
【高校生物】代謝③「光合成とはどのような反応か?」

~プロローグ~
「ランプが燃えれば油の汚れができ、ろうそくが燃えれば燃えさしが残る。天の光だけが清らかに輝いてしみを残さない。」ゲーテ『箴言と省察』より
★テストに出やすいワード
①光化学系Ⅱ
②光化学系Ⅰ
③クロロフィルa
④電子伝達系
⑤カルビン・ベンソン回路
要点:光合成には、様々な光合成色素が使われる。
● 光合成を行う生物(植物・藻類・シアノバクテリア・光合成細菌)に存在し、光合成のエネルギー源として光エネルギーを吸収する色素の総称を光合成色素という。クロロフィル、カロテノイド、フィコビリンの3つに分類できる。
● 酸素発生型の光合成を行う生物はクロロフィルaをもつ。
雑談:クロロフィルは、アミノ酸などを材料にして、複雑な反応を経て合成される。
雑談:クロロフィルのクロロは「緑」、フィルは「葉」の意味。
● 植物は、カロテノイド(カロテン・キサントフィル)などの補助色素を(多くのクロロフィルaも補助色素として働く)持つ。補助色素は、吸収した光エネルギーを反応中心にある特別なクロロフィルaに伝える。
*カロテノイドには、カロテンとキサントフィルの2種類がある。
雑談:カロテノイドの内、炭化水素のみのものをカロテン、水酸基、カルボニル基などの形で酸素原子を含むものをキサントフィルという。
雑談:下図は、補助色素が吸収した光のエネルギーが反応中心のクロロフィルaまで伝達されるイメージ。

*実際は、補助色素(アンテナ色素とも呼ばれる)は、タンパク質と結合し、集光性複合体として存在している。
雑談:カロテノイドは、補助色素として、吸収した光エネルギーをクロロフィルに移行させている。また、活性酸素を除去する機能も持っている。
雑談:かつては、クロロフィルa以外の色素を補助色素と呼んでいた。しかし、現在では、クロロフィルaも、その大部分は補助色素として働き、エネルギーを反応中心のクロロフィルaに伝えていることがわかった。そのため、色素の特定の分子種を指して、補助色素と呼ぶのは適当ではない(たとえば、「クロロフィルaという色素は反応中心色素で、それ以外の色素は補助色素である」という説明は適当ではない)。
発展:色々な光合成色素
(今慌てて覚えなくてよい。系統・分類を学んだ後、参考程度に見ればよい。出題頻度は低い。表には例外があるからである。)
● カロテンは橙黄色(わかりやすく言えばオレンジ色。問題文によっては黄色と表現されることもある)、キサントフィルは黄色、クロロフィルは緑色とだけ知っておけばよい。(厳密には、クロロフィルのうちクロロフィルaが青緑、クロロフィルbが黄緑、キサントフィルのうち、フコキサンチンは褐色。ただ、色の見え方には個人差があるので、今はほぼ入試では問われない)。
雑談:フィコシアニン(phycocyanin)は青色(cyanoは藍色を表す)。フィコエリトリン(phycoerythrin)は赤色または赤橙色(erythroは赤を表す)。

● クロロフィルにはNやMgなどの元素が含まれる。
● クロロフィルは青紫色(短い波長)と赤色(長い波長)をよく吸収する。緑色の光をよく反射するので、その緑色の光が我々の目に入り、葉が緑色に見える。

雑談:実際は、葉の内部に入った光は散乱を繰り返す。その中で、赤色と青紫色の光の多くはクロロフィルに吸収される。その結果、緑色の光のみが外部に漏れ出ることになる。なお、緑色の光も、葉の中で反射を繰り返し、効率的に吸収されると考えられている。
雑談:「葉の色は、本当は何色なのか?」と思うかもしれない。しかし、「葉」は、究極的には電子など素粒子の集合から成っており、その「本当の色」を問うことは無意味である。我々が「色」と呼んでるものは、視細胞が受けた様々な波長の電磁波の情報を、脳が処理した結果である。
雑談:青色の光は短波長側、赤色の光は長波長側の電磁波である。赤色の光は長い波長で直進性が高く、遠くまで届く。なので朝焼けや夕焼けは赤い。青色の光は波長が短く、よく散乱する。なので昼の空は青い。
● 色素それぞれがよく吸収する光の波長を示したものを吸収スペクトルという。
● クロロフィルaのほうがbに比べて、より青紫側(短波長側)と、より赤側(長波長側)の光を吸収する。「aは外堀」と覚えておこう。
下図は吸収スペクトルのイメージ。

● 光合成速度と光の波長の関係を示したものを作用スペクトルという(下図は作用スペクトルのイメージ)。作用スペクトルの形はクロロフィルaの吸収スペクトルに似ているが、完全に一致はしない。その理由の一つは、クロロフィルa以外のクロロフィルbやカロテノイドなどの補助色素が様々な波長の光を吸収するからである。

雑談:葉に照射される光強度の上昇に伴い、葉が吸収する光エネルギーは増えていく。光が強すぎると、光エネルギーの吸収量が、光合成による消費量を上回るようになる。過剰な光エネルギーは、無駄になるばかりでなく、活性酸素の生成につながってしまう。活性酸素は、タンパク質の破壊、膜脂質の過酸化などによって葉緑体の機能を低下させる。植物は、このような光阻害(光によって光合成の速度が低下する現象)を防ぐための様々なしくみをもつ。たとえば、葉緑体は、弱光化では光吸収が最大になるように配置されるが、強光下では光吸収が小さくなるように配置される(このような葉緑体光定位運動には、後に学ぶフォトトロピンという光受容体が関わっている)。
要点:ペーパークロマトグラフィーによって、光合成色素を分離することができる。
講義動画【ペーパークロマトグラフィー】
● ペーパークロマトグラフィーでは、溶媒前線に近い順に、カロテン、キサントフィル、クロロフィルa、クロロフィル bの順で、光合成色素が分離される。
語呂「カキクa b(ペーパークロマトグラフィーでは、溶媒前線に近い順に、カロテン、キサントフィル、クロロフィルa、クロロフィル bの順で分離する。)」


