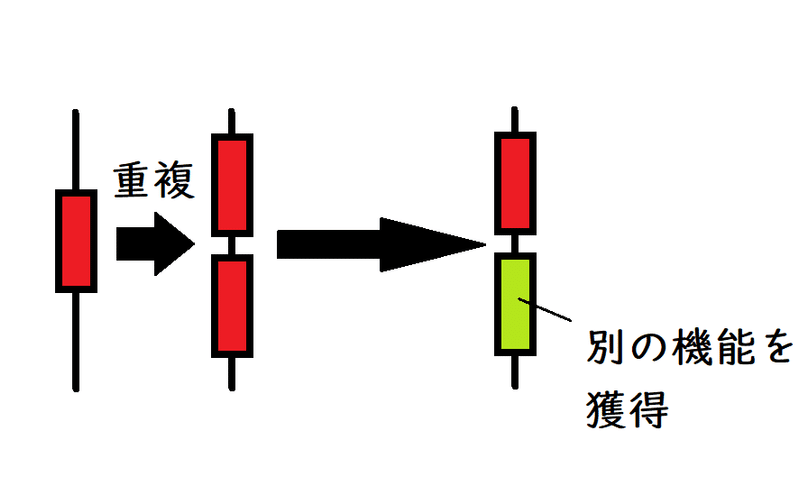
発展:遺伝子重複
同じ遺伝子がゲノム内に重複して存在する場合がある。これを遺伝子重複(いでんしじゅうふく、いでんしちょうふく)という(染色体突然変異や減数分裂時の染色体の乗換えの異常が遺伝子重複の原因になり得る)。遺伝子重複がある場合、一方の遺伝子に突然変異が生じても、他方が補うことがあるため、自然選択に不利とならない。したがって、重複している遺伝子に生じた突然変異は蓄積されやすく、その結果、似た機能をもつ別の遺伝子になっている場合も多い(遺伝子としての機能を失い偽遺伝子になる場合もある)。
例)ヒトの赤オプシン遺伝子と緑オプシン遺伝子の相同性は非常に高い。また、両者はX染色体上に隣接して配置されている。よって、この2つの遺伝子は遺伝子重複によって生じたと考えられる(祖先型のオプシン遺伝子≒赤オプシン遺伝子が重複して2つになった後、突然変異によって緑オプシン遺伝子が誕生したと考えられている。なお、原猿類のアイアイやロリスは青・赤色を認識する2色型色覚だが、ヒトとゴリラを含む狭鼻猿類は青・赤・緑を認識する3色型色覚である)。下図はイメージ。
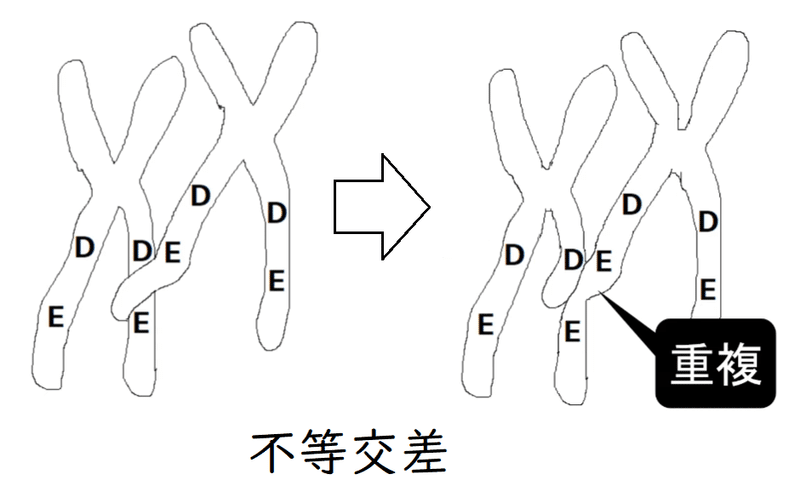
遺伝子重複は不等交差(不等乗換え)によって起きる可能性がある。下図は不等交差のイメージ(相同染色体が、正常な位置から少しずれて対合し、乗換えを起こしたせいで、遺伝子Eの重複が[その隣では遺伝子Eの欠失が]起きている)。
雑談:「ところで、遺伝子重複機構というと体裁がよいが、ひとくちに言って一創造百盗作ということである。」「自然淘汰は、その遺伝子が作る蛋白質のなかの機能的にもっとも大切な活性部位を犯すようなアミノ酸置換を、汲々と排除していかねばならない。・・・新しい機能を得るためには、既存の活性部位を変えなければいけないが、自然淘汰が活性部位の変化を許さないところに問題がある。・・・自然淘汰の極端に保守的な監視眼から逃れたところで、初めて進化の望みが湧いてくる。この自然淘汰の監視から逃れる手は、1つしかない。自身を重複してエキストラコピーを作ることである。そうすると、監視の目はそのなかの1つに集中するから、もう1つは勝手に突然変異によるアミノ酸置換を集積できる。・・・当時は、これはちょっと言い過ぎではないかという友人の方が多かったが、今はすでに、遺伝子重複による進化は常識となってしまった。」大野乾『生命の誕生と進化』より
*大野乾は遺伝子重複が進化の原動力になると考えた生物学者。ジャンクDNAの概念も提唱している。また、ライオンと同時期に、哺乳類の雌でX染色体が不活化することを発見している。
雑談:異なる遺伝子のイントロン領域の間で交差が起こることで、新しい遺伝子が創生される場合がある(新しいエキソンの組み合わせを持った、新規の遺伝子が生まれる。覚えなくてよいが、これをエキソンシャッフリング[エキソンのかきまぜ]という。部品[エキソン]は破壊せずに、その組み合わせを変えることで、新たな器械[遺伝子]を生み出すようなものである)。
雑談:不等交差が起きなくても、染色体の乗換え自体が進化に寄与しているという考えもある。染色体の乗換えは、様々な遺伝子の組み合わせをもつ多様な個体を生じさせる(自然選択がかかる個体群の多様性が高まる)。集団の多様性を高めることは、適応的な進化の速度を速めることにつながると考えられている(自然選択は「多様な個体が生じ得る」ということを前提としている[多様な個体の中で、環境に適応した有利な者が増えていく])。
染色体の乗換えについての解説はこちら↓
*もちろん、そもそもの減数分裂による配偶子形成と配偶子同士の接合によっても、膨大な種類の遺伝子の組み合わせをもった子が生じ得る。たとえばヒトは、23対の染色体それぞれから1本を選んで配偶子に入れる。その選び方は2の23乗ある(2本ある相同染色体のうちのどちらか1本を配偶子に入れる。それが[性染色体も含めれば]23対分ある)。両親で同じことが起こっているので、2の23乗種類の卵と、2の23乗種類の精子が生じ得る。それらが受精するとすると、できる受精卵の種類は(2の23乗)×(2の23乗)=約70兆にもなる。実際は減数分裂時に組換えが起こるわけだから、もっと多様な配偶子ができ、もっと多様な子が生じ得る。
配偶子の種類については、以下の資料で少し詳しく解説している。
(3)収束進化
● 収束進化(しゅうそくしんか):系統の異なる複数の生物が、類似する形質を個別に進化させることを収束進化(収れん進化、収れん)という。
*相似器官が見られるのも、収束進化の結果である。
雑談:フクロモモンガ(有袋類)とムササビ(真獣類)は非常によく似ている。これは収束進化の結果である(両者は見た目が非常に似ているが、塩基配列は似ていない[近縁なわけではない]。フクロモモンガとムササビの滑空する能力は、はるか遠く離れた2つの哺乳類で独自に進化したと考えられている)。
雑談:収束進化は、進化における"他人のそら似"にたとえられる。
(4)擬態(ぎたい)
● 擬態=周囲の環境や他の生物と見分けがつきにくくなること。擬態は、自衛や攻撃に有利になるような自然選択の結果と考えられる。
