
【高校生物】動物生理⑤「筋肉はどのように収縮するのか?」

~プロローグ~
筋肉は収縮の機能をもつ組織であり、主にミオシンとアクチンというタンパク質から構成されている。動物は、筋肉を用いて、体のさまざまな部位を動かす。
ヒトの場合、筋肉は全体重の40~50%を占める。筋肉は力を生み出す(化学エネルギーが力学的エネルギーに変換される)だけでなく、熱を生産したり、各器官の体積を保持したりする働きも持っている。
「人間にできるのは動かすということであり、言葉をつぶやくのであれ、森の木を切るのであれ、それを実行する唯一の主体は筋肉である。」シェリントン(イギリスの神経生理学者)
★テストに出やすいワード
①筋繊維
②筋原繊維
③滑り説
④暗帯・明帯
⑤サルコメア
要点:筋肉は筋繊維(筋細胞)からなり、筋繊維には筋原繊維という微細構造が見られる。
(1)筋繊維(きんせんい)と筋原繊維(きんげんせんい)
● 骨格筋などの筋肉は、筋繊維(筋細胞ともいう)からなる。
● 筋繊維の細胞質には細長い筋原繊維(筋細胞内に存在する微細構造)が束になって存在する。
*筋原繊維は細胞の名前ではないので注意。筋原繊維は、筋細胞の収縮を担う細胞小器官であり、主にアクチンフィラメント、ミオシンフィラメントの束からなる(筋原繊維は、筋繊維内に多数ある直径1μmくらいの円筒状の微細構造である)。両フィラメントの滑りから力を発生する(筋原繊維を「細胞小器官」とは呼ばないことも多いが、今回は東京化学同人『生物学辞典』の解釈に従った)。筋原繊維は筋小胞体の網目によって包まれている。
*めっちゃ大雑把に言えば、筋肉は筋繊維(筋細胞)の集まり、筋繊維(筋細胞)は筋原繊維の集まりみたいなイメージ(あまり正確な言い方ではないが)。

(2)明帯と暗帯
● 横紋筋の筋原繊維には明るい部分(明帯[めいたい])と暗い部分(暗帯[あんたい]:収縮しても長さは変わらない)が交互に並んでいる。
(3)アクチンフィラメントとミオシンフィラメント
● 筋原繊維にはアクチンフィラメントやミオシンフィラメントが含まれている。
雑談:アクチンフィラメントはアクチンというタンパク質から、ミオシンフィラメントはミオシンというタンパク質から成る。「フィラメント」は繊維状の構造を意味する。ミオシンは、以下のような形(2本のゴルフクラブを合体させたような形)をしている。ATPアーゼ活性とアクチンへの結合能は頭部に存在する。

雑談:アクチン(actin)の語源は「act(行動する)」に由来する。また、ミオシンのミオ(myo-)は「筋肉の~」に由来する。
● 明帯の中央には、Z膜とよばれる仕切りがあり、Z膜からZ膜までをサルコメア(筋節[きんせつ])という。
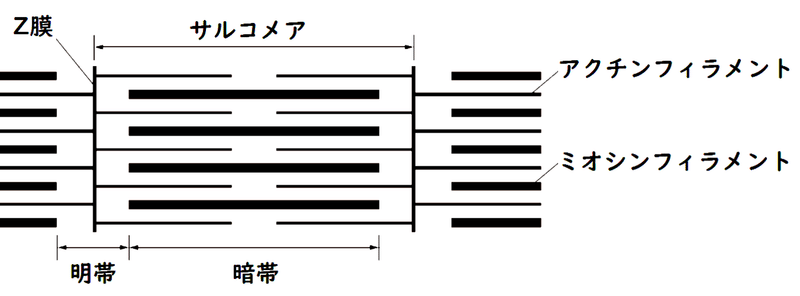
雑談:Z膜のZの由来はドイツ語のZwischenscheibeで、between discの意。Z膜は、明帯の中間に、黒い線として観察される。
