
【高校生物】生物の生活①「生物はどのように増えていくのか?」

~プロローグ~
「朝がくると東風はいなごを運んできた。いなごはエジプト全土を襲い、エジプトの領土全体にい坐った。これ程沢山のいなごはこれより先にもなかったし、これから以後もないであろう。いなごは全地の表面をおおい、地はそれで黒くなった。」『旧約聖書 出エジプト記』より
「私のたてた前提が承認されたものと仮定して話を続けるなら、人口が増える力は、土地が人間の食糧を生産する力よりもはるかに大きい。人口は、何の抑制もなければ、等比級数的に増加する。・・・今この地点に含まれる生命の種に、それをどこまでも成長させるに足る食糧とスペースが与えられたならば、数千年のうちに地球数百万個分の生命があふれるだろう。必然性、すなわち厳然と全体を支配する自然の法則が、生命の数をあらかじめ定められた範囲内に制限するのである。」マルサス『人口論』より
★テストに出やすいワード
①成長曲線・環境収容力
②密度効果
③相変異
④標識再捕法
⑤生命表・生存曲線
要点:「環境収容力」以上に個体群が大きくなれないのは、種内競争の激化や、排せつ物の蓄積による。
● 一定地域に生活する同種の生物の集まりを個体群(こたいぐん)という。
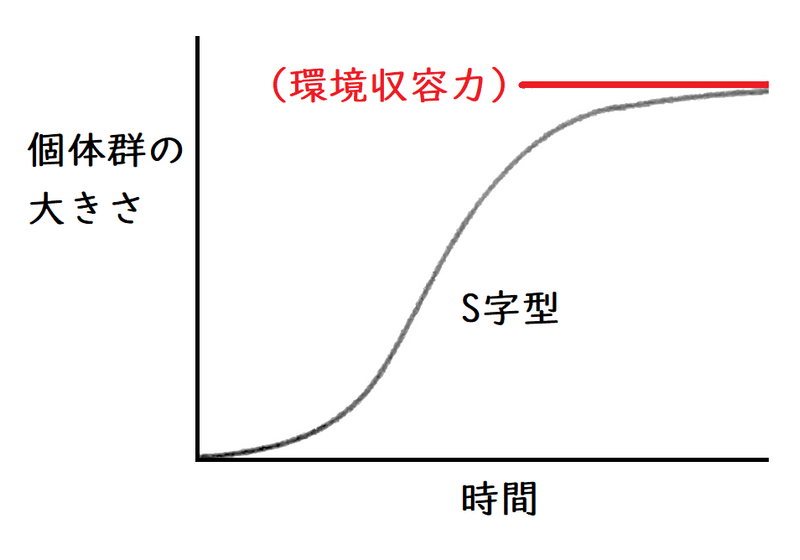
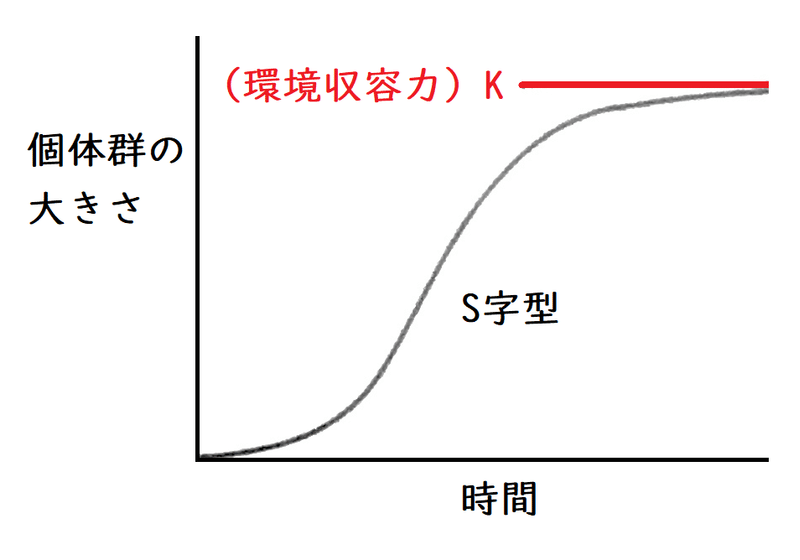
● 増殖を抑制する要因のない理想状態では、(理論上)個体数は指数関数的に増加するはずだが、実際には個体数の増加に伴って個体群の成長速度はにぶり、やがて個体群の大きさはほぼ一定の値に落ちつく。
したがって、個体群の大きさの時間変化をグラフにすると、下図のようになる。これを成長曲線(せいちょうきょくせん)という。成長曲線はS字型になる。
*個体群の成長を抑制する要因としては(1個体当たりの食物や生活空間の減少による)種内競争(しゅないきょうそう。同種個体間の競争のこと)の激化、排出物の増加による生活環境の悪化などがある(よく問われる)。
*ある環境で存在できる最大の個体数を環境収容力(かんきょうしゅうようりょく。物理で習う「力」とは全然違う意味なので注意)という。
雑談:手塚治虫の漫画『ブラック・ジャック』には、ある架空の病が登場する。漫画中の登場人物、戸隠によれば、その病は、「飢饉」と「食料不足」に対する神の警告であり、正体などないのだという。それを聞いたブラック・ジャックは天に向かって絶叫する。「医者は人間の病気をなおして命を助ける!その結果世界じゅうに人間がバクハツ的にふえ食料危機がきて何億人も飢えて死んでいく・・・そいつがあなたのおぼしめしなら・・・医者はなんのためにあるんだ」
