
【高校生物】遺伝①「減数分裂とはどのような分裂か?」

~プロローグ~
「遺伝を司る法則についてはまったくわかっていない。・・・生まれた子の特徴が、親ではなく祖父母やさらに遠い祖先に似る、いわゆる先祖返りがしばしば生じる理由もわかっていない。」ダーウィン『種の起源』より
先祖返りの一種に、隔世遺伝(ある個体の形質が祖父または祖母に似ること)がある(珍しい現象ではない。身の回りで目にすることもあるだろう)。F2における劣性形質の分離は隔世遺伝の例である。たとえば、遺伝子型AAのオスと、遺伝子型aaのメスから生じた子(遺伝子型Aa)同士を交配させるとする。すると、遺伝子型AA:Aa:aaの個体が1:2:1の比で生じてくる。孫の代の遺伝子型aaの個体は、親(遺伝子型Aa)よりも、むしろ祖母(遺伝子型aa)に似ている。遺伝する「因子」、すなわち遺伝子を考えることで、このような隔世遺伝を簡単に説明できる。
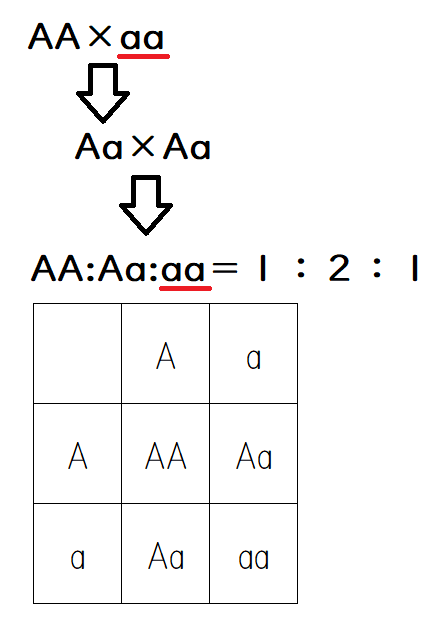
*学校によっては、遺伝子型Aa同士の交配を、上のような表ではなく
(A+a)²=(A+a)(A+a)=AA+2Aa+aa
のように表記して解説することもある。やっていることは変わらない。
もし、「混血」や「純血」という概念が正しいとするならば(交配が繰り返されるたびに血が混ざっていくと考えるならば)、隔世遺伝のような現象を説明することは難しい。オレンジジュース(遺伝子型AAの個体)とリンゴジュース(遺伝子型aaの個体)を混ぜると(交配させると)ミックスジュース(遺伝子型Aaの個体)ができる。通常、生じたミックスジュース同士を混ぜても、ミックスジュースしか生じないはずである。しかし、現実には、生じたミックスジュース同士から(遺伝子型Aa同士の交配から)リンゴジュース(遺伝子型aaの個体)が生じるのである。遺伝現象を、(ジュースや血などの)何らかの液体の混ざり合いと考えている限り、この現象に適切な説明を与えることはできない。実際は、DNAに乗った遺伝子(塩基配列)が後世に伝わっていくのである。血がその人の体の特徴を決めるのではない。体(主成分はタンパク質)の設計図は、DNAの塩基配列だったのである。
「減数分裂によって生命のサイクルは完成する。身体は朽ちて、死ぬ。しかし、減数分裂によって形成された配偶子は親の死を生き残り、次の世代をつくる。有性生殖、進化に伴う変化、世代間の形質の伝達、これらはすべて減数分裂に帰着する。」ギルバート『ギルバート発生生物学』より
★テストに出やすいワード
①二価染色体
②対合
③精原細胞・卵原細胞
④第一極体・第二極体
⑤重複受精
要点:減数分裂は間に間期を挟まない2回の連続した分裂であり、DNA量、染色体数が半減する。
● 間期
S期にDNA量が2(相対値)から4に複製される。
*核当たりのDNA量はG1期を2とする。
*DNA複製を終えた染色体は、2本の姉妹染色分体から構成されるようになる。これらの染色分体は、コヒーシンというタンパク質によって接着している(下図はイメージ。1本の染色分体には、1本のDNAが含まれる)。

以下、
①核相
②核当たりのDNA量(相対値)
③特徴
の順に記す。
(1)第一分裂
●前 期
①2n
②4
③相同染色体が対合して、二価染色体が形成される。相同染色体の間で染色体の一部が交換される(減数分裂第一分裂前期に染色体の乗換えが起こる)。

●中 期
①2n
②4
③二価染色体が赤道面に並ぶ。
●後 期
①2n
②4
③二価染色体が対合面で分かれ、染色体がそれぞれ両極へ移動する。

●終期
①2n(終了とともにnになる)
②4(終了とともに2になる)
③第一分裂が終わると、娘核の染色体数は半減(n)する。続いて、DNAの複製を行わずに第二分裂の前期に入る。
<間期なし。そのまま第二分裂に入る(DNA量を減らそうとしているのに、第一分裂と第二分裂の間にDNA合成期なんてあったら、何をしようとしてるかわからない)。>
(2)第 二 分 裂
●前 期
①n
②2
●中 期
①n
②2
③染色体が赤道面に並ぶ(中期と言えば「赤道面に並ぶ」)。
●後 期
①n
②2
③2本の染色体(染色分体)が縦裂面で分離して、両極へ移動する。
*この過程(染色分体が縦裂面で分離する)は体細胞分裂に似ている。

●終 期
①n
②2(終了とともに1になる)
③核膜が形成される。
● 減数分裂の結果、(DNA複製前の細胞に比べて、)染色体数・DNA量・核相が半分になった細胞が4個生じる。
*体細胞分裂と見比べよ(下図)。
*注意:DNA複製が終わった時点のDNA量を4としている(複製が起きる前のDNA量は、細胞当たり2)。


