
【高校生物】進化⑤「遺伝子頻度はどのように変化するのか?」

~プロローグ~
進化遺伝学では、「集団内の遺伝子頻度の変化」を進化の定義の一つとしている。そのような進化が起きているかを調べる際に、ハーディー・ワインベルグの法則が利用できると考えられている。この法則は、遺伝的変異が起こらない集団をモデル化しているため、実際に起こっている変異を算出する場合には、逆に、理論上の標準として用いることができる。
「宇宙のなかに存在するものはすべて、偶然と必然との果実である。」デモクリトス
「進化は何かしら集団に起るものなのであるから、集団内の現象と個体での減少とを関連づける数学上の学説なしには、この問題に関しはっきりした考察はあり得ない。」ライト(遺伝学者。遺伝的浮動に関する研究が有名で、古い文献では、遺伝的浮動をライト効果と呼ぶこともある。)
「遺伝学は進化説と同義語ではない。一方進化説も決して遺伝学の一分科ではない。けれども遺伝学は進化機構の問題と密接な関係を持っているから、しっかりした遺伝の学説を無視した進化説はその根底において誤ったものである。」ドブジャンスキー(遺伝学者)『遺伝学と種の起原』より
★テストに出やすいワード
①遺伝的浮動
②遺伝子頻度
③ハーディー・ワインベルグの法則
④分子時計
⑤分子進化
要点:偶然により遺伝子頻度が変化することを、遺伝的浮動(いでんてきふどう)という。
(1)遺伝子頻度
● 集団がもつ遺伝子の集合全体を遺伝子プールといい、遺伝子プールにおけるそれぞれの対立遺伝子の割合を遺伝子頻度という。
● 遺伝子頻度とは、遺伝子Aとaが多数あった場合に、遺伝子全体のうち、Aやaがどの程度の割合で存在しているかを示す値である。
例)遺伝子型がAaの個体が100匹いるとすると(遺伝子数は200個)、Aとaの遺伝子頻度はどちらも0.5 (=100/200)である(遺伝子型Aaの個体1匹はAとaの2個の遺伝子を持っていることに注意)。
*この集団では、ハーディー・ワインベルグの法則が「成り立っていない」らしいことが推察できる(Aの頻度をp、aの頻度をqとした時、上の例では、遺伝子型AAの個体の頻度がp2に、遺伝子型Aaの個体の頻度が2pqに、遺伝子型aaの個体の頻度がq2になっていない。したがって、ハーディー・ワインベルグの法則が成り立っている集団ではないと推測できる)。
例)遺伝子型AAの個体が2個体、遺伝子型Aaの個体が1個体、遺伝子型aaの個体が1個体いるとすると(下図)、Aの遺伝子頻度は5/8=0.625である。
(なお、aの遺伝子頻度は0.375。3/8を計算してもよいし、Aの遺伝子頻度0.625を1から引いてもよい。全体を1としていて、遺伝子はAとaのどちらかしかないのだから、Aの遺伝子頻度を1から引けばaの遺伝子頻度が求まる。クジにおいて、当たりクジの出る確率が0.3なら、ハズレクジの出る確率が1-0.3=0.7になるのと考え方は同じ)。

(2)遺伝的浮動
● 『偶然』により遺伝子頻度が変化することを遺伝的浮動(いでんてきふどう)という。
雑談:次の世代をつくりだす配偶子は、現在の集団に存在する対立遺伝子の内、選ばれたもののみを含んでいる(遺伝子型Aaの男性がいたとする。しかし、その人が子供をつくった時、子供に伝わる遺伝子はAだけということはあり得る。たまたまAが入った精子によって子供が生じ、男性がその子以外に子を残さなければ、はじめ男性のもっていたa遺伝子は次世代に伝わらない。子をつくる男女全てでこういったランダムな遺伝子の選択がなされている)。したがって、ある対立遺伝子の集団における頻度は、その世代の親世代の集団と比べてわずかに増加しているかもしれないし、減少しているかもしれない。もともと頻度が低かった対立遺伝子は、偶然の結果、次世代に伝わらない可能性もある。このように、突然変異や自然選択が起こらなくても、遺伝子頻度は偶然により変動するのである(遺伝的浮動)。
