
【高校生物】植物生理③「植物は花を咲かせる時期をどのように知るのか?」

~プロローグ~
「そもそも、花といふに、万木千草において、四季折節に咲くものなれば、その時を得てめづらしきゆゑに、もてあそぶなり。」世阿弥『花伝第七 別紙口伝』より
「目にも見えず、言葉にもならぬ美を湛えたる至上の神から摂理が下り地上の万物を満たしたることを、プラトン主義者プロティノスは木々の葉、木々の花をもって証明する。曰く、これら儚き命が、かくも完璧にかくも繊細に仕上げられた美を有するとは、それが神より降り下りたる証拠でなくて何であろう。」聖アウグスティヌス『神の国』より
植物は、花を咲かせる季節を、どのように知るのだろうか?長い間植物学者は「植物は、日が長い時期に光合成産物を蓄え、それを用いて花を咲かせるのだ」と考えていた。この仮説が正しくないことは、1920年、メリーランド・マンモスを用いた実験により示された。この植物は、夏の一般的な条件の下では、旺盛に成長して5mもの高さに達するものの、花を咲かせることはない。しかし、秋に挿木して温室で育てると、わずか1.5mで開花し、種子を付ける。また、あるダイズの品種についても、不思議な現象が発見されていた。5月から7月にかけて、2週間ごとにこの品種の種を蒔いたところ、どの時期に種を蒔いても、花は9月になって”一斉に”現れた。あたかも、ダイズは「花を咲かせるための合図」を待っていたかのようであった。
★テストに出やすいワード
①限界暗期
②フロリゲン
③フィトクロム
④クリプトクロム
⑤フォトトロピン
要点:短日植物は限界暗期より短い暗期で花芽形成を行う。
(1)光周性
● 連続した暗期(自然界では「夜」)の長短によって、花芽(かが)の形成などが左右されることが知られている。このような性質を光周性(こうしゅうせい)という。
*光周性:1日の明期・暗期の長さ(光周期)に対する生物の応答あるいは応答性。
*花芽(かが、はなめ):展開して花になる芽。
雑談:花芽形成では、頂端分裂組織が花序分裂組織へと転換し、花序分裂組織から花芽分裂組織が分化してくる。花芽分裂組織では、がく、花弁、雄しべ、雌しべなどの花器官原基が分化する。その後、成熟した花芽になり、最終的に花芽が開いて開花に至る。
雑談:今からおよそ100年前、アメリカの農務省に努めるGarnerとAllardは、タバコやダイズが、アメリカ北部では開花しない原因を研究し、これらの品種が日長が一定以下になると花芽を形成し開花することを見出した。これが光周性の発見となった。
● 花芽形成と日長との関係から、種子植物は、短日植物・長日植物・中性植物に分けられる。
(2)短日植物
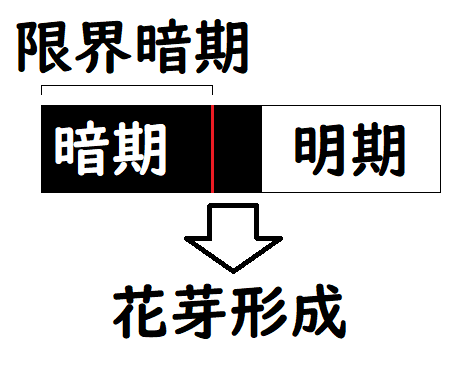
・短日(たんじつ)植物・・・暗期が限界暗期以上で花芽形成。
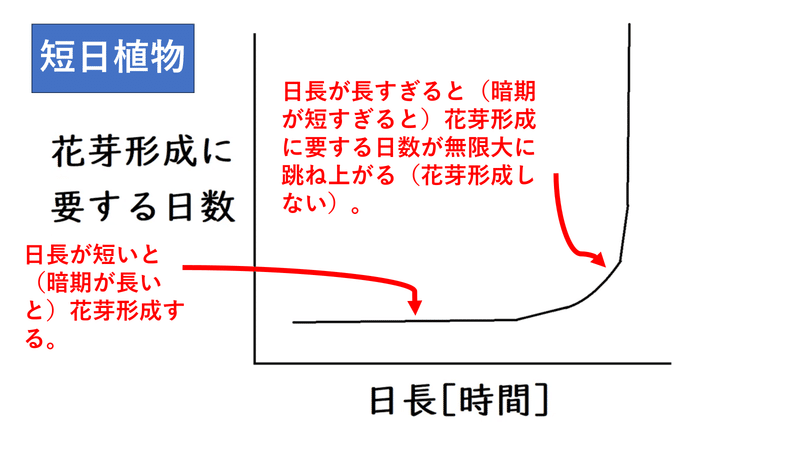
・短日植物の「花芽形成に要する日数」と「日長」のグラフは以下のようになる。日長が短い(暗期が長い)と花芽形成する。
短日植物の例)アサガオ、キク、コスモス、オナモミ、イネ、ダイズ
語呂「朝はキックで起こすてくれる気の短い女がいーね大好き(アサガオ、キク、コスモス、短日、オナモミ、イネ、ダイズ)」
注意!!:短日植物とは、「限界暗期の短い植物」ではない!限界暗期の長さは植物の種類ごとにおおよそ決まっており、「その時間の長さ(限界暗期の長さ)より暗期が長くなると花芽形成する」のが短日植物である。
雑談:なお、高校生は全く気にしなくてよいが(テストでは無視する)、ほぼすべての植物は、一定以下の日長では開花しない(そのような日長を「下位の限界日長」と言う)。したがって、一定限界以下の日長で開花しないのは長日植物だけではない。ただし、短日植物の「下位の限界日長」は非常に短く、ふつうは5時間以下である。そのような状況は、植物が生活するような自然界にはない日長である。短日植物の「花芽形成に要する日数」と「日長」のグラフは、本当はU字型になる(日が長すぎても花芽形成しないし、日が短すぎても、すなわち下位の限界日長を下回っても、花芽形成しない)。上で示したグラフは、本当のグラフを部分的に切り取ったものである。
