
【高校生物】遺伝子②「リーディング鎖とラギング鎖の違いは何か?」

~プロローグ~
「実にその驚異の情こそ智を愛し求める者の情なのだからね。つまり求智の始まりはこれよりほかにはないのだ。」ソクラテス
半保存的複製には、5’→3’方向へのヌクレオチドの重合とともに、3’→5’ 方向へのヌクレオチドの重合が不可欠であると考えられていた。5’→3’方向に働くDNAポリメラーゼが発見された時、生物学者達は考えた。「3’→5’ 方向に働くDNAポリメラーゼもすぐに発見されるだろう。」しかし、3’→5’ 方向に働くDNAポリメラーゼは一向に見つからなかった。実際は、そのような酵素は地球上にはなかったのである。
DNAの複製は、図1ではなく、図2のように行われていた。

★テストに出やすいワード
①ラギング鎖
②岡崎フラグメント
③リーディング鎖
④密度勾配遠心法
⑤メセルソンとスタール
要点:ラギング鎖は、岡崎フラグメントというDNA断片をつなげることで合成される。
● DNAが複製されるとき、DNAポリメラーゼは5’→3’の方向にヌクレオチド鎖を伸長させる(鎖の3'末端にヌクレオチドを付加する)。
したがって、2本の鋳型鎖の一方では、もとのDNAがほどけていく方向と同じ向きに連続的に新生鎖が伸長する。これをリーディング鎖という。
● もう一方の鋳型鎖では、もとのDNAがほどけていく向きと、新生鎖が伸長できる向きが逆となる。そこではいくつもの岡崎フラグメントとよばれる短いDNA断片ができ、その断片がDNAリガーゼという酵素によってつながれながら、不連続的に新生鎖が伸長している。これをラギング鎖という。


*ラギング鎖は、リーディング鎖に比べて伸長が遅い、つまり伸長にタイムラグがあることからその名が付いた。その原因が「不連続な短いDNA断片をつなげているから」だと示したのが岡崎令治(おかざきれいじ)である。
雑談:岡崎令治は、恒子夫人らの協力を得て、1966年、生体内でのDNA合成前駆体である短鎖DNAの生成、伸長、結合を明らかにし、DNAの不連続的複製機構の解明に大きく貢献した。岡崎博士は中学時代に広島郊外で原爆に遭っており、慢性骨髄性白血病となり44歳で急逝された。
雑談:DNAポリメラーゼによる5'→3'方向のDNA合成と、巨視的に見た場合の片方の新鎖の3'→5'方向への伸長という矛盾(DNAジレンマ)は、不連続複製機構(岡崎フラグメントごとの不連続な複製)の発見によって解決された。
雑談:岡崎フラグメントは、原核生物では1000~2000ヌクレオチド、真核生物では100~200ヌクレオチドの長さをもつ。
雑談:複製起点周辺の、DNAが開いた構造を複製バブルと言う。
● (複製バブルの両側の)Y字状の構造を複製フォークと言う。

講義動画【半保存的複製】
雑談:高校ではあまり注目しないが、一般に、ポリメラーゼが働くためには、必ず「鋳型の鎖」を必要とする。たとえば、DNAポリメラーゼは、鋳型無しで、バラバラのヌクレオチドから自由な配列をもつDNAを作り出すことはできない(ポリメラーゼを画家にたとえれば、その画家は、もともと存在する絵と似た作品は描けても、0からの真の創作はできないのである)。
<Q.岡崎フラグメント=ラギング鎖?…正確には違う。もともと「フラグメント」は「断片」を表す語である。岡崎フラグメントは、ラギング鎖が伸長する際につくられる、「短いDNA断片」を指す。岡崎フラグメントがいくつもつながって、ラギング鎖が伸びていくイメージ。ラギング鎖が発見された時は、どのようにしてラギング鎖が伸びるのかわからなかった(少しリーディング鎖に遅れて[タイムラグがある]合成されることはわかっていた)。やがて、岡崎フラグメント同士がDNAリガーゼで連結されることにより伸びていることが分かった。>
問題:以下の図はDNAの複製の様子を模式的に表したものである(新鎖は描いていない)。プライマーが設置される可能性があるのは①と②のどちらか。