雑談:クロマは色、グラフィは記録というギリシャ語が語源。
雑談:色々な成分が含まれる試料中の各成分を、固定相と移動相という性質の異なる相の間で繰り返し相互作用させ、それぞれの相に対する親和性の違いによって分離する方法をクロマトグラフィーという。
試料の成分は、固定相と移動相との間を往復しながら、移動相と共に移動するが、固定相に対する親和性の高い成分ほど移動速度は遅くなるため、その差によって分離が達成される。
雑談:厳密には、ペーパークロマトグラフィーは、濾紙を固定相とし、主に有機溶媒を移動相として、毛細管現象による展開液の浸透によって、試料成分の展開・分離を引き起こす手法である。
● 固定相に濾紙ではなく薄層プレート(シリカゲル薄層プレートなど)を使う場合は、薄層(はくそう)クロマトグラフィーといい、前線に近い順から、カロテン、クロロフィルa、クロロフィルb、キサントフィルの順番に分離される。 (固定相が異なると化学的な分離条件が変わるので、このように濾紙を使った場合と差が出てくる。)
● 色素の移動距離(原点からその色素までの距離)÷溶媒の移動距離(原点から溶媒前線までの距離)の値をRf値という。
Rf値はrate of flow の略。「その物質は、どの程度流れ動きますか?」というニュアンスの意味。条件さえ揃えれば、光合成色素ごとに決まったRf値になるので、その色素がどの光合成色素か推定できる。
● 当たり前だが原点を展開液の中に沈めてはならない。サンプルが展開液に拡散してどこかに行ってしまう。
● 色素の移動距離を測定する時は、色素の「中央」から原点までを測る。
● 植物などから色素を抽出するときに使う石油エーテル、トルエンなどの抽出液や、ジエチルエーテル、キシレンなどの展開液は、有機溶媒で、すぐに蒸発し、引火性も持つ。なので溶液にはフタをする。
● R f値をなるべく正確に測定することができるように、原点は直径5mm以内にすることが望ましい。
雑談: ペーパークロマトグラフィーは、1944年にアミノ酸分析に用いられてから広まった(アミノ酸の場合は色がついていないので、ニンヒドリン反応を起こして着色する)。 クロマトグラフィー自体は、1903年にツヴェットが発明し、カロテンとキサントフィルの分離に成功していたが、長い間化学者の注目は受けなかった。シリカゲルを用いた薄層クロマトグラフィーが1956年にシュタールによって行われ、その精度の高さから、急速に普及した。
雑談:カロテンには様々な異性体がある。その中でも最も広く分布しているのはβ-カロテンで、ニンジンや柿の色もβ-カロテンに由来している。β-カロテンは活性酸素の発生を抑制し、結果として強すぎる光から植物体を保護している(強すぎる光は過剰な還元力を生み、その結果、活性酸素が発生することが知られている[また、光エネルギーがクロロフィルから速やかに移動しせず、特殊な状態になったクロロフィルが酸素と反応し、活性酸素が発生してしまう]。活性酸素は、色素やタンパク質を破壊する)。
雑談:老化によって葉の色が緑色から「黄色」に変化するのは、一般に、クロロフィルが分解され(窒素の回収に伴いクロロフィルが分解される)、カロテノイド(カロテノイドには窒素が含まれず回収されない)の色が目立つようになるからである(また、アントシアンが合成され、「紅葉」になることもある。アントシアンは過剰な光を吸収し光阻害を防ぐということが知られているが、紅葉の生理的な意義についてはまだよくわかっていない)。
挿絵:光合成色素のようなファッション

要点:光合成では、光エネルギー用いてATPとNADPHがつくられる。それらは、カルビン・ベンソン回路で炭水化物の合成に使われる。
(1)葉緑体の構造
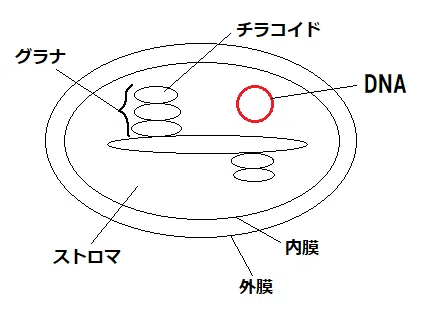
*チラコイド:葉緑体にある扁平な袋状の構造(葉緑体は、外膜と内膜に加え、チラコイドと言う第3の膜系を持っている)。チラコイドは「袋状のもの」という意味。
*グラナ:チラコイドが積み重なっている部分。
覚え方「グラナは積み重ねた座布団みたい。グラナの上に座ったらグラグラしそう。グラグラ。グラナ。」
*ストロマ:チラコイドを含む流動性の部分。
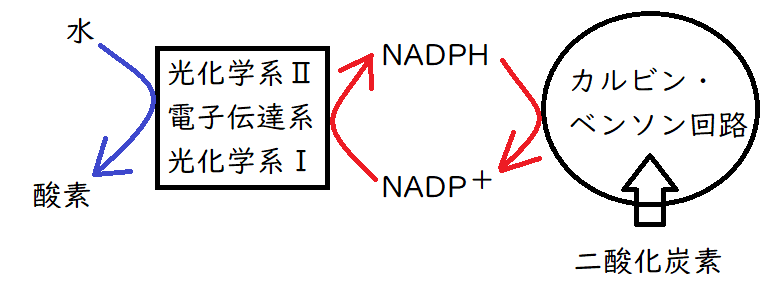
(2)光合成の概要
①光エネルギーを用いて、ATPと還元力(NADPH)がつくられる。その過程で、水が分解され、分解産物である酸素が廃棄される。(光化学系Ⅱ・Ⅰと電子伝達系:チラコイド膜で起こる反応)
②①で得たATPとNADPH(還元力)を用いて、CO2が固定され、炭水化物(糖)が合成される。(カルビン・ベンソン回路:ストロマで起こる反応)

*「カルビン・ベンソン回路」は「カルビン回路」ともいう。昔から日本の高校生物の教科書には「カルビン・ベンソン回路」と書かれてきたが、最新の教科書では「カルビン回路」と書かれることが多くなった(「カルビン回路」の方が一般的であるので、良い流れであると思う。大学では「還元的ペントースリン酸回路」「光合成的炭素還元回路」などとも呼ばれる)。
(3)光合成の詳細
● 光化学系Ⅱ、光化学系Ⅰは葉緑体のチラコイド膜にある。
● 以下はイメージ。①、②、③についてこれから説明する。
①光化学系Ⅱ:水の分解→酸素の発生。
②電子伝達系: H+がチラコイド内腔に能動輸送される。そして、H+の濃度勾配を利用して、ATP合成酵素でATPが生成される。
③光化学系Ⅰ: NADPHの生成。