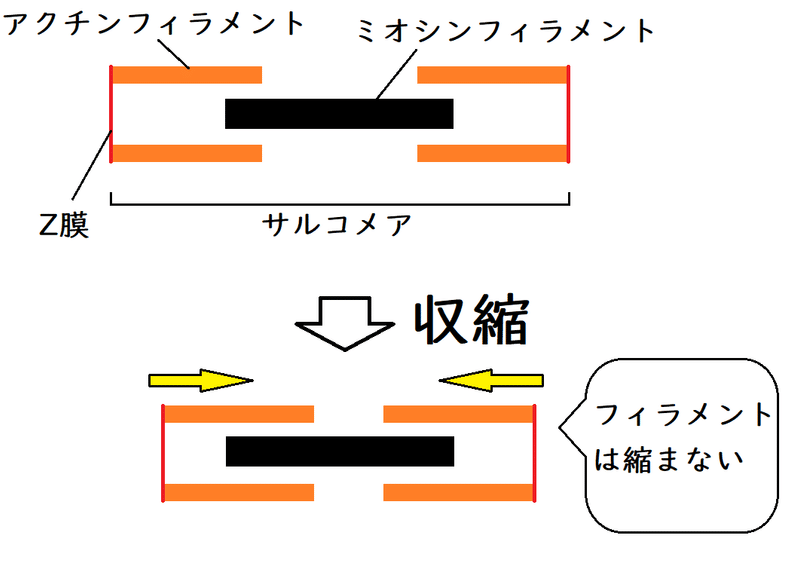
以下はサルコメアが収縮する時のイメージ。フィラメント(オレンジ色や黒色の棒)の長さは変化していないことに注意(フィラメントの滑りが起きている[フィラメントがスライドしている])。
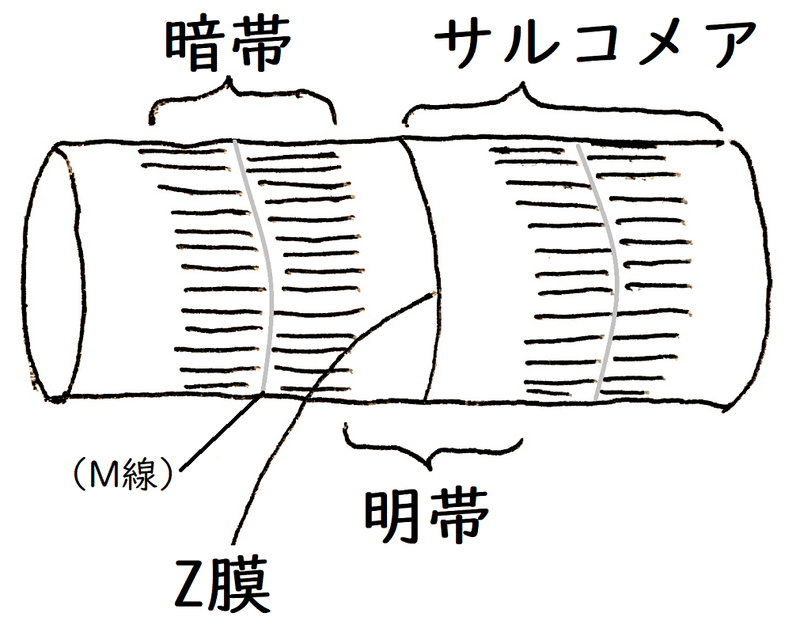
雑談:高校生は知らなくてよいが、暗帯の中央にはM線という仕切りがあり、濃い線に見える。M線には色々なタンパク質があり、ここでミオシンフィラメントが連結している。下図は筋原繊維のイメージ。
発展:筋原繊維の断面
①アクチンフィラメントしかない所の断面、
②ミオシンフィラメントしかない所の断面、
③両方のフィラメントがある所(ミオシンフィラメントの間にアクチンフィラメントが入り込んでいる所)の断面
は、以下の図のようになる。
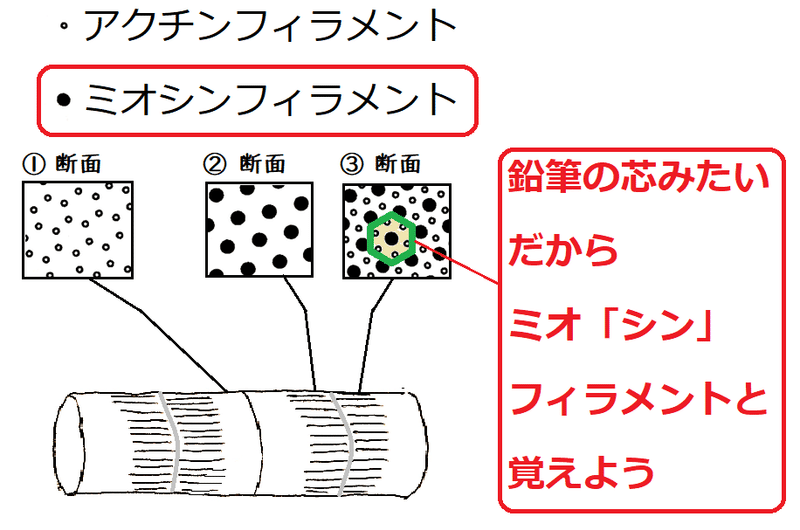
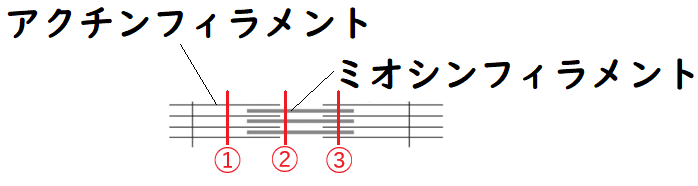
②の断面:ミオシンフィラメントのみ見える。
③の断面:両方のフィラメントが見える。
(4)筋肉の種類
● 筋肉には以下のような種類がある。骨格筋の細胞が多核であること、骨格筋と心筋は横紋筋である(横紋と呼ばれる縞模様がある)ことはよく出題される。
*意志によって働く筋肉を随意筋、意志とは無関係に働く筋肉を不随意筋という。
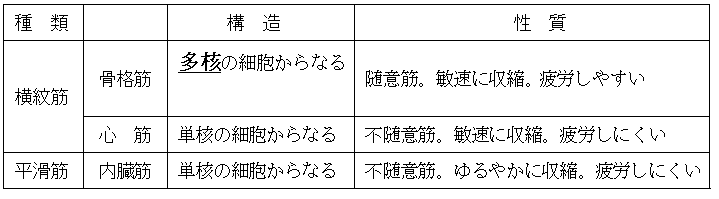
*心筋を内臓筋に含む(内臓筋を、内臓にある平滑筋および心筋の総称と定義する)ことも多い。高校生は今学校で使っている教材に解釈を合わせること。
雑談:骨格筋の筋繊維は、数百個の筋芽細胞が融合して生じる(したがって骨格筋の筋細胞は多核にある)。
雑談:平滑筋では、筋節が横紋筋のようには整然と配列されていない。
講義動画【筋収縮】
(5)暗帯の長さは変化しない
● ミオシンフィラメントの存在する部分は、暗帯(あんたい)として観察される。収縮の前後で暗帯の長さが変化しない。
語呂「変化しなければ安泰だ(暗帯の長さは収縮の前後で変化しない)」
語呂「あんた、変わらないね(暗帯の長さは収縮の前後で変化しない)」


収縮後・・・
暗帯の長さ:変化しない(非常によく問われる)
明帯の長さ:短くなる
サルコメアの長さ:短くなる
問題:筋収縮の前後で、長さが変化しないものを1つ選べ。
①明帯 ②暗帯 ③サルコメア
答え:②
要点:筋肉が収縮するしくみは「滑り説(すべりせつ)」で説明される。
● 筋収縮はミオシンフィラメントの間にアクチンフィラメントが滑りこむことによって起こる(滑り説[すべりせつ])。
雑談:正確には「横紋筋の収縮を筋原繊維のアクチンフィラメントとミオシンフィラメントの相対的な滑りで説明する学説」を滑り説という。