答え:①
解説:このような問題は、鋳型鎖の3'や5'に注意する。
まず、図を半分に分ける。

鋳型鎖の5'末端に向かっていく鎖がリーディング鎖である(鎖が伸びる末端[矢印の先端]は必ず3'である。そして、新しい鎖の3'-5'の方向性は、鋳型鎖と逆になっている)。
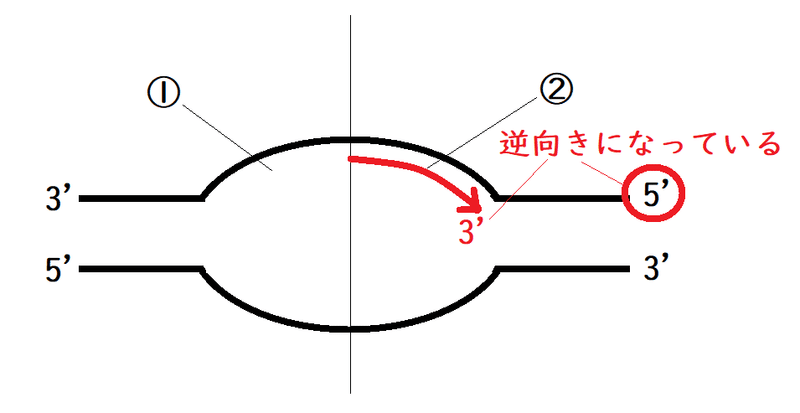
上の図より、②にプライマーは設置されない(②にはリーディング鎖がある)。
一方、鋳型鎖の3'末端方向には、ラギング鎖が形成されていく(ラギング鎖では、何個もプライマーが設置され、岡崎フラグメントが形成されていく)。

よって正解は①である。
下半分も描けば、以下のような図になる。鋳型鎖(黒)と新鎖(赤)の3'---5'の方向性が逆向きになっていることと、新鎖(赤)の3'末端側が伸長していることに注意。自分で作図できるように。

挿絵:ドライフラワーのような複製フォーク

要点:メセルソンとスタールは、塩化セシウムを使った密勾配遠心法によってDNAの半保存的複製を証明した。
● 半保存的複製では、DNA二本鎖のそれぞれの一本鎖が鋳型となり、それぞれに相補的な新しい一本鎖が合成されて、もとと同じ二本鎖DNAが合成される。
● DNAの二重らせん構造が明らかになったとき、DNAの複製の方式について、(a)~(c)の3つの可能性が考えられた。
(a)半保存的複製:DNA二本鎖のそれぞれの一本鎖が鋳型となり、それぞれに相補的な新しい一本鎖が合成されて、もとと同じ二本鎖DNAが合成される。(正しい仮説。DNAは半保存的複製によって複製される。)

(b) 保存的複製:古い鎖は分離することなく完全に保存される。(誤った仮説)

(c) 分散的複製:古い鎖と新しい鎖がランダムに混ざり合う。(誤った仮説)

これらの可能性のどれが正しいかを明らかにするために、メセルソンとスタールは密度勾配遠心法(みつどこうばいえんしんほう)を行った(密度勾配遠心法は、密度勾配遠心分離法ともいう)。

雑談:この実験は生物学史上最も美しい実験と言われる。簡単な手法で、見事に保存的複製と分散的複製を否定し、半保存的複製を証明したからである。
(1)実験
大腸菌を、重い窒素(15N)を含むNH4Clの存在下で培養し、細胞内DNAを15Nで標識した。
その後、軽い窒素(14N)を含むNH4Clを含む培地に移して培養し、1回複製後、および2回複製後に大腸菌からDNAを精製し、密度勾配遠心法により解析した(大腸菌が1回分裂したタイミング、2回分裂したタイミングでDNAを取り出し、密度勾配遠心分離にかけた)。