①光化学系Ⅱ(水の分解・酸素の発生)
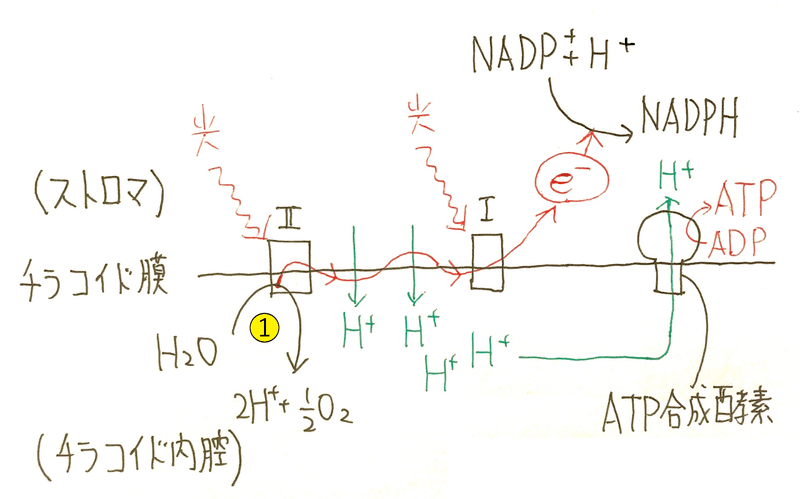
水が分解される→副産物として酸素が発生する。
(電子が水から引き抜かれ、副次的な現象として、酸素が発生する。)
光化学系Ⅱから飛び出た電子(e‐)は電子伝達系に伝えられる。電子を失った光化学系Ⅱのクロロフィルaは、水から電子を受け取る(水を分解し、水の電子を奪い取る)。
*「副次的な現象として、酸素が発生する」とは、つまり、酸素の発生は主目的ではないということである。酸素は副産物である。必要なのは水の電子である。酸素はいらないので捨てる(酸素は、ある意味、排気ガスである)。
下図は光化学系Ⅱのイメージ。

②電子伝達系(チラコイド膜上を電子が伝達する)

電子が伝達される際に、H+がチラコイド内腔に能動輸送される。そして、H+の濃度勾配を利用して、ATP合成酵素でATPが生成される(チラコイド内からストロマにH+が流れ出る勢いを使って、ATPが合成される[このしくみを光リン酸化という])。

③光化学系Ⅰ(NADPHの生成)

NADPHが生成する(NADP+とH+と電子が結合し、NADPHが生成する)。
電子を失った光化学系Ⅰのクロロフィルaは、電子伝達系を伝わってきた電子を受け取る。
雑談:実際は、電子は、フェレドキシン(Fdと略される。植物などに広く分布し、電子伝達を行うタンパク質)を介して伝えられる。そして、フェレドキシン‐NADP+還元酵素という酵素がフェレドキシンからNADP+に電子を伝達して還元する。下図はイメージ。

発展:光化学系Ⅱ・電子伝達系・光化学系Ⅰについて
● 光は電磁波なので、電子を揺さぶる。光化学系の反応中心にあるクロロフィルaの電子が光によって揺さぶられ、十分なエネルギーを得ると、クロロフィルaから電子が飛び出す。
● 光化学系Ⅱから飛び出た電子は電子伝達系を伝わる。その際、電子のエネルギーがH+のチラコイド内腔への能動輸送に使われる。電子は最終的に光化学系Ⅰに渡される(光化学系Ⅰが失った電子の穴を埋める)。
● 電子を失った光化学系Ⅱのクロロフィルaは、水から電子を受け取る(水の電子を奪い取る)。
● 光化学系Ⅰでは、飛び出た電子はNADP+に渡され、NADPHが生じる。
● 電子を失った光化学系Ⅰのクロロフィルaは、電子伝達系を伝わってきた電子を受け取る。
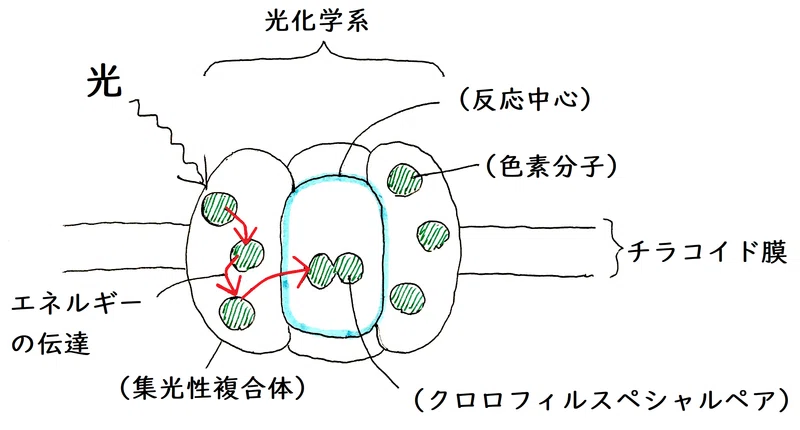
● 光化学系とは、「光化学反応を起こすのに必要な(色素などを含む)システム全体」を指す(植物の光化学系の反応中心には、特別なクロロフィルaの分子対が存在する)。
*色素が光エネルギーを吸収して化学反応を起こすことを総称して光化学反応(こうかがくはんのう)という。
● 光化学系Ⅰ、Ⅱは起こる順番とは関係ない。Ⅰ、Ⅱの順に解明されたというだけ。
● 光合成での電子伝達系によるATP生成のしくみを光リン酸化(こうりんさんか)という(「光」エネルギーでADPを「リン酸化」)。
雑談:電子の動きのイメージ


*光化学系と電子伝達系でつくられたNADPHやATPは、カルビン・ベンソン回路(におけるPGAからGAPへの変換、GAPからのRuBPの再生)で消費される。詳しくは後述する。
雑談:光化学系には補助色素(光を吸収しそのエネルギーを反応中心に伝える役割をする色素の総称)がたくさん含まれている。それらの色素が吸収したエネルギーは、最終的に光化学系の中心部(反応中心)にある、特別なクロロフィルa分子対に集約される。
<Q.光合成の光リン酸化と呼吸の酸化的リン酸化は同じ?…確かに似ているが、異なる反応である。光リン酸化は、光合成において、電子伝達系により形成されたH+の勾配を利用して駆動されるATPを合成する反応。酸化的リン酸化は、呼吸において、有機物を酸素により酸化して得られるエネルギーをATPのエネルギーに変換する反応。2つとも同じような反応だが、たとえば、エネルギーの源は異なる。光リン酸化のエネルギー源は、元をたどれば光エネルギーである。酸化的リン酸化のエネルギー源は、元をたどれば有機物である(まあ、その有機物内のエネルギーも、もっと元をたどっていくと、生産者がとらえた光エネルギーに由来しているのだが)。>
講義動画【光化学系のクロロフィルa】
雑談:光化学系は反応中心1個と数個の集光性複合体(アンテナ複合体とも呼ばれる。タンパク質と補助色素が結合したもの)からできている。光エネルギーは集光性複合体によって集められる。エネルギーは共鳴エネルギー移動によって、分子から分子へと移動し(電子ではなくエネルギーが移動する)、反応中心にたどり着く。そして、反応中心では、特別なクロロフィルaの分子対(クロロフィルスペシャルペア)の電子が1個励起される。
挿絵:ATP合成酵素のようなランタン