(3)長日植物
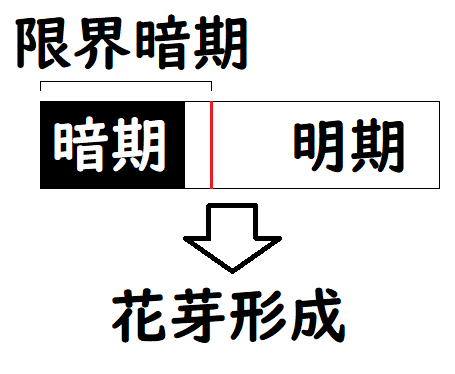
・長日(ちょうじつ)植物・・・暗期が限界暗期以下で花芽形成。
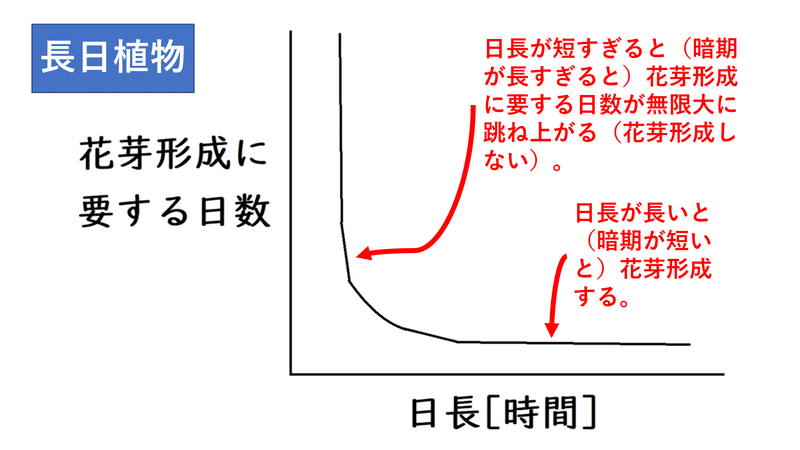
・長日植物の「花芽形成に要する日数」と「日長」のグラフは以下のようになる。日長が長いと(暗期が短いと)花芽形成する。
長日植物の例)アヤメ、コムギ、アブラナ、ダイコン、ホウレンソウ、シロイヌナズナ
語呂「アヤメ、長い小麦色の危ない大根足放り出すな(アヤメ、長日、コムギ、アブラナ、ダイコン、ホウレンソウ、シロイヌナズナ)」
*限界暗期の長さは植物によって異なるが、10~14時間程度のものが多い。

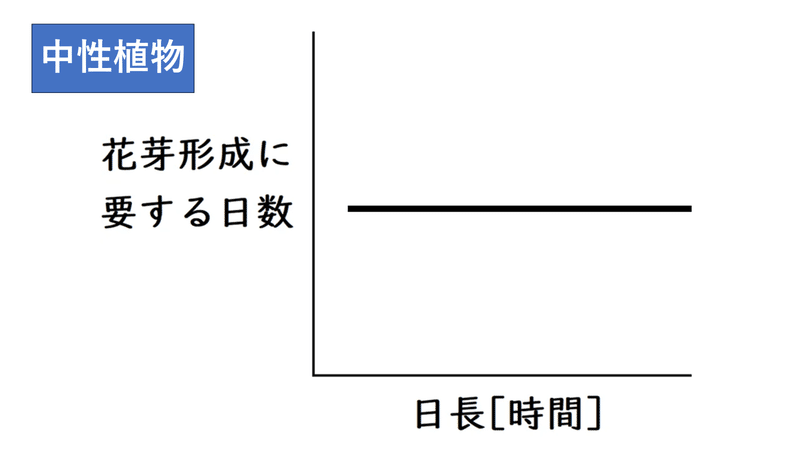
(4)中性植物
・中性植物・・・日長と無関係に花芽形成が起こる。
中性植物の例)トマト・ヒマワリ・セイヨウタンポポ・エンドウ・ソバ・トウモロコシ・ナス・キュウリ・メロン
覚え方「おいしいのが中性植物(トマト・エンドウ・ソバ・トウモロコシ・ナス・キュウリ・メロン)」
・中性植物の「花芽形成に要する日数」と「日長」のグラフは以下のようになる。日長と無関係に花芽形成が起こる。
(5)光中断
● 短日植物では、短日処理中の暗期をごく短時間の光照射で中断(光中断)すると、花芽形成が起こらなくなる。このことから、花芽形成には一定時間以上の連続した『暗期』が必要であることがわかる(光中断の前後で、明期の長さは変えていないことに注意)。
この時の反応には光受容体であるフィトクロムが関わる。暗期中に赤色光を当てると光中断の効果が現れるが、すぐに遠赤色光を当てるとその効果は打ち消される。
下図は光中断のイメージ(極端に描いている。実際は光中断は短時間である)。
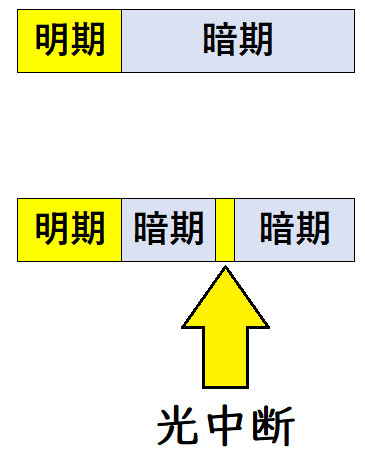
雑談:暗期の適当な時期に短時間の光を照射すると、期待される光周性の効果と逆の結果がもたらされる場合がある。その時、この光処理を光中断(light break, light interruption, night break)という(「光中断」という名前が非常にわかりにくい。僕は高校生の頃「光を消すことを光中断と言うんだろうな」と勘違いしていた。光中断は、 英語では「night break」の語が用いられることが多く、「暗中断」の訳語を当てることもある。入試では書いてはいけないが、このほうが自然な表現であるように思う)。
講義動画【短日植物・長日植物】
雑談:厳密に短日植物、長日植物、中性植物を分けることはできない。中性植物と呼ばれるものでも、長日か短日のどちらかによって開花が促進される傾向をもつことがある。また、「短ー長日植物」や「長ー短日植物」など、正しい順番で日長条件が必要な植物もいる。
雑談:植物は暗期の長さを感知しているのであるから、厳密には短日植物ではなく長夜植物とすべきである。しかし、ふつう地球上では光周期は24時間サイクルであり、我々は夜寝てしまうので、夜の長さはイメージしにくい。そのため、短日植物、長日植物の語が定着した。
雑談:開花までのステップには、
①催花(栄養成長からの転換)②花芽創始③花芽分化④花芽発達⑤花芽成熟⑥開花がある。②~④を花芽形成と呼ぶことが多い。そして、植物によって、それぞれの段階で、どのような日長条件のシグナルをどの程度キャッチするかは異なる。ただし、開花に特別な条件が必要な場合を除き、花芽が形成されれば、すみやかに開花が起こると考えてよい。
問題:以下のような性質を示す植物は短日植物かと長日植物のどちらか。
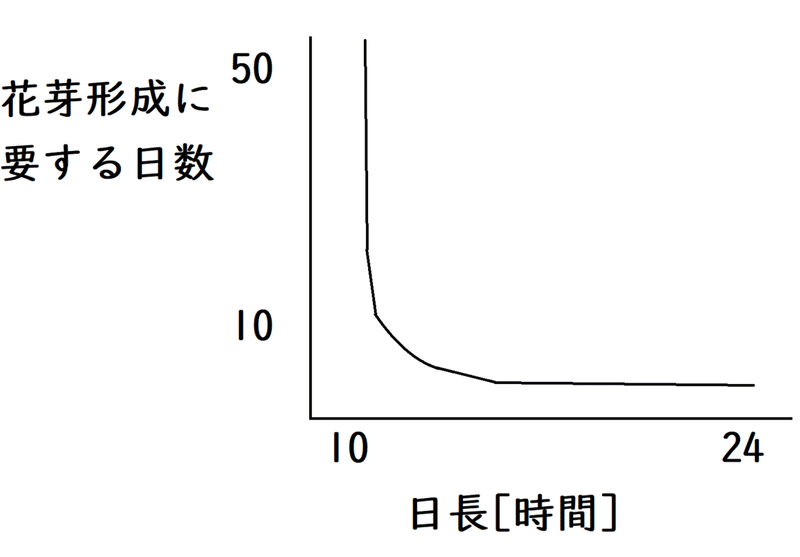
答え:長日植物
解説:横軸が暗期ではなく日長(日の長さ)になっていることに注意。日長が短いと、何日たっても花芽形成しない(短い日長で、グラフが上に跳ね上がっているのは、ものすごい日数その条件で育てても、花を咲かせないことを意味する)。つまり、日長が長くないと花芽形成しない→長い日長で花芽形成→長日植物。