● 減数分裂におけるDNA量の変化
*2であったDNA量の相対値は、1になって(半減して)終わる(精子や卵などの生殖細胞になる)。

<参考>下図は体細胞分裂におけるDNA量の変化(体細胞分裂では、減数分裂と違って、分裂の前後でDNA量は半減しない。2から始まったDNA量の相対値は、また2に戻って終わる。体細胞分裂を繰り返している細胞では、同じ変化が繰り返し起こっている)。

講義動画【減数分裂に伴うDNA量の変化】
● 二価染色体は相同染色体が対合したものである。複製した2つのDNAはそれぞれ姉妹染色分体という。

上の図を見よ。第一分裂では、染色体の数が半減しているが、第二分裂では、染色体の数は半減していない(染色分体は分かれている)。染色体の数え方にはいろいろあるが、上の図で、第一分裂前が2本、第一分裂後は1本、第二分裂後は1本と数える数え方が一般的。

「染色体が何本あるか」「染色体が何本観察されるか」という問題は今はあまり出ない。数え方が統一されていないからである(同じ教科書の中でも数え方が異なることもある)。問題文で何も指示がなければ上の図のように考える。
<Q.どうして減数分裂第二分裂で、核相がnから半減しないの?…核相は、染色体が何組あるか?を表す数値である。簡単に判断するには、染色体に同形同大のペアがいれば2n、いなければnである。減数分裂前は、相同染色体(同形同大の染色体)はペアになって存在していた(同じ大きさで同じ形のX字の染色体が2本ずつあった)。しかし、第一分裂後は同形同大のペアがいないから核相はn。弟二分裂後も核相はn。>
雑談:一般に、有性生殖では、減数分裂や配偶子の接合によって、遺伝子の多様な組み合わせが生じ得る。環境が変化した時、有性生殖を行うことで、その環境に、より適応した新たな遺伝子の組み合わせを生み出すことができるかもしれない。一方、自身のクローンを生み出す無性生殖は、安定した環境で、すでに成功している遺伝子の組み合わせを確実に存続させることができる。また、無性生殖では交配相手が必要ないため、手間なく数を増やすという点では、有性生殖より有利であると考えられる。しかし、常に無性生殖を行う動物は少なく、多くの動物が有性生殖を行っている。遺伝的多様性を増大させる機構は、生物の存続にとってとても重要であるらしい。下図は有性生殖と無性生殖の大雑把なイメージ。

発展:ダウン症
減数分裂の目的の一つは、染色体数を半減させることである。しかし、時には染色体の分離がうまくいかず、異数体が生じることがある。ダウン症候群は21番染色体が1本多いトリソミー(3個の相同染色体をもつ)である。21番染色体以外のトリソミーは、出生前に致死となる場合が多い。
雑談:正確には、対合面は下図1のようになる(図1には染色体の乗換えは描いていない。二価染色体には4本の染色分体[姉妹染色分体が2本ずつ]が含まれる。乗換えについては後に学ぶ)。
図1

下図2はキアズマ(下図の染色体が交差している部分)や動原体を描き足して少しリアルにしたイメージ。
図2

要点:染色体の構成を表したものを核相という。
(1)核相
● 染色体の構成を表したものを核相という。
● ほとんどの動物と数多くの植物は、すべての体細胞が対になった相同染色体をもっているので、2倍体と言われる。
● ヒトの体細胞の染色体の総数は46本である(精子や卵にはその半分の23本が入っている)。
● ヒトの体細胞の核相は2n=46である(=の左には、染色体のセット数、=の右には、染色体がいくつあるかを書く)。卵や精子ではn=23である。
雑談:上のような表し方では染色体構成を表せないような細胞も存在する。たとえばがん細胞などは、異常な数と形の染色体を不規則に持つことが知られている。
雑談:nを基本的な単位と考えている。そこに遺伝子の1シリーズがあるからである(たとえば、ヒトの卵の核相はn=23だが、この23本の染色体には、すべての種類の遺伝子が入っている。受精卵は同じ種類の遺伝子を2つずつもつ)。
講義動画【核相】
<Q.無性生殖って何?…自身のクローンをつくる、配偶子の接合によらない生殖。アメーバやイソギンチャクの『分裂』、酵母やヒドラの『出芽』、ジャガイモやユキノシタの『栄養生殖(根・茎・葉などの栄養器官の一部から新個体ができる)』などがある(さし木も栄養生殖)。最近はあまり問われない。>
発展:栄養生殖
栄養生殖に使われる構造には以下のようなものがある。昔はよく問われたが、最近はあまり問われない(ただし、いつも食べているジャガイモが茎、サツマイモが根であることは知っておくとよいかも)。
①ユキノシタの走出枝(ほふく茎、ほふく枝と呼ばれることもある。茎が地面に沿って長く伸び。その先が新しい個体になる)。
②オニユリのむかご(親植物体の一部から無性的に形成され、その後独立した個体となるものの総称を「むかご」という。むかごの起源は様々だが、オニユリの場合は肉質の鱗片葉が茎をとりまいたもの[鱗芽]である)。
③ジャガイモの塊茎(「かいけい」と読む。地下茎の一部あるいは全体が異常肥大したもの)。
④サツマイモの塊根(「かいこん」と読む。塊状根ともいう。根の一部が異常に肥大して塊状になったもの)。
● 1組の染色体をもつ細胞は1倍体(n)とよばれる。配偶子である卵と精子は1倍体である。
● 1倍体を単相、2倍体を複相などともいう。
● 多くの真核生物の体細胞の核相は2nである。
● 核相3nの細胞は、たとえば被子植物のつくる胚乳などがある(後に学ぶ)。
雑談:体細胞が3倍体の生物も存在するが、奇数に染色体をもつ生物は、正常な減数分裂を行えないことが多い(奇数は半分に割れない)。食用のバナナは3倍体で、染色体を3nに持ち、無性生殖で増える。
雑談:どうして核相などという概念を使うのか理解に苦しむだろうが、「この細胞の中には、同じ形の染色体が、それぞれ2本ずつ入っていて、それで、全部で4本ある。」などと言うより、「2n=4」と言った方が簡単である。
(2)配偶子の種類
● 一般に、減数分裂で生じ得る配偶子の種類は2のn乗種類である。たとえば、2n=4の生物の場合、(2n=4は数式ではないのだが、数式のように、無理矢理両辺を2で割ったとして)n=2なので、2の2乗=4種類の配偶子が生じる。
*相同染色体2本のうち、どちらかがランダムに精子や卵に入ると考える。相同染色体がn対あれば、その選び方は2のn乗である(2本の相同染色体はほとんど同じDNA配列をもつが、完全に同一ではない)。
*性染色体については考えていない。しかし、やはり2本ある性染色体も、相同染色体と同じように1本ずつ配偶子に入る(ヒトのオスでは、X染色体が入った精子とY染色体が入った精子が作られる)。XO型のオスやZO型のメスなど、性染色体の数が奇数になっているような場合も、性染色体が入る配偶子と入らない配偶子が生じる(性染色体については後に学ぶ)。
*ヒトは2n=46(n=23)なので、2の23乗種類の配偶子(卵または精子)をつくる(実際は、後で学ぶ組換えが起きるので、もっと種類は多くなる)。膨大な種類(2の23乗種類)の卵と精子が生じ、さらにそれらが受精するので、組換えは考えないとしても、
(2の23乗)×(2の23乗)=約70兆種類の受精卵が生じ得ることになる。
● 組換えが1回起こったら2の(n+1)乗種類になる(組換えについては後に学ぶ)。
試験中忘れたら2n=4の細胞などを想定して、下図のように、実際に減数分裂させてみればよい。
①2n=4(n=2)で、組換えが起こらない場合