④カルビン・ベンソン回路(カルビン・ベンソン回路はATPとNADPHを使って二酸化炭素を還元し、糖を合成する)
● カルビン・ベンソン回路:二酸化炭素を炭水化物に還元する反応経路。ストロマで起こる。
雑談:すべての高等植物は、カルビン・ベンソン回路で固定した炭素を、主にデンプンやセルロースに変換する。
雑談:CO2はエネルギー的に低い状態の分子である。CO2を有機分子に取り込む反応には、エネルギーと還元力が必要である(C-C結合の形成にはATPの供給するエネルギーが必要である。また、CO2を炭水化物に変換するためには、強力な電子供与体が、高いエネルギー状態の電子を供給しなければならない。すなわち、電子を押し付ける力、還元力が必要である。植物では、NADPHがこの電子を供給する)。
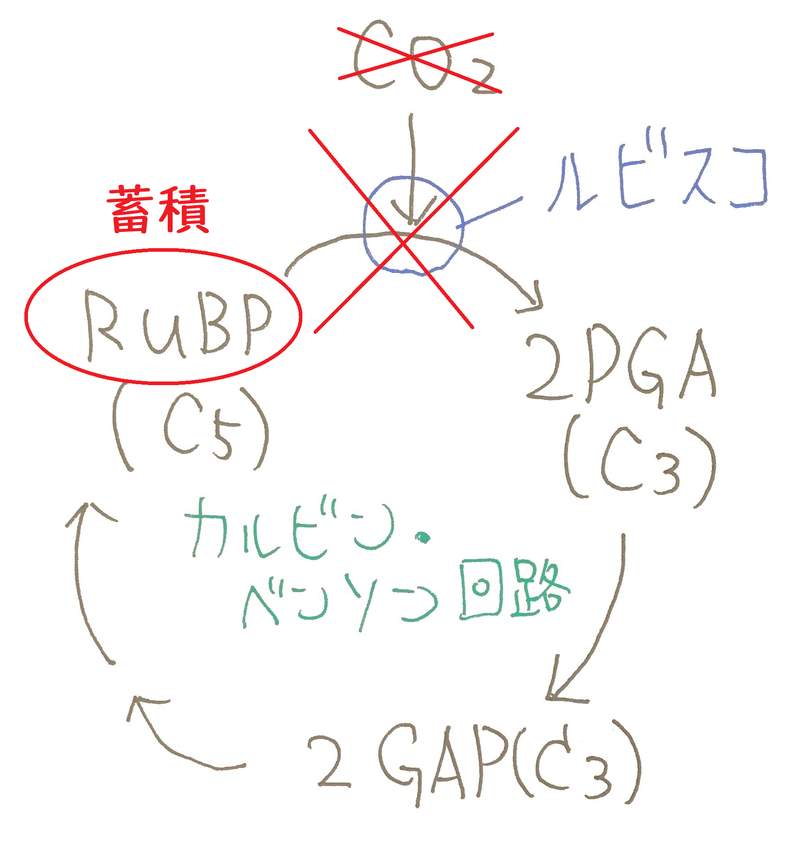
● まずチェックすべき重要な反応はRuBP→2PGAの反応である。この過程でCO2が固定されるからである。
● RuBPとCO2の反応を触媒する酵素をルビスコという(ルビスコは地球上で最も量が多い酵素とも言われる)。

● RuBP(リブロースビスリン酸)、PGA(ホスホグリセリン酸)、GAP(グリセルアルデヒド三リン酸)は、正式名称を知っておくとよい。正式名称をぼんやりとでも覚えておけば、略称の書き間違いななくなる(PGAをPAGなどのように誤って書いてしまうミスを減らせる)。
・RuBP ( ribulose bisphosphate ):リブロースビスリン酸(リブロース二リン酸とも言う。「ビス」は2の意味)
・PGA ( phosphoglyceric acid、phosphoglycerate ):ホスホグリセリン酸(PGAはC3植物のCO2固定の初期産物)
・GAP ( glyceraldehyde-3-phosphate ):グリセルアルデヒド三リン酸
● RuBP(C5と書いてあるのは、「炭素を5個もつ化合物」という意味)は、CO2と結合する(この反応はルビスコが触媒する)。
カルビン・ベンソン回路は、CO2分子を、RuBP(CO2の受容体といえる)に結合させることによって、1回に1分子ずつ取り込む。
● C5(Cを5個もつ)のRuBPと、CO2(Cを1個もつ)が結合し、一瞬、C6の不安定な化合物ができるが、すぐに2つに分解し、PGA(C3)が2分子生じる。
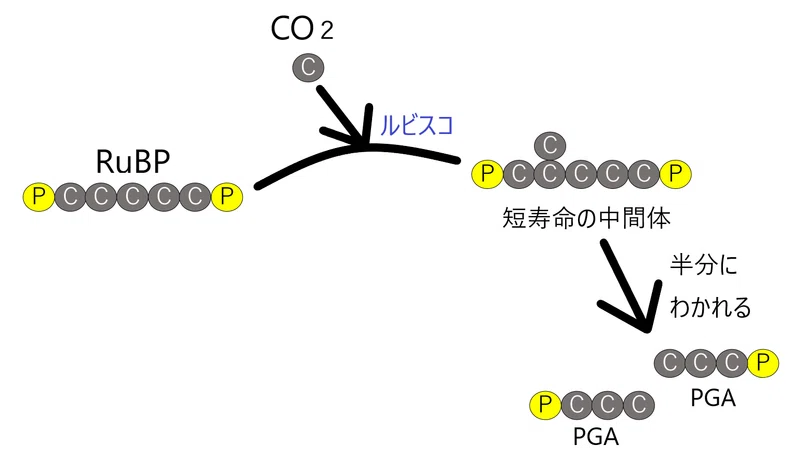
● PGAから、いくつかの反応を経て、GAPが生じる(この反応ではATPとNADPHが使われる)。

● 実際は、たくさんのGAPが生じているが、その中のGAPの一部がカルビン・ベンソン回路から抜け、糖の合成に使われる。その他のGAPはRuBPの再生に使われる(この再生にはATPが使われる。このRuBPの再生には、実際にはいくつもの反応が関わる)。
*GAPからRuBPが再生するステップは、実際は複雑である。簡単に結論だけ言うと、5分子のGAP(5分子×3個の炭素=15個の炭素)から、3分子のRuBP(3分子×5個の炭素=15個の炭素)が生じる。5分子のGAPに含まれる炭素の数と、3分子のRuBPに踏まれる炭素の数は同じである(15個)。下図はイメージ(リン酸などは描いていない)。