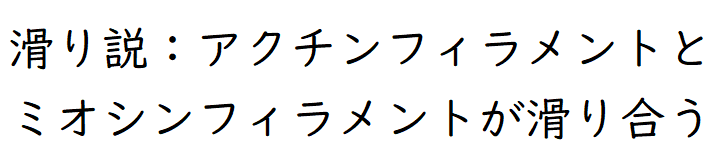
*筋収縮の流れ
① 神経筋接合部(しんけいきんせつごうぶ)において、運動神経の軸索末端からアセチルコリンが分泌される。
語呂「運動して汗が散る(運動ニューロンがアセチルコリンを分泌)」
*下図には描いていないが、アセチルコリンは筋細胞の細胞膜上の受容体に結合する。これにより筋細胞内にNa+が流入し、活動電位が発生する(②が起こる)。
*運動ニューロンと筋細胞との接合部を神経筋接合部(しんけいきんせつごうぶ)という。
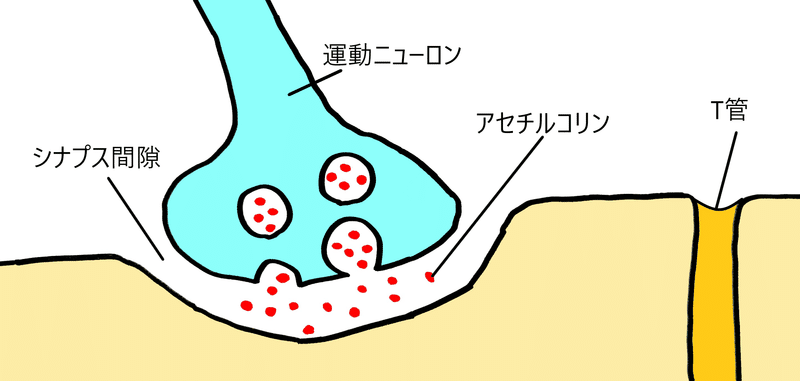
② 筋細胞内にNa+が流入し、活動電位が発生する。膜電位の変化はT管を介して筋細胞の奥まで伝わる。
*筋細胞の細胞膜は深く陥入し、筋小胞体(きんしょうほうたい。特殊化した滑面小胞体)に達している。この筋細胞の細胞膜が陥入しているところをT管(てぃーかん)という。
③ T管の活動電位に伴って、筋小胞体からCa2+が細胞質に放出される。
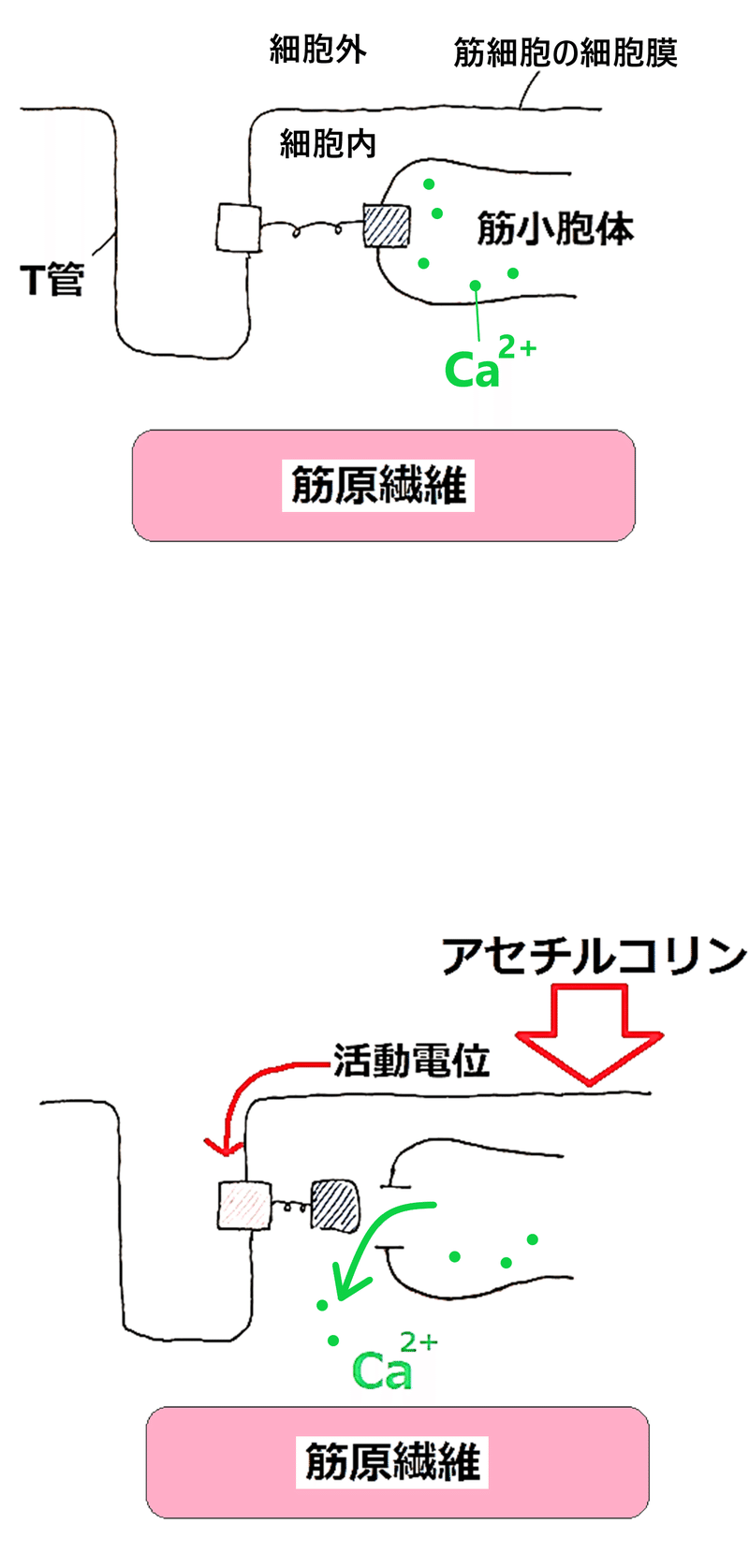
④ Ca2+がトロポニンと結合する。すると、トロポミオシンの構造が変化する。
⑤ トロポミオシンによって塞がれていたアクチンフィラメントのミオシン結合部位が露出し、アクチンフィラメントとミオシン頭部が結合できるようになる(アクチンのミオシン結合部位が露出する)。図はイメージ。
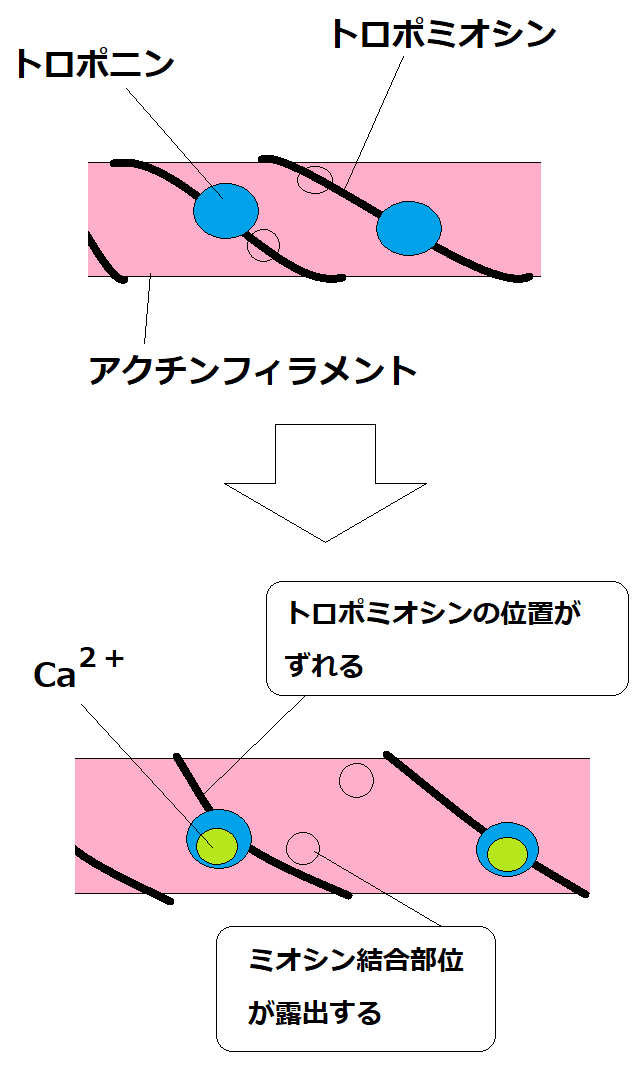
*下図はイメージ(下図はあまり正確ではない。トロポミオシンはミオシン頭部とアクチンフィラメントの結合を阻止している。トロポミオシンにくっついているトロポニンにCa2+が結合すると、トロポミオシンの構造が変化し、ミオシン頭部とアクチンが結合できるようになると考えられている)。
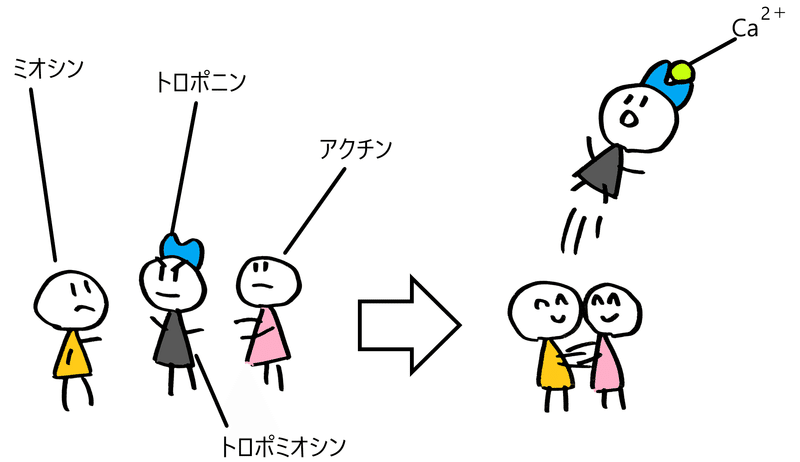
⑥ ミオシン頭部がアクチンフィラメントに結合する。
⑦ ミオシン頭部がリン酸とADPを放出する。ミオシン頭部の立体構造が変化してアクチンフィラメントをたぐり寄せる。これにより筋肉が収縮する。
⑧ ATPがミオシン頭部に結合すると(アクチンとミオシン頭部の親和性が低下し)、ミオシン頭部がアクチンフィラメントから離れ、筋肉がし緩する。
⑨ ATPを分解したミオシン頭部は、立体構造を回復する。
⑩ 筋小胞体は『能動輸送』でCa2+を回収する。(めちゃめちゃ良く問われる!!!)

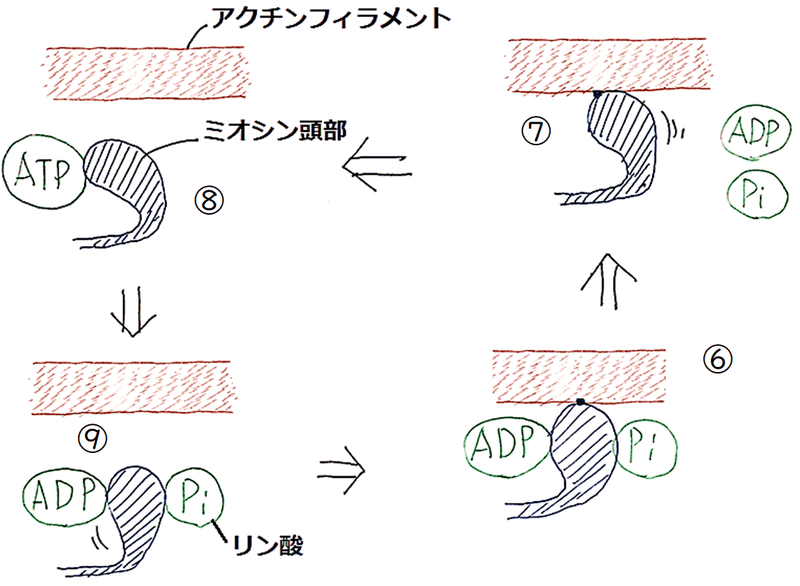
⑦ ミオシン頭部がリン酸とADPを放出し、アクチンフィラメントをたぐり寄せる。
⑧ ATPがミオシン頭部に結合するとミオシン頭部がアクチンフィラメントから離れる。
⑨ ATPを分解したミオシン頭部は、立体構造を回復する。
講義動画【滑り説】
講義動画【筋収縮】