雑談:擬態にはいくつか種類がある。
①隠ぺい擬態(いんぺいぎたい。コノハチョウは木の葉に似た姿となって、攻撃者から発見されないようにしている)
*動物個体の存在を、背景の中に埋没させてしまうような効果を持つ体色を隠ぺい色という。特に、被食者がもつ隠ぺい色を保護色(ほごしょく)という(ただし、捕食者の隠ぺい色も保護色と呼ぶ場合がある)。
②攻撃擬態(こうげきぎたい。ハナカマキリは姿を花に似せ、被食者を待ち伏せする)
③ベイツ型擬態(べいつがたぎたい。トラカミキリは、アシナガバチに擬態している=毒を持つ生物に見られる警告色で捕食されないようにしている)
④ミュラー型擬態(みゅらーがたぎたい。ハチ類は、お互いに似た体色を持ち、捕食のリスクを少なくしている)
*ベイツ型擬態・・・他の動物に捕食されやすい味の良い種類の動物が、不味で警告色をもつ他の動物に姿を似せることで捕食者を欺いている。
*ミュラー型擬態・・・2種以上の動物のもつ警告色が、相似た斑紋や色彩に収束進化し、それにより未経験の捕食者に食われる率を低めている。ハチの多くの種類が同じく黄と黒の模様をもつなどがその例。
(5)共進化
● 共進化(きょうしんか)=異なる種が相互的に作用しながら進化すること。
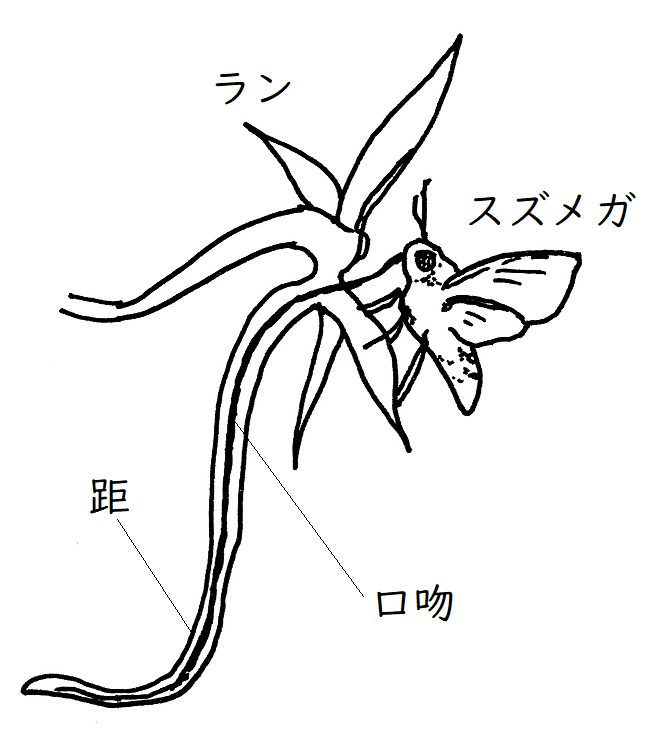
例)ランの距(きょ)と、その蜜を吸うスズメガのストロー状の口吻(口器の一部)は、ともに長くなるように進化してきた(蜜を貯めるランの距が長くなると、スズメガに花粉を付着させやすくなる。それに合わせてスズメガの口吻の長いものが自然選択される[口吻が長い方が蜜が吸いやすくなり有利になる]。これを繰り返した結果、ランの距もスズメガの口吻も長くなったと考えられている)。
*距:花弁の基部から突出した細長い筒状の構造で、奥に蜜をためている。
雑談:ダーウィンは長い距を持つ花を見て、28cmの長い口吻をもつガの存在を予測した。そのようなガは、ダーウィンの死後20年経てから発見された(もし、花粉を付着させることなく、蜜を飲むのに十分な長さの口吻をもつ虫がいたとする。その場合、虫は花に接触することなく蜜だけを吸うことができる。虫に花粉が付着しないので花粉は運ばれず、植物にとって不利である。よって自然選択は、より長い距をもつ植物に有利に働くだろう。一方、虫も、効率よく蜜を得るためには、長い口吻をもつほうが有利である[短すぎる口吻では蜜を得られない]。このような関係があるので、距と口吻はともに長くなるように進化[共進化]してきたらしいと考えられている[まるで形質の軍拡競争である])。
雑談:スズメガは高い飛翔能力をもち、優れた送粉者となっている。夜活動するガの天敵はコウモリである。様々なガで、コウモリに対抗するための特性が発見されている(コウモリの発する超音波をとらえるための耳が発達していたり、自身がコウモリの探索を阻害する超音波を出していたりする)。スズメガの高い飛翔能力も、コウモリとの攻防の結果進化してきたものと考えられている。
発展:イチジクとイチジクコバチ
イチジク類では、雌花と雄花(咲くタイミングは異なる)が「花のう」と呼ばれる袋状の構造の内側に形成される(袋状の構造の内側に雌花と雄花があるので、雌花や雄花は外からは見えない。「イチジク」は漢字で書くと「無花果」である)。
このままでは(袋の内側に花が咲いていては)イチジクは花粉を散布することができない。
しかし、イチジクコバチとの巧妙な共進化の結果、子孫を残すことができている。
①花粉を携えたイチジクコバチの雌が、花のうの上部に空いた隙間から侵入する。
②イチジクコバチの雌は花のうの内側に咲いた雌花の雌しべに花粉を付けて回る。そしてそれと同時に、雌花の子房に卵を産み付ける。ただし、雌花には雌しべの長さが長いものと短いものがあり、(産卵管が届く)雌しべの短い方の雌花の子房にしか卵を産み付けることはできない。
*もし、すべての雌花で産卵が行われれば、(これから生まれてくるイチジクコバチの幼虫にすべての胚珠が食べられてしまって)イチジクは種子を残すことができなくなるだろう。イチジクコバチに食べられなかった分が、イチジクの取り分になるのである(イチジクは、何割かの雌しべを短くすることで、イチジクコバチに餌となる胚珠を提供している。その代わりとして、イチジクコバチに花粉を運んでもらうのである)。
③卵から孵化した幼虫は胚珠1個を消費して成長する。
④先に翅がない雄のイチジクコバチが羽化する。彼らは花のうの中を徘徊し、遅れて羽化する雌のイチジクコバチと交尾する。また、雄のイチジクコバチは、花のうに穴をあける(これが雌のイチジクコバチの脱出口となる)。
*なお、雄のイチジクコバチは、花のうの中で一生を過ごす。
⑤交尾が済んだ雌のイチジクコバチは、遅れて成熟したイチジクの雄花の花粉を携えて、産卵のため、次の花のうに向けて飛び立っていく(花のうの成熟に伴い、上の隙間はふさがってしまっているので、雌バチは、雄バチの空けた壁の穴から花のうの外に出る)。
→再び①へ。
*イチジクの雌花と雄花の成熟の時期はずれている。雌のイチジクコバチが⑤で花のうから出ていく時期に合わせて雄花が咲くようになっている。