雑談:ミトコンドリアで発生したCO2は、通常の大気圧条件では、最もCO2分圧の低い葉緑体のストロマのルビスコに向かって拡散する。
● GAPから直接グルコースができるわけではないが、できる化合物をグルコースに換算すると、光合成は以下のような反応式になる。
6CO2+12H2O → 6O2+6H2O+C6H12O6
(あたかも、呼吸の逆反応であるかのようである。もちろん実際には別の酵素、仕組みが関わっている)
語呂「カルビー好こ(カルビン・ベンソン回路、ルビスコ)」
語呂「RPG(RuBP→PGA→GAP)」

雑談:GAP正味1分子の合成のために、カルビン・ベンソン回路は合計で9分子のATPと6分子のNADPHを消費する。光化学系、電子伝達系がこれらのATPやNADPHを再生させる。
雑談:カルビン・ベンソン回路から出たGAPは、グルコースやほかの糖など、さまざまな有機化合物を合成する代謝経路の出発物質となる。
講義動画【炭素数の確認】
問題:空気を抜いて(二酸化炭素を取り除いて)、取り出した葉緑体を含む液に光照射を行った。その結果、ほとんど酸素の発生は見られなかった。酸素が発生しなかった理由を説明した次の文章の空欄を埋めよ。
「二酸化炭素がないので、( ① )回路が進行せず、NADPHが消費されない。その結果、光化学反応の進行に必要な電子受容体であるNADPH+が供給されず、( ② )の分解が起きなかったから」
答え:①カルビン・ベンソン ②水
解説

①空気を抜いたので二酸化炭素がなくなり、カルビン・ベンソン回路が停止した。
②光化学反応に必要な電子受容体であるNADP+の産生がストップした。
③NADP+がないので光化学反応が停止し、酸素が発生しなくなった(光化学系Ⅱ、電子伝達系、光化学系Ⅰは協調して働いているので、光化学系ⅠにNADP+が供給されなくなると、光化学系Ⅱにおける水の分解も停止する)。
*なお、空気を抜いた条件でも、シュウ酸鉄(Ⅲ)など、NADPH+の代わりに電子を受け取ってくれる物質を加えれば、酸素は発生する(この資料の最後の方にあるヒルの実験を参照)。
● 一般に糖はスクロースの形で維管束を通って転流する。
*スクロースは、いわゆる普通の「お砂糖」である。
*転流:植物体内で光合成産物などがある組織から離れた組織に運搬されること。
雑談:スクロースは自然界で、最も多い二糖類である。
雑談:αーグルコースの1位のヒドロキシ基と、βーフルクトースの2位のヒドロキシ基が脱水縮合してできた二糖がスクロースである。スクロースは還元性を示さない。そのグリコシド結合はアミラーゼなど広く存在する加水分解酵素による影響を受けないので、植物体内で炭素の輸送に使うのに都合がよいと考えられている。
● 急に光を消したり、CO2の供給を止めたりする実験は超頻出。「通行止めの直前が渋滞する」と考えればよい。
(1)二酸化炭素の供給が止まると、RuBP+CO2→2PGAが阻害される。
よって、急に二酸化炭素の供給を止めるとRuBPが蓄積する(PGAは減っていく)。
以下はCO2の供給を止めた時のイメージ。

(2)PGAからRuBPが再生する反応には、光照射のもとでつくられるATPとNADPHが必要である。よって、急に光の供給を止めるとPGAが蓄積する(RuBPは減っていく)。
以下は光の供給を停止した時のイメージ。

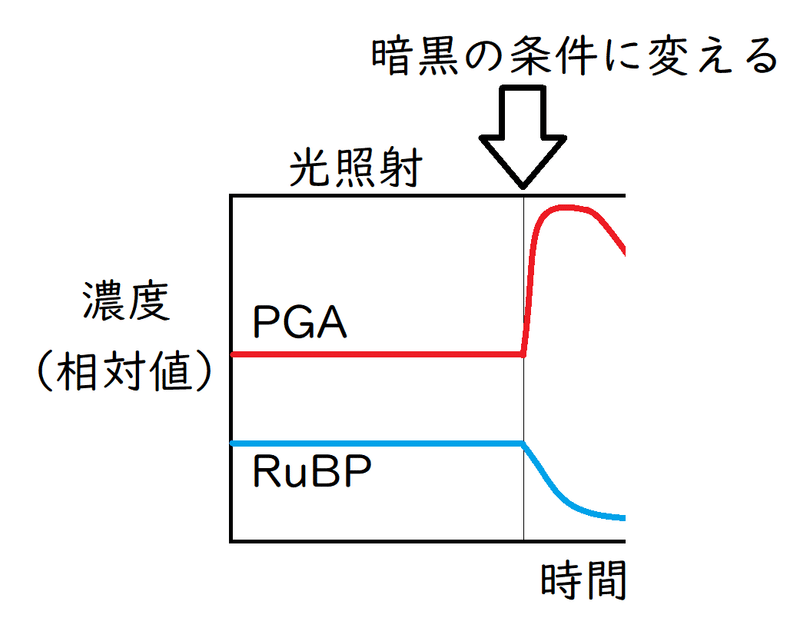
雑談:上の図で、PGAが暗黒条件に変えて少し経った時に減少し始めているのは、PGAの酸化的な分解や、他の物質への変換が原因であると考えられる。
● 一般的な植物はC3植物である。CO2を取り込んではじめにできる化合物は炭素を3つもつ(C3化合物である)PGA(ホスホグリセリン酸)だからである(この後、特別なC4植物について学ぶ)。
講義動画【光合成】
要点:C4植物はCO2の固定とカルビン・ベンソン回路を空間的に分けている。CAM植物はCO2の固定とカルビン・ベンソン回路を時間的に分けている。
● 二酸化炭素濃度が薄いと、ルビスコがうまく働かず、光合成が阻害される(ルビスコが酸素と反応してしまう。これを光呼吸という)。
雑談:ルビスコ(rubisco)の正式名称は「ribulose bisphosphate carboxylase/oxygenase」である。「ribulose bisphosphate(RuBP)」と、 「carboxylase(炭酸固定酵素)/oxygenase(酸素化酵素)」という語がくっついている。この名前は、「RuBPとCO2の反応を触媒するだけでなく、RuBPとO2の反応も触媒し得る」ということを表している。なお、ルビスコは、アメリカの製菓会社ナビスコを文字って提唱された(当時ルビスコの食品化の研究が進められていた)。
*光呼吸は、酸素を消費され、さらに、エネルギーが消費されてしまう反応である(光呼吸には活性酸素を処理する意義があるとも言われている)。
● 熱い地域や、乾燥の激しい地域に住む植物は、活発に気孔を開くと、水分が失われてしまうため、活発に気孔を開くことができない。
● 細胞内のCO2濃度が低くなると、光呼吸が起きてしまう。それを避けるために、いくつかの植物種では、第二の炭酸固定経路を用意することによって光呼吸が抑制されている。
(1)C4植物
例(必ず問われる):トウモロコシ、サトウキビ
雑談:トウモロコシは世界で最も生産量が多い穀物と言われる。C4植物は人類の生活を支えている。