②2n=4(n=2)で、1回乗換えが起こった場合

講義動画【体細胞分裂の復習】
要点:精子や卵は始原生殖細胞からつくられる。1個の一次精母細胞から4個の精子が生じ、1個の一次卵母細胞から1個の卵が生じる。
(1)配偶子の接合
● 合体することで新個体になる細胞を配偶子(はいぐうし)という。配偶子同士が合体することを接合という。
● 同形配偶子接合では、同形同大の配偶子が合体して新個体になる(クラミドモナスなどが行う)。
● 異形配偶子接合では、形や大きさが異なる配偶子が合体して新個体になる(アオサ、ミルなどが行う)。
● 配偶子が精子と卵といいうる程度にまで分化している場合、それらの配偶子の接合を、特に『受精』と呼ぶ。その場合、接合子を特に『受精卵』と呼ぶ(ヒトなどなどが行う)。

<Q.異形配偶子接合=受精でよい?…よくない。異形配偶子には、様々な段階があり、上図のように、雌性配偶子が運動性を持ち、卵にまで分化していないものもある。異形配偶子が卵(卵はふつう運動性を失っている)と精子にまで分化している場合、その接合を、特に受精と言う。卵と精子は異形配偶子のひとつと考えていいのだが、よほど特殊な文脈で議論しない限り、卵や精子を異形配偶子と呼んだり、受精を異形配偶子接合と呼んだりすることは少ない。>
<Q.精子や卵の定義は?…厳密には決まっていない。「多細胞生物の雄性配偶子のうち、雌性配偶子と形態的・機能的に著しい差を示し、小形で運動性をもつもの」を精子と言い、「雄性配偶子が精子と呼ばれる場合に、それに対応する雌性配偶子」を卵と言うことが多い。生物種ごとに様々な卵・精子が存在し、どこまでを卵、精子と呼ぶかは難しい議論なのだが、高校生は気にしなくていい。>
雑談:「これから減数分裂をはじめるよ!」という細胞の名前には「母」が付くことが多い。
例)一次精母細胞、一次卵母細胞、花粉母細胞、胚のう母細胞
(2)精子の形成
● 始原生殖細胞(2n)→精原細胞(2n)(体細胞分裂で数を増やす)→一次精母細胞(2n)→(減数分裂第一分裂)→二次精母細胞(n)→(減数分裂第二分裂)→精細胞(n)→(変態)→精子(n)
*精原細胞の一部が一次精母細胞になる。
*精細胞は変態して精子になる。
雑談:精子への変態については、よくわかっていないことも多い(ゴルジ体から先体が形成される。中心体からつくられる鞭毛が伸びていく。細胞質の大部分は捨てられる)。


(3)精子の構造
● 精子の頭部の前方には先体(先体胞)が、後方には核がある。先体はゴルジ体に由来する。
雑談:核の先端、または核を覆うようにある構造を先体という。ウニなどの海産動物では、先体は、前方のゴルジ体由来の膜に囲まれた先体胞(先体胞の核側の膜を先体内膜、外側に面している膜を先体外膜という)と、後方の未重合のアクチン(アクチンモノマー)が存在する部分からなる(アクチンは受精の際に重合してアクチンフィラメントになる。ウニの受精については後述する)。なお、先体胞と先体を同義とすることも多い。覚えなくてよいが、下図はイメージ。