雑談:生物集団の個体数は常に有限である。遺伝的浮動は、親世代から子世代に遺伝子が伝わる時に有限数の標本抽出が起こるために生ずる偶然の効果の現れである。
● 一般に、集団が小さいと、遺伝的浮動の影響が大きくなる。
雑談:遺伝的浮動とは、集団の遺伝子組成の確率論的に起こる変動のことである。集団が小さいほど、または形成される配偶子数(生産される子孫数)が少ないほど、(環境への適応などとは無関係に)次世代の遺伝子構成が抽出誤差によって決定される機会が増す(こう書くと難しく感じるが、イメージは全然難しくない。1/2で当たるクジを1億回[多数]引けば、当たりクジとハズレクジはおよそ1:1で出る。しかし、3回[少数]しか引かないと、誤差の影響が非常に大きくなり、すべてハズレ[もしくはすべて当たり]といったようなことが十分あり得る。そんなイメージ)。遺伝的浮動は進化の原動力の一つになっていると考えられている。
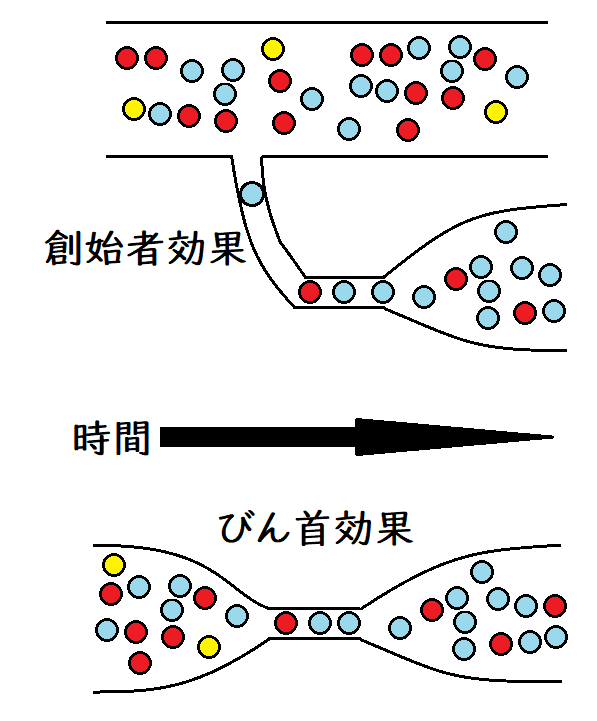
雑談:大きな集団から小さな集団(創始者集団)が隔離され、その小さな集団が新たな集団を確立することがある(移民などがその例である)。その時、新たな集団の遺伝子プールは、もとの大きな集団の遺伝子プールと異なる場合がある(創始者集団[小さな集団]では遺伝的浮動が起こりやすい)。これを創始者効果(そうししゃこうか)という。
「ペンシルベニア州東部のアーミッシュ共同体に住む1万4,000人は、18世紀初頭にドイツから移民した200人の子孫である。アーミッシュ住民の躁うつ病の発症率が欧州人集団に比べてはるかに高いのは、この病気の原因対立遺伝子を数人の創始者が保因していたためである可能性が高い。」ハートウェルら著『ハートウェル遺伝学』(メディカル・サイエンス・インターナショナル)より引用
雑談:はじめにアメリカ大陸に移住した祖先集団(創始者集団、少数の集団)では、偶然にO型の人数が多かった。アメリカ先住民の集団はその子孫から形成されたため、アメリカ先住民のABO式血液型では、O型の頻度が極端に高くなったと考えられている。
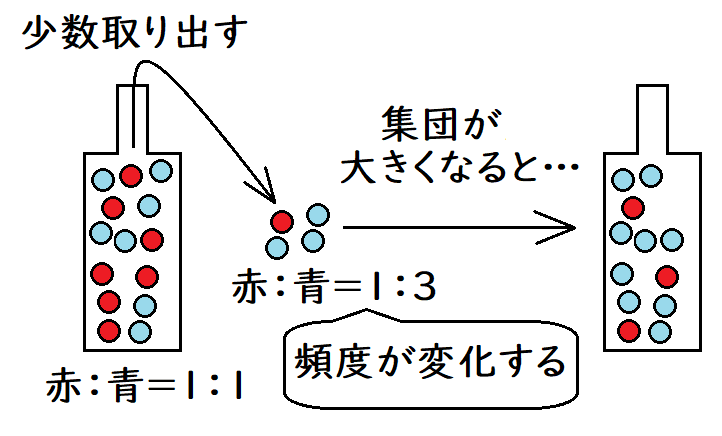
● びん首効果(びんくびこうか):集団の個体数がある短い期間にわたって極端に減少することをびん首効果という。この個体数が減少した期間では、遺伝的浮動の作用が強くなる。びん首効果が現れる状態はさまざまであるが、環境の激変によって個体数が激減する場合などが考えられる。
*たとえば、赤い玉100個と青い玉100個が入ったビンがあるとする(赤い球、青い球は異なる対立遺伝子のモデルである)。ここから100個の球を取り出せば、およそ赤:青=1:1で出てくるだろう。しかし、少数を取り出す場合は、アンバランスが生じやすい。たとえば、4個しか球を取り出さない場合は、3個が青で1個だけ赤などというアンバランスが生じることは十分にあり得る(狭いびん首を通して少数の球を取り出すことは、集団サイズの減少の比喩である)。びん首効果以降に集団の数が回復しても、元の集団と同じ遺伝子頻度(赤:青=1:1)にはならない可能性が高い。
雑談:以下は創始者効果とびん首効果のイメージ。高校生はまったく気にしなくてよい(創始者効果は高校範囲外。なお、創始者効果を、びん首効果の例の1つと見なすことも多い)。集団の数が回復しても、元の集団と同じ遺伝子頻度にはならない場合がある。黄色の球のように、新しい集団では消失する対立遺伝子もある。
要点:ハーディー・ワインベルグの法則が成り立っている集団では、遺伝子頻度が変化しない。