講義動画【花芽形成に関するグラフ】
要点:花成ホルモン(フロリゲン)は葉で作られ師管を通る。
(1)フロリゲン
● 花芽形成の促進にはたらく物質であるフロリゲン(花成ホルモンとも呼ばれる)は、一定時間の暗期が条件となって葉でつくられ、師管を通って茎頂に運ばれる。(葉でつくられたたフロリゲンは全身に輸送される)。
*まだわかっていないことも多い。
雑談:茎頂は、ざっくり茎の先端と考えればよい(厳密には、茎頂はシュートの最先端部分である。茎頂分裂組織と、これに付随する若い葉、および若い茎の組織からなる)。
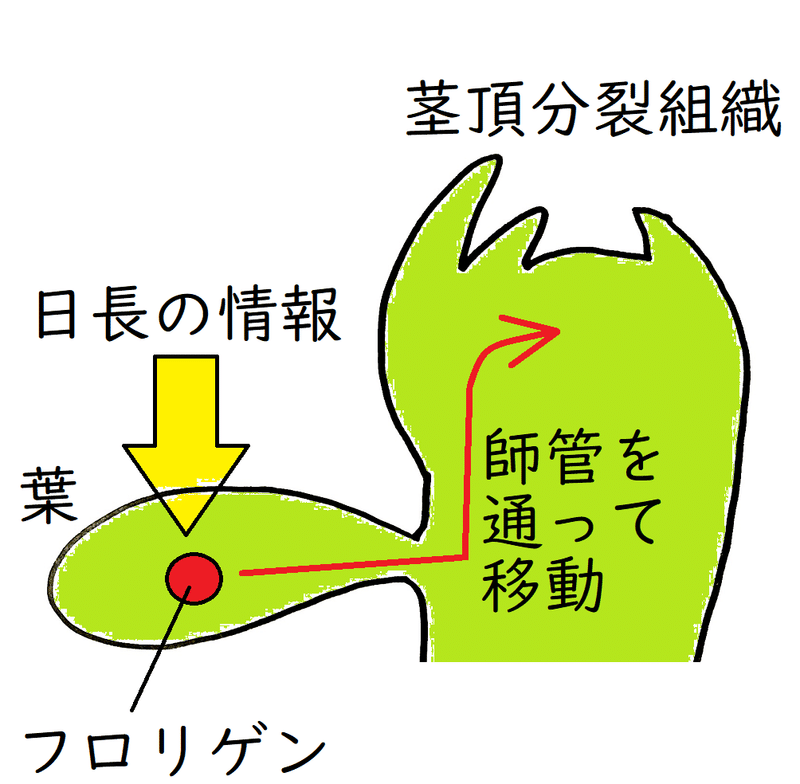
*上図では、葉が「日長の情報」を感知するように描いている(一般にそのように表現される)が、日長の情報を植物は「暗期の長さ」で測定していると考えられているので注意せよ。
● 植物は、一般に、葉で日長を感じ取る。
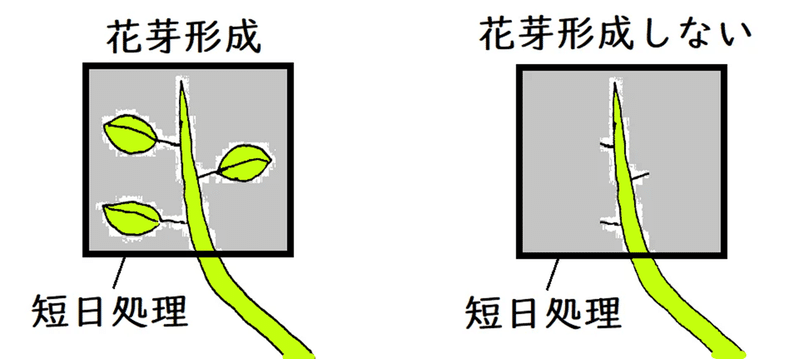
● 葉のついた短日植物の枝を短日処理(人工的に連続暗期を限界暗期よりも長くする処理)すると、花芽が形成される。しかし、葉を除いた枝を短日処理しても、花芽は形成されない(葉がなくては日長を感じ取れない)。
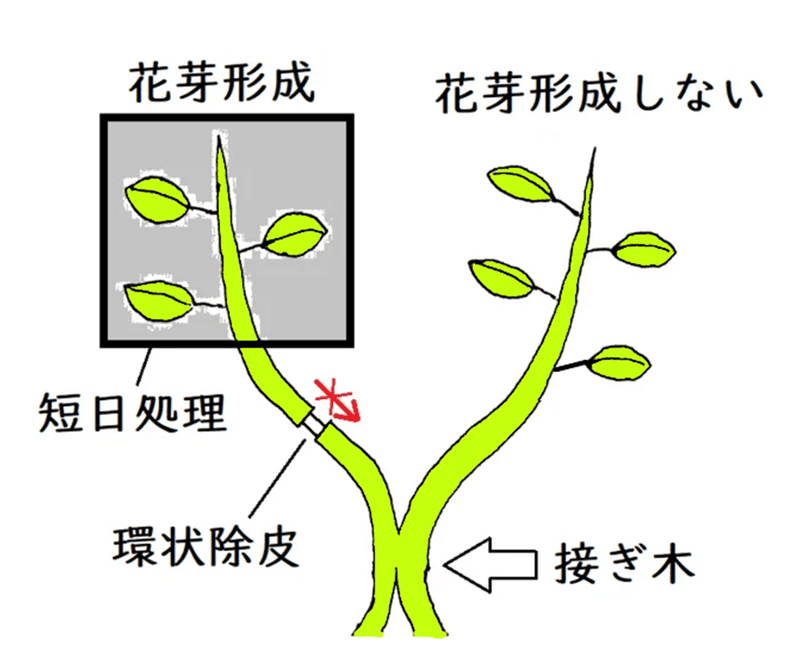
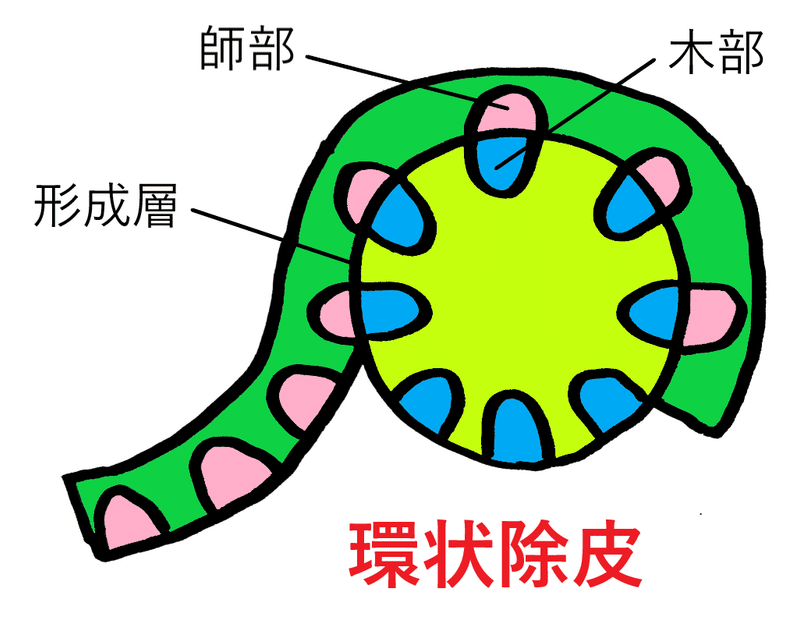
● 環状除皮(かんじょうじょひ)によって師部を除くと、そこでフロリゲンの移動は止まることから、フロリゲンは師管を移動すると考えられるようになった。
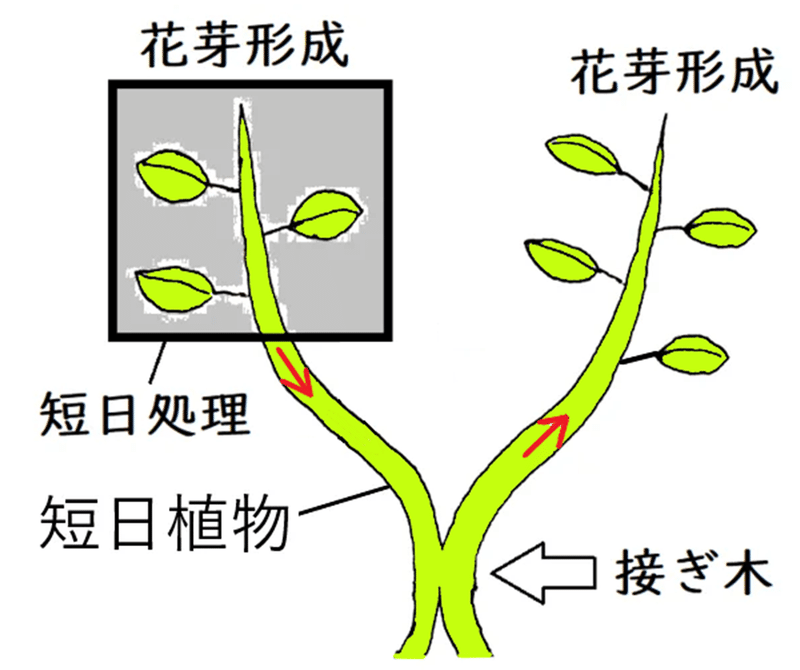
● <下図>短日植物を接木(つぎき)する。片方の枝を短日処理すると、短日処理してない方(長日条件下)の枝にも花芽が形成される。これは、フロリゲンが、短日処理された方の枝の葉から、(短日処理していない)もう片方の枝に(師管を通って)輸送されたためと考えられる。
*接木:植物の二つ以上の部分を組織間の接着と癒合によって一体化させる方法。
<下図>しかし下図の所で、師管を環状除皮によって取り除くと、短日処理していない方の枝では花芽が形成されない(環状除皮によって師管が除去されたため、フロリゲンの移動がそこで停止したと考えられる)。
*赤矢印は、この実験から推測される、フロリゲンの移動方向のイメージ(フロリゲンの実体・輸送・作用機序については、わかっていないことが多い)。
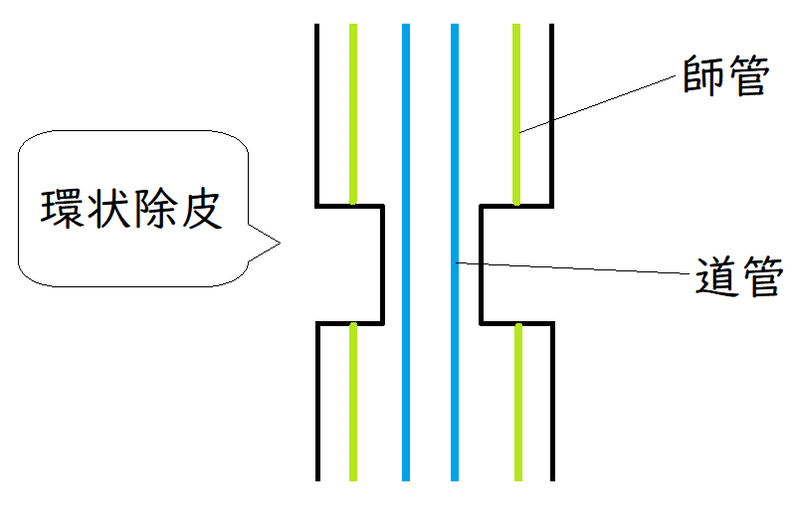
*茎の形成層(師部と木部の間にある)より外側をはぎ取る操作を環状除皮という。環状除皮を行うと、道管(木部にある)は残るが師管(師部にある)は除かれる。環状除皮は師管を通した物質の輸送を妨げる目的で行われる。下図はイメージ。
雑談:短日植物と長日植物を接木(たとえば短日性のタバコと長日性のタバコを接木)して長日条件に置くと、短日植物にも花芽ができることがある。このことから、短日植物と長日植物のフロリゲンは相互に置き換え可能であると考えられる。
雑談:チャイラヒャンは、葉で花成ホルモンが作られ、これが茎頂に輸送されることによって花成が始まると言う説を提唱した。この説は、「花成を誘導した個体を別の個体に接木すると、接木された個体は、非誘導的な光周条件下でも花成が誘導される」という事実によって支持された(異なる種の間の接木でも同じ結果が得られることから、フロリゲンは種に特異的ではないとされる)。フロリゲンの研究は困難を極め、その単離の難しさから、「幻のフロリゲン」とも呼ばれた。僕が高校生の頃、色々な本に「フロリゲンはまだ発見されていない」と書いてあったのを覚えている。
雑談:ある物質をフロリゲンとみなす条件には、以下のようなものがある(ただし議論は続いている)。 ①花芽形成を誘導する日長依存的に葉で合成される。 ②師管を通って茎頂に輸送される。 ③茎頂で花芽形成を引き起こす。 ④他の植物ホルモンのように植物種を超えて共通である。 ⑤接木面を介して移動できる。