● 頭部と中片部の間には中心体がある。べん毛は中心体から伸長する。
● 中片部にはミトコンドリアがある。
講義動画【精子の形成】
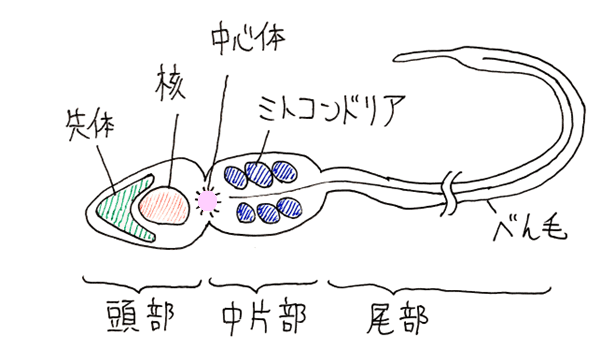
雑談:上図の中心体から伸びている線は軸糸。軸糸は微小管によって形作られている(9+2構造をとっている)。
(4)卵の形成
● 始原生殖細胞(2n)→卵原細胞(2n)(体細胞分裂で数を増やす)→一次卵母細胞(2n)→(減数分裂第一分裂)→二次卵母細胞(n)→(減数分裂第二分裂)→卵(n)
*卵原細胞の一部が一次卵母細胞になる。
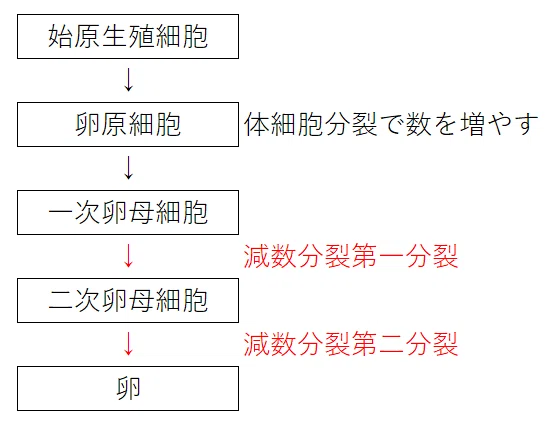

● 一次卵母細胞から減数分裂第一分裂で第一極体と二次卵母細胞が生じる。これは不等分裂である。栄養が二次卵母細胞に集中する。
● 二次卵母細胞から減数分裂第二分裂で第二極体と二次卵母細胞が生じる。これは不等分裂である。栄養が卵に集中する。
*第二極体が生じた位置を動物極、その反対を植物極という。
雑談:動物極側には卵黄があまり含まれていないので、よく卵割が起きる。そのため、よく『動く』という意味で、『動物極』と名付けられた。その反対側の極は『植物極』という。
*第一極体は分裂する場合もあればしない場合もある。
● 第一極体、第二極体は退化・消失する。(良く問われる)

雑談:ヒトの女性では、一次卵母細胞はすぐに減数分裂に入るが、「第一分裂前期」の段階で休止状態となる。一次卵母細胞は、第一分裂前期にとどまったまま卵黄を蓄積し、卵とほぼ同じ大きさにまで成長する。女性が成長し、思春期が始まると、また減数分裂を再開する。第一分裂が完了すると「第二分裂中期」へと進行するが、この段階で二次卵母細胞が排卵される。そして、受精した場合に、また減数分裂を再開する。
雑談:成長を開始した一次卵母細胞の核は、次第に大きくなり、卵核胞(らんかくほう)と呼ばれるようになる。
講義動画【卵の形成】
問題:100個の一次卵母細胞から、理論上、何個の卵ができるか。
答え:100個
問題:100個の一次精母細胞から、理論上、何個の精子ができるか。
答え:400個
要点:ウニの受精では、卵黄膜が受精膜となる。
● ウニの受精
(ウニの場合、減数分裂の終わった卵に精子が進入する。)
①ゼリー層に精子が到達すると、精子の頭部にある先体胞(先体)がエキソサイトーシスを起こし、中身(タンパク質を分解する加水分解酵素など)を放出する。タンパク質分解酵素によってゼリー層が分解される。
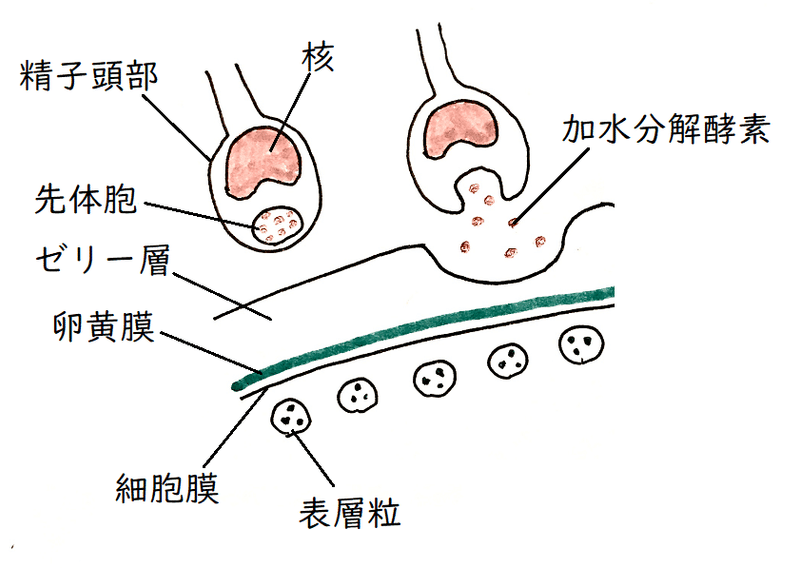
②精子の先端から先体突起が伸びる(細胞膜の先端が伸びて突起状に変化する)。先体の中身の放出と、先体突起の伸長を先体反応という。
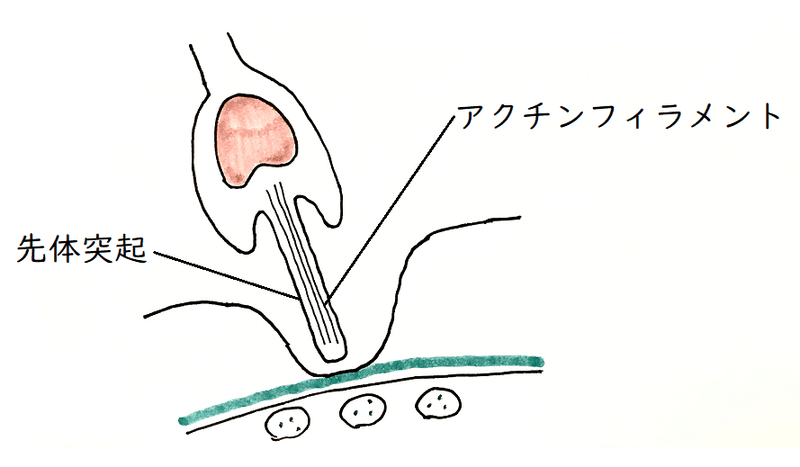
雑談:ウニでは、核の前に蓄えられていたアクチンが重合してアクチンフィラメントが形成され、細胞膜(先体胞の核側であった部分)を押し上げながら伸長する。そうして、精子の先端から伸びる糸状の構造ができる。その構造を先体突起という。先体突起の先端には、バインディンと呼ばれるタンパク質が存在し、バインディンはウニの卵黄膜に存在するバインディン受容体と結合する(バインディンは同種・異種の認識に関わる。「バインディンとバインディン受容体」の「鍵と鍵穴」の関係は重要である。特に体外受精を行うウニの場合、周囲の水には他種の配偶子が存在するかもしれない)。
③精子と卵の細胞膜が融合する。精子と卵の細胞膜が融合する部位が盛り上がる。その部分を受精丘(じゅせいきゅう)という。
下図はイメージ。