● 次の条件を満たす集団では、集団内の遺伝子頻度は、世代を経ても変化しない。これをハーディー・ワインベルグの法則という。
<条件>
① 自由な交配が行われる。
② 突然変異が起こらない。
③ 注目する形質に対する自然選択がはたらいていない。
④ 集団の個体数が大きく、遺伝的浮動の影響を無視できる。
⑤ 移入や移出がない。
*逆に、これらの条件が成り立っていない場合、遺伝子頻度が変化する場合がある。それは、つまり進化である(進化の定義の一つは、遺伝子頻度の変化である)。
<証明>
遺伝子Aの遺伝子頻度をp、遺伝子aの遺伝子頻度をqとする(ただしp+q=1とする)
*p+q=1は当たり前。全体が100%(=1)で、Aが70%(=0.7)なら、残りのaは30%(=0.3)なのは当然。
この集団では、配偶子(精子や卵)の遺伝子型の存在比率は、A:a=p:qである。任意交配すると(精子と卵がランダムで受精すると)、次世代の集団の遺伝子型の割合は、下表より、AA:Aa:aa=p2:2pq:q2

p+q=1に注意すると(p+qをつくるように式変形していくと)、次世代の遺伝子Aの遺伝子頻度は
(Aの数) /( 遺伝子の総数)
=(2p2+2pq) / 2(p2+2pq+q2)・・・式①
=(2p2+2pq) / 2(p+q)2 ←p+p=1を使う
=p2+pq
=p(p+q) ←p+p=1を使う
=p
よって、次世代のAの遺伝子頻度はpになる。同様に、遺伝子aの遺伝子頻度はqとなり、最初の世代と同じである。
*上の式①について:AAの個体はAを2個、Aaの個体はAを1個もつので、Aの数は(2p2+2pq)個。また、すべての個体は2個ずつ遺伝子を持つので、遺伝子の総数は2(p2+2pq+q2)個。
発展:ハーディー・ワインベルグの法則について
実際は、ハーディー・ワインベルグの法則は、以下の事実を主張する。
対立遺伝子A、aそれぞれの遺伝子頻度をp、qとすると
(i) 自然淘汰、突然変異、移住、遺伝的浮動などの要因が働かずに任意交配の行われる集団ではp、qは世代とともに変化しない。
(ii) 任意交配の下では、遺伝子型AA、Aa、aaの個体の存在頻度はp2:2pq:p2となり、以後任意交配の下でこの割合は不変に保たれる。
高校教科書や参考書では、(i)のみが強調され、(i)が大発見であるかのように書かれることが多い。
しかし、(i)はそもそも当たり前(先ほどのような仰々しい計算・式変形をしなくても、わかり切っていること)である。
遺伝子は自己増殖していく因子だからである(かつて、メンデル遺伝を理解していない人が「メンデルの遺伝の法則が正しいならば、優性の表現型の頻度は必ず75%生じるようになるはずだ。」「メンデルの遺伝の法則が正しいならば、集団内で優性形質が増加し、劣性形質が減少していくはずだ。」「だからメンデルの法則は間違っている。」という見当違いの批判をしていた。(i)は、そのような誤解を解くのに役に立った)。
対して、(ii)は有用である。ハーディー・ワインベルグの法則が成り立つような集団では、遺伝子型の頻度が簡単な掛け算で推定できるからである(AA:Aa:aa=p2:2pq:q2)。
*下表のうち、遺伝子型aaの頻度q2は、(aを劣性遺伝子として、)劣性形質をもつ個体数を直接数えることで得ることができ(遺伝子型aaの個体数/全体の数)、そこからqが求まる。
*劣性形質をもつ個体の遺伝子型はaaで確定なので、遺伝子型aaの個体数は直接数えることができる。
qを1から引けばpも求まる。
p、qが求まれば、遺伝子型AAの頻度(=p2)、遺伝子型Aaの頻度(=2pq)がそれぞれ推定できる。