雑談:コナン・ドイルによる小説『シャーロック・ホームズ』には、クラーレという毒が登場する。クラーレは、南米の原住民によって用いられた矢毒で、有効成分の一つであるd-ツボクラリンは、運動神経終末から分泌されるアセチルコリンと拮抗して、終板(運動終板ともいう。神経筋接合部に形成される筋表面の板状の構造のこと)に存在するアセチルコリン受容体を占拠し、アセチルコリンの作用をブロックする。結果、神経筋遮断を引き起こす。
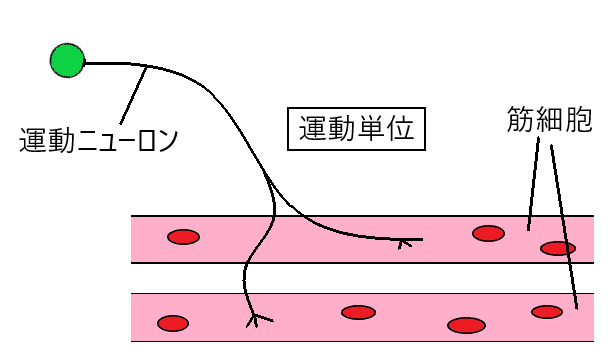
雑談:1本の運動ニューロンは、複数の筋細胞(筋繊維)を支配している。1本の運動ニューロンと、その運動ニューロンが支配するすべての筋細胞をまとめて「運動単位(うんどうたんい)」という。運動ニューロンに活動電位が発生すると、その運動単位のすべての筋細胞が1つのまとまりとして収縮する(興奮した運動ニューロンの支配する筋細胞が同期的に興奮・収縮する)。
雑談:江橋節郎(えばし せつろう)は、筋収縮のメカニズムについて、筋小胞体から放出されるカルシウムイオンによって収縮が引き起こされるという説を提唱した(当時、筋収縮がカルシウムイオンによって制御されているという説は大反対を受けたが、現在では正しいことがわかっている)。さらに江橋はカルシウムイオンと結合するタンパク質「トロポニン」を発見し、カルシウムイオンによる筋収縮・弛緩制御の分子機構を明らかにした。
雑談:モーターやエンジンでは、スイッチのオンオフは、エネルギーの供給により行われることが多い(電流を流すことでモーターは回転を始める)。筋肉の場合、燃料であるATPは常に存在する。したがって、別のスイッチが必要となる。トロポニンなどを用いた筋収縮の調節は、筋収縮を制御するスイッチのようなシステムとも見なすことができる(トロポニンは、細胞内カルシウム濃度によって収縮のON/OFFするスイッチのように働いている)。
雑談:筋肉において、力を発揮する過程の終わりになっても、アクチンとミオシン頭部の架橋は解消されない。新たなATP分子との結合が架橋の解消には必要である。細胞の膜構造は、死後、その密閉性を失い、カルシウムイオンが壊れた筋小胞体から漏出する。これがきっかけとなってミオシン頭部がアクチンに結合する。しかし、ATP合成が停止しているので、ミオシン頭部とアクチンの連結が解消されない。その結果、筋は硬くなり、収縮も伸展もしなくなる。これが「死後硬直(死体硬直)」である。死後硬直は死後3~4時間に始まり、約24時間続くが、リソソームからのタンパク質分解酵素が架橋を分解し消失する。なお、死後硬直の評価は主観的であり、死亡時刻の推定に利用するには、あくまでも参考程度にとどめておく必要がある。
雑談:かつて、筋繊維は、ゼンマイのようにぐるぐる巻かれたもの(引き延ばすことができるが、放すとすぐにぐるぐる巻きにもどるようなもの)であると考えられていた。