*③のように、イチジクコバチの幼虫は、熟した胚珠を食べて育つ。したがって、イチジクコバチにとって、確実に花粉を運び、イチジクの胚珠を成熟させることは、自身の子孫を残すために必要である。
*イチジクコバチの多くの種は、花粉ポケットと呼ばれる花粉をためる特殊な構造をもっている。羽化して交尾を終えた雌は、前脚を使って積極的に花粉ポケットに花粉を集める。そして、羽化した花のうを飛び立って新しい花のうに入ると、今度は花粉ポケットにためた花粉を使って雌花を受粉させていく(送粉者自身が能動的に花粉を運ぶ、驚くべき適応である)。もちろん、イチジクは、イチジクコバチに花粉を運んでもらうことで子孫を残すわけだから、イチジク側もメリットを得ている(イチジクとイチジクコバチは相利共生の関係にある)。
下図はイチジクの花のうのイメージ。
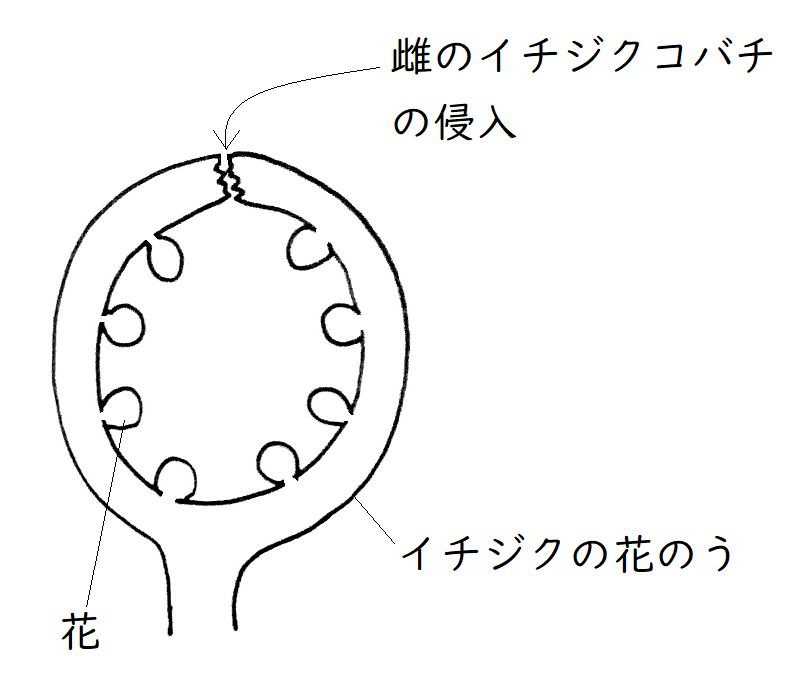
②イチジクコバチの雌は花のうの内側に咲いた雌花の雌しべに花粉を付けて回る。そしてそれと同時に、雌花の子房に卵を産み付ける。
③卵から孵化した幼虫は胚珠1個を消費して成長する。
④先に翅がない雄のイチジクコバチが羽化する。彼らは遅れて羽化する雌のイチジクコバチと交尾する。また、雄のイチジクコバチは、花のうに穴をあける
⑤交尾が済んだ雌のイチジクコバチは、イチジクの雄花の花粉を携えて、産卵のため、次の花のうに向けて飛び立っていく
→再び①へ。
(6)示準化石・示相化石
● 示準(しじゅん)化石:ある特定の時代にかぎって産出される化石で、年代決定に用いられる。 三葉虫(さんようちゅう)、フズリナ、筆石(ふでいし)、ウミユリは古生代の、アンモナイトは中生代の、貨幣石(かへいせき)、マンモス、ビカリア(巻貝の仲間)は新生代の示準化石である。
雑談:筆石(ふでいし)類(graptolite:ギリシャ語の「描かれた・文字が記されている」に由来)は、カンブリア紀中期から石炭紀前期に生存した化石動物(古生代の示準化石)。特にオルドビス紀、シルル紀に大繁栄した。原索動物に近い半索動物に属する。筆石の化石がこの名前で呼ばれているのは、硬い頁岩に残された痕跡が石版筆の跡に似ているからだとも、また羽根ペンの形に似ているからだとも言う。
雑談:フズリナ(fusulinid)は紡錘虫とも呼ばれる。古生代後期の石炭紀とペルム紀に繫栄した原生生物・有孔虫類の絶滅分類群の総称(およそ5000種が知られている)。紡錘・円盤・球状の石灰質の殻を持つ。石灰質の殻の壁に発達する隔壁、孔、その他の構造の詳しい研究によって系統樹が組み立てられており、形態進化が著しく(一般に、内部構造が単純なものから複雑なものへと形態が変化している)、汎世界的な分布のために、重要な示準化石となっている(サンゴのような浅海性の化石とともに産することが多く、示相化石としても価値が高い)。
雑談:ウミユリは棘皮動物(植物ではない)。オルドビス紀に出現し現在にいたる(現生種は650種ほど)が、古生代では特に主要な海生動物の一つとして大繁栄していた。
雑談:アンモナイト(ammonite)は、広義には、デボン紀前期に出現し、白亜紀末に絶滅した化石頭足類の一群を指す。狭義には、ジュラ紀・白亜紀のアンモナイト目(Ammonitida)を指す(これが示準化石に用いられる)。
雑談:貨幣石(ヌムリテス)は原生生物・有孔虫類の1科Nummulitidaeに属する絶滅した大型の底生有孔虫類(もちろん石ではなく生物である。その形態から貨幣石と呼ばれている)。エジプトのピラミッドなどの石材にも含まれる。
雑談:今から1500万年前(新第三紀)の日本には、熱帯の海が広がっていた。内湾の岸辺や河口付近にはマングローブが生い茂り、海底にはたくさんのビカリア(腹足類)が生息していた。
雑談:現代でも、東南アジアやオーストラリアのマングローブの浅海には、ビカリアに近い仲間のセンニンガイが多く見られる。
● 示相(しそう)化石:その当時の環境を示す化石を示相化石という。サンゴの化石は、その環境が当時暖かい浅い海であったことを示している。
雑談:ほかにも、ブナの葉の化石は、やや冷涼な気候であったことを示すと考えられる。
雑談:化石を、示準化石や示相化石として、科学に用いることができるということは、我々にとっては当たり前の話である。しかし、昔は当たり前ではなかった。化石は宗教的な議論に利用されてきた。1726年に、博物学者ショイヒツァーはノアの洪水の証拠として、洪水以前の人間の標本(溺死した証人)を提示した(しかし、それは実際は、イクチオサウルス[魚竜類]の化石だった)。化石から祖先の大きさを算定し、アダムとイヴの身長を数十mと推定した人もいた。化石は、創造主による仕事場の屑、途中で放棄された計画であるとする人もいた(ただし、そのような熱心な化石の収集・観察が、古生物学の発展、そして進化論につながっていったということも事実である)。
雑談:「化石はこの惑星の記憶である。地球は化石で岩石に自叙伝を綴っているのだ。」スミソニアン国立自然史博物館 館長 カーク・ジョンソン
要点:隔離によって種分化が起こる。