● 近年、短日植物であるイネではHd3a(Heading date 3a)とよばれるタンパク質が、長日植物であるシロイヌナズナではFT(FLOWERING LOCUS T)とよばれるタンパク質が、それぞれフロリゲンの実体であることが明らかになった。
*FTやHd3aは、茎頂分裂組織に移動し、花芽形成を誘導すると考えられている。
*シロイヌナズナは長日植物、イネは短日植物である。
語呂「太った白い犬(FTタンパク質、シロイヌナズナ)」

雑談:2005年、シロイヌナズナのFT遺伝子の産物が、フロリゲンの実体であることが提唱され、2007年には、イネにおけるFT相同タンパク質である Hd3aが葉から茎頂分裂組織へ輸送されることが示された。
雑談:日長に応じて葉で作られたFTタンパク質は、茎頂分裂組織で待ち受けるFDタンパク質と複合体(FT-FD複合体)を形成し、花芽形成を開始すると考えられている。
雑談:シロイヌナズナの場合、長日条件におけるCO遺伝子という遺伝子の発現が、FTmRNAの増大をもたらすことが明らかになっている。すなわち、FT遺伝子はCO遺伝子の下流の標的遺伝子である。
講義動画【フロリゲン(花成ホルモン)】
(2)春化処理
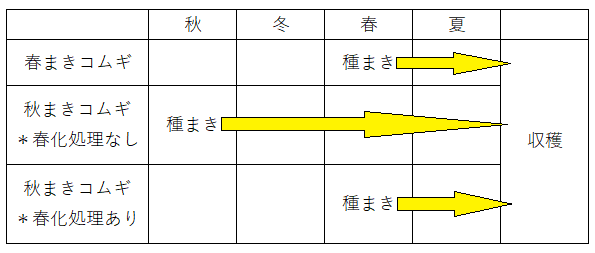
● 秋まきコムギの開花・結実には、冬の低温に一定期間さらされることが必要である。低温状態の経験によって花芽形成などが促進される現象を春化(しゅんか、バーナリゼーション)という。人為的に低温状態におくことで花芽形成を促進することを春化処理という。
*春化処理:花成を誘導するために、植物を人為的に一定期間低温で処理すること。
*春化処理を行うことにより、秋まきコムギを春にまいて収穫することができる(つまり、秋まきコムギに、人為的に冬と同じような低温を経験させることで、冬を経なくても収穫できるようにすることができる)。
雑談:ロシアのT.D.Lysenko(ルイセンコ)は、秋まきコムギを低温処理することにより春まきコムギの性質をもたせることができると主張した。ルイセンコはこれを獲得形質の遺伝と解釈し、小麦の播き性を人為的に変化させ遺伝的に固定させ得るという説を提唱した(このルイセンコ説は誤りである。一般に、獲得形質は遺伝しない。ルイセンコは本来の遺伝学を攻撃し、ロシアの生物学の発展を遅らせたと言われている)。
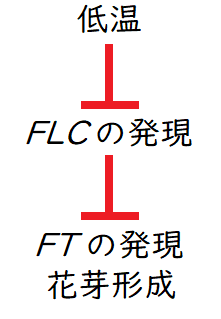
雑談:シロイヌナズナでは、MADSボックスをもつ開花抑制遺伝子(FLC[FLOWERING LOCUS C])が、春化処理により春化処理により様々なヒストン修飾を受ける(コムギでは、別の遺伝子が春化処理によりエピジェネティックな遺伝子発現調節を受けていると考えられている)。その結果、発現が抑制される。すると、下流のFT遺伝子(FTタンパク質はフロリゲンの実体である)の発現が誘導され、花芽形成に至ると考えられている。下図はイメージ(春化処理の詳細については未解明である)。
*エピジェネティクス:DNAの塩基配列に変化を起こすことなく、DNA複製・細胞分裂を経て伝達される遺伝子機能の変化、ならびにその制御機構を研究する学問領域のこと。
要点:光発芽種子の発芽には光が必要である。
● レタスやタバコの種子は、発芽に光を必要とする。このような種子を光発芽種子(めちゃめちゃよく問われる)という。

● 光発芽種子に赤色光を当てると発芽が促進されるが、遠赤色光を当てるとその効果が打ち消される(太陽光には様々な波長の[色の]光が含まれている。一方、葉を透過してきた光には、遠赤色光が多く含まれる[葉で光合成に使われる光の色は『赤』と青であったことを思い出そう。赤色の光は葉で吸収され、光合成に利用されてしまう]。つまり、種子は、遠赤色光を多く浴びる時、自分のいる位置が『葉の下』であり、そこで発芽しても光不足で死んでしまう可能性が高いことを察知しているのである)。