要点:バッタの相変異は、密度効果の例である。
● 個体群密度が個体(の形態や生理・行動など)に影響を及ぼすことを密度効果(みつどこうか)という。
● 密度効果の例
①ショウジョウバエなどでは、個体群密度が高くなると、雌1匹あたりの産卵数が減少する。
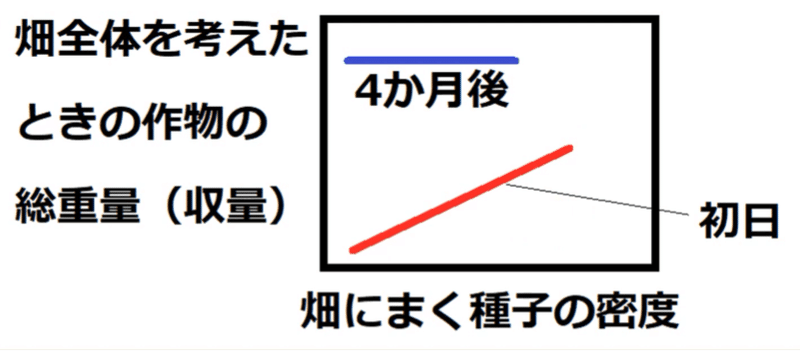
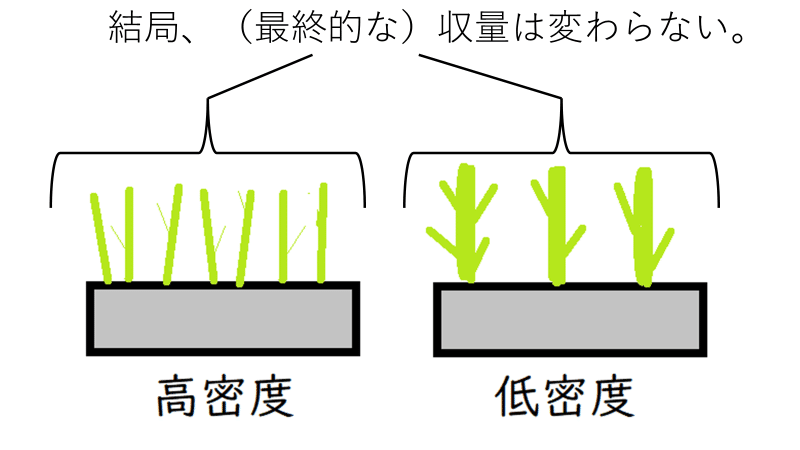
②ダイズなどをいろいろな密度で育てると、密度が高いほど光や栄養などをめぐる個体間の競い合い(種内競争)が激しくなる。結果、個体の成長は悪くなる。
→最初の密度(まばらに植えるか密に植えるか)にかかわらず個体群全体の重量は最終的にほぼ一定になる。
これを、最終収量一定の法則(さいしゅうしゅうりょういっていのほうそく)という。
*たとえば、「畑に種を高密度にまけばまくほど、収穫量は増える」などということにはならない。高密度に種子をまくと植物体1個1個は成長が悪くなり、最終的な収量(畑全体の植物体の重量)は、低密度で種をまいた時と同じ程度になってしまう。
下図はイメージ。
雑談:最終収量一定の法則は、「種を蒔く時の密度や、苗木を植える密度を変えても、十分生育が進んだ時の単位面積当たりの植物体の現存量が、初めの密度に関わりなく一定になるという密度効果の法則」である(文字通り「最終」の「収量」が「一定」になる法則)。なお、厳密には「収量」は植物体全体の現存量を表しており、穂や種子などの実用部分のみを指すわけではない。
雑談:最終収量一定の法則は、吉良竜夫らによって世界に先駆けて発見された。
③相変異(そうへんい):ある種の昆虫は、幼虫時などの個体群密度によって形態・生理・行動などが変化する。この現象を相変異という。
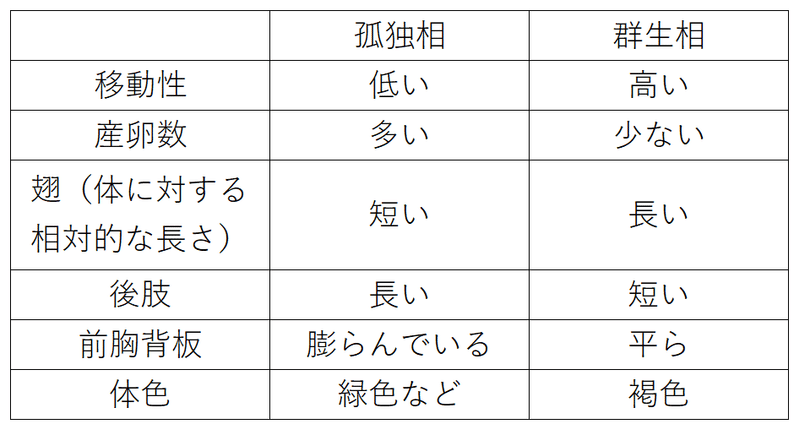
*バッタの相変異が有名である。低密度の時出現する型を孤独相(こどくそう)、高密度の時出現する型を群生相(ぐんせいそう)という。
語呂「そう?へん?茶色の軍手(相変異、茶色の群生相)」
*群生相は、「少数の卵を産むことで、体の大きな個体を残し、多数いる他個体との競争に負けないようにする」「新たな餌場を求めて、大移動しやすい形態をもつ」イメージを考えるとよい(ただし、群生相の形態が力学的に飛翔に有利であるかは、完全には明らかになっていない)。
雑談:相変異は、突然変異ではなく、進化ではない。相変異のように、1つの遺伝子型が環境条件に反応して表現型を変化させる能力は、『表現型の可塑性(かそせい)』と呼ばれている。