*密度勾配遠心法:密度勾配遠心分離法ともいう。何種類か方法があるが、たとえば、塩化セシウム(CsCl)などの密度勾配を遠心によって形成させ、生体分子を固有の(その分子の密度に応じた)位置に移動させる方法などがある(詳しい原理は問われないが、「塩化セシウム」を使用するということは問われる)。
雑談:密度勾配遠心法の詳しい原理
塩化セシウムは、長時間遠心すると、液面から底に向かって密度が増す勾配をつくる。この密度勾配に様々な密度をもつ物質(たとえば様々な密度をもつDNA)を加えておくと、その密度に応じて勾配の中の異なる位置に沈降する。加えた物質の密度が溶媒の密度より大きい部分では沈降が、その逆の部分では浮上が起こり、最終的には両者がつり合った位置に加えた物質がバンド状に集まることになる(たとえば、密度が均一な水の中に物質をポチャンと入れた場合は、その物質の密度が、水の密度より小さいと液面まで浮かび上がり、水の密度より大きいと底まで沈む。これでは、様々な密度を持つ物質を分離することはできない[水の密度より大きいか小さいかでしか分けることができない]。塩化セシウムを用いた密度勾配遠心法では、塩化セシウムが密度の勾配を容器の中で作っている[水のように液面から底まで密度が均一ではなく、液面に近い方が密度が小さく、底に行くほど密度が大きくなっている]。そのような溶媒の中では、物質は、自身の密度に応じた位置に留まる[密度が大きいものほど底に近い位置に留まる])。

講義動画【細胞分画法と密度勾配遠心法】
雑談:密度勾配遠心法には、ショ糖を用いて行う方法もある。それを、ショ糖密度勾配遠心法という(対して、上で説明した塩化セシウムを使う密度勾配遠心法は、平衡密度勾配遠心法という)。 まず、遠心管内に、あらかじめショ糖の密度勾配を作製しておく。その上に分離したい試料を加えて遠心を行うと、沈降速度(下に落ちていく速度)の差によって、試料を成分ごとに分離することができる(試料中の粒子は、おのおのの沈降速度に従って密度勾配中をバンドになって移動する。さらに、この方法では、遠心終了後、遠心管の底に穴をあけて、試料を下から取り出し、一定量ずつ採取して、分析することもできる)。
(2)結果
*以下、15Nを含む鎖(重い鎖)のみからなるDNA(15N-DNA)を重いDNA、
15Nを含む鎖(重い鎖)と14Nを含む鎖(軽い鎖)からなるDNAを中間の重さのDNA、
14Nを含む鎖(軽い鎖)のみからなるDNAを軽いDNAと呼ぶ。
はじめ(軽い窒素の培地に切り替えた直後)、すべてのDNAは重いDNAであった。


1回目の複製後、中間の重さのDNAのバンド(重い鎖+軽い鎖)が見られた(保存的複製の否定)。
*もし保存的複製が正しいなら、重いDNAのバンドがずっと残っているはずである。


2回目の複製後、中間の重さのDNAと、軽い重さのDNAのバンド(軽い鎖+軽い鎖)が見られた(分散的複製の否定)。
*もし分散的複製が正しいなら、何度複製を繰り返しても、軽い重さのDNAは生じないはずである(醤油とオレンジジュースを加えて割っていっても、完全なオレンジジュースは生じないように)。