(1)地理的隔離
● 地理的な障害などにより同種の集団の間で自由な交配ができなくなったとき、これを地理的隔離(ちりてきかくり)という。地理的隔離が種分化を促すと考えられている。
雑談:厳密には、地理的隔離とは「海峡など生物の移動を妨げる外的障壁によって生じる、集団間の遺伝子の移動の減少あるいは阻止」を指す。
雑談:「隔離」とは、「集団間の遺伝子交流が何らかの要因により妨げられること」をいう。
(2)生殖的隔離
● 地理的隔離などが原因となり、交配が行われず子孫を残せなくなった状態を生殖的隔離(せいしょくてきかくり)という(もしくは生殖隔離という)。生殖的隔離は新しい種が形成された状態である。
雑談:厳密には、生殖的隔離とは「集団内の遺伝的な基盤をもつ差異によって、その間の遺伝子の移動が著しくあるいは完全に制限されていること」を指す。生物学的種の概念においては、生殖的隔離の有無が同種か別種かの判断基準とされる。たとえば(極端な例だが)、ヒトとチューリップは交配して子を残すことはできない。それは、地理的隔離が起きているからではない(ヒトとチューリップの間に山脈とか海峡とかがあって交配できないわけではない)。遺伝的な基盤に基づく原因で(体の構造が異なることが原因で)、同じ場所に暮らしていても、交配不可能なのである。無論、ヒトとチューリップは別種である。
(3)生物学的種(せいぶつがくてきしゅ)
● 種の定義は色々あるが、「その集合の内部で、交配が可能な子孫をつくり続けていくことができる個体の集合」を種(生物学的種)とすることが多い。
雑談:「猫からライオンが生まれることも、またライオンから猫が生まれることも決してない。したがって種とは単なる分類上の群とか範疇とかに止まるものではない。同時にそれは、共通の形態的特性を具えていると否とにかかわらず、個体を超えた生物学的実在である。」ドブジャンスキー『遺伝学と種の起原』より
雑談:厳密には、生物学的種は"自然条件下で実際に、あるいは潜在的に交配し、ほかの群とは生殖的に隔離されている生物の一群"である。異所的な集団(異なる場所にいる集団)あるいは異時的な集団(生殖時期が異なる集団など)に関しては、生殖的隔離の存在を直接検証することができないため、様々な間接的な証拠に基づいて生殖的隔離の有無を推測するのが一般的である(なお、生殖的隔離の程度は、しばしば連続性を示すので、同種か別種かの区別は絶対的なものではない。また、もっぱら無性生殖を行う生物に対しては生殖的隔離の概念を適用できない)。
雑談:歴史的には、種の区別は、形態的な観察(見た目)に基づいて行われてきた。形態の似た(見た目が似た)生物集団を一つの単位と見なし、固有の名前が与えられてきた。このような(ある意味"素朴"な)種の概念を形態的種概念という。形態的種概念は、種として区別する基準があいまいで、主観的である。
雑談:たまに、生物基礎の問題で、「次の生物のうち、最も近縁なものを選びなさい。」とか言って、「似た形」の生物のイラストを選ばせる問題がある。これはとんでもない話で、形態が似ているからと言って近縁であるとは限らないし、形態が似ていなくても同種であるということはあり得る。たとえば、アシナシイモリとヘビには一般に四肢がなく似ているが、アシナシイモリは両生類、ヘビはハ虫類である。クジラとマグロは似ているが、クジラは哺乳類、マグロは魚類である。
(4)種分化(しゅぶんか)
● 地理的隔離による種分化のイメージは以下の通り。
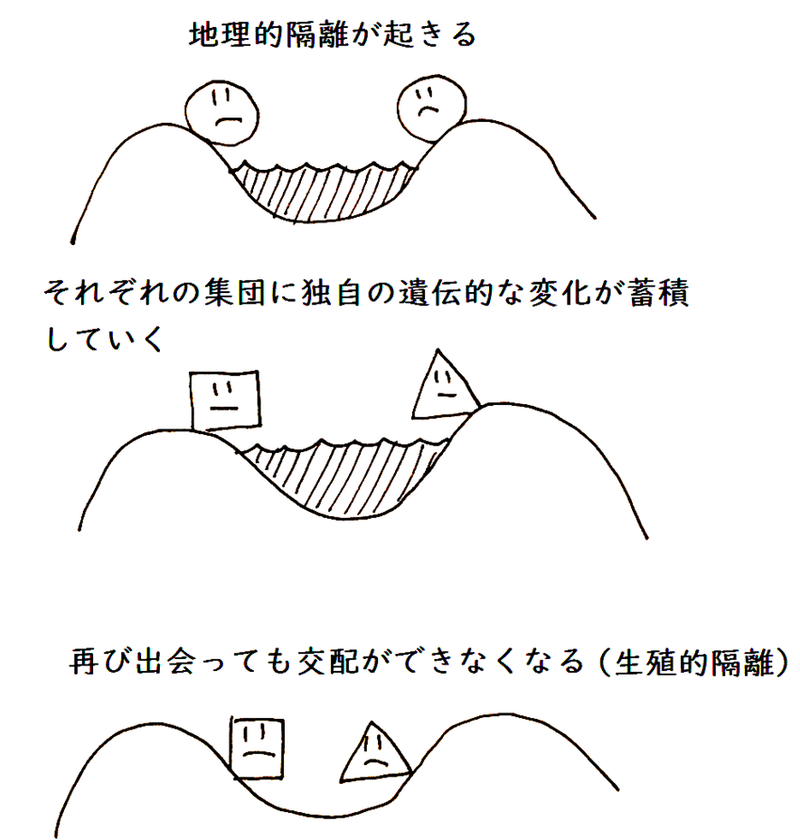
雑談:ワグナーは、上記のように、種を分化させるものはまず地理的隔離だと考えた(隔離説)。
● 地理的隔離がきっかけとなって起きる種分化を異所的種分化(いしょてきしゅぶんか)という。
● 隔離は、地理的でない場合もある。たとえば、性成熟の時期の違いや、鳴き声の違いにより隔離が生じ得る。そのような違いを引き起こす突然変異をもつ集団ともたない集団との間に、生殖的隔離が生じ、種分化が起こることもある。このような、「地理的隔離を伴わずに」起きる種分化を同所的種分化(どうしょてきしゅぶんか)という。
雑談:「おそらく隔離機構は一回の突然変異によってでき上がるものではなく、複雑な補足遺伝子系が築き上げられることによって生ずるものであろう。仮に或る集団が aabb という遺伝子構成を持ち、 a および b がそれぞれ単一の遺伝子もしくは遺伝子群であると考え、なお、この集団が二つの異所性の、すなわち地理的に隔離された部分に分かたれるものとする。そして、一方では a が A に突然変異して地方品種 AAbb を生じ、他方では b が B に変わって aaBB という品種になったとする。この場合 aabb, Aabb および AAbb という構成の個体は自由に相互交配を行うから、A遺伝子が集団中に固定されることは困難ではない。また aabb, aabB, aaBB も相互に交配できるから、B遺伝子も同様に固定される。しかし AAbb×aaBB という交雑は、 A と B との相互作用により一種の生殖的隔離機構が生ずるから、まったく起こらないかはなはだ起り難い。そしてたとえ AAbb および aaBB 遺伝子型を持つものがそれらを隔てている障壁を乗り越えることがあっても、両群はもはや相互に交雑することなく、同一地内に共存し得るようににあるであろう。」ドブジャンスキー『遺伝学と種の起原』より