雑談:イギリスの生物物理学であるハンソンは、ミオシンとアクチンのフィラメント構造を明らかにし、同じイギリスの生物物理学者ハクスリとともに筋収縮の滑り説を発表した。また、ハンガリーの生理化学者セント・ジェルジは、アクチンとミオシンの複合体(アクトミオシン)が、ATPによって、ガラス管内で収縮することを示した。
雑談:サリン (メチルホスホノフルオリド酸イソプロピル)は化学兵器用剤の神経ガスである。1930年代からドイツにおいて殺虫剤の研究の中で開発された。
サリンは、1994年の松本サリン事件、1995年の東京地下鉄サリン事件でテロに使用された。
サリンはアセチルコリンエステラーゼ活性阻害作用(アセチルコリンの分解を阻害する作用)を有し、極めて毒性が高い。
神経終末はアセチルコリンをシナプスに放出し筋肉を収縮させる。筋肉はアセチルコリンが存在するかぎり、収縮を持続する。そのため、通常は、アセチルコリンエステラーゼが、直ちにアセチルコリンを分解している。
サリンが組織のアセチルコリンエステラーゼの作用を阻害すると、アセチルコリンエステラーゼはアセチルコリンを分解することができなくなる。すると、アセチルコリンがシナプスに過剰に蓄積し、筋収縮をコントロールできなくなる。その後、短時間で様々な症状が現れ、呼吸筋が麻痺状態になり死亡する。
発展:クレアチンリン酸
筋収縮によってATPが減少すると、筋細胞に含まれるクレアチンリン酸(クレアチンリン酸はエネルギーを貯蔵している)が分解され、素早くATPが合成される。
クレアチンリン酸+ADP⇄クレアチン+ATP
*筋収縮のエネルギーはすべてATPから供給される。
*ATPは呼吸や解糖の反応によって生産される。
*筋細胞には、エネルギーがクレアチンリン酸として蓄えられている。
*長時間収縮が続くと、呼吸の反応に必要な酸素の供給が間に合わなくなる。その場合でも、解糖によってATPが供給される(解糖は酸素を必要としない)が、乳酸が蓄積する(乳酸は解糖の副産物である)とともに、エネルギーの供給が不足し、十分な収縮ができなくなる。この状態を疲労という。
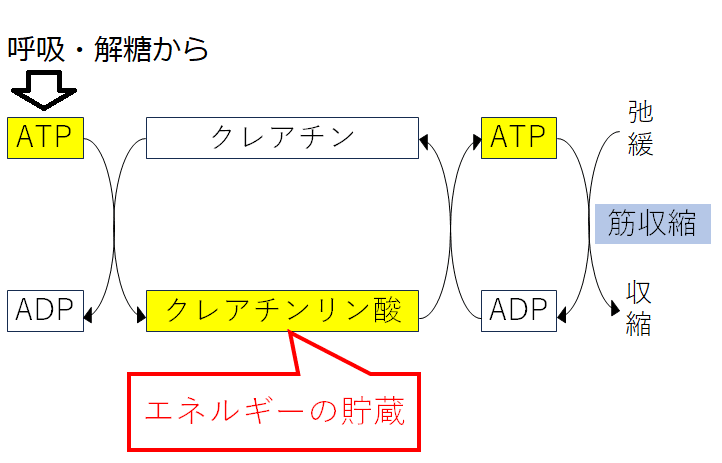
雑談:ATPとは異なり、クレアチンリン酸は細胞機能システムとの間の直接の共役を担えない。しかし、クレアチンリン酸は、補助的なエネルギー貯蔵庫として機能している。筋肉において、クレアチンリン酸とATPは一緒になって、最大約15秒間の最大収縮力を発揮するためのエネルギーを供給する。筋細胞内のクレアチンリン酸が使い果たされた時、グルコースが分解され、ATPが産生される。
雑談:筋収縮の研究が始まったころ、筋収縮の前後で、ATP濃度が変化しないという報告がなされ、研究者達は困惑した。どうしてATPが減っていないように見えたのか?実際は、ATPは分解されているが、すばやくクレアチンリン酸によって再合成されていたのであった。
要点:筋肉の収縮は「単収縮」「不完全強縮」「完全強縮」に分けられる。