④表層粒の中身が、卵の細胞膜と卵黄膜の間に放出され、卵黄膜を押し上げる(表層反応)。卵黄膜は受精卵になる(細胞膜が受精膜になるのではない)。受精膜の形成により、他の精子による受精が防がれる・・・遅い多精拒否(多精、すなわち2個以上の精子が1個の卵と受精してしまう事態を防ぐ仕組み)



雑談:表層粒のエキソサイトーシスによって、ムコ多糖類や、様々な酵素が分泌される。セリンプロテアーゼという酵素は、卵黄膜と細胞膜を連結しているタンパク質を切断する。さらに、ムコ多糖類が卵の細胞膜と卵黄膜との間に放出されることによって、(浸透圧の勾配が生じ、水が入り込むことで)卵黄膜が押し上げられる。また、表層粒から放出されるトランスグルタミナーゼという酵素は、卵黄膜を硬化させ、受精膜に変える反応に関与する。
発展:早い多精拒否
早い多精拒否は、膜電位の変化による。精子が進入すると、Na+が高速で流入し、細胞内の電位が細胞外に比べて、マイナスからプラスへ変化する。この電位の変化が多精拒否に働く(受精前は、卵細胞内外の電位差はー70mVである。精子が卵に到着した数秒後には、膜電位は正の方向に変化する。この膜電位は1分程度しか維持されない。その間に遅い多精拒否[受精膜の形成]を完成させる)。
下図は受精前後のウニ卵の膜電位のイメージ。

講義動画【ウニの受精】
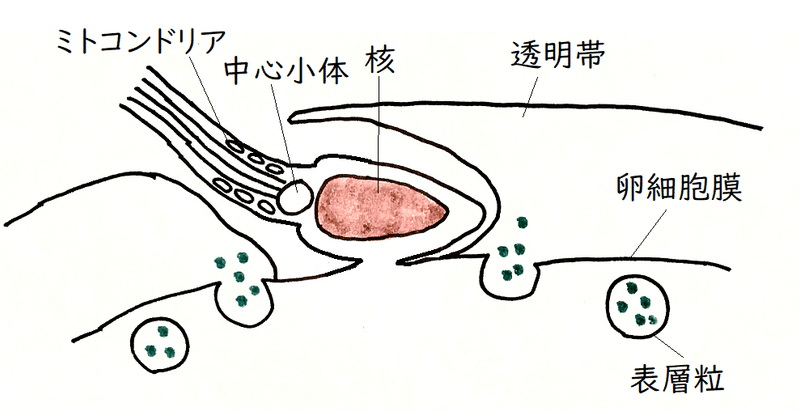
雑談:哺乳類の受精
哺乳類では、下図のように、卵と結合するのは、精子の頭部の先(ウニでは精子は卵に垂直に侵入する)ではなく、精子の頭部の側面である。高校生は知る必要はないが、マウスの受精では、以下のようなことが起きていると考えられている(わかっていないことも多い。哺乳類は体内受精を行うので、受精の研究がしにくい面もある)。
①透明帯は、ZP1、ZP2、ZP3という3つの糖タンパク質などからできている。ZP3と精子が結合することにより、精子における先体反応が誘導される(と長い間考えられてきたが、近年では、透明帯に精子が結合する前に先体反応が起こっているらしいことが明らかになっており、ZP3の役割の見直しが行われている。透明帯のどの糖タンパク質がどのような機能を持っているのかについては、完全には解明されていない)。先体反応によって精子頭部に含まれていたさまざまな酵素を含む内容物が放出されると、それらの酵素などの働きによって、精子は透明帯を通過しやすくなる。
②表層粒からさまざまな酵素が放出(エキソサイトーシス)される。放出された酵素は、透明帯の性質の変化を触媒する。透明帯の性質の変化は、「遅い多精拒否」として機能する(1つの精子が卵に入ると、他の精子は透明帯と結合することも結合したままでいることもできなくなり、速やかに払い落とされてしまう。なお、哺乳類の「早い多精拒否」についてはよくわかっていない[哺乳類では、ウニで見られたような電気的な早い多精拒否機構は検出されていない])。
*多くの哺乳類(及び両生類)の精子は、減数分裂第二分裂の中期で停止している卵母細胞に進入する(ウニなどの棘皮動物では、精子は減数分裂が完了した卵に進入する)。
要点:1個の花粉母細胞から4個の花粉が生じ、1個の胚のう母細胞から1個の胚のうが生じる。
● 被子植物の配偶子形成について
(1)花粉の形成
● 花粉母細胞(2n)から、減数分裂によって花粉四分子(n)(かふんしぶんし)が生じる。花粉四分子の4つの細胞はそれぞれ成熟した花粉になっていく。
*花粉母細胞の減数分裂によって生じた4つの細胞を、まとめて花粉四分子という。
● 花粉四分子(n)の細胞は特殊な体細胞分裂を行い、雄原細胞(n)(ゆうげんさいぼう)と花粉管核(n)(かふんかんかく)が生じる。
*成熟した花粉は、雄原細胞と花粉管核を含む。花粉の細胞質の中に、雄原細胞が遊離している状態になっている。
下図は成熟花粉のイメージ。
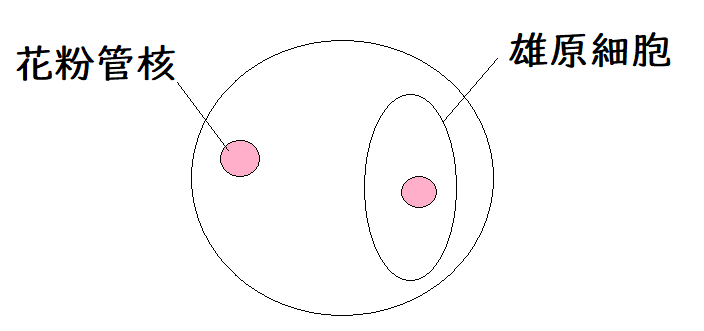
● 雄原細胞(n)は体細胞分裂を行い、2個の精細胞(n)が生じる。花粉管の先端には花粉管核がある。