要点:中立の突然変異は、遺伝的浮動によって集団内に広がる。自然選択だけでなく、遺伝的浮動によっても遺伝子頻度は変化する。
● 中立説(ちゅうりつせつ):「突然変異の多くは自然選択に有利でも不利でもない中立的なものであり、中立的な突然変異は遺伝的浮動(いでんてきふどう。偶然により遺伝子頻度が変化すること)によって集団内に広がる」という考え方。木村資生(きむらもとお)によって提唱された。
雑談:「「中立説」という語は、「分子レベルにおける進化的変化と多型は、主に、自然淘汰に関して、ほとんど中立でその行動と運命が主として突然変異と偶然的浮動によって決定されるような突然変異遺伝子によるものであるとする説」の省略であることを理解しておいていただきたい。」木村資生『分子進化の中立説』より
*木村資生は、『分子進化の中立説』の中で、自身の中立説を「分子進化の中立突然変異浮動仮説」と言い換えている。また、「「中立説」という言葉がすでに広く使われており、途中で変えない方が良いように思う。」とも語っている。
講義動画【中立説】
<Q.中立説って?…厳密に学ぶと難しい。上に書いたように「突然変異の多くは自然選択に有利でも不利でもない中立的なものであり、それが遺伝的浮動によって集団内に広がるという考え方」とだけ知っておけばよい。もともとは、ダーウィン進化論では説明できないアミノ酸の変異速度を説明するために木村資生が考えた説である。もし、有利な突然変異のみが進化の原動力だと言うのなら、同義置換の頻度が非常に多い理由が説明できない(同義置換が非常に有利であるとは思えない)。また、住んでいる環境の大きく異なる動物種(たとえば海に住む動物と陸に住む動物)の持つそれぞれのヘモグロビンを解析した時に、どちらにも一定の確率で変異が起こっているように見える理由が説明できない(環境が大きく違うのだから、変異の速度に違いが出てもよいはずである)。それに、常識的に考えて、環境条件が一定ならば、有利な突然変異の出現は、さらなる有利な突然変異の出現する確率を減ずるはずである[改善は、さらなる改善の余地を減らすだろう]。なお、中立説は、自然選択を否定していないことに注意(正の自然選択は無視できるほどにまれであるとは主張しているが、自然選択説を完全に否定する説ではない)。>
● 中立説は、自然選択を否定するものではない。自然選択が起きることは認めている。
雑談:表現型レベルでの、進化に関する様々なことがら、たとえば、眼の形態や、行動、生態の進化を扱う研究分野では、未解明な部分が多い。
雑談:進化において、正の自然選択が働かずに中立突然変異が次々に固定してゆくことで、遺伝子や形質の時間的変化が生じることを、中立進化という。
問題:ある遺伝子座の対立遺伝子頻度が世代を経るにしたがって偶然上昇したり下降したりすることを何というか。
答え:遺伝的浮動(「偶然」がキーワード)
要点:寒い地域に住む動物ほど突起が小さく、体が大きい傾向がある。
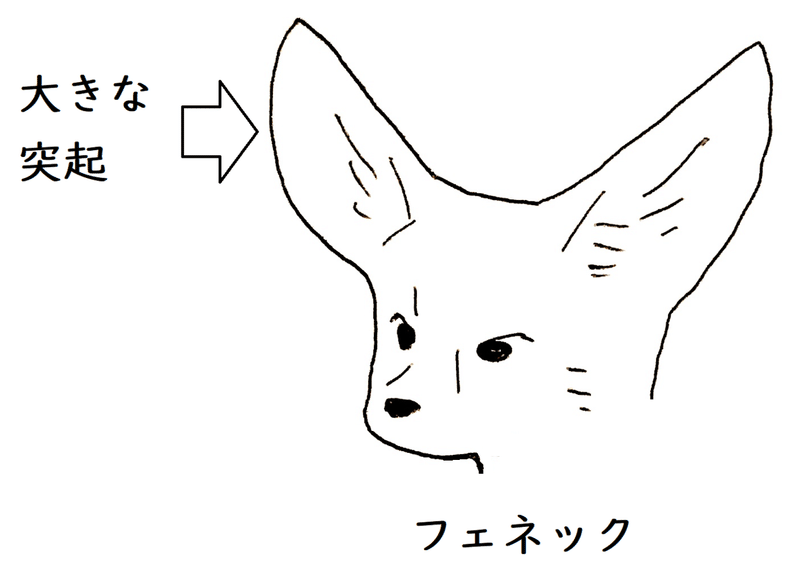
(1)アレンの規則
● アレンの規則:寒冷な地域に住む恒温動物ほど突起部が小さいという傾向(同じ種の個体あるいは近縁の異種の間には、寒冷な地域に生活するものほど、耳、肢、翼、尾などの突起部が短くなるという傾向がある)。
例)ホッキョクギツネの耳より、フェネック(砂漠などに住む)の耳の方が大きい。下図は大きな耳をもつフェネック。
(2)ベルクマンの規則
● ベルクマンの規則:寒地に生息する恒温動物ほど体が大きくなるという傾向(同じ種でも、寒冷な地方に生活する個体の方が温暖な地方に生活する個体よりも体重が大きい傾向がある。また、近縁な異種間では、大形の種ほど寒冷な地方に生息する傾向がみられる)。
例)ホッキョクグマはマレーグマ(熱帯雨林などに住む。世界最小のクマとも言われる)より大きい。下図は大きな体をもつホッキョクグマ。

● 熱は体の体積部分で作られ、表面から逃げていく。体積が大きいほど熱産生の程度が大きくなり、表面積が大きいほど放熱量が大きくなる。動物の体を球だとすると、体積(熱産生)は半径の3乗に比例し、表面積(放熱)は半径の2乗に比例する。したがって、寒い地域では、体が大きい(半径が大きい)方が有利ということになる。
*ただし、アレンの規則、ベルクマンの規則には例外も多い。
要点:相同器官は適応放散の結果であり、相似器官は収束進化の結果である。
● 外観やはたらきが異なっても、同じ発生起源をもち、同じ基本構造をもつ器官を相同器官(そうどうきかん)という。相同器官が見られるのは、共通の祖先をもつ生物がさまざまな環境に適応して多様化(適応放散[てきおうほうさん])した結果であると考えられる。
相同器官…ヒトの手とコウモリの翼、魚類のうきぶくろと両生類の肺、ふつうの植物の葉とサボテンのとげなど
● 相似(そうじ)器官:発生上の起源は異なるが、外観やはたらきが『よく似ている』器官を相似器官という。