● C4植物は、葉肉細胞(ようにくさいぼう)と維管束鞘細胞(いかんそくしょうさいぼう)という2つの細胞を用いて光合成を行う(葉肉細胞と維管束鞘細胞は原形質連絡でつながっており、物質が行き来できる)。
● C4植物は、葉肉細胞で固定したCO2を、維管束鞘細胞で放出する(C4植物の葉肉細胞はポンプのようにCO2を維管束鞘細胞に送る)。それにより、維管束鞘細胞の中のCO2濃度を高濃度に保っている(CO2を濃縮している)。
● C4植物の行うCO2の濃縮には、ATPのエネルギーが必要である。しかし、光呼吸の抑制という点では、C3植物より有利である。薄い二酸化炭素濃度の条件では、C4植物は、物質生産において(C3植物よりも)有利である。
● まず、二酸化炭素の固定が葉肉細胞で行われる。
● C3植物の維管束鞘細胞には葉緑体はほぼないが、C4植物の維管束鞘細胞には葉緑体がある。
● C4植物では、ホスホエノールピルビン酸(PEP)とCO2が反応し、オキサロ酢酸(C4化合物である。なのでC4植物と呼ばれる)が生成する。
オキサロ酢酸はリンゴ酸になり、維管束鞘細胞に送られる。そしてリンゴ酸はピルビン酸に変わる。その際に生じるCO2がカルビン・ベンソン回路に送られる。
*ホスホエノールピルビン酸カルボキシラーゼ(PEPカルボキシラーゼ)が、ホスホエノールピルビン酸と炭酸水素イオンからオキサロ酢酸を生成する反応を触媒する。PEPカルボキシラーゼは、CO2との親和性が非常に高い。
*維管束鞘細胞の細胞壁は厚く、CO2が逃げにくいので、維管束鞘細胞内に高いCO2濃度が保たれる。
*上記の反応は一例で、C4光合成経路の反応は植物種によって異なる。
*厳密には、大気CO2によって供給される炭酸イオンがPEPと反応する。


雑談:ハワイのサトウキビ会社のフーゴ―・コ―チャックは、サトウキビを研究材料にして14Cトレーサー実験を行ってみた。すると、その初期産物は、PGA(C3化合物)ではなく、Cを4つもつC4化合物であった。当時、この結果は受け入れられなかった。その後、ハッチとスラックが、サトウキビの光合成初期産物がC4化合物であることを確認した。
(2)CAM植物(かむしょくぶつ)
例(必ず問われる):ベンケイソウ、サボテン、パイナップル
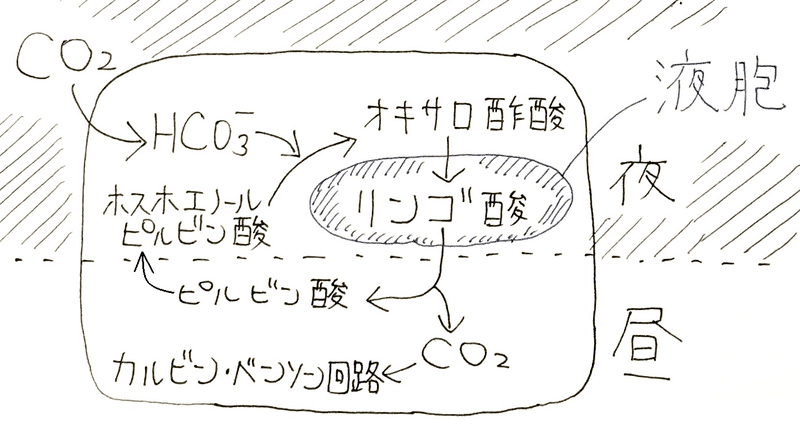
● CAM植物は、「気孔を開けてCO2を取り込む時間(夜)」と、「気孔をかたく閉じてカルビン・ベンソン回路を働かせる時間(昼)」を分けている。
● 夜間に液胞に蓄えたリンゴ酸が、昼間に炭素固定のためのCO2を供給する(非常によく問われる)。
*CAM植物は、夜間に気孔を開いてCO2を取り込み、C4化合物であるオキサロ酢酸として固定する(知らなくてよいが、正確には大気中のCO2は細胞内でHCO3ーとなる。HCO3ーはPEPカルボキシラーゼという酵素によりホスホエノールピルビン酸と反応する。その結果オキサロ酢酸が生じる)。
オキサロ酢酸はリンゴ酸に変換され、液胞中に貯蔵される。昼間に気孔を閉じ、リンゴ酸からCO2を放出して(リンゴ酸からCO2とピルビン酸ができいる。ピルビン酸は様々な反応を経てホスホエノールピルビン酸に戻る)カルビン・ベンソン回路によって有機物を合成する。乾燥の激しい昼間に気孔を閉じることができるので、蒸散による水分の損失を低く抑えることができ、乾燥地帯でも生育できる。
雑談:CAM植物は朝方にすっぱくなる。夜の間、リンゴ酸が液胞に蓄積されているからである。