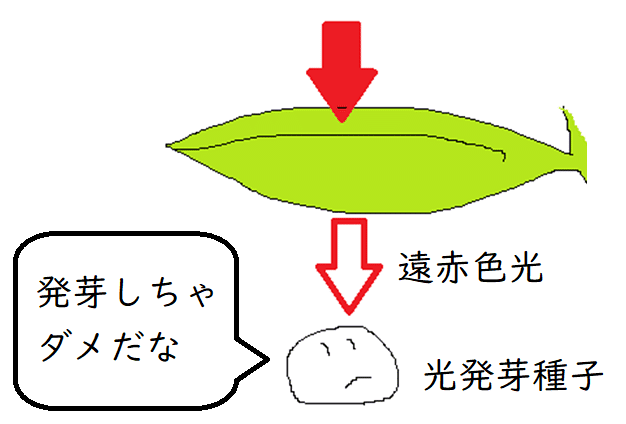
*光発芽種子には種子の中の栄養分が少ない小形の種子が多い。このような種子は、他の植物の陰になっている場合など光が当たらない環境で発芽を控えることで、芽ばえの生存率を上げていると考えられる。
● 赤色光と遠赤色光の光を交互に種子に照射した場合、最後に照射した光が赤色光であれば発芽が促進され、遠赤色光であれば(赤色光を照射した場合に比べて)発芽が抑制される。
この実験から、2つの型を行き来するような光受容体の存在が予測されていた(まるでON⇄OFFを繰り返すスイッチのように)。やがてそのような性質をもつ光受容体フィトクロムが発見された。
● カボチャやケイトウなどの種子は、光があると発芽しないので暗発芽種子とよばれる(その意義については様々な説がある。乾燥が激しい場所での発芽を抑制している可能性がある)。
● 発芽に低温を要求する種子(低温要求種子)もある。
雑談:リンゴなどの種子は発芽のために低温を要求する低温要求種子であり、採種した時点ですでに形態的には完成しているが、生理的には未熟であり、そのままでは好条件に置いても発芽しない(発芽が抑制される原因は、植物ごとに様々である)。一部の種子が冬の低温を受けた後に発芽することは、古くから農家で知られていた。発芽に低温を要求する種子は亜寒帯に原生する植物の種子に多い。そのような寒い地域に住む植物の種子は、土壌の中で冬を越し、春に発芽する。そうすることで、冬前に発芽して幼植物が冬の寒害を受けるような事態を避けることができる。
要点:植物は、いくつかの光受容体によって光刺激を受容している。
(1)フィトクロム
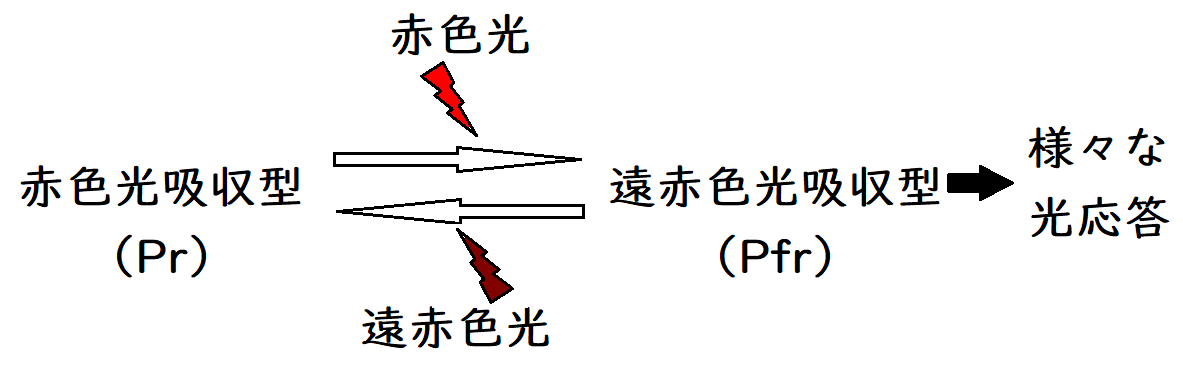
● 光発芽種子や花芽形成における光条件の感知には、フィトクロム(phyto=植物、chrome=色素)とよばれる色素タンパク質が光受容体(光エネルギーを吸収して生物に一定の機能を果たす物質の総称)として関与している。光周性における光の感知にもフィトクロムが関与している。
● フィトクロムには、赤色光吸収型(Pr)と遠赤色光吸収型(Pfr)がある。遠赤色光吸収型のフィトクロムは、遠赤色光(波長730nm)をよく『吸収』して赤色光吸収型に変化する。同様に、赤色光吸収型のフィトクロムは、赤色光(波長660nm)をよく『吸収』して遠赤色光吸収型に変化する。
*赤色光を当てると赤色光吸収型が増えるのではないことに注意。
*生理学的にはPfr型が活性型で、Pr型は活性をもたないとされる。
*英語で赤色光は「red light」、遠赤色光は「far red light」と書く。
下図はイメージ。
(2)クリプトクロム
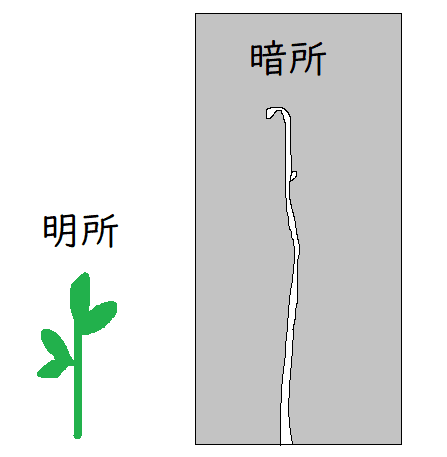
● クリプトクロム:青色光を受容する色素タンパク質。茎の伸長成長の抑制に関与(長年性質が不明だったため、隠れた=クリプト、色素=クロムから名づけられた)。
詳細:クリプトクロムは光形態形成(環境の光情報によって発生や分化の過程が制御される現象)に関わる。薄暗いところで発芽した植物は「もやし状」になるが、光を照射すると「もやし状」になる反応が阻害される(下図はイメージ。暗所で発芽させた植物は緑色にならず、黄白色でひょろひょろと長くなり、葉も発達しない)。
クリプトクロム欠損突然変異体では、明るいところでも、青色光を感知できず、「もやし状」になる。
*光がない所で太くなったり、葉を展開したりしても、無駄である。暗い所では、まずはひょろひょろと細く伸びて、明るい所に出た方がよい。
*もやし状=胚軸が長い。子葉を広げない。子葉に葉緑体が発達しない(黄色になる)。
雑談:クリプトクロムは動植物の体内時計の調節にも関与する。
語呂「もやしと栗と(モヤシ状、クリプトクロム)」
雑談:スーパーなどでよく「モヤシ」を見かけるだろうが、「モヤシ」という植物種があるわけではない。種子が、暗所で、十分な水を与えられて生育すると「モヤシ」になるのである。市販されている「モヤシ」は、ダイズなど種子に十分な水を与えて暗所で発芽させてしばらく成長させたものである。イネやムギの種子からも「モヤシ」は得られる。