雑談:バッタにおいて、群生相の個体の子供はお互いに惹かれ合い、群れを成す。この高密度による正のフィードバックを介して、さらに群生相の特徴が増幅される(群生相は2~3世代にわたって維持される)。相変異は、個体間の相互刺激によって内分泌活動が変化する結果生じると考えられている。
雑談:一般に、相変異するものをバッタ、相変異しないものをイナゴと呼ぶ。『旧約聖書 出エジプト記』には「沢山のいなご」が登場するが、もし、その時現れたのが群生相なら、「沢山のいなご」ではなく、沢山の"バッタ"と言った方がよいかもしれない(冗談です)。
雑談:オンブバッタやショウリョウバッタは相変異を基本的に起こさないので、本来「イナゴ」と呼ぶべきである。
雑談:表現型に多型があることは珍しいことではない。(密度効果ではないが)たとえばミジンコは、ハエの一種に遭遇すると、巨大なヘルメットのような構造を頭に持つようになる。また、ある種の巻貝は、捕食者であるカニの発する化学物質にさらされると、貝殻が厚くなり、貝殻に歯のような構造が形成されるようになる。これらの変化は内分泌系によって引き起こされるとされているが、その仕組みは完全には明らかになっていない。
講義動画【密度効果】
雑談:相変異の原因として、視覚・匂い、物理的な他個体との接触・他個体の体表に存在する化学物質との接触など、さまざまな原因が考えられてきたが、現在では、主に他個体の体表面にある化学物質との接触や、視覚による個体群密度の認識などが関係していると考えられている(その後、体内におけるホルモン分泌の変化が、様々な形態や行動の変化、つまり相変異に繋がっていくとされる)。面白いことに、その体表面に存在する化学物質は、種を超えた共通性があるらしい。
雑談:相変異がどの程度の時間で起きるのかについても研究が進んでいる。かつては数世代にわたり少しずつ何らかの生理物質の蓄積が起こると考えられていたが、現在では一世代で相変異が起こることが知られている(ただし、短期間の混み合いの効果や、混み合いを感受する厳密な時期については、よくわかっていないことも多い)。
雑談:ロシアの昆虫学者ウバロフ卿は、ワタリバッタ類について、それまで別種とされていた大発生時にみられる群移動性の個体と、平年にみられる非集合性の定住的な個体とが同種であることを明らかにした。そして、それぞれを群生相、孤独相と名付け、孤独相から群生相への相の転換が大発生と大移動の機構を解く鍵であるとの説を提唱した。
雑談:相の変化(孤独相⇄群生相)は、どの発育ステージでも起こることが知られている。