雑談:CAMはベンケイソウ型酸代謝(crassulacean acid metabolism)の略である。
雑談:上に挙げた例の他にも、様々なCAM植物が知られている。着生植物であるランの仲間にもCAM型の光合成を行っているものがいる(コチョウランなど)。樹上や岩場に着生して生育する着生植物は、土壌から十分に水分を吸収できないため、常に水分ストレスにさらされていることが多い(水不足に陥っている)。よって乾燥に適応したCAM型の光合成を行うように進化したと考えられている。
問題: CAM植物の光合成のしくみには、どのような利点があるか。
答え:乾燥の激しい昼間に気孔を閉じることで、蒸散による水分の損失を防ぎ、乾燥地帯での生育に適応できる。
講義動画【C4植物】
発展:限定要因について
● 光合成において、①光の強さ②温度③CO2濃度などの要因うち、最も不足していて、光合成速度を制限している要因を「限定要因」という。
● 限定要因を改善しない限り(他の条件を良くしても)光合成速度は上昇しない。
雑談:「眠いよー」と言っている人に、「ほら!ご飯だよ!」と言っても喜ばない。「光の強さが足りないよー」と言っている植物に「ほら!高いCO2濃度だよ!」と言っても喜ばない(光合成速度は上がらない)。そんなイメージ。
雑談:ある要因が不足し、ある反応が滞ると、代謝経路全体が滞る。その滞りは、別の反応経路の効率を改善しても変わらない。最も遅い反応に代謝経路全体の速度が左右されている。これは文化祭の準備に似ている。最も遅く仕事をしているグループの仕事効率が上がらない限り、全体の仕事の速度は上がらない。たとえば、段ボールで教室を装飾する場合、段ボールを切る作業が非常に遅ければ、いくら色塗りグループや、飾りつけグループの人数を増やしても、装飾のスピードは上がらない。「段ボールを切るグループの人数」が文化祭準備の限定要因になっているのである。限定要因となっている要因を改善しない限り、文化祭の準備速度は上がらない。
● たとえば、下図のAの範囲(光の強さが★までの範囲)では「光の強さ」が限定要因である(光の強さを改善したら、光合成速度が上がっているから)。
● Bの範囲では、光の強さ以外(「CO2濃度」か「温度」)が限定要因である(光の強さを強くしても[改善しても]、光合成速度が上がっていないから)。

● また、下のグラフで、光の強さ☆、温度10℃の時の限定要因は温度である。
*下の10℃の方のグラフ(青いグラフ)を見る。光の強さを☆より強くしても、光合成速度は上がらない。しかし、温度を10℃から30℃に変えると、光合成速度は上がるから、温度が限定要因だとわかる(限定要因となっている要因を改善すると、光合成速度が上昇する。限定要因となっていない要因を改善しても、光合成速度が上昇しない)。

問題:ある植物を用いて実験を行った。光の強さをAからBに増やしたところ、光合成速度が上昇した(つまり、光の強さがAの時、光の強さが限定要因だった)。
さて、光の強さがAの時、CO2濃度をCからDに上昇させたら、光合成速度(光合成によるCO2吸収速度)はどうなるか。選べ。ただし、CO2濃度はCよりもDの方が光合成にとって好ましい条件であるとする。
①大きくなる ②変わらない ③小さくなる
答え:②変わらない(光の強さが限定要因の時、光の強さ以外の条件を良くしても光合成速度は大きくならない)
問題(発展):上の問題と同じ植物を用いて実験を行った。光の強さをAからBに増やしたところ、光合成速度が上昇した(つまり、光の強さがAの時、光の強さが限定要因だった)。
光の強さがAの時、温度をEからFに上昇させたら、この植物のCO2吸収速度(見かけの光合成速度)はどうなるか。選べ。ただし、温度は、EよりもFの方が、光合成にとっても呼吸にとっても好ましい条件であるとする。
①大きくなる ②変わらない ③小さくなる
答え:③小さくなる(光の強さがAの時は光の強さが限定要因なので、温度を良い条件に変えても光合成によるCO2吸収速度は変わらない。しかし、温度を高くしたことで、呼吸速度が大きくなり、CO2を呼吸でたくさん放出するようになる。よってCO2吸収速度[見かけの光合成速度]は小さくなると考えられる。)
雑談:限定要因の概念を提唱したのはイギリスの植物生理学者ブラックマンである。ブラックマンは、光合成が光化学反応のみによる反応であれば、光の強さに比例して、どこまでも光合成速度が大きくなるはずであると考えた。しかし、実験の結果、光の強さがある一定の値に達すると、それ以上光の強さを大きくしても光合成速度が増加しなくなることがわかった。ブラックマンは、光合成には、光化学反応だけでなく、別の化学反応も含まれていると考え、光化学反応を明反応、それ以外の(光化学反応ではない)化学反応を暗反応と呼んだ。ただし、今では明反応や暗反応という用語はあまり使われない(カルビン・ベンソン回路は直接光エネルギーを利用しないが、暗反応と呼ぶのはよくない。カルビン・ベンソン回路の駆動には、光エネルギーを使って作られたATPやNADPHが必要だからである。そもそも、本当の意味での明反応は光化学反応のみであり、その他はすべて暗反応と呼ぶことになるが、これでは議論しにくい。明反応、暗反応という語はあまり使わないほうが良い)。
発展:ヒルの実験
● ヒルは、葉緑体懸濁液にシュウ酸鉄(Ⅲ)(電子を受け取りやすい物質[還元されやすい物質、Hを受け取りやすい物質])を加え、光を当てることで酸素を発生させた(試験管の中で、光合成における「水の分解」を再現して見せた)。
● ヒルの実験から、光合成において、以下のような反応が起こるということが示された(Aは電子を受け取りやすい物質である)。
H2O + A → 1/2O2 + AH2
この反応をヒル反応という。
*ヒル反応:葉緑体が光エネルギーによって電子受容体を還元し、同時に酸素を発生する反応(一般に、ヒル反応と言えば、光化学的な酸素発生反応を指す)。

● 溶液にCO2は与えなくても、酸素が発生した。このことから、酸素は水由来であることがわかる。

● 入試では、以下のように考えて問題を解く場合がある。
「CO2がない条件で実験をしているのでカルビン・ベンソン回路は動かず、NADPHはNADP+に戻ることができない。本来電子を受け取ってくれるはずのNADP+がないので、代わりに電子を受け取ってくれる、電子受容体のシュウ酸鉄(Ⅲ)が必要になる。」
(なお、知らなくてよいが、シュウ酸鉄(Ⅲ)が電子を受け取るとシュウ酸鉄(Ⅱ)になる。)
*ふつうは(自然条件下では)、NADP+が電子を受け取る。

*CO2がないと、NADP+が再生しない。

*NADP+がないと水の分解(酸素の発生)が起きない(最終的な電子の受け取り手がいないため、電子が流れない)。

*NADP+がなくても、NADP+の代わりとなる電子受容体(たとえばシュウ酸鉄(Ⅲ)など)があれば、電子が流れ、水の分解(酸素の発生)が起こる(電子伝達のどの段階で電子受容体が電子を受け取るかは、使用する電子受容体によって様々である)。