相似器官…昆虫の翅(表皮から分化した突起)と鳥類の翼(前肢が変化したもの)、エンドウの巻きひげ(葉)とブドウの巻きひげ(茎)、サツマイモのいも(根)とジャガイモのいも(地下茎の変化した塊茎)、バラのとげ(茎)とサボテンのとげ(葉)など
*相似器官は、似てるだけという意味。たいてい似てるものを選べば相似器官である。出題者は「こんなに似ているのに、起源が異なるんだよ」という驚きを受験生に伝えたいのである。
● ヒトの尾骨や虫垂などのように、退化してほとんど機能をもたない器官(痕跡的に留まる器官)を痕跡(こんせき)器官という。
雑談:進化の過程で、ある器官の退化が進むと、最終的にはその器官は消失に至ると考えられている。痕跡器官はその途上にあると考えられる。
雑談:虫垂は、哺乳類の盲腸先端部にある大腸の一部である。虫垂は草食動物では盲腸とともによく発達するが、ヒトや類人猿では萎縮する。草食動物では、虫垂の中に大量の腸内細菌が存在し、セルロースの代謝に非常に重要な役割を果たしている。ヒトの虫垂には重要な生理作用・機能はないとされる(ただし、ヒトにおいて、虫垂で発達しているリンパ組織が腸内の免疫機能の維持に重要な役割を果たしているという考えもある)。
発展:ラマルクの用不用説
(今はほぼ問われない)
ラマルクは『動物哲学』の中で用不用説を提唱した。
不用説:生物にとって必要で使用する頻度の高い器官の大きさや機能は発達するが、使用しない器官のそれらは退化するという考え。
たとえば、用不用説の立場をとれば、キリンの首が長い理由は、以下のように解釈できる。
「キリンの首は高い樹木の葉を食べるために背伸びし続けた結果である。」これはもちろん誤った説である。
用不用説の背景には獲得形質は遺伝すると言う考えがある(「用」とは、積極的に器官を使用するということである。各世代のキリンは、積極的に首を伸ばそうとした。その世代世代の努力の積み重ねにより、首が長くなったと考えるのが用不用説である)。
現在では、獲得形質は遺伝しないことがわかっている(生殖細胞に起きた突然変異が遺伝する)。たとえば、あなたが、一生のうちに、トレーニングをして、後天的にマッチョになったとしても、それ(マッチョであること)は子に遺伝しない。

雑談:「その変異は遺伝することが、自然淘汰が作用する上では不可欠である。」ダーウィン『種の起源』より
雑談:「ダーウィンに比べて、ラマルクは大したことないな。キリンが各世代の努力で首を長くしたわけないことは、小学生でも知っているよ。」と思うだろうか?
確かに、高校生物の教材はラマルクを上記のようなイメージで扱ってきた。
しかし、進化学の歴史を変えたのは彼である。
多くの生物学者が、種を、神が創った不変のものと見なしている中、ラマルクは、明確に環境と進化を結び付けている。ラマルクはこれをダーウィンより50年はやく見出していた。
ラマルクは不遇のまま死んだが、ラマルクの娘コルネリは「後の世の人がきっとほめてくれるでしょう。そしてお父さまの仇をとってくれますよ」と言ったという(イヴ・ドゥランジュ『ラマルク伝』より)。
ラマルクは『動物哲学』の中で次のように述べている。
「生物の間の種は、相対的な安定性しか有せず、かつある時期の間だけ不変であるに過ぎない。」
また、ラマルクは、『動物哲学』の中でキュビエらの唱えた天変地異説を批判している。
「大部分の動物が時の推移とともに変化を受け得るということを認めなかった博物学者たちは、観察された化石並びに地表のさまざまな地点に認められる変動に関する事実を説明しようとして、地球に全世界的変災が発生し、・・・当時存在していた種は大部分絶滅したと想定した。・・・何を苦しんで証拠もなしに、世界的大変異などというものを想定する必要があるのか。」
当時は、神の創造した種は不変であるという説が主流であった。たとえば、分類学の父とも呼ばれるリンネすら「最初に創造されただけの種が存在する」と言っている。キュビエは、種が不変であるということと、不思議な形をした化石の存在に整合性を持たせるため、定期的に天変地異が起きたと仮定した。キュビエは、現在の生物は、人を含め、最新の天変地異が起きた後につくられたと考えた。信仰心の強いキュビエは、最後まで進化論を認めず、ラマルクに対して痛烈な批判を続けた(キュビエほどの偉大な科学者が、進化論だけは頑なに認めなかった)。ちなみに、ラマルクは、全生物に共通する法則性の探究が必要だとして、「生物学(Biology)」という語を提案した。
講義動画【生物学史】
(動画の「系統・進化」のところで少しだけ進化学の歴史について話している。すべて雑談である。)
発展:ヘッケルの提唱した反復説
動物学者ヘッケルは、「個体発生は系統発生を繰り返す(受精卵から誕生までの発生過程は、その生物がたどってきた進化の過程を短縮して再現している)」と考えた(この考えを反復説[はんぷくせつ]という)。以下の①、②が反復説の根拠として有名である。
①哺乳類の胚に鰓裂(鰓孔ともいう。魚類などではこの部位に鰓が形成される)のような構造がみられる(魚類から哺乳類が進化した証拠とされた)。②鳥類の窒素排出物がアンモニア→尿素→尿酸と変化していく(鳥類が魚類から両生類を経て進化した証拠とされた。魚類はアンモニアを排出し、両生類の成体は尿素を排出する)。