(2)胚のうの形成
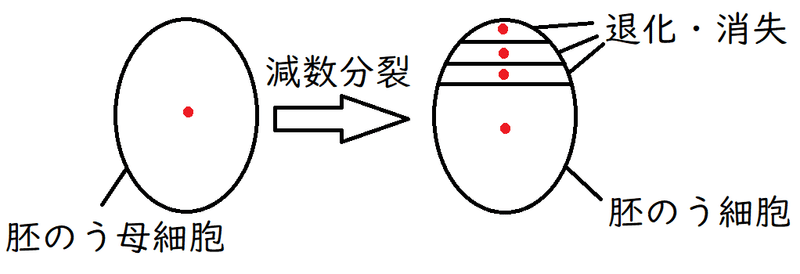
● 胚のう母細胞(2n)から、減数分裂によって胚のう細胞(n)が生じる。4つの細胞が生じるが、生き残り成長するのは1個だけで、残り3個の細胞は退化・消失する。
● 胚のう細胞は体細胞分裂を行い、胚のうが生じる。
詳細:胚のう細胞は、核だけの分裂(特殊な体細胞分裂)を3回行う(核が1→2→4→8個に増える)。そして(細胞膜や細胞壁が形成され)、3つの反足細胞(n)、2つの助細胞(n)、1個の卵細胞(n)、1個の中央細胞が生じる。中央細胞には2個の極核(n)(n)がある。
下図は完成した胚のうのイメージ。7個の細胞がある(核は8個ある)。

覚え方:だんだん文字が減っていく。
「胚のう母細胞 → 胚のう細胞 → 胚のう」
*助細胞(じょさいぼう)は、花粉管の誘引に働く(トレニアのルアーが花粉管誘因タンパク質として有名である)。
雑談:反足細胞(はんそくさいぼう)の役割は不明である。受精の前、あるいは直後に消失する。何の機能もない可能性もあるが、胚のうへの栄養供給に何らかの役割を果たしているとも言われる。
発展:ルアー
トレニアという植物を用いた実験によって、助細胞に花粉管を誘因するはたらきがあることが明らかになった(レーザーで、胚のうの細胞の1~2個の細胞を破壊し、花粉管の誘引頻度を調べた。結果、下の表のようになった。表から、助細胞が花粉管を誘導していることが示唆される)。
後に、助細胞から分泌される花粉管を誘因するはたらきをもつタンパク質が同定され、ルアー(英単語のlure[釣りのルアーと同じ]から命名)と名付けられた。
*助細胞が破壊されると花粉管の誘引頻度が減る。

雑談:名古屋大学の東山哲也教授らによってルアーが同定された。2009年3月19日発行の『Nature』(Nature 458, 7236)の表紙には、花粉管がルアーの跡をたどって引き寄せられ、Natureの頭文字である「N」の形に伸長している所が写っている。驚くべき写真である。検索して見てみてほしい。
雑談:ルアー様のタンパク質はシロイヌナズナでも発見されており、シロイヌナズナのルアーを発現させたトレニアの胚珠は、シロイヌナズナの花粉管を選択的に誘引することが明らかになっている。
雑談:ほとんどの被子植物では、胚のうを珠皮から取り外すことは不可能である。しかし、トレニアなどの一部の植物では、胚のうの一部が胚珠から飛び出して、むき出しになっている(卵細胞、2つの助細胞、そして中央細胞の約半分が胚珠の外部に出ている。下図はイメージ)。胚珠を植物体から切り離し単離すると、むき出しの胚のうが直接培地上に置かれる状況になるので実験に便利である。

(3)卵細胞ができるまでのDNA量の変化
● 下図は、花粉母細胞から精細胞が形成されるまでのDNA量の変化を表したもの。

①花粉母細胞が減数分裂を行い、4つの細胞(花粉四分子)が生じる。
②花粉四分子の4つの細胞はそれぞれ体細胞分裂を行い、花粉管核と雄原細胞が生じる(1回目の体細胞分裂)。
③雄原細胞は体細胞分裂を行い、2個の精細胞が生じる(2回目の体細胞分裂)。
● 下図は、胚のう細胞から卵細胞が形成され、受精に至るまでのDNA量の変化を表したもの。

①胚のう母細胞が減数分裂を行い、胚のう細胞が生じる(4つの細胞が生じるが、3つは退化・消失する)。
②胚のう細胞は核のみの分裂を3回行い(3回の特殊な体細胞分裂)、その後、卵細胞が形成される。
講義動画【植物の配偶子形成】
要点:被子植物は重複受精を行う。
● 被子植物は重複受精を行う。重複受精は、同時に二か所で行われる受精形式である。
雑談:現代(新生代)は被子植物の時代である。重複受精のしくみが、被子植物が繁栄した原因の一つと考えられている(もちろん、もう一つの原因は、花を咲かせ、昆虫を利用して花粉を運ばせるという戦略である)。 赤ちゃん(受精卵)と栄養(胚乳)を同時につくる重複受精は効率的である。結婚の予定もない、彼女・彼氏もつくる予定のない友達の部屋に、たくさんの粉ミルクだけが蓄えてあったら変である(冗談です)。胚と胚乳は、同時につくる方が、無駄がなくて良い。
● 重複受精は、被子植物特有の受精形式である。2個の精細胞が、それぞれ卵細胞、中央細胞と受精する。
①精細胞(n)+卵細胞(n)→受精卵(2n)・・・胚になる
②精細胞(n)+中央細胞(n)(n)→胚乳(3n)・・・胚の栄養になる
★精細胞(n)と中央細胞の2個の極核(n)(n)が受精するので、胚乳は3倍体(胚乳の核相は3nである)になる(非常によくテストに出る)。
*発展だが、裸子植物の胚乳の核相はnである。