雑談:「ハーディー‐ワインベルグの法則は有用ではあるが理論的には簡単なことで、これを多くの教科書にあるように、もったいぶってむずかしそうに説明する必要はない。…ハーディー‐ワインベルグの法則が集団遺伝学のまるで根本であるかのように述べているのは、筆者には実にばかげたことのように思われる。…遺伝子頻度を変化させる特別の要因(突然変異、…自然淘汰など)が存在しないかぎり、メンデル性分離だけでは遺伝子頻度には変化は起きない…遺伝子が自己増殖分子であることからも当然であろう。…もしヒトが半数生物だとしたらハーディー‐ワインベルグの法則など誰も問題にしないと思われる。」木村資生『生物進化を考える』より
雑談:なお、ランダムに交配が起これば、どのような遺伝子型頻度の集団から始めても、1代で上の表のような遺伝子型頻度の状態に達する(ランダムな交配は、まるで、ランダムに遺伝子を混ぜ合わせ、各個体が持つ遺伝子の組み合わせを新たに決めるようなものである。はじめ、だれがどのように遺伝子を持っていようが、ランダムな混ぜ合わせが1回でも起これば上表の遺伝子型頻度になるのは当然である。たとえば、はじめ、AAとaaが同数ずついるとしよう。この集団のAの頻度は0.5、aの頻度は0.5である。ランダムな交配が起きると、次世代の遺伝子型頻度はどうなるか?0.5の頻度で精子と卵がそれぞれ存在し、ランダムに受精するのだから、遺伝子型AAが生じる頻度は0.5×0.5、遺伝子型Aaが生じる頻度は2×0.5×0.5、遺伝子型aaが生じる頻度は0.5×0.5となる[下の表において、Aの遺伝子頻度p=0.5、aの遺伝子頻度q=0.5とすればよい]。たった1回のランダムな交配で、遺伝子型AAの頻度がp2に、Aaの頻度が2pqに、aaの頻度がq2になった。これは、はじめの集団にどのような遺伝子型頻度を想定しても変わらない。結局、pの頻度でAが、qの頻度でaが存在しており、ランダムな混ぜ合わせが起こるからである)。

問題:遺伝子Aとaについて考える。遺伝子Aはaに対して顕性(優性)である。遺伝子型AA、Aaの人は正常だが、遺伝子型aaの人は疾患を持つとする。あるハーディー・ワインベルグの法則が成り立つ集団において、疾患を持たない人が75%、疾患を持つ人が25%だった。
(1)aの遺伝子頻度qを求めよ。
(2)Aの遺伝子頻度pを求めよ。
答え:
(1)ハーディー・ワインベルグの法則が成り立つ集団の話をしているので、以下の表のようになっている。
「疾患を持たない人が75%、疾患を持つ人が25%」と言われているが、疾患を持たない人75%のデータはあまり役に立たない。なぜなら、この75%のうち、何%が遺伝子型AAで、何%が遺伝子型Aaなのか、すぐにわからないからである。

だから、こういう問題では潜性(劣性)形質をもつ人に注目する。

疾患の人は遺伝子型aaで確定(aが劣性だから)。
しかも、疾患の人の頻度(遺伝子型aaの人の頻度)はq2である(ハーディー・ワインベルグの法則が成り立つ集団だから)。
よって、25%は0.25だから、
0.25=(0.5)2=q2
q=0.5
(ちなみに、基本的に、ハーディー・ワインベルグの法則が成り立つ集団についてのすべての問題で、ちゃんと劣性の人の頻度は、何かの数字の綺麗な2乗になっている。25%[0.5の2乗]とか、16%[0.4の2乗]とか、36%[0.6の2乗]とか。)
したがってaの遺伝子頻度qは0.5
(2)
くじ全体の内、はずれくじの頻度がわかっているのなら、1からそれを引けば当たりくじの頻度である。
たとえば、30%(=0.3)ではずれるくじがあったなら、当たる確率は70%(=1-0.3=0.7)である。
aの遺伝子頻度が(1)で0.5とわかっているので、A(当たりくじ)の遺伝子頻度pは、1からa(はずれくじ)の遺伝子頻度qを引けば求められる。
p=1-q=1-0.5=0.5
したがってAの遺伝子頻度pは0.5
講義動画【自由交配】