雑談:大学入試では上記のように考えることがあるが、実際は、葉をすりつぶして葉緑体を含む溶液を作る際に、葉緑体の内膜・外膜が壊れ、ストロマ中のカルビン・ベンソン回路に関わる酵素や、NADP+が失われることが多い。特にヒルが行った実験では、葉緑体の外膜、内膜は壊れていた(すなわち、ヒルが光を当てていたのは、無傷の葉緑体ではなく、葉緑体の中身[グラナ]であった。なお、後に、イエンセンとバッシャムがCO2固定能力を持つ無傷の葉緑体を調製することに成功している)。カルビン・ベンソン回路の酵素やNADP+は、周囲の溶液に拡散し失われてしまっていた。
この場合、NADP+の代わりとなる(シュウ酸鉄(Ⅲ)などの)電子受容体を加えれば、電子が流れ、(水の分解が起こり)酸素が発生する。
ただし、無傷の葉緑体を用いた場合でも、CO2がなければ、(上の図で説明したように)NADPHがNADP+に戻ることができず、水の分解は起きないと考えることができる。
● ヒルの実験によって、重大な事実がいくつか明らかになった。
①CO2の存在なしに酸素が発生することで、酸素は二酸化炭素由来ではなく水由来であることが明らかになった。
②単離した葉緑体でも、光合成の重要な部分反応を行えることが明らかになった。
③光合成の反応において、ある物質から別の物質への電子の伝達が光エネルギーによって駆動されることが明らかにされた。
その後、ルーベンらは、酸素の同位体である18Oを用いた実験を行った。18Oを含む水の中でクロレラに光合成を行わせたところ、発生した酸素の中には18Oが含まれていた(水の分解によって酸素が発生する[酸素は水由来である]ことが明確に証明された)。
雑談:ルーベンは、第二次世界大戦中、化学兵器の研究中に事故により死亡した。まだ29歳の青年であった。世界大戦の中で、純粋科学の研究の支援は後回しにされていた。
講義動画【ヒルの実験・ヒル反応】
発展:光合成の解明の歴史
以下に、光合成の解明に関する歴史的な実験を、(実験):(その実験で主張されたこと)のように記す。
①ファン・ヘルモントの実験:植物は土を養分としない(この主張は正しい)。植物は水からつくられる(この主張は正しいとは言えない)。
(雑談:ヘルモント[ベルギーの化学者]のヤナギの木の実験は有名である。ヤナギを数年育てたところ、ヤナギの重さはかなり大きくなったが、土の重さはほとんど減っていなかった。これは大発見であった。植物体をつくる栄養はすべて土からくるというアリストテレスの主張が間違っていたことを示したのである。ただし、もちろん、彼の、植物が水からつくられるという主張は行き過ぎで、正しくない。植物は二酸化炭素と水を用いて有機物をつくっている。しかし、彼の時代、まだ気体に関する本格的な研究が始まっていなかったので、大気中の二酸化炭素に注目できなかったのは仕方がなかった。それに、彼は、「水は万物のもとである」という主張を持っていた。)
語呂「え?植物が成長したら土の重さが減るもんと思ってた(ヘルモント、植物の成長は土に由来しているわけではない)」
②プリーストリーの実験:植物は酸素を放出する。
(雑談:実際には、プリーストリー[イギリスの牧師で、化学、植物生理を研究した]は、植物が空気を綺麗にすると考えていた。確かに、植物を入れた容器に入れたロウソクは燃え続けたが、植物を入れない容器にロウソクを入れた場合は、まるで空気が汚れたかのように、ロウソクはすぐ消えてしまった。プリーストリーは、その"植物による空気の浄化"に光が関係するとまでは突き止めなかった。なお、プリーストリーは実験で「脱フロギストン空気」を発見している。それこそ酸素であった[今では、フロギストンという元素は存在しないことがわかっている。詳しくは述べないが、昔は、燃焼の際に生じる炎は、フロギストンというものが逃げる際に行う旋回運動だと考えられていた。長い間、この誤ったフロギストン説は、物理学・化学界を悩ませた。プリーストリーも、多くの偉大な化学者と同様に、フロギストンの存在を信じていた]。実際は、スウェーデンの化学者シェーレの方が先に酸素を発見していたが[シェーレはそれを"火の空気"と呼んだ]、シェーレの発表は遅れたため、プリーストリーが酸素の発見者として有名になっている。余談であるが、シェーレは、プリーストリーの植物の実験を追試してる。しかし、うす暗い部屋で実験したため、プリーストリーのいうような結果は得られなかった。)
語呂「植物が空気を綺麗にするなんてプリティーストーリーだね(植物が空気を浄化、プリーストリー)」
③インゲンホウスの実験:植物は光のある所で酸素を放出する。
(雑談:実際には、インゲンホウス[オランダの医師、植物生理学者]は、「脱フロギストン空気が放出されるのは、緑色の葉に日光が当たるときだけである」と主張した。)
語呂「インゲン豆のビニールハウスには光が必要(インゲンホウス、光が必要)」
⓸セネビエの実験:植物は二酸化炭素があるところで光が当たると光合成を行い、酸素を発生する。
(雑談:セネビエ[スイスの植物生理学者]は、石だらけの土地でも、水中でも植物が成長することに興味を持った。彼は、植物は空気中にあるものを使って体をつくるに違いないと考えた。二酸化炭素を含む水中に水草を入れ、光を当てると、気泡が発生した[その気泡が脱フロギストン空気、すなわち酸素であることも確かめた]ことから、セネビエは、植物の成長のもとが、土の中ではなく、空気中の二酸化炭素にあると考えた。ただし、彼は、植物は、二酸化炭素をとかした水を根から吸収するものと考えた[もちろん誤っている。植物は一般に葉から二酸化炭素を吸収する]。)
⑤ソシュールの実験:光合成によって、空気中の二酸化炭素が取り込まれる。
(雑談:ソシュール[スイスの植物生理学者]は、植物体の炭素源が根から吸収されるというセネビエの説に疑問をもった。そして、ソラマメが空気中の二酸化炭素を吸収して炭素を得ていることを示した。)
⑥ザックスの実験:植物は日光によってデンプンをつくっている。
(雑談:ザックス[ドイツの植物生理学者]は近代植物生理学と植物栄養学の創始者とされる。)
*以下の動画の光合成のところで少し光合成の解明の歴史について話している。
講義動画【生物学史】
まだわかっていないこと
● 光化学系の構造(色素やタンパク質の配置・機能など)について、完全には明らかになっていない。
● 地球のCO2濃度の上昇は、C3植物とC4植物の生育にどのような影響を与えるのか。
● 水中に生息する藻類と陸上植物では、光合成の機構はどのように異なるのか。
● コケ植物の光合成の機構は、種子植物とどう異なるのか。
● 光呼吸の意義は何か(活性酸素の原因となる過剰な酸素の消費、ADPとNADP+の再生のためとも言われている)。