雑談:胚乳は、たとえば白米の部分である。胚乳は人類の食糧の主な供給源である。胚乳はココナッツミルクの原料にもなっている。
講義動画【重複受精】
雑談:重複受精の詳細
重複受精については、わかっていないことが多い(また、植物によって少しずつ様式が異なる)。以下にシロイヌナズナの重複受精について詳しく記す。
①花粉管の破裂と内容物の放出が起こる。花粉管を受け入れる助細胞は崩壊する(ように見える)。
②2つの精細胞は卵細胞と中央細胞の境界付近に約7分間留まる。
③1つの精細胞は卵細胞と融合し、もう1つの精細胞は中央細胞と融合する。精細胞の核は、卵細胞の核、中央細胞の核に向かって移動する。
*なお、シロイヌナズナでは、中央細胞の2つの極核は受精前に融合している。
*精細胞の移送の様式についてはよくわかっていない。

問題:
(1)重複受精の利点は何か。
(2)重複受精を行う植物を以下から選べ。
①被子植物 ②裸子植物 ③シダ植物 ④コケ植物
答え:(1)胚乳と受精卵が同時に生じるため、栄養が無駄にならない。(2)①
● 受精卵は胚に、胚珠は種子に、珠皮(しゅひ:胚珠を包んでいる)は種皮(しゅひ)になる。
● 珠皮は雌親の体の一部である。すなわち、種皮は雌親の体細胞由来である。
問題:ある被子植物を用いて以下交配実験を行った。
遺伝子型AAの個体(雄親)から生じた花粉を、遺伝子型aaの個体(雌親)に受粉させ、種子を得た。
(1)得られた種子の種皮の遺伝子型は何か。
(2)得られた種子の胚乳の遺伝子型は何か。
答え:
(1)aa(種皮は雌親の体の一部[珠皮]からできる。したがって種皮の遺伝子型は、雌親の遺伝子型と等しい。)
(2)aaA(遺伝子型aaの雌親のつくる胚のうの極核の遺伝子型はa。2個の極核[a+a]と、1個の精細胞[遺伝子型A]が受精して胚乳が生じるので、胚乳の遺伝子型はaaA。)
講義動画【重複受精と胚乳の遺伝子型】
<以下、植物発生学について触れる。ここから先に進むのは、動物の発生を学んでからでもよい。>
要点:胚球は胚になる。胚柄は退化・消失する。
● 被子植物の発生
・受精卵は体細胞分裂を繰り返し、胚球(はいきゅう)と胚柄(はいへい)が生じる。
・胚球は胚になる。
・胚柄は退化・消失する。(非常によく問われる)

雑談:胚柄(はいへい)は通常、短命の構造である。胚柄は、完熟種子では、痕跡だけが残る。 胚柄は、胚を胚乳の中に深く押し込む。それによって、胚は栄養的に恵まれた環境に配置されることになるとされる。また、いくつかの植物で、胚珠以外の組織から栄養を吸収し、それを胚へ運搬していると考えられている。
要点:無胚乳種子は子葉に栄養を蓄える。
(1)無胚乳種子:栄養は子葉に蓄えられている。
例)アサガオ、クリ、ナズナ、アブラナ、マメ
語呂「朝からクリーナーで危ない豆拾い無理(アサガオ、クリ、ナズナ、アブラナ、マメ、無胚乳種子)」

雑談:クリの食べる部分は、子葉である。
*下図は無胚乳種子のイメージ。(( ))の中は雑談。

(2)有胚乳種子:栄養は胚乳に蓄えられている。
例)イネ、カキ
*白米の部分が胚乳である(イネは有胚乳種子)。
*下図は有胚乳種子のイメージ。

講義動画【被子植物の発生】
発展:胚乳の核相について
①被子植物の種子の胚乳:3n
②裸子植物の種子の胚乳:n
(裸子植物では、核相nの胚のう細胞が体細胞分裂を繰り返し、胚乳になる。したがって裸子植物の胚乳の核相はnである。)
雑談:「裸子植物では、核相nの胚のう細胞が体細胞分裂を繰り返し、胚乳になる。」と高校では教えることが多い。しかし、狭義の胚乳は「被子植物の」栄養組織を指すので、裸子植物に対して使用する用語としてはあまり適切ではない。
雑談:実は、真に胚乳をつくらない被子植物はラン科、カワゴケソウ科、ヒシ科など、限られたものだけである。無胚乳種子をつくる植物も、胚乳を形成する。しかし、発達中の胚によって胚乳が消費され、最終的に無胚乳種子となるのである(子葉が肥大して栄養を蓄えるものが多い)。胚乳が完熟種子まで持ちこたえているものを有胚乳種子と呼んでいるのである。
発展:イチョウの受精
(まずテストに出ない。ほぼ雑談と思ってよい。ただし、ソテツやイチョウが精子をつくることは知っておくこと。)
● イチョウやソテツは精子をつくる(他の裸子植物は精子をつくらず、精細胞を形成する)。
● イチョウを含め、裸子植物の配偶子形成、受精の過程は複雑である。だいたい以下のようなイメージを持っておけばよい。
● 花粉四分子は分裂を行い、花粉となり(図には示していない)、後に精子(n)を形成する(1個の花粉は4つの細胞[雄原細胞・花粉管細胞・2個の配偶体細胞]からなるが、このうち雄原細胞が受粉後に分裂して、2つの精子になる)。
● 胚のう細胞(n)は多細胞化し、卵細胞と、その周りの多数の細胞(やがて胚乳となる。したがって胚乳の核相はnである)を生じる(下図には卵細胞しか示していないが、卵細胞の周りには多数の細胞がある)。
● 花粉は花粉管を伸ばす。花粉管から放出された精子は、繊毛を使って水(粘液)の中を泳ぎ、自力で卵細胞まで移動する(胚珠では周辺組織から粘液が分泌されている)。
● 通常、2個の卵細胞のうち、1個だけが胚にまで成長する。
● 裸子植物では重複受精は起きない。裸子植物の胚乳となる部分は受精前につくられており、核相はnである(ただし、狭義の胚乳は「被子植物の」栄養組織を指すので、裸子植物に対して使用する用語としてはあまり適切ではない)。