講義動画【遺伝子頻度】
講義動画【ハーディー・ワインベルグの法則】
問題:ハーディー・ワインベルグの法則が成り立たつ条件を5つ答えよ。
答え:①集団が十分に大きい②個体の移出・移入がない③突然変異が起こらない④自然選択が働かない⑤交配が任意(ランダム)に行われる
雑談:数学者ハーディーは映画『奇蹟がくれた数式』の主人公の一人(もう一人の主人公はラマヌジャン。ハーディーはラマヌジャンを支援した)。
雑談:「数学の証明は、単純で明確な星座のようであるべきで、銀河に散らばる星くずの集まりのようであってはならない。」ハーディー『ある数学者の生涯と弁明』より
雑談:遺伝子型の頻度がわかれば、遺伝子頻度は必ず求まる。たとえば、遺伝子型AAが30%、遺伝子型Aaが40%、遺伝子型aaが30%の頻度で存在しているなら、
遺伝子Aの頻度pは(遺伝子Aの数)÷(遺伝子の総数)
={(30×2)+40}÷200
=0.5
よってaの頻度qも0.5(=1-0.5)
しかし、逆に、『遺伝子頻度がわかっても、遺伝子型の頻度はわからない』。
Aの頻度が0.5で、aの頻度が0.5の集団には、以下のような集団もある→①AA:aa=1:1に存在する集団。②すべてがAaの集団 など
したがって、遺伝子頻度から遺伝子型頻度を求めるには、なんらかの条件(仮定)が必要となる。それが『ハーディー・ワインベルグの法則が成り立つ集団』という条件である。この狭い条件の集団では、遺伝子型AAの頻度はp2であり、遺伝子型Aaの頻度は2pq、遺伝子型aaの頻度はq2である。
『ハーディー・ワインベルグの法則が成り立つ集団』において、Aの頻度が0.5(p=0.5)、aの頻度が0.5(q=0.5)ならば、AAは0.25の頻度で、Aaは0.5の頻度で、aaは0.25の頻度で存在することになる。
AAの遺伝子型頻度 = p2 = 0.5×0.5 = 0.25
Aaの遺伝子型頻度 = 2pq = 2×0.5×0.5 = 0.5
aaの遺伝子型頻度 = q2 = 0.5×0.5 = 0.25
<Q.遺伝子頻度と遺伝子型頻度ってなにが違うの?…文字通りである。遺伝子頻度は、(ある遺伝子の数÷遺伝子の総数)で求められ、遺伝子「型」頻度は(ある遺伝子「型」÷個体数)で求められる。
AA AA Aa aa
の集団がいるとすると、Aの遺伝子頻度は5/8(全8個の遺伝子のうちAは5個)。AAの遺伝子型頻度は2/4(全4個体のうちAAは2個体)。>
<Q.ハーディー・ワインベルグの法則って何の役に立つの?…遺伝子型の頻度は、見た目ではわからないことが多い(AAとAaがどちらも優性形質で区別がつかない)。そこで、(ハーディー・ワインベルグの法則が成り立っている集団ならば、)aaの頻度(=q2)から、遺伝子頻度qやpを求めることで、Aaの存在頻度(=2pq)などを推察できる。
劣性形質をもつ個体の遺伝子型はaaとわかる。遺伝子型aaの個体を数え、その頻度を求める。それがq2と等しいことから、qが求まる。すると、1ーq=pより、pも求まる。ハーディー・ワインベルグの法則が成り立っている集団ならば、遺伝子型AAがp2、遺伝子型Aaが2pqの頻度で存在するので、理論上の遺伝子型AAの頻度、遺伝子型Aaの頻度が求まる。また、ハーディー・ワインベルグの法則が成り立っていると仮定して出した理論値との"ずれ"から、新しい知見が得られる(自由な交配が起きていなかったり、自然選択が起きている場合などは、理論値と異なってくる)。>
*何度も注意しておくが、「遺伝子型aaの個体の頻度がq2」などと判断できるのは、ハーディー・ワインベルグの法則が成り立っている集団の場合だけである。
講義動画【ハーディー・ワインベルグの法則(難問対策:ABO式血液型)】