雑談:密度効果における相変異は、一般に、昆虫類をはじめとした動物に対して使用する用語である。たとえば植物が密度効果によって成長に差が出るような現象を指して「植物の相変異」などという用語は使うべきではない。
雑談:成長曲線について
個体数が非常に小さい状態から、個体群が増加する時、その変化は一般にS字型の曲線になる。それは、式①のような微分方程式を用いて説明できる。
式①
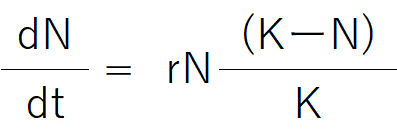
r:内的自然増加率(環境が制限的でない、つまり空間・食物・他の生物が制限要因として働いていないときの固有増加率)
N:個体数
K:環境収容力(増加率0に達したときの個体数上限値)
t:時間。
(dN/dtは、個体数が増えようとする勢いのようなイメージ)
NがKに達したとき、つまりN=Kの時、式①の(K-N)=0になるので、右辺は0になる。dN/dt=0となり、時間に対するNの増減の勢いは停止する(時間が経っても個体数が変わらなくなる)。KがS字の上部漸近線(頭打ちとなった値、個体数の最大値)であることが確かめられる。
資源が十分あるとき(Kが非常に大きい時、あるいはNが非常に小さい時)は、「(K-N)/K」=「1ーN/K」は1に近くなり、dN/dt=rNで近似できる(Nはどんどん増えていく)。
少し詳しく解説するが、高校生は知らなくてよい。
まず、何の制約もなければ、個体数の増加率dN/dtは現在の集団のサイズNに比例する(比例定数をmとする)。
dN/dt=mN・・・式②
このmは、人口が指数的に増加するという傾向をもつと主張した経済学者マルサスにちなんで、マルサス係数と呼ばれる(そもそも、この理論は、人口の変動の推定に用いられたものである)。
しかし、現実には、生物集団がいつまでも指数的に増えていくのは不可能である。環境の悪化や餌の不足などが起こる。
そこで、高密度になると、増殖が停止するという、現実に近い仮定を考える。
この仮定を表すために、式②のmの代わりに、「Nとともに減少する関数」として、「r(1-N/K)」を置こう。すると式②は
dN/dt=r(1ーN/K)N・・・式③
=rN{(K-N)/K}
となり、式①が導かれる。
式③のrは、個体数Nが十分に小さい時(1-N/Kが1に近い時)の増加率を表しており、内的自然増加率と呼ばれる(正の定数である)。
NがKに近づくほど、式③の「r(1-N/K)」は減っていく(0に近づく)。
Kは環境収容力と呼ばれる。
N=Kとなる時
r(1-K/K)=r(1-1)=0
となり、dN/dt=0となる。これは、個体数NがKとなった時、Nが変化しようとする勢いがなくなることを表している。