以上の結果から、DNAは半保存的複製によって複製されることが証明された(DNAが半保存的複製によって複製されているとすれば、以上の結果を合理的に説明できる)。
講義動画【半保存的複製の証明】
発展:テロメア
染色体の末端をテロメアという。テロメア配列は、特徴的な短い塩基配列の繰り返しで構成されている(たとえば哺乳類の場合は5'ーTTAGGCー3'が数百回)。染色体の末端では、プライマーが除去された後、その空白を埋めることができない(他の場所では、その空白を埋めることができる。まず、プライマーRNAを除去してDNAに置き換える[まずリボヌクレアーゼHという酵素によりRNAプライマーが取り除かれ、できた空白をDNAポリメラーゼが埋める]。空白の部分を、後ろから伸びてくるDNAが埋めるイメージ。その後、DNAリガーゼが2つの断片の切れ目をつなぐ。図1)。
したがって細胞分裂を繰り返すごとに、テロメアは短くなって行く(図2。これが有名な末端複製問題である。原核生物はこの問題をDNAを環状にすることで回避している)。これが、細胞分裂の回数に一定の限度があること、すなわち細胞寿命と関係しているとされる。
テロメラーゼというテロメアを伸ばす酵素が存在する。テロメラーゼは造血幹細胞など、細胞分裂を繰り返す必要のある細胞でよく発現していることが知られている。
図1
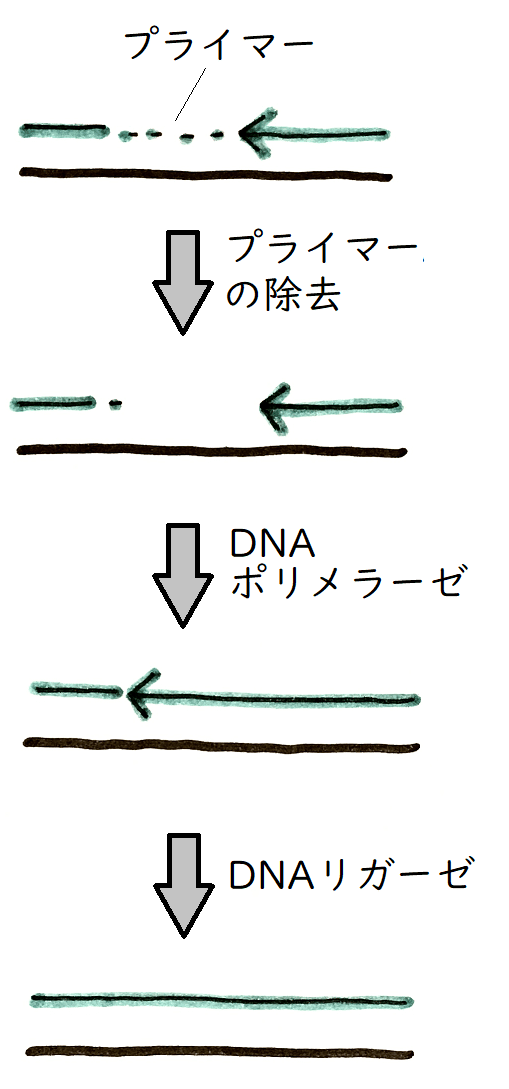
図2

雑談:「死が起きるのは、磨り減った組織がいつまでも再生できるわけではないからであり、そして、細胞分裂という手段で増える能力は無限ではなく、限界があるからだ。」ワイズマン(ヴァイスマン、ワイスマンとも。ドイツの動物学者で、生殖質説の提唱者。)
まだわかっていないこと
● ヒトの寿命はどのように決まるのか。細胞の寿命はどのように決まるのか。それにはテロメアが関係しているのだろうか。
落語『死神』では、男が、死神に、「寿命のロウソク」が消えれば死ぬと宣告される。「震えると消えるよ。消えると死ぬよ。」「あぁ〜もう消えそうだ・・・消えてしまう・・・」「あぁ・・・消えた。」
「寿命のロウソク」は、DNAのテロメアととてもよく似ている。
クローン羊ドリーはなぜはやくに死んでしまったのか。テロメアが関係しているのか。それとも一般的な疾患が原因だったのか。
● DNA複製の校正について、完全には解明されていない。DNAの複製は、素早く、かつ、驚くべき正確さで実行される。
付加する塩基10000000000個あたり、誤りは1個程度に維持されている。1000冊の本を高速で書き写す時に、1文字の書き間違いしか起こさないようなものである(DNAポリメラーゼも、低い頻度だが、間違いは起こす。しかし、たとえば、誤ったヌクレオチドは、DNAポリメラーゼと同じポリペプチドに存在する校正エキソヌクレアーゼにより除去され、正される)。
なお、RNAポリメラーゼの行う転写では、もっと間違えが起こりやすい。RNAは分解されてしまうので、それほど慎重になる必要はないのかもしれない。