(1)単収縮と潜伏期・収縮期・弛緩期
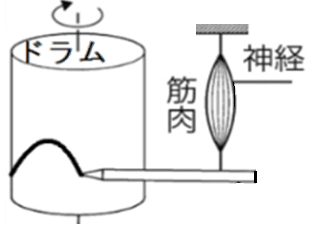
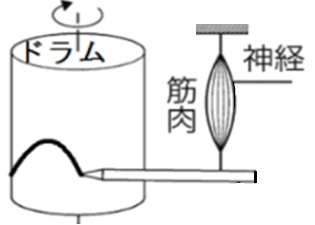
● 筋肉の収縮のようすはキモグラフという装置で観測できる。
筋肉が収縮すると、筋肉の下についたペンが、その様子を筒状の紙に描き出す(筋肉が縮むとペンが上に上がる。筋肉が伸びると、ペンが下に下がる)。
雑談:運動を曲線として描記する装置の総称はキモグラフと呼ばれる。筋肉の単収縮のように特に速やかな運動の記録装置はミオグラフ(筋運動記録器)と呼ばれる(ミオグラフではドラムの回転が速い)。
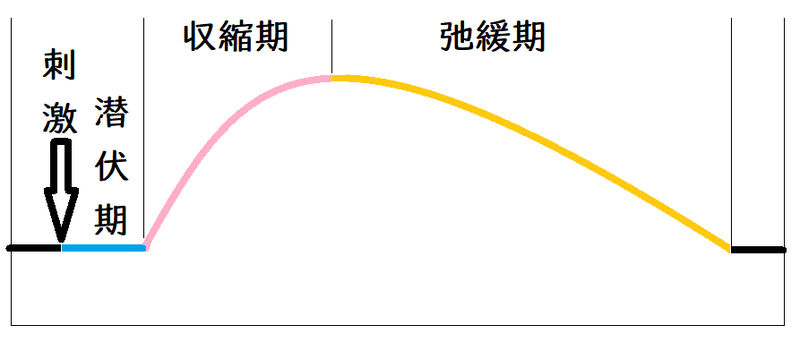
● 単収縮(たんしゅうしゅく):潜伏期(せんぷくき)・収縮期(しゅうしゅくき)・弛緩期(しかんき)に分けられる。
*単収縮:単一の刺激で生じる収縮。
下図は単収縮のイメージ(ミオグラフによる記録)。
(2)全か無かの法則
● 骨格筋においては、刺激強度を増すにつれ、単収縮の大きさが増大するが、これは、興奮する筋細胞(筋繊維)の数の増加によるものである。個々の筋細胞の単収縮は全か無かの法則に従う。

(3)強縮(不完全強縮・完全強縮)
● 強縮(きょうしゅく):連続的な刺激で起こる単収縮の重なった大きな収縮で、刺激の頻度によって以下の2つに分けられる。
①不完全強縮(ふかんぜんきょうしゅく。毎秒約15回程度刺激したときにおこる強縮)
②完全強縮 (かんぜんきょうしゅく。毎秒約30回以上刺激したときにおこる強縮)
下図はいろいろな収縮を記録した時のイメージ(キモグラフによる記録)。
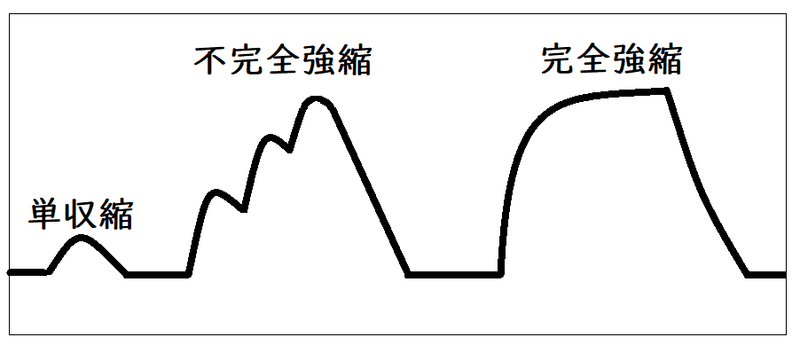
雑談:筋肉に適当な頻度の反復刺激を与えた時、相次ぐ単収縮に加重が起こって大きな持続性収縮を生じた状態を「強縮」という。上で説明したのは骨格筋の例だが、平滑筋では毎秒6回以下の刺激でも強縮が起こる。心筋は不応期が長いため、強縮を生じ得ない。また、セミの発音筋などは、単収縮の経過が極めて速やかで、毎秒100回の刺激でも加重を来さない。なお、生体の随意運動や反射運動の多くは、運動神経からの反復衝撃に基づいて生じた強縮である。
発展:いろいろな効果器(発電器官・発光器官・色素胞)
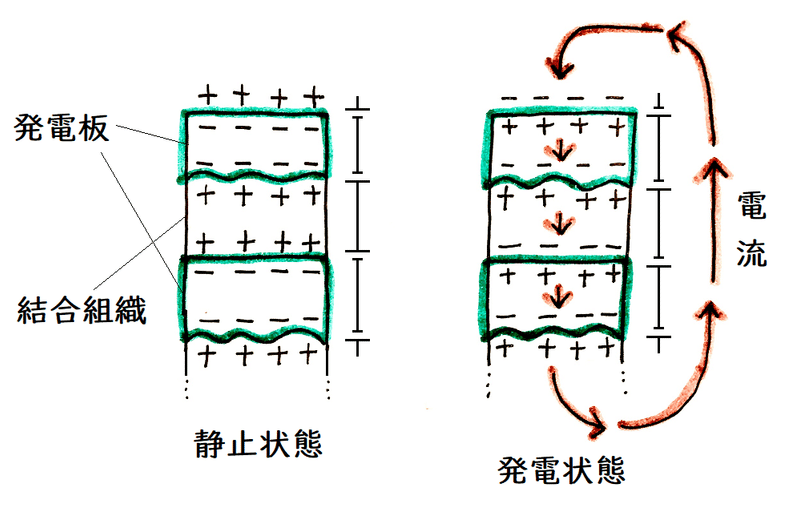
①発電器官:筋細胞が変化した細胞(発電板[はつでんばん])が多数積み重なっている。神経からの刺激で電流が発生する。
例 )シビレエイ
雑談:発電魚の発電器官は、発電板という扁平な細胞が集まったものである。下図は発電板の機能のイメージ。静止状態では電流は流れない。(図には示していないが、発電板の平らな面の細胞膜には運動神経が分布しており、)神経からの刺激により発電板に活動電位が発生すると、(まるで電池を直列につないだ時のようになり)、電流が生ずる(発電状態では、静止状態に比べ、発電板の上側の平らな膜の所で電位が逆転していることに注目)。