雑談:ヘッケルの反復説には例外も多く、観察結果の一つの解釈に過ぎないとも言われている。発生初期には自然選択が強くかからないため、他種の生物の胚が似ているのは自然であるとする考えもある。ちなみに、人類学者・社会学者であるベ―ツソン(連鎖を発見した遺伝学者ベーツソンの息子)は以下のように論じている。
「胚の発生のより早い時期に衝撃を与えることは、それだけ長く複雑な出来事の連鎖をかき乱すことに等しい」「道筋の初期段階で逸脱する方が、後の段階で逸脱するよりより難しい(成功の確率が低い)」「仮に進化のエンジニアになったとして・・・個体発生のあまり早い時期に強く介入することは避けた方が賢明だ」ベ―ツソン『精神と自然』より
発展:定向進化説(ていこうしんかせつ)
動物学者アイマ―や古生物学者コープは、「進化において、形質の変化は一定の方向をとる」とする定向進化説を提唱した。たとえば、ウマは、体が大型化するような進化をしてきた。また、指は中指のみが発達を続け、残りの指は退化した(他にも、マンモスの牙の巨大化などが根拠として挙げられることがある)。しかし、一定方向への進化を誘導する内的要因の具体的な機構が説明されていないこと、進化は直進的ではなく分岐をもつことが普通であることなどから、定向進化説は現在ではほとんど支持されていない。
雑談:定向進化説の議論では、何らかの神秘的な要因が関わっているかどうかが問題とされてきた。アイマーらが、物理的・化学的過程を考えたのに対し、一部の定向進化論者は、「完成化の力」なるものを想定し、生物の発達が一定の方向へ導かれると考えた。
発展:コムギの進化(染色体の倍数化による種分化)
AAのゲノム(2n=14)をもつ一粒系コムギとBBのゲノム(2n=14)をもつコムギ(おそらくクサビコムギ。未確定)が、それぞれA(n=7)が入った配偶子、B(n=7)が入った配偶子をつくり、それらが受精した。
そして、ABのゲノム(n=14)をもつ子供が生じた。
【A+B→受精→AB】
この子供は子孫を残すことができない。同形同大の染色体のペアがない(ABに含まれる14本の染色体の形はどれもバラバラ。塩基配列もバラバラ)ので、対合できず、減数分裂ができない(種子を残せない)。
しかし、倍数化が起こり(おそらく、減数分裂時のミスで、ABがそのまま入った配偶子が生じ、それらが受精した)、AABBのゲノム(2n=28)をもつ個体(二粒系コムギ[デュラムコムギ。マカロニコムギとも呼ばれる])が生じた。
【AB→倍数化→AABB】
*下の図では、染色体を省略して描いている。たとえば、AAのゲノムを持つ一粒系コムギ(図左上)の染色体を1対の青い棒で表現しているが、実際は、一粒系コムギは7対(14本)の染色体をもつ。
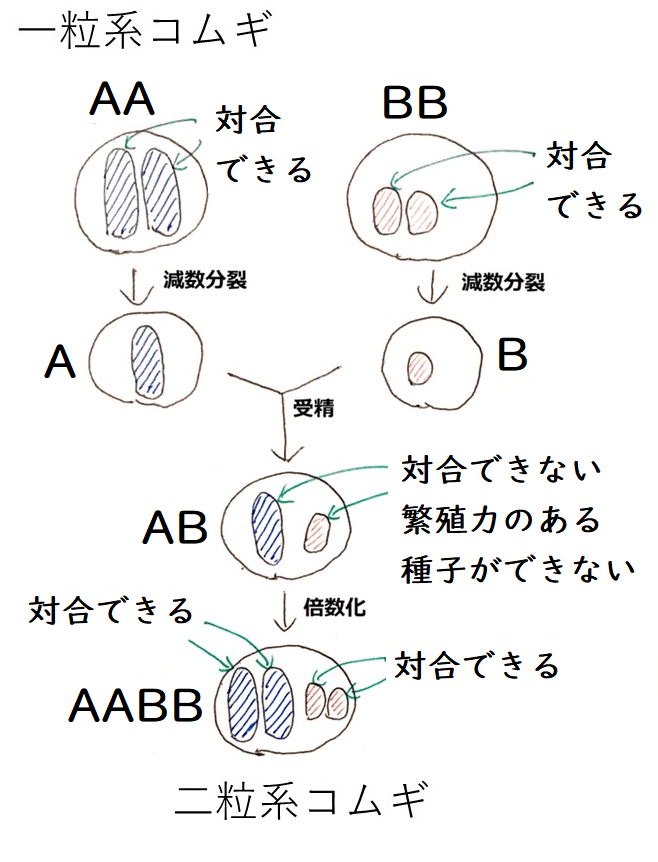
このAABBのゲノムを持つコムギ(二粒系コムギ)は、AB(n=14)の配偶子をつくる。
この配偶子と、DDのゲノム(2n=14)をもつコムギ(タルホコムギ)のつくった、D(n=7)が入った配偶子が受精した。
【AB+D→受精→ABD】
生じた子供はABD(n=21)だが、このままでは子孫を残せない(ABDの21本の染色体は形がバラバラで対合できず、減数分裂ができない)。
しかしこれが倍数化して2n=42のパンコムギ(AABBDDのゲノムを持つ)が生じた。
【ABD→倍数化→AABBDD】
パンコムギは、パンやうどんの材料となっている。
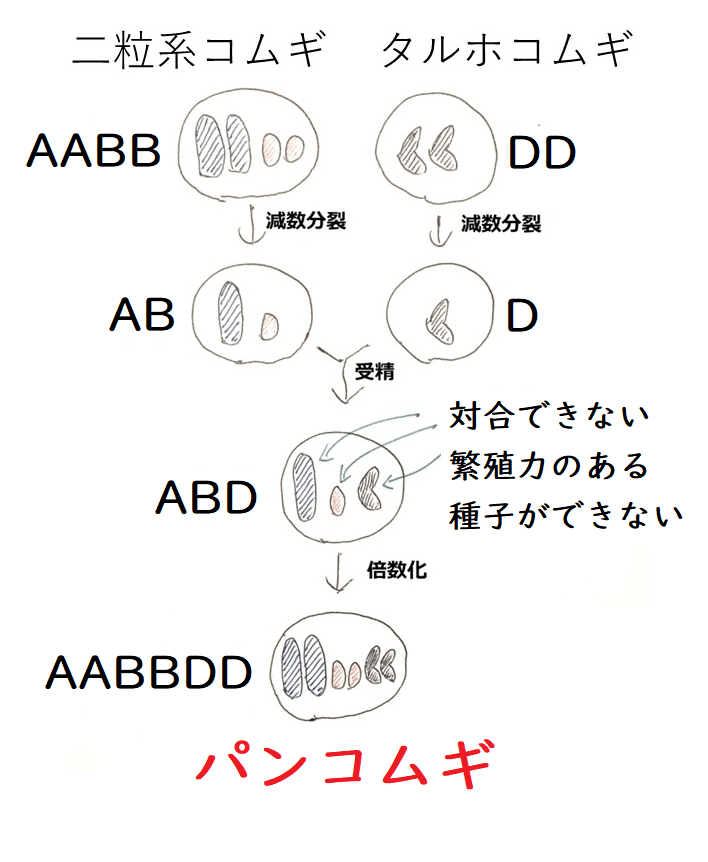
*AやBやDを基本的な単位と数えれば、パンコムギはそれらを6セットもつことになる。このような場合、「パンコムギは6倍体である」と言うことがある(覚えなくてよいが、今回のように、1そろいの染色体の組を、近縁ではあるが別種の2種以上の生物から受け継いだ雑種を、特に『異質倍数体』という)。x=7を基本的な数として、パンコムギのゲノム(AABBDD)を2n=6x=42と表現することがある(この表現のほうがよくゲノムの構成をよく表している)。
*なお、A、B、Dのゲノムをまったく別物のように説明しているが、実際は、遺伝子、遺伝子間領域の塩基配列が類似している部分も多い。
雑談:木原均(きはらひとし)は、上記の仮説を立証した。様々な種を集め、交雑させ、染色体が対合するかどうかを分析することでパンコムギの祖先を突き止めた。この方法はゲノム分析と呼ばれる。
*AとA、BとB、DとDが対合する。
たとえば、DDのゲノムをもつタルホコムギを持ってきて、パンコムギと交配させ、ABDDの個体を作るとする。すると、減数分裂時にDとDが対合するので、DDのタルホコムギはパンコムギの祖先の1つであるとわかる。木原は、染色体が対合するかしないかを調べ、パンコムギのゲノムの祖先を突き止めたのである。
「ゲノム分析による小麦属の分類が完成した事から見て小麦の研究は一先づ一段落ついたと見る事も出来るが、ゲノムの研究は之から初まるのである。」木原均『小麦の研究』より