要点:重要な機能を持つ分子ほど、変異の蓄積速度(進化の速度)は遅くなる。
● DNAの塩基配列やタンパク質のアミノ酸配列に関する進化を分子進化という。
● タンパク質やDNA分子の定速的な経時変化(もしくは、そのような分子の変異が特定遺伝子に蓄積する速度の一定性)を分子時計という。
<Q.分子時計ってどこが時計なの?…DNAの塩基配列や、タンパク質のアミノ酸配列が『一定の速度で』変化していく。そのため、これらの分子の情報は、種の分岐年代など、生物の進化の過程を分析するための「時計」となり得、分子時計と呼ばれる。『一定の速度をもつ』ということが時計の本質である。確かに、君の持っている時計の針も、一定の速度をもつ。この速度の一定性が、時計の条件である。「針が3つ進んだから3秒経ったことになる」、などと推定できるように、「種Aと種Bのヘモグロビンを比べると、アミノ酸が**個異なっているから、種Aと種Bが分かれてから**億年たったことになる」、などと推定できる。>
● 特定の遺伝子の塩基配列や、特定のタンパク質のアミノ酸配列についての種間での違いは、それらの種が共通の祖先から分かれてからの時間を反映している。また、この違いは、同じ種間でも重要な機能に関係する遺伝子やタンパク質ほど少ない(変異が遅い)傾向にある。
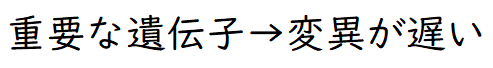
● DNAの塩基配列やタンパク質のアミノ酸配列など、生体を構成する物質の分子データを用いてつくられた系統樹を分子系統樹(ぶんしけいとうじゅ)という。
講義動画【分子時計】
講義動画【系統樹の作成】
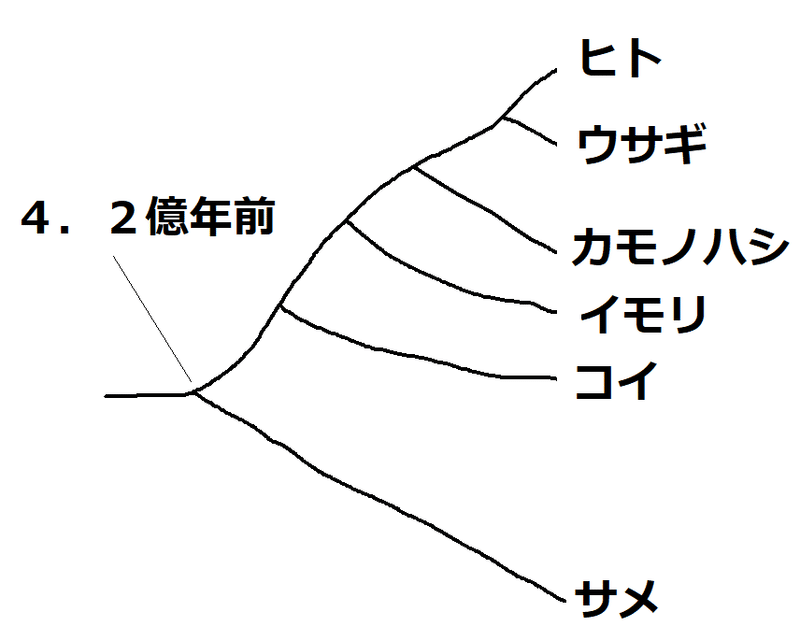
問題:いくつかの脊椎動物の間で、ヘモグロビンα鎖(141個のアミノ酸からなる)のアミノ酸配列を比較し、アミノ酸の違いの数を表にまとめた。サメの祖先が他の種の動物の共通祖先から分かれた時期は4.2億年前と考えられている。
カモノハシの祖先が、ヒトやウサギの共通祖先から分かれたのは何億年前か。最も近いものを選べ。
①4.2億年前 ②2.2億年前 ③1億年前

下図は系統樹のイメージ
答え:②
このような問題では、各生物の間に、『別々の位置に、均等に変異(置換)が起こった』と考える(問題文で言われなくても)。
サメは、他の動物と均等に離れているはずであるから、変異の数を平均する。
(たとえば、サメとヒトの遠さと、サメとウサギの遠さは、理論上同じであるはずである。サメと他の動物の差がばらついているのは、誤差[のようなもの]と考え、平均してしまう。問題文で説明がなくても自分で平均を取る。)
サメと、他の動物種の違いの数を平均する。サメと他の動物の違いは
(75+84++84+85+79)÷5=81.4個
さらに、その半分の数の変異が、それぞれの動物種に起こったと考える。
よって、サメと、他の動物に、それぞれ
81.4÷2=40.7個の置換が起こったと考えられる。
(それぞれ、別々の場所に40.7個の置換が起こったから、違いは81.4 個になった。それぞれ異なる位置に置換が起きると仮定してよい[問題文で言われなくても])
*僕とあなたが100ページの同じ本を持っているとする。僕がどれか1ページを書き換える。あなたもどれか1ページを書き換える。僕の本とあなたの本を比べたとき、違いは2ページになる。
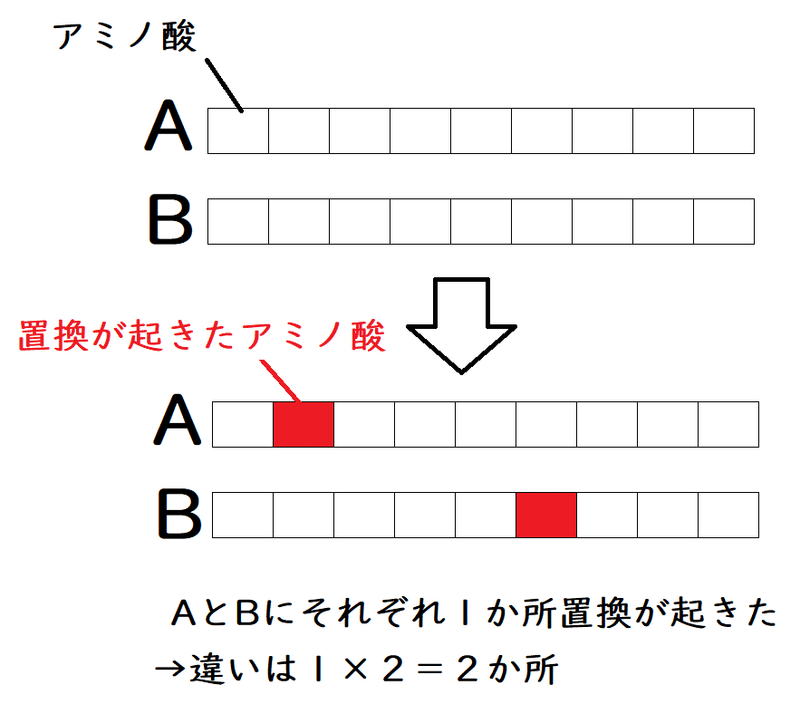
40.7個の置換が起きるのに4.2億年かかった。
一方、カモノハシと(ウサギ、ヒト)の違いは、同じように平均を取って
(49+37)÷2=43個
(カモノハシとウサギ、カモノハシとヒトは、理論上同じくらい離れてるはずである。表のデータは49と37でばらつきがあるから、誤差[みたいなもの]と考えて、平均を取る。)