②発光器官:発光物質(ルシフェリン)が酸化されると発光する。ATPを用いる。ルシフェラーゼという酵素が関与する。
例)ホタル
雑談:ルシフェラーゼの語源は堕天使ルシファー。
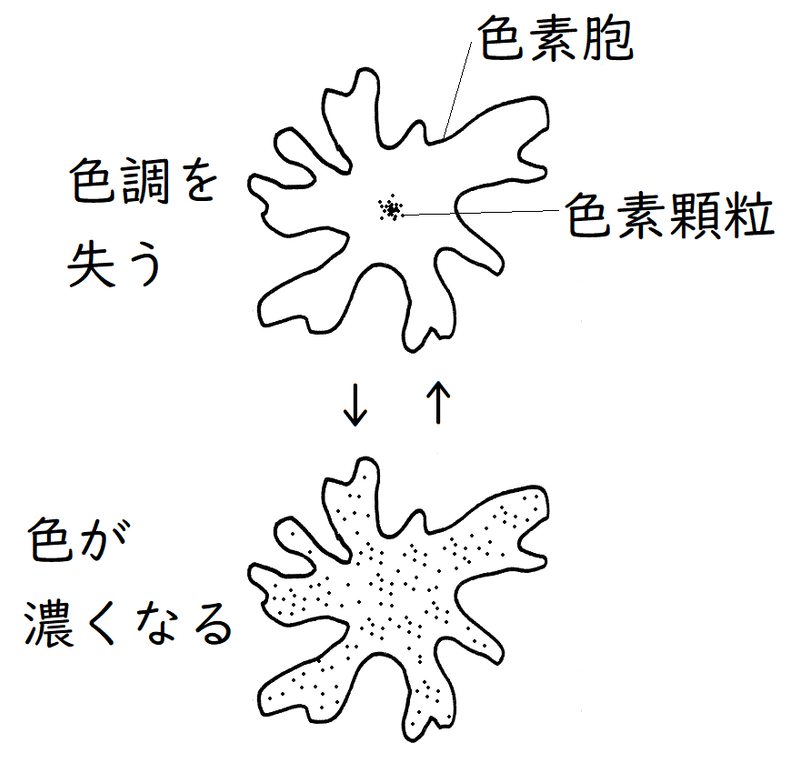
③色素胞(しきそほう):色素胞中の色素顆粒(しきそかりゅう)の拡散・凝集により、体色変化が起こる(色素顆粒の輸送には微小管がかかわることが知られている)。色素顆粒が拡散すると体色が暗くなる。
例)メダカ(周囲が明るいと、色素顆粒が凝集し、体色が明るくなる。周囲が暗いと、色素顆粒が拡散し、体色が暗くなる)
雑談:色素胞は、多くの魚類、両生類、爬虫類および無脊椎動物の外皮などに存在する細胞であり、内部に色素顆粒をもつ。通常、突起が放射状に発達している。色素胞は色素細胞と呼ばれることもある(色素胞は「細胞」であり、液胞などのような「細胞小器官」ではないので注意)。色素顆粒が中央部に凝集すると皮膚は色調を失う。逆に、色素顆粒が細胞内に広く拡散しているときには皮膚の色が濃くなる。色素顆粒の凝集・拡散を引き起こす刺激(神経やホルモンによる刺激など)の種類は、動物によって異なる(たとえばメダカでは、色素胞内の色素顆粒は、交感神経によって凝集し、脳下垂体から分泌される黒色素胞刺激ホルモンによって拡散する。メダカは、周囲の明るさに応じて体色を変え、外敵に発見されにくくしている)。
<Q.筋繊維と筋原繊維と筋細胞の違いは?…筋繊維=筋細胞としてよい(テストではどちらを書いても○。集まって筋肉になる)。筋原繊維は細胞内構造(細胞の中にある構造)で、細胞ではない。>
<Q.教科書に出てくる下図のような装置は一体なんなの?…筋肉が縮む様子を記録する装置。筋肉の端にペンが付いていて、筋肉が収縮・弛緩すると、回転しているドラム型の記録紙に山の形の線が描かれる。なお、よく問題で出てくる「座骨神経」は多数のニューロンの集まりであり、「筋肉」は多数の筋細胞の集まりである(したがって全か無の法則にしたがわない。全か無かの法則に従うのは1個のニューロン、1個の筋細胞である)ので注意。>
講義動画【伝導速度に関する計算問題】
雑談:上の動画のように、筋肉に繋がった神経上の2点を刺激し、神経を走るインパルスの平均速度(伝導速度)を求める方法を考案したのは、エネルギー保存則を打ち立てたことで有名なヘルムホルツ(ドイツの物理学者・生理学者)である(ヘルムホルツが上の動画で解説したような方法で伝導速度を測定するまでは、伝導速度は、光速に匹敵するほど速く、計測は不可能に近いと言われていた)。
発展:サルコメアの長さに関するグラフ
以下のグラフから、アクチンフィラメント1つと、ミオシンフィラメントの長さを求めてみる。