「地球の歴史は地層に、生物の歴史は染色体に書いてある。」木原均『小麦の祖先』より
雑談:木原は、パンコムギ(AABBDD:このゲノムの由来は当時不明であった)と、二粒系コムギ(AABB)を交配し、その雑種(F1)植物(AABBD)の減数分裂において、14対の二価染色体(AとAが、BとBが二価染色体を形成した)と、7個の一価染色体を観察した。このことから、二粒系コムギ(AABB)はパンコムギの祖先のひとつだとわかった。その後、パンコムギ(AABBDD)とタルホコムギ(DD)を交配して得たF1植物の減数分裂での染色体の対合を調べたところ、7対の二価染色体(DとD同士が対合した)と14個の一価染色体が観察された。このことから、パンコムギのゲノムDは、タルホコムギ(DD)に由来すると判断した。さらに、二粒系コムギ(AABB)とタルホコムギ(DD)を交配し、作成したF1植物(ABD)を用いて染色体倍加実験を行い、AABBDDのゲノムを持つ植物を作成した。この植物は、普通系コムギにきわめて類似していた。
雑談:木原均は、ゲノムを「生活機能の調和を保つうえで欠くことのできない染色体の一組」と見なし、現代的なゲノムの概念を確立させた。
雑談:コムギは、世界で最も多く栽培されている作物の一つであり、一年中世界のどこかで収穫されている。
雑談:パンコムギ(普通コムギ、Triticum aestivum)は、単に「コムギ」とも呼ばれる。世界で広く栽培され、パン、めん、菓子など、様々な食物の原料になっている。世界のコムギ栽培面積の9割はパンコムギである。
雑談:コムギ属は、一粒系コムギ(1穎花だけが稔る)、二粒系コムギ(2穎花が稔る)、普通系コムギ(3~4穎花が稔る)に分けられる。
*穎花:「えいか」と読む。イネ科植物の花。
雑談:現在主に栽培されているのは、二粒系コムギ(デュラムコムギ[マカロニコムギ]。マカロニやスパゲティに適する)と普通系コムギ(パンコムギ、クラブコムギ)である。
雑談:コムギの起源となる種は、野生型の一粒系コムギ(AA)であると考えられている。これが、未確定のコムギ(おそらく野生していたクサビコムギ[BB]。クサビコムギは今も雑草として見られる)と交雑して、野生型の二粒系コムギ(AABB)が誕生した。二粒系コムギは栽培型の二粒系コムギであるエンマコムギ(AABB)へと発達した(エンマコムギからやがてデュラムコムギ[マカロニコムギ]などの二粒系コムギが発達していった)。そして、エンマコムギが雑草であったタルホコムギ(DD)と交雑し、普通系コムギ(AABBDD)が誕生した(この普通系コムギからパンコムギが発達した)。
*タルホコムギのゲノムを表す記号がDなのは、「普通系」をドイツ語で「Dinkelreihe」と書くことから(Dゲノムは普通系コムギ特有のゲノムとして研究されてきた。木原均はその由来を突き止めたのである)。
*普通系コムギが誕生したのは、紀元前5000年頃ではないかと考えられている。
講義動画【コムギの進化】
まだわかっていないこと
● 種をどう定義すればよいか。無性生殖を繰り返す生物について、種の概念をどう適用すればよいか。また、自然では交配しないが、実験室では交配する場合は、どう考えればよいか。さらに、化石でしか見つからない生物については、どう考えればよいか(ホモ・ネアンデルターレンシスとホモ・サピエンスは交配し子を残したらしいと考えられているが、では、この2つの人類は同種としてよいか)。
● 大進化に関する確かな証拠は、どのように得ればよいのか。
● 自然選択は「どのような単位に」働いているのか(自然選択が生じる生物学的な実体あるいは単位、すなわち「淘汰の単位」は何か)については、結論が出ていない。自然選択は遺伝子に対して働いているのか(より多くの子孫を次世代に残すことができる要因となった遺伝子の頻度は増加する)?個体に対して働いているのか(適応度の高い個体が次世代により多くの子孫を残す)?または家族、個体群、種など、個体のグループ全体に対して働いているのか?結論は出ていない(分子生物学者・遺伝学者・生態学者の間で議論が続いている)。
● ダーウィンが種の起源の中で自ら認めているように、自然選択説では説明に困ることもある。例えば、眼のように複雑な受容器の形成である。突然変異と自然選択の繰り返しのみで、眼のような複雑な器官が生じ得るだろうか。人類史上最高の天才とも言われる物理学者フォン・ノイマンは、以下のようにダーウィン理論を批判(疑問を提起)したと、ハイゼンベルクは記している。
「フォン・ノイマンは悲観論者だった。この数学者は生物学者を自分の研究室の窓の所へ連れて行って言った。"あそこの丘の上にこじんまりした別荘が見えますか?あれは偶然によってできたのです。数百万年の間に丘が地質学的な過程によってつくられました。木が生えて腐り、倒れてから、再び生長し、そして、たまたま風が丘の頂を砂で覆いました。石は噴火によって吹き飛ばされてきて、偶然によって秩序正しく置かれました。大部分は何か他のものになりましたが、たった一度、別荘がそこにでき、そこに人間が引っ越してきて、今住んでいます。"」ハイゼンベルク『部分と全体』より
あなたはフォン・ノイマンに何と答えるか。