(3)フォトトロピン
● フォトトロピン:青色光を受容する色素タンパク質。光屈性に関わる光受容体はフォトトロピンである(「フォトトロピン」という名の由来はphoto[光]tropis[屈性]である)。
● フォトトロピンは他にも気孔の開口や、葉緑体光定位運動(葉緑体が光環境に応じて細胞内を移動する現象)にも関与していると考えられている。
雑談:フォトトロピンが、オーキシン輸送体の配置(局在)を、具体的にどう制御しているのか、わかっていない(フォトトロピンは、細胞膜の細胞質側に結合している。フォトトロピンが光を受け、活性化されると、オーキシンを排出する輸送体のはたらきを阻害するようになるらしい。ただし、様々なことがよくわかっていない)。
(4)まとめ

講義動画【植物の光受容体】
<Q.短日植物って限界暗期が短い植物?…ちがう。限界暗期の長さは「短日」「長日」の区分けと関係ない。植物ごとに固有に決まっている。「限界暗期」の長さより短い暗期で花芽形成する(長日植物)か、「限界暗期」の長さより長い暗期で花芽形成する(短日植物)かで判断する。>
<Q.光中断って光照射を中断すること?…ちがう。暗期の中で短時間光を照射すること。暗期を「光」で「中断」することである。>
雑談
要点:ABCモデルは花の形成についてのモデルである。
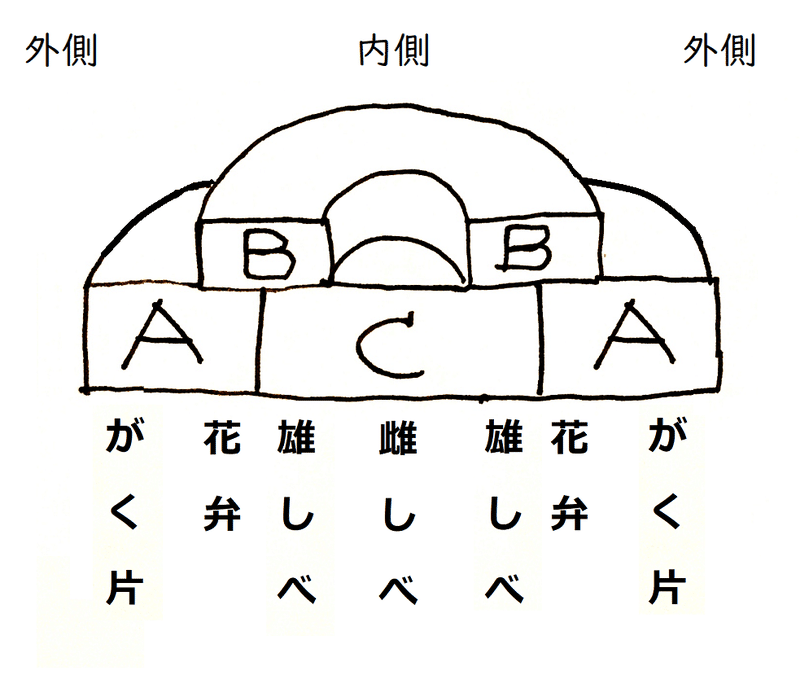
● 下図は仮想的な花の構造である。がく片、花弁、雄しべ、雌しべだけチェックすればよい。
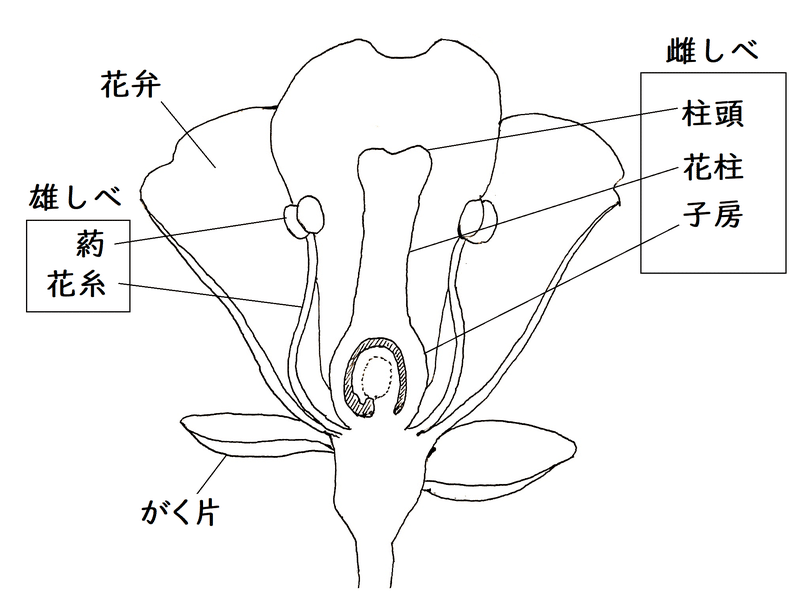
雑談:萼(がく)片は一般に緑色か目立たない色であり、葉状で、本来、つぼみを守る働きをする。それらは通常、開花後すぐに落ちてしまう。
「ぼんやり窓の外を眺めていた。窓一ぱいにあんなに見事に咲いていた桜の花も、おおかた散ってしまって、いまは赤黒い萼だけが意地わるそうに残っている。」太宰治『正義と微笑』より 芹川進の言葉
● 様々な花器官決定に関わる遺伝子が、3つのクラス(A、B、C)に分類されている。
● 茎頂分裂組織の同心円状の各領域ではたらく3つのクラスの遺伝子(A、B、C)の組み合わせによって、分化する構造の種類(がく片・花弁・雄しべ・雌しべ)が決まってくる(同心円状の領域が想定され、外側から内側に向かい、がく片・花弁・雄しべ・雌しべが形成される)。
*「がく片」の集まりを「がく」という。
● ABC遺伝子はホメオティック遺伝子(体のどこに何を作るのかを制御する)である。
上の図は、AとCの台に乗ったBというバームクーヘンを切った断面のようなイメージ。
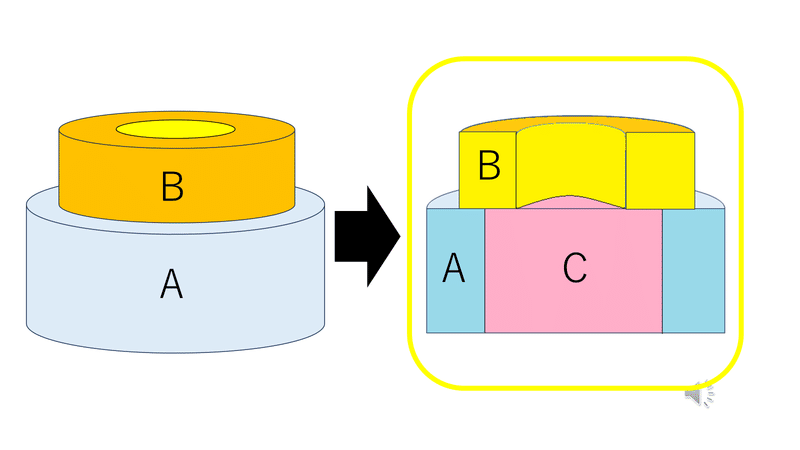
● クラスAは単独でがく片を形成する。
● クラスAとBは花弁を形成する。
● クラスBとCは雄しべを形成する。
● クラスCは単独で雌しべを形成する。
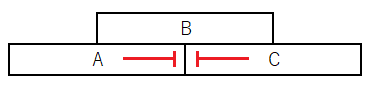
*クラスAとCは互いに相手を抑制し合う(つまり、AとCは器官の属性決定における機能に加えて、相手の発現を抑制しその活性を排除する機能をもつ)。
*下図は、A、B、C遺伝子が発現する領域と、形成される構造のイメージ。
これから、A、B、C遺伝子の内の1つの活性を喪失させた場合に、図の赤い所に何ができるかを見てく。
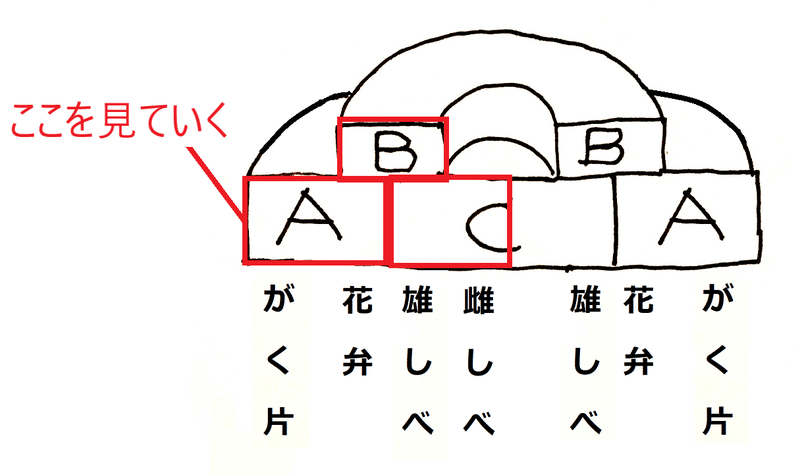
*下図は、A、B、C遺伝子の内の1つの活性を喪失させた時にできる花の構造のイメージ。
クラスC活性喪失変異体では、外側から内側に向かって
「がく片・花弁・花弁・がく片」ができる。
● クラスAは単独でがく片を形成する。
● クラスAとBは花弁を形成する。
ことを思い出そう。
クラスA活性喪失変異体では、外側から内側に向かって
「雌しべ・雄しべ・雄しべ・雌しべ」ができる。
● クラスCは単独で雌しべを形成する。