要点:個体数を調査する方法には、区画法や標識再捕法がある。
● 個体数の調査方法
① 区画法:生息地域内の一定の広さの区画内の個体数をかぞえ、その結果から全体の個体数を推定する方法。植物や、動きの遅い動物の個体数の調査に適している。
② 標識再捕法(ひょうしきさいほほう):1回目に捕獲した全個体に標識をつけてから放す。
標識した個体と標識していない個体が混ざりあった後に、再び捕獲(2回目の捕獲)し、捕獲した個体数に対する標識個体数の割合から、全体の個体数を求める。移動性の高い動物などの個体数の調査に適している。
(1回目に標識した個体数)/(そのエリアにいる全個体数)
=(2回目に捕獲した時に標識されていた個体数)/(2回目に捕獲した個体数)
の式に当てはめて、(そのエリアにいる全個体数)を求める。
式を暗記するより、以下の動画でイメージを理解しておくとよい。
講義動画【標識再捕法】
● 一般に、以下のような条件が標識再捕法を行うために必要な条件とされる。
①標識個体と未標識個体の捕獲のされやすさに差がないこと。
②調査期間中に個体の出入り・出生・死亡がないこと。
③標識が個体の行動・生理に影響しないこと。
④個体の移動が活発で、標識個体と未標識個体が均等に混ざり合うこと。
⑤標識が消失しないこと。
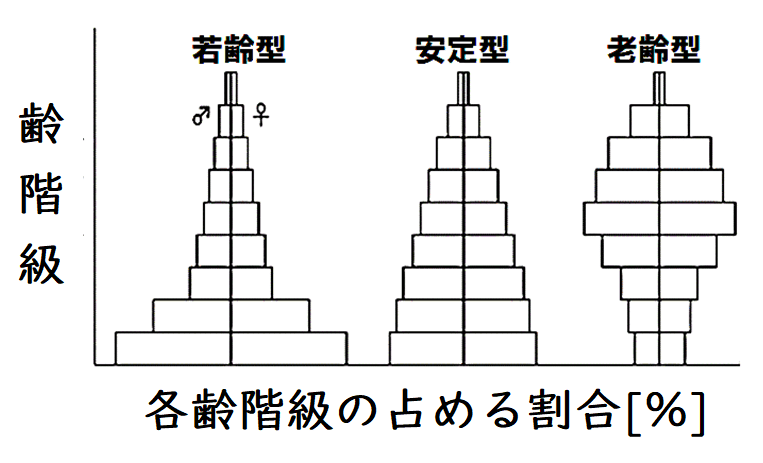
要点:年齢ピラミッドには3つの型がある。
● 総個体数に対する各齢層の割合を示したものを齢構成(れいこうせい)という。齢構成は下図のような年齢別に積み上げた年齢ピラミッドで表され、 3つの型に分けられる(難問はほぼ出ない。見ておくだけでOK)。
● 一般に、急速に拡大している個体群は、若齢個体を多く含み(若齢型)、安定した個体群は各年齢段階が比較的均一であり(安定型)、衰退している個体群は老齢個体を多く含む(老齢型)。
要点:生命表を用いることで、生存曲線が描ける。
(1)生命表
● 生まれた卵や子が、その後の時間経過とともにどれだけ生き残るかを示した表を生命表といい、これをグラフにしたものを生存曲線という。
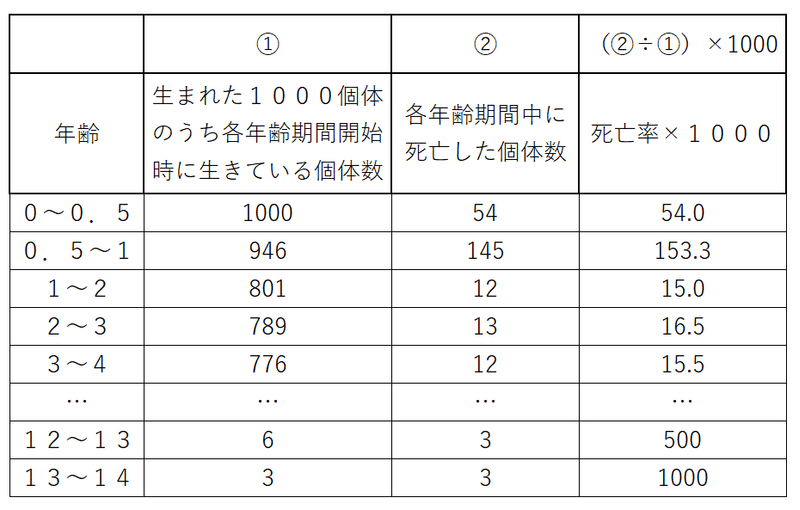
*下表はヒツジの生命表のイメージ(生命表の様式は問題ごとに異なるので、試験中に数値の意味をつかむこと。複雑なものは問われない)。
なお、死亡率は一般に「単位時間当たりの死亡数をその期間の最初の個体数で割った値」である。
たとえば、下表において、年齢「0.5~1」の死亡率は
145÷946=0.15327696=約0.1533