よって、その半分の、43÷2=21.5個の置換が、カモノハシと(ウサギ、ヒト)にそれぞれ起こったことがわかる。
*カモノハシはカモノハシで21.5か所置換が起きた。(ウサギ、ヒト)は(ウサギ、ヒト)で21.5か所置換が起きた。なので、違いが43か所になった。
問題文より、4.2億年前にサメの祖先が他の種の動物の共通祖先から分かれたのだから、変異の速度は一定であるとして
4.2億年:40.7個=?億年:21.5個
?=約2.2億年前・・・答え
(「4.2億年で40.7か所に置換が起きた。ならば、?億年で21.5か所に置換が起きる」と考えている。)
(それぞれの種に起こった置換の数を求めなくても、
4.2:81.4=?:43で解けるのだが、今回は他の問題にも対応できるように、置換の数まで求めた。)
たとえば、以下のような問題が続くこともある。
(難しい人はとばしてもよい)
追加問題(発展):上の問題の条件で、アミノ酸座位1個に置換が起きる確率は、1年あたり、どれくらいになるか。サメのデータから求めよ。
解説:
アミノ酸座位1個あたり、1年あたりの、置換が起きる確率をnとする。
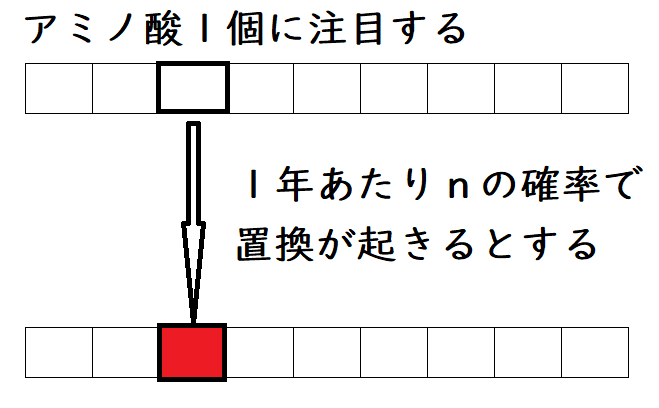
つまり、「1年あたり、アミノ酸1個に置換が起きる確率」を求めようとしている。
今回、ヘモグロビンα鎖は141個のアミノ酸からなる(問題文より)。
宝クジが141個あるようなものである。1年あたり、宝クジ1枚が当たる(変異が起きる)確率がnと考えてよい。
当たる確率がnの宝クジが141枚ある(1年に1回抽選会がある)。4.2億年たったら、141枚のうち、40.7枚が当たっていた(40.7か所に置換が起きた)と考える。
*同じ宝くじは2回当たらないと考えてよい(同じ場所に2度置換が起きる場合は考慮しなくてよい[問題文で言われなくても])。
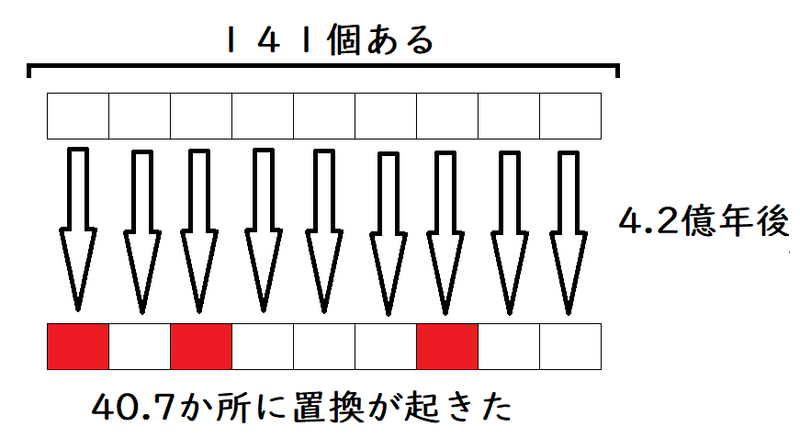
n × (141個) × (4.2億年) = 40.7個
*nはアミノ酸座位1個あたり、1年あたりの置換が起きる確率。
(「当たる確率がnの宝くじが141枚ある。抽選会は1年に1回。4.2億年たったら、40.7枚が当たっていた。」正確に言えば、「アミノ酸座位1個あたり、1年あたりの置換が起きる確率をnとする。αヘモグロビンは141個のアミノ酸からなり、4.2億年で40.7か所に変異が起きた。」)
n=約6.8×10⁻¹⁰ ・・・答え
発展:最節約法(さいせつやくほう)
可能な系統樹のうち、変化の回数(塩基配列であれば塩基置換の回数)が最も少なくなる系統樹を選択する方法を最節約法という。基本的に、一度変化が起きた場所は元に戻ることはないと仮定してよい(「一度塩基が置換した場所に、もう一度置換が起こって、元に戻った可能性」は考えない。無駄に多くの可能性を考えることになるからである)。
高校生はそれほど深く考えなくてもいい。系統樹を推定する時に、余計に複雑にしようとしなければいいだけである。ここでは大雑把な最節約法のイメージだけ解説する。
たとえば、種①~③とその祖先種の、ある位置の塩基配列が、以下の表のようになっていたとする(祖先種の配列は様々な別のデータから明らかにしたものとする)。
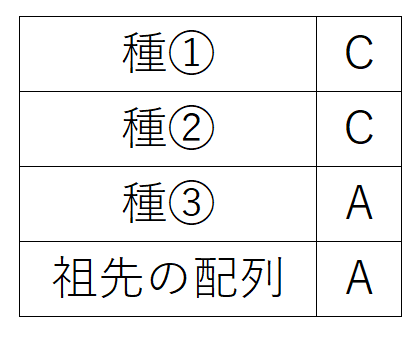
以下の3つの系統樹の可能性を考える。最節約法を採用した場合、選択すべき系統樹は、一番上の系統樹である(仮定している変化の回数が最も少ないから)。赤矢印は塩基の置換(A→C)が起きたことを表す。
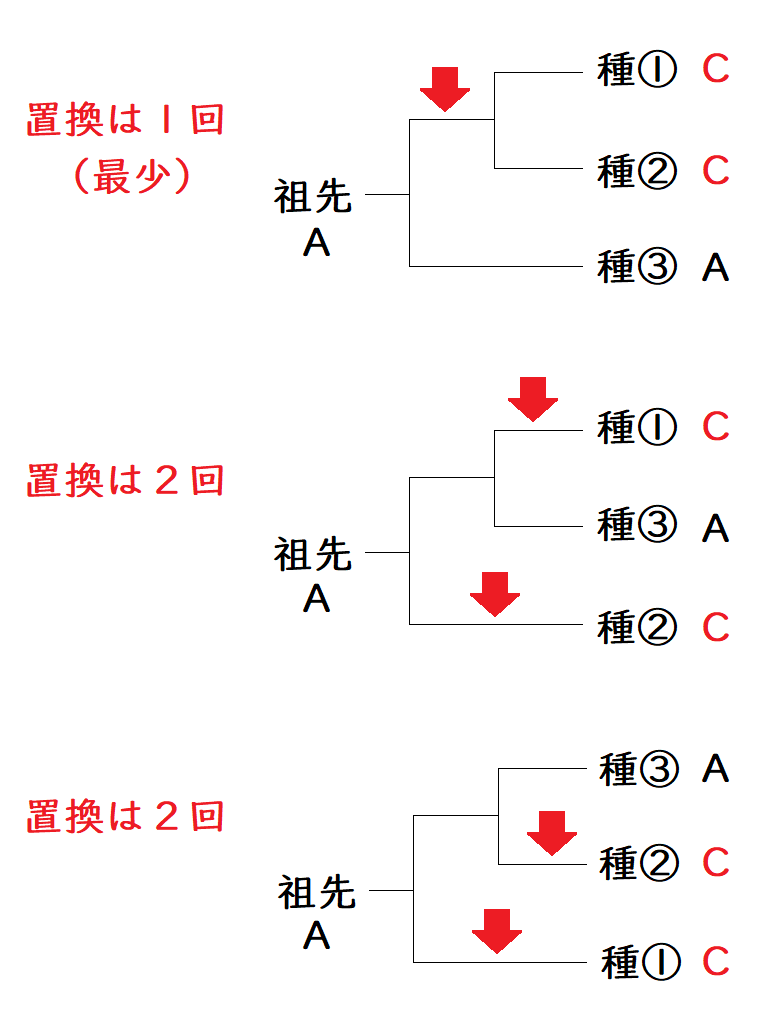