● クラスBとCは雄しべを形成する。
ことを思い出そう。
クラスB活性喪失変異体では、外側から内側に向かって
「がく片・がく片・雌しべ・雌しべ」ができる。
● クラスAは単独でがく片を形成する。
● クラスCは単独で雌しべを形成する。
ことを思い出そう。
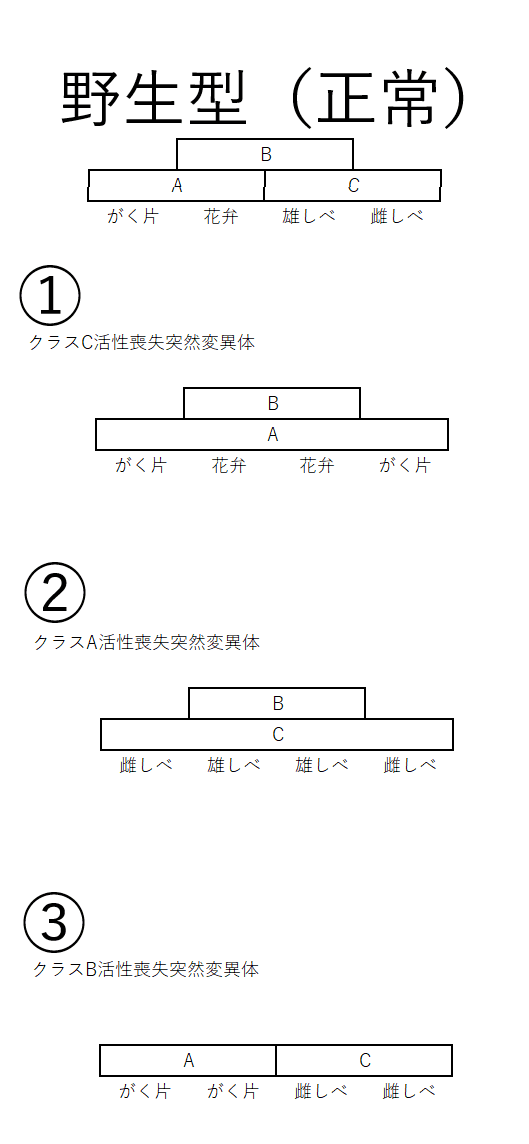
雑談:今では、胚珠形成に必要なクラスD遺伝子や、クラスA・B・C遺伝子の活性に必要なクラスE遺伝子が発見されている。クラスE遺伝子にはSEP1~SEP4の四つの遺伝子があり、これらの四重突然変異体は、すべての領域に葉が形成される(これは、クラスA遺伝子、B遺伝子、C遺伝子のすべてが働かない突然変異体と同様の変化である)。下図はクラスE遺伝子の発現領域のイメージ(4つの領域で発現している)。
クラスA、B、C、D、E遺伝子が、下流の様々な遺伝子の発現を制御していると考えられているが、その詳細はほとんど明らかになっていない。
雑談:クラスC遺伝子には、茎頂分裂組織での新たな細胞の形成を抑制する働きもあることが知られている。クラスC遺伝子の突然変異により、「がく片ー花弁ー花弁」の単位が繰り返されることがある(外側から内側に向かって、がく片ー花弁ー花弁ーがく片ー花弁ー花弁・・・となる)。八重咲の花(本来より多く花弁を持っている花)の中には、クラスC遺伝子の突然変異が原因と考えられるものもある。
雑談:シロイヌナズナの花芽分裂組織は、4種類の花器官(がく片、花弁、雄しべ、心皮)の形成を開始するが、これらの器官の発生は、「環域(ワール whorl)」と呼ばれる環状領域で始まる。シロイヌナズナでは、4つの環域は以下のように配置されている。
・第1環域(最も外側):4枚のがく片。
・第2環域:4枚の花弁。
・第3環域:6本の雄しべ(その内2本は他の4本より短い)。
・第4環域(最も内側):雌しべ(単一の複合器官[雌の生殖器官])。雌しべは、多数の胚珠を含む2枚の心皮が融合してできた子房、花柱、その上の柱頭からなる。
下図はイメージ。
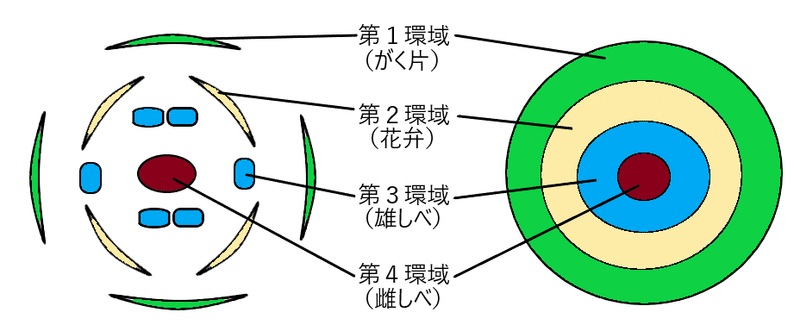
右:4つの環域の模式図。
雑談:シロイヌナズナでは、Aクラス遺伝子にAP1、AP2が、Bクラス遺伝子にはAP3、PIが、Cクラ ス遺伝子にはAGが同定されている。なお、これらのほとんど(AP2以外)はMADSボックス(マッズボックス)と呼ばれる領域(ショウジョウバエのホメオティック遺伝子がもつホメオボックスとは異なる)を持つMADSボックス遺伝子である。
雑談:ABCモデルがどの程度普遍性をもっているかについては、今まさに研究が進んでいる所である。ただし、A、B、Cの各活性をもった遺伝子は、シロイヌナズナやペチュニアをはじめ、イネ科を含む単子葉植物からも見つかっている。そして、その発現領域はABCモデルから期待される通りの領域で発現している。また、チューリップのようにがく片が花弁化している花では、Bクラス遺伝子が一番外側の領域でも発現していることがわかっている。
講義動画【ABCモデル】
発展:植物のからだ
(生物基礎の資料第1講にも植物のからだについての詳しい資料がある。)
(1)組織系
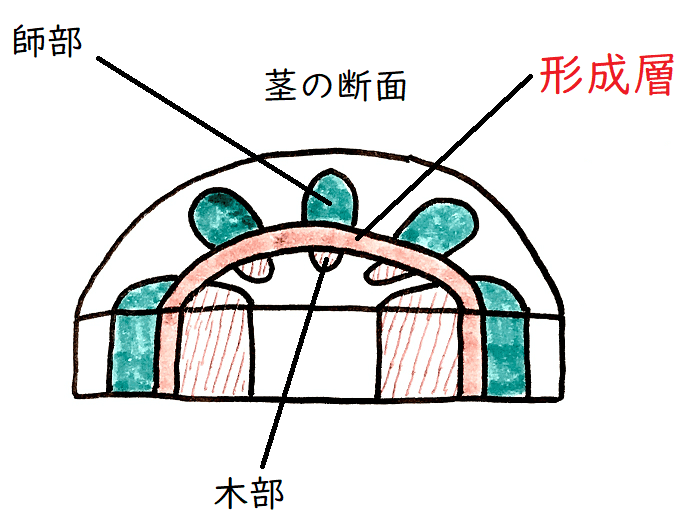
● 維管束植物には3つの組織系、すなわち①表皮系②維管束系③基本組織系がある。
*組織系:関連のあるいくつかの組織の集団。ふつう、維管束植物についての議論で使われる用語。
● 表皮系を構成する組織は、単に表皮と呼ぶことが多い。表皮細胞はクチクラ(動物の同様の働きをする構造を呼ぶ場合はキューティクルということが多い)で覆われている。クチクラには、水の損失を防いだり、病原体から植物体を保護したりする役割がある。 表皮細胞には葉緑体がない(虫に食われ、無駄になることが多いためか)が、孔辺細胞には葉緑体がある(気孔開閉に関わる生命現象を活発に行うためか)。
● 維管束系は木部と師部からなる。木部の道管は主に水や無機塩類が移動する通路、師部の師管は主に葉の同化物質が移動する通路である。