であり、これに1000をかけた値(=153.3)が表に記載してある(別に死亡率に1000をかけなければならないという決まりはない。書き方は論文ごとに異なる)。
問題:以下の生命表の空欄x、yに入る値として最も適切なものを、それぞれ以下の候補から選び答えよ。
候補【 1、5、12、24、50、75、100、500、800 】
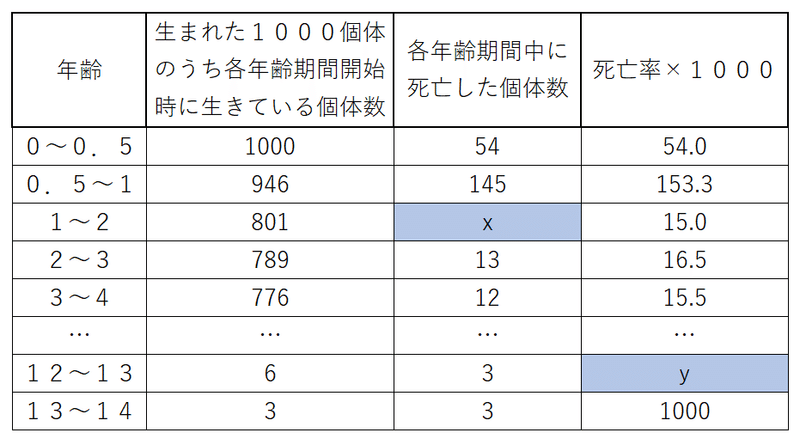
答えと解説
x:12
その年齢期間のはじめに801匹いた。次の年齢期間のはじめには789しかいなくなっていた。死亡数は
801-789=12匹
死亡率から求めてもよい。
x/801×1000=15
x=約12
y:500
死亡率は3÷6=0.5
これに1000をかけた値は
0.5×1000=500
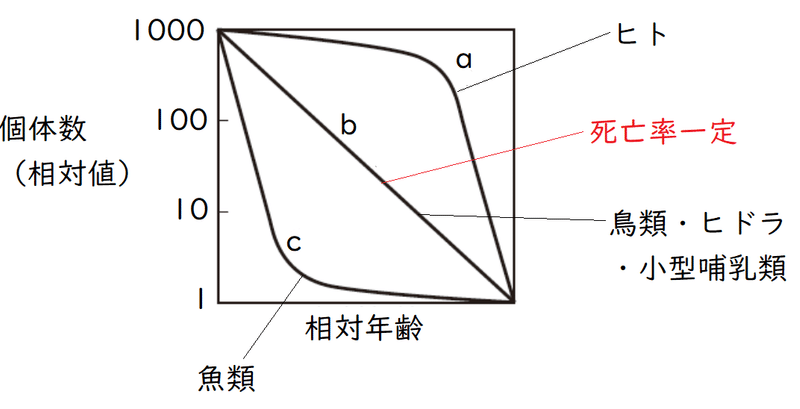
(2)生存曲線
● 生存曲線は次の3つの型に分けられる。生物例が問われる。
(a) 晩死型:幼時期に親の保護が厚く、幼時期の死亡率は低い。多くのものは理想的条件下での寿命(生理的寿命)近くまで生きる。
例 ヒト、サル、社会性昆虫など
(b) 平均型:一生を通じて死亡率がほぼ一定(死亡「数」一定ではない!!!)。
*たとえば、一定の期間ごとに、生存数が1/10になっている。
例 鳥類、小型の哺乳類、ヒドラなど
*ヒドラは、出芽で増えるため、新個体が親個体に守られることになる。
雑談:現実には、全寿命にわたって死亡率が一定の生物は存在しないが、多くの鳥やハツカネズミ、ウサギなどは直線に近い。
(c) 早死型:産卵数は多いが、幼時期に親の保護がなく死亡率は高い。
例 多くの無脊椎動物(カキなど)・魚類
(魚類がものすごい数産卵することは有名ですね。)
講義動画【生命表・生存曲線】
雑談:生命保険のシステムが発明された時、保険会社は、ある年齢の人が、平均であとどれくらい生きることができるかを調べ始めた。そして、人口統計学者によって、『生命表』が開発された。その後、Raymond Pearl はショウジョウバエの実験データに生命表を適用した。現在では、様々な生物について生命表が作成されてる。
雑談:蝶のような完全変態昆虫の生存曲線は、階段状になる。死亡率の高い卵の時期、蛹化の時期、短命な成虫の時期に、急激に傾斜する。
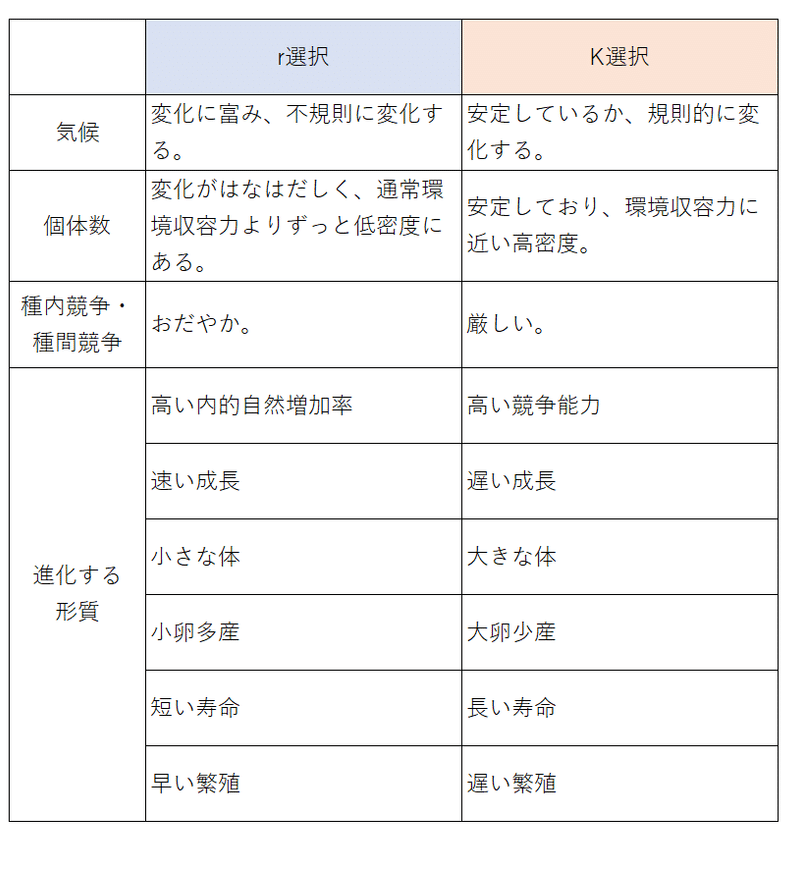
雑談:r選択とK選択、r戦略とK戦略について
・r選択とは、種内競争が激しくないとき、個体の潜在的な増殖能力を高めるように働く自然選択である(集団が環境収容力より低い密度にある条件下で作用する淘汰)。