*上のイメージでは、種の数が少ないので、「ちょっと系統樹を描くだけで何を大げさな!たった3つの可能性しかないのに、あれこれ考えるなんて!」と思ったかもしれない。しかし、種の数が10種になるだけで、可能な系統樹は数千万種類になる。普通、複数の位置の塩基のデータを比較し、コンピューターを用いて、最もあり得そうな系統樹を選択する。
雑談:最節約法は、科学全般で重視される最節約原理(オッカムのかみそり[不必要な複雑性をそぎ落とした、最も単純な問題解決手段を提唱した哲学者オッカムにちなむ])に基づいた方法である。
雑談:最節約法は長い間分類学において神聖視されてきたが、現在では、最節約法がどのような場合でも最善というわけではないことが明らかになり、よく用いられる系統樹作成法の一つとして位置付けられている。
雑談:共通祖先からの由来に基づかない類似性が存在する場合がある(系統の異なる複数の生物が類似する形質を個別に進化させた場合。すなわち収束進化[収れん]が起きた場合)。そのような場合、最節約的でない系統樹を最節約的だと見誤ってしまうこともある。
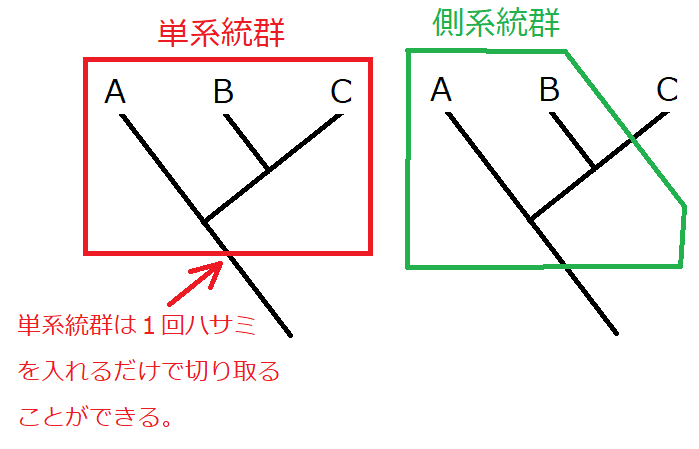
発展:単系統群と側系統群
ある1つの共通祖先とそれから派生した種のすべてを含むようなグループを単系統群(たんけいとうぐん)という(単系統群は、系統樹に1回ハサミを入れるだけで切り取ることができる)。単系統群から1つ、または複数の単系統群を除いて構成されるようなグループを側系統群(そくけいとうぐん)という。
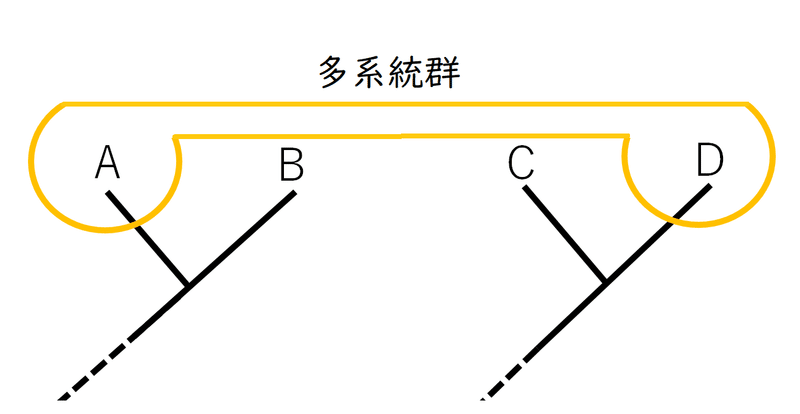
雑談:系統樹上で連続していない種をまとめたグループを多系統群と呼ぶ場合がある。たとえば、同じような形質を収束進化によって(独立に)得たいくつかの種を1つにまとめてグループを作ってしまっていた場合、その群は多系統群と見なされる(ただし、高校では多系統群は分類群には含めないことが多い。実際の研究でも、ある分類群が多系統群であることが判明した場合、その群の分類を再検討するのが普通である。また、現代では、側系統群と多系統群は系統樹上では区別できないとされることもある[どちらも単系統ではないという点で同じであり、概念的な区別の有効性はないという考えがある])。
講義動画【最節約法・単系統群・側系統群】
雑談
まだわかっていないこと
● 分子進化(タンパク質や塩基配列の進化)と、表現型の進化はどう関係しているのか。それぞれの進化の相違点は何か(それぞれの傾向に違いはあるか)。そして、それらの進化の証拠は、何を観察・解析することで得られるのか。
*中立説は、中立(か、ほぼ中立)な変異が起き、それらのものの中で幸運なものが、自然選択ではなくて、遺伝的浮動により集団に固定するということを主張する(負の自然選択も起きるが、正の自然選択は無視できるほど少ない)。一方で、生物の表現型の進化には、自然選択が働いていることは、歴然とした事実である。どのようなデータがマクロな進化とミクロな進化を繋ぐのか。
● 集団遺伝学についてのパラメータは、正確な測定が非常に困難である(突然変異率や、淘汰係数、集団の大きさ、移住率などを正確に測定するのは困難である)。ならば、正確な研究はどのように行えばよいか。木村資生は『分子進化の中立説』で、以下のように述べている。
「これらの集団遺伝学的パラメータは決して正確にはわからず、だからそんなパラメータを含む理論は役に立たないと主張する強い傾向があった。私は反対の意見を持っていて、もしわれわれが本当に分子レベルでの進化機構を理解したいならば、これらのパラメータは研究され測定されなければならないと信じている。天文学者や宇宙論学者は、各種の天文学的パラメータを正確に推定するのが困難であるという理由でそんなパラメータを含む理論は避けるべきだと主張するのであろうか?」