● 表皮系と維管束系以外を基本組織系という。柔組織、厚壁組織、厚角組織などから成る(厚壁組織や厚角組織の細胞の細胞壁は厚い)。
雑談:厚壁組織は、梨のざらざらした舌触りの原因になっている。
雑談:厚角組織や厚壁組織などの集合を機械組織という(植物体を「機械」的に支持する)。
(2)分裂組織
● 維管束植物には、2つの分裂組織がある。
①頂端分裂組織:茎頂分裂組織と根端分裂組織。伸張成長に関わる。
②形成層:木部と師部の間にある。肥大成長に関わる。
雑談:植物では、体細胞分裂を行う幹細胞の位置が明確である。対して、動物では、幹細胞は体の色々な部位に散在する(動物の成体の幹細胞については、まだまだ解明されていないことが多い)。
雑談:形成層の細胞が分裂して師部と木部がつくられるが、師部の方は壊され、木部の方が蓄積する。木部の部分が肥大することで、木は太くなる(木部には、"木"の本体の"部"分といった意味が込められている)。その際、季節によって増殖する細胞の大きさや数に差が出る(形成層の活動は温度などの環境要因に支配されやすいことが知られている)ので、温帯の地域の木の多くには、一般に、明瞭な年輪が見られる(明瞭な季節が存在しない熱帯の樹種には年輪が認められないものが多い)。
講義動画【植物の体】
雑談:基本組織系はザックス(光合成でデンプンができることを発見した植物学者)が命名した。維管束植物について、表皮系と維管束系を除いたすべての部分を指す。
雑談:木部には、木部柔組織などが含まれる。木部柔組織はデンプンや樹脂を多く含んでおり、貯蔵の枠割をもつと言われている。
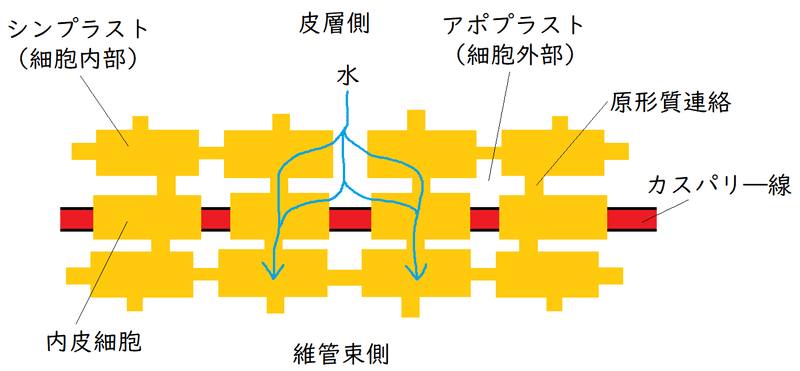
雑談:内皮は、カスパリ―線を持つ特殊な細胞層であり、根の維管束を取り囲んでいる(各内皮細胞の上下と側面の壁にカスパリー線は存在する)。カスパリー線は疎水性のスベリンを多量に含むため、水溶液を透過させない。内皮細胞の間にカスパリ―線が存在するため、水と溶質は、内皮を横切るために、必ず細胞膜を通過しなければならない(水は一度は必ず内皮細胞を通らなければならない)。 いったん維管束内に入った水や溶質は、カスパリ―線があるため、外に出ることはできない。それにより、根圧が生じると考えられている。下図は根の断面のイメージ。
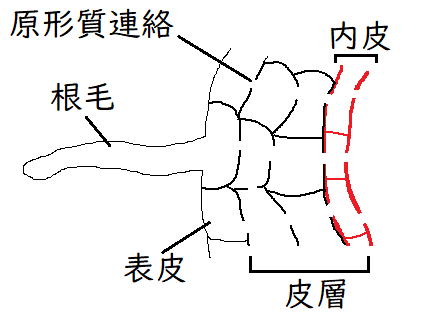
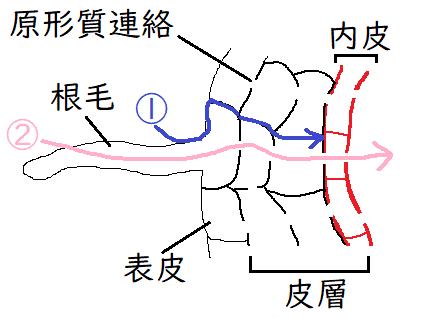
雑談:根の表皮から内皮まで水が流れる経路には、アポプラストとシンプラスとがある。アポプラスト、シンプラストの相対的な重要性についてはよくわかっていない。
①アポプラスト経路:膜を横断することなく、細胞壁や細胞間隙を通って移動する。ただし、アポプラスト経路は、内皮を通過する際、カスパリ―線によって遮られるので、結局は水の移動経路は②のシンプラスト経路のみになる(シンプラストにすでに入っているか、内皮の細胞膜を通過した水と溶質だけが、維管束まで到達できる)。
②シンプラスト経路:原形質連絡でつながり合った細胞質のネットワーク全体からなる。この経路では、水は、原形質連絡を介して、根の皮層を横切って移動する。
雑談:皮層とは、主に、根では表皮と内皮の間、茎では表皮と維管束の間にある部分を指す。植物の基本組織系の主要素である。樹木においては、皮層はコルク形成層が分化してしばらくは生きつづけることもあるが、やがて樹皮として剥離する。 内皮を皮層に含めることもある。
まだわかっていないこと
● 栄養成長から生殖成長への転換は、どのような遺伝子、分子が関わるのか。
● フィトクロム、フォトトロピン、クリプトクロムが光を受容してから、植物体が反応するまでに、どのような分子が、どのように関わるか(光に対する応答には、素早い反応や、ゆっくりとした反応がある。また、必要光量も様々である)。
● フィトクロムはどのように植物の発生に関わっているのか(同じ被子植物の間でも異なる)。
● 光受容体の進化的な起源は何か。
● フロリゲンの進化的な起源は何か。