雑談:イチョウやソテツでは、まず早朝に珠孔に分泌された受粉滴が、日の出とともに濃縮されて、表面についた花粉粒を胚珠の上部の花粉室内に引き込む。引き込まれた花粉は花粉管を伸ばし、養分を吸収しながら成熟し、2個の精子を完成させる。花粉粒から放出された精子は、造卵器室の液の中を泳いで、造卵器に到達する。受粉から受精までは数か月を要する。
雑談:イチョウの精子は植物学者の平瀬作五郎(ひらせさくごろう)が発見した。また、池野成一郎(いけのせいいちろう)はソテツの精子を発見した。
雑談:イチョウの精子を発見したのは、東京帝国大学(現在の東京大学)の植物学教室の助手であった平瀬作五郎である。平瀬は当初、それを寄生虫だと考えたが、当時助教授であった池野成一郎に見せたところ、池野はそれが精子であると直感したという。池野は平瀬のイチョウの精子発見の経過を終止応援していた(池野は平瀬の論文執筆を助けた)。その後、池野自身もソテツの精子を発見した。池野に帝国学士院賞恩賜賞授与の話があった時、池野は、平瀬の論文の方が先だったからと言って、平瀬がもらわないなら、自分ももらわないと主張した。
下図は平瀬作五郎が精子を発見したイチョウと、池野成一朗が研究に用いた鹿児島県に現存するソテツの株の分株。いずれも小石川植物園で撮影。


雑談:クロマツ(裸子植物)の受精
クロマツは雌雄同株である。花粉管中の精細胞が胚のう中の卵細胞と受精する(受粉した花粉は花粉管を伸ばし、精細胞を放出する)。精子は形成しない。受精に先立って卵の成熟と同時に胚乳が形成される(胚乳は胚のう細胞の分裂によって生じる)ため、胚乳の核相はnである。

雑談:ヒトの卵形成
● 卵形成は、その女性が誕生する前から始まっている。
● ヒト胚では、発生途上の卵巣に到達している数千個の卵原細胞が急速に分裂し、およそ700万個の卵原細胞を生み出す。
● 卵原細胞のほとんどは死んでしまうが、生き残った細胞は減数分裂に入り、一次卵母細胞となる。一次卵母細胞は、長い時間分裂を休止する(長い間、減数分裂第一分裂前期に留まる)。
そして、女性が成長し、思春期が始まると、また減数分裂を再開する。
● 第一分裂が完了すると第二分裂中期へと進行するが、この段階で二次卵母細胞が排卵される。そして、受精した場合に、また減数分裂を再開する。
下図はイメージ。知らなくてよい。

雑談:卵形成の過程で、減数分裂を開始した一次卵母細胞は、カエルで1~2年、ヒトでは十数年、減数分裂第一分裂前期で分裂を停止する(この時期をディプロテン期という)。その時、核は通常より大きくなっている。この大型の核を卵核胞という。卵核胞には、発生初期で働くmRNAを合成するランプブラシ染色体(多数の突起が生じていて、ランプを磨くブラシのように見える状態の染色体)や、リボソームを合成する多数の核小体が見られる。
発展:精巣と精子
● 脊椎動物の精巣に存在する、精子をつくるための曲がりくねった管状の構造を精細管という(ヒトでは、1日3000万個の精子が精細管でつくられている)。精細管においては、内腔に向かって、精母細胞、精細胞、精子が配列している。それらの細胞はセルトリ細胞という細胞に付着している(下図はイメージ)。セルトリ細胞には様々な役割がある。たとえば、セルトリ細胞は精細胞を覆い、保護し、栄養を与えている(その他、セルトリ細胞は、変性した生殖細胞の貪食や、精細胞の分化の制御などに関わる)。
*たまに発展問題のテーマとして扱われるが、知識問題としてはほぼ問われない。「セルトリ細胞は精細胞を守り、育てているんだなあ」というイメージを持っておけば良い。

雑談:精子形成の過程で、組換えが起こる。すると、(組換えによって遺伝的な情報が改変されるので)精細胞は非自己としての抗原性をもつことになる。何の対策もしないと、精細胞が非自己として認識され、自己免疫疾患が引き起こされる危険がある。セルトリ細胞は、精細管の内外をしっかり隔てることによって、そのような事態を防いでいると考えられている(また、同時に、セルトリ細胞は、有害な物質が精細管内に侵入し、精細胞のゲノムに影響を与えることを防いでいる)。
まだわかっていないこと
● 生殖細胞は、発生の過程でどのように体細胞から区別・分離されるのか。
● ウニの受精において、膜電位の変化が、なぜ第二の精子の進入を防ぎ、多精拒否にはたらくのか、完全には明らかになっていない。
● 脊椎動物の(始原生殖細胞はどうして生殖腺とは異なる場所で生じるのか。そして、)どのようなシグナルが始原生殖細胞を(発生途上の)生殖腺まで遊走させるのか、完全には解明されていない。
● 脊椎動物の配偶子成熟と排卵の複雑な過程の制御は、どのように行われているのか、完全には解明されていない。