
【高校生物】遺伝子③「遺伝子はどのように発現するのか?」

~プロローグ~
「生物体のすべての構造と働きがこれを構成しているタンパク質の構造と活性の結果として生ずるものであるかぎり、生物全体を遺伝的メッセージそのものの最終的な後成的表現であるとみなさなくてはならない。」モノー『偶然と必然』より
★テストに出やすいワード
①転写
②RNAポリメラーゼ
③翻訳
④リボソーム
⑤セントラルドグマ
(生物基礎の復習)
講義動画【染色体・DNA・遺伝子・ゲノム】
要点:遺伝情報の流れは一方向的である。
● DNAからRNAへ、RNAからタンパク質へ、『一方向的に』遺伝情報が流れるという原則を、セントラルドグマという。

*以下はセントラルドグマのイメージ。DNAは、プラモデルの作り方が書かれた説明書の原本、RNAがそのコピー、タンパク質がプラモデル。

発展:逆転写
セントラルドグマの例外として、逆転写がある。特殊なウイルスがもつ逆転写酵素(RNA依存DNAポリメラーゼ)という酵素はRNAを鋳型にしてDNAを合成する。RNAを遺伝物質としてもつある種のウイルスは、逆転写酵素により自身のRNAを鋳型にしてDNAをつくり、宿主のDNAに組み込む(レトロウイルスというウイルスは、感染細胞において自身のRNAから二本鎖DNAを合成する。この反応はウイルス粒子内にある逆転転写酵素によって行われる。逆転写酵素によってつくられたDNAは、感染細胞のDNAに組み込まれる。そして、感染細胞のRNAポリメラーゼによってmRNAが合成され、それをもとにウイルスタンパク質が翻訳される。なお、HIVはレトロウイルスの一種である)。
雑談:セントラルドグマの提唱者、クリックは、すでにセントラルドグマの例外を予言していた。彼はRNAの塩基配列情報からDNAの塩基配列情報へ情報が流れ得ると考えた。実際、その予言は的中した。RNAを鋳型にDNAを合成する特殊な酵素、「逆転写酵素」が発見されている。また、「RNA依存性RNAポリメラーゼ(RNAを鋳型としてRNAを合成する酵素)」も発見されている。なお、「一度タンパク質に入った情報はもう外へ出ることはない」というクリックの主張については、例外は見つかっていない。タンパク質のアミノ酸配列情報に翻訳された情報は、もうタンパク質から出ていかない。アミノ酸配列情報が塩基配列情報に戻る『逆翻訳(こんな用語はない)』は地球上に存在しない。以下は拡張されたセントラルドグマのイメージ。

要点:DNAにはタンパク質の情報をもつ領域(ヒトゲノムでは全体の1.5%程度)と、持たない領域がある。
● 現在、遺伝子に決まった定義はない。高校生は、まずは「遺伝子とは、転写され得るDNA領域である」と考えておけばよい(タンパク質のアミノ酸配列を指定する領域のみを遺伝子ととらえることもある。遺伝子発現を調節している領域も遺伝子に含めることもある)。
● タンパク質情報を指定する領域はヒトゲノムの1.5%程度である。遺伝子領域は染色体上に「点在」している。
● 細胞の種類ごとに、また、時期ごとに発現している遺伝子は異なる(選択的遺伝子発現)。下表はイメージ。

雑談:以下の写真は僕が大学生時代に撮ったもの。アフリカツメガエルの胚の、nrp-1という遺伝子が発現している細胞を染めている。st.(ステージ)13~15というのは発生の段階を示している。nrp-1という遺伝子は、st.14の時期から発現し始めていることがわかる。また、特定の細胞だけで発現している[胚の一部だけが染まっている]こともわかる(ちなみに、nrp-1は神経をつくる細胞[神経幹細胞]で発現する遺伝子なので、神経が形成される領域で発現している)。

● ほぼすべての体細胞は同じ遺伝子のセットを持つ(例外はT細胞とB細胞。これらのリンパ球は遺伝子の再編成を行っているため、持つ遺伝子が異なる)が、発現している遺伝子は細胞ごとに異なる。つまり、たとえるなら、遺伝子にはスイッチON、OFFの状態があり、細胞ごとに、ON(発現する)、OFF(発現しない)が調整されているのである。
● どの細胞でも常に発現し、細胞の生命活動に必須な機能を果たすタンパク質をコードする遺伝子を総称してハウスキーピング遺伝子という(上表の遺伝子Cのような遺伝子がハウスキーピング遺伝子。どの細胞でも常に発現している)。
*ハウスキーピング遺伝子には、たとえばrRNAの遺伝子などがある(基本的にすべての細胞にリボソームは必要である)。
要点:mRNAにはタンパク質の情報が含まれ、tRNAはアミノ酸を運び、rRNAはリボソームの構成要素となる。
● 連続する塩基3個の配列をトリプレットとよぶ。
● RNAには以下のような種類がある。
①mRNA(伝令RNA)はタンパク質のアミノ酸配列についての情報を含む。
*mRNAのトリプレットはアミノ酸を指定しており、コドンと呼ばれる(ただし終止コドンはアミノ酸を指定しない)。
(コドン=トリプレットとすることもある。)
*コドン:mRNA(またはDNA)のトリプレット(3塩基)で、特定のアミノ酸を指定する遺伝暗号の単位。ただし、終止コドンはアミノ酸を指定せず、タンパク質の合成の終止を指示している。4×4×4=64種のコドンがある。
*塩基は4種類(ATCG)ある。塩基2つの並びでは4×4=16種類のアミノ酸しか指定できない(タンパク質を構成するアミノ酸は20種)ので、コドンは3文字ではないかと予想されていた(知らなくてよいが、ビッグバン理論で有名なガモフによって予測されていた)。
②tRNA(転移RNA)はアンチコドン(mRNAのコドンに対応するトリプレット)をもち、特定のアミノ酸をリボソームまで運ぶ。

*tRNAとアミノ酸の結合反応を触媒する酵素を、アミノアシルtRNA合成酵素という(アミノ酸と結合したtRNAをアミノアシルtRNAという)。

③rRNA(リボソームRNA)はリボソームの成分である(リボソームはrRNAとタンパク質からなり、膜で包まれていない。rRNAの部分に主な触媒活性があるらしいことがわかっている。すなわち、rRNAはリボザイム[酵素活性をもつRNA]である)。
講義動画【遺伝暗号表】
要点:転写では(RNAポリメラーゼにより)mRNAが合成され、翻訳では(リボソームにより)タンパク質が合成される。
● DNAの一方の鎖が鋳型となって、その塩基配列に対応したRNAがRNAポリメラーゼによって合成される。これを転写という。
(1)転写の流れ
(DNAの塩基配列をRNAの塩基配列に『転写』する)
①真核生物では、RNAポリメラーゼが基本転写因子とともにプロモーターに結合して転写が開始される(よくテストに出る)。
*DNAには、プロモーターとよばれる領域がある(プロモーターは転写をここから開始せよ、という合図になっている。転写の速度[DNAとRNAポリメラーゼの親和性]はプロモーターの種類で決まる)。

雑談:真核細胞の転写因子のうち、転写反応に必須なものを基本転写因子という(原核細胞ではほぼRNAポリメラーゼのみで転写が起こり得るが、真核細胞ではRNAポリメラーゼに加えていくつかの基本転写因子が正確な転写開始点の認識及び転写に不可欠である)。
RNAポリメラーゼの種類によって異なる基本転写因子が関与する(高校では習わないが、RNAポリメラーゼにはいくつか