・K選択は、種内競争が激しいとき、種内競争を勝ち抜くように働く自然選択である(集団が常に環境収容力に近い高密度で維持されている場合に作用すると考えられる淘汰)。
・rは内的自然増加率、Kは環境収容力を表している。
・このrーK選択説(マッカーサーとウィルソンが提唱)は、生物進化の方向について、以下のような方向性があるということを提案している。
①不安定な環境では、個体数の増殖能力(r)を進化させる方向に選択圧がはたらきやすい。
②安定した環境では、個体数の限界値(K)を進化させる方向に選択圧がはたらきやすい。
・ピアンカという学者は、r選択とK選択によって進化した生物(r戦略者、K戦略者)が、それぞれどのような特徴をもって、環境へ適応しようとするかについての仮説を提唱した。
・r選択は、不安定で変動が大きい環境で作用しやすく、K選択は、安定な環境で作用しやすいとされる。
・r選択が作用しやすい変動の大きい環境では、潜在的な増殖能力が非常に重要になる(正確な表現ではないが、このような環境では、最も対策しなければならない敵は、「環境の変動」である。「厳しい環境」が生き延びる上で最大の敵である。死亡率は密度とはあまり関係なく、不規則に変動する気候などに依存する)。
・K選択が作用しやすい安定した環境(個体数はK[限界値]に近い所にまで増加している)では、種内競争能力を高めることが重要になる(正確な表現ではないが、このような環境では、最も対策しなければならない敵は、「周りの奴等」である。自分が安定して住みやすい、増えやすい環境だということは、周りの奴等にとっても住みやすい、増えやすい環境だということである。なので、種内競争が厳しくなる。強い子を残し、競争に勝たなければならない)。
・ピアンカは、
「r選択のもとでは、小さい卵で多産・早熟で早い繁殖・短い世代時間・小さな体(サイズ)などの性質が進化」し(どんどん増える方が有利)、
「K選択のもとでは、大きい卵で少産・晩熟で遅い繁殖・長い世代時間・大きな体などの性質が進化する」と予測した(周りの奴らよりも強い方が有利)。
前者のような性質を r戦略、後者のような性質をK戦略と言う。いくつかの生物種で、この仮説を支持する現象が報告されている。
・たとえば、マウスはr戦略者(多数の小さい子供をつくり、寿命が短い)、ゾウはK戦略者(少数の大きい子供をつくり、寿命が長い)と言える。また、荒原の植物はr戦略者(小さく、寿命が短い)、極相林の高木はK戦略者(大きく、寿命が長い)と言える。
(同じ種の中にもr戦略者とK戦略者がいる場合もある。ある種のトカゲのメスには、遺伝的に決まっている2つのタイプが存在し、片方のタイプは多数の小さな卵を産むr戦略者であり、もう一方のタイプは少数の大きな卵を産むK戦略者である。)
*r―K戦略に関しては、わかっていないことが多く、高校範囲外である。
*実際に個体の特徴がどのような方向に「進化」していくのかについては、影響される要因が非常に複雑で多いので、簡単に予想することはできない。
雑談:島の生物地理学
● マッカーサーとウィルソンは島の生物地理学理論(島の平衡モデル)を提唱した。
ポイントは以下の通り。
① 島に存在する種の数は、大陸からの新たな種の移入・定着と、島にいる種のランダムな絶滅の平衡関係で決まる。
②島にすでに生物種がいるほど、移入速度は小さくなり(ある生物が島にたどり着いた時、その島にすでに同じ種がいる場合は移入に数えない)、絶滅速度は大きくなる。
③島が小さいほど種の絶滅速度は大きい。(島が大きければ、個体群も大きくなるし、様々な環境が島内にあるので、絶滅を避けることができる)。
④大陸から遠い島ほど種の移入・定着速度は小さい。逆に近い島ほど大きい。
*一般に、 小さい島では移入速度が低くなること(たとえば、とても小さい島に植物の種が運ばれてきて、偶然その小さい島に落ちるのは稀である)、 近い島では消失速度が小さくなること(島で消失した種が陸から再侵入することがあるので) も知られている。
下図には、島の大小・島の大陸からの遠近の組み合わせによる4つの平衡点が示されている。たとえば、陸から遠くて、小さい島では、①のところの種数(A)がその島に存在すると予想される。
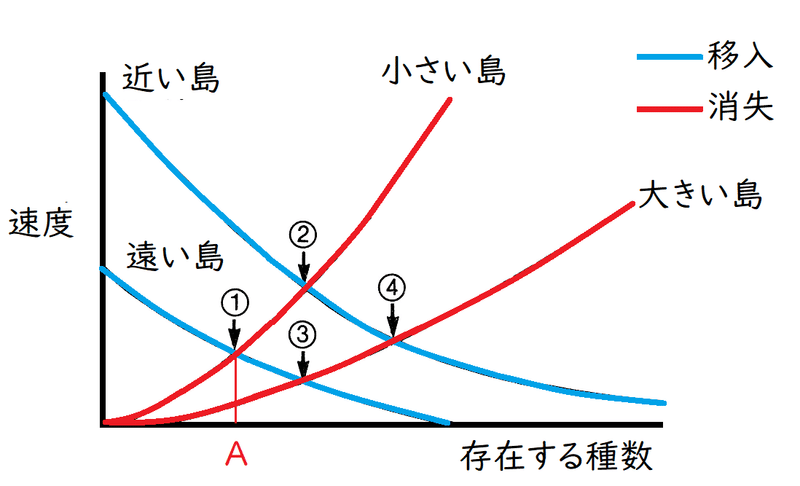
*もちろん、このモデルでは、撹乱、進化的適応などの様々な要因を無視している。 しかし、このモデルは、島でなくとも、島のように孤立した生息地に当てはまる。たとえば、我々人の影響で分断化された森林や公園などの環境が、どのくらい多くの種を保存できるかという予測を可能にする。
講義動画【島の生物地理学理論】
まだわかっていないこと
● バッタの相変異の原因について、完全には明らかになっていない。
● 医学の進歩や栄養状態の改善によって、人類の生存曲線はどのように変化していくか。そしてそれは社会にどのような影響を与えるか。
● 繁殖様式や出産時期の変化は生存曲線にどう影響を与えるか。
● 魚などの生物では、急に、卵や幼魚の生存数が異常に多い年齢階級(団塊の世代)が現れることがある。その原因は何か。
● ギャレット・ハーディン(生態学者)は、「たとえどれほど小さくとも、指数的に増えるものには最大限の敬意を払わねばならぬ。」と言った。今後、世界の人口はどのように変化していくか(人口の成長曲線はどうなっていくのか)。