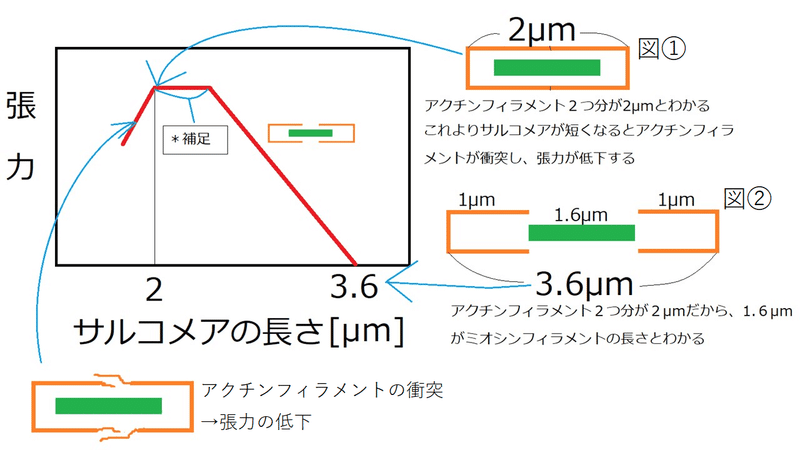
アクチンフィラメントが衝突する瞬間(サルコメアが短くなり過ぎて張力が低下する瞬間:図①)、サルコメアの長さが2μmになっている。したがってアクチンフィラメント1つ分の長さは1μm。
張力0になる瞬間(サルコメアの長さ3.6μmの時)は、アクチンフィラメントとミオシンフィラメントの重なりがなくなる瞬間(図②)である。したがって図②より、ミオシンフィラメントの長さは1.6μm。
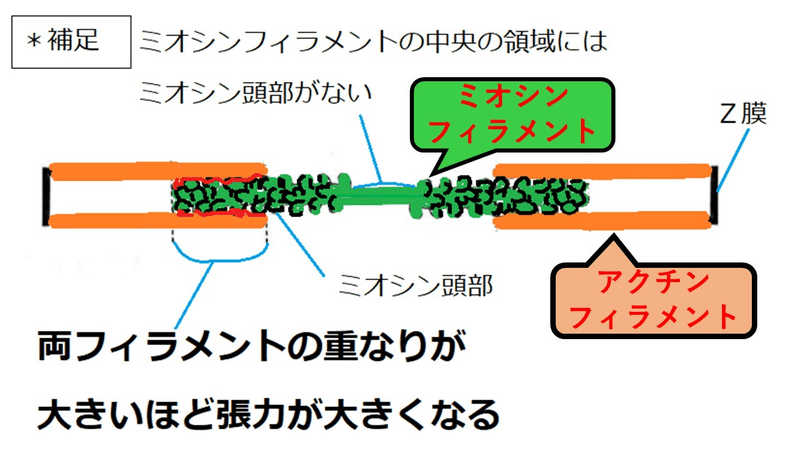
補足:サルコメアの長さが変化しても張力が変化していないところがある。これは、ミオシンフィラメントの中央部にミオシン頭部がない領域があることが原因である(張力はミオシン頭部とアクチンフィラメントが結合することで発生する[ミオシン頭部がある領域とアクチンフィラメントが重なり合えば合うほど張力が大きくなる]。ミオシン頭部がない領域で重なり合っても張力が増えない)。知らなくてよい。
講義動画【サルコメアの長さと張力のグラフ】
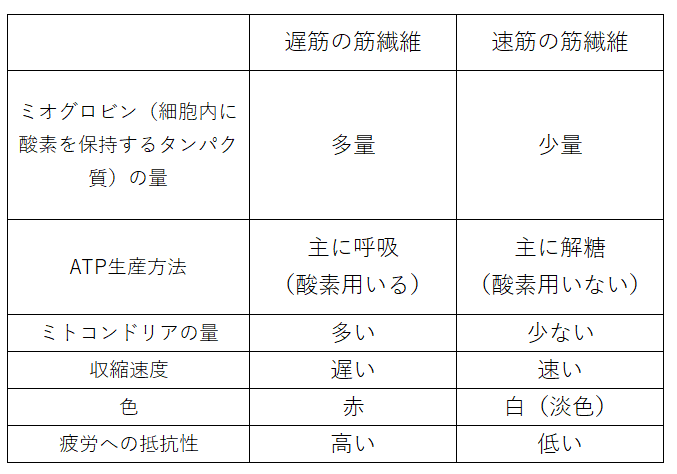
発展:遅筋と速筋
骨格筋には、遅筋(ちきん)と速筋(そくきん)がある。
覚え方「遅筋は時間をかけてエネルギーを生産=時間がかかるエネルギー生産方法は呼吸(よってミトコンドリア多量)=呼吸には酸素が必要=ミオグロビンがたくさんの酸素を保持=赤色になる」
発展:グリセリン筋
筋肉を棒などに結びつけ、50%のグリセリンに浸したものをグリセリン筋(ぐりせりんきん)という。グリセリン筋は、細胞膜の構造が壊れており、電気刺激には応じないが、ATPを加えると収縮する(アクチン・ミオシンなどによる収縮機能は保持している)。グリセリン筋は筋肉のモデルとして、滑り説の研究に大いに役に立った(―20℃で保存でき、劣化が非常に少ない)。
*本来、生体内での筋収縮には、筋小胞体から放出されるCa2+が必要である(ミオシンとアクチンが結合するには、トロポニンにCa2+が結合し、トロポミオシン[アクチンのミオシン結合部位を覆っている]の構造が変化する必要がある)。しかし、グリセリン筋では、(ミオシンとアクチンの結合を阻止している)トロポミオシンがなくなっているので、ATPを加えただけで収縮が起こると考えられている。
まだわかっていないこと
● 心筋の細胞、および、平滑筋の細胞では、どのように興奮が制御され、どのように収縮が起こっているのか。特に心筋は、どのように心臓の自動性に関係しているのか
● 滑り説について、両フィラメントの相対的な滑りは、ミオシン頭部が、ATPの加水分解とアクチンとの反応を繰り返すことにより生じると考えられているが、完全には解明されていない(滑り「説」は非常に有力な仮説であるが、まだ仮説である)。
● 筋肉には、動物種によって、どのような多様性があるのか。
● 筋肉の進化的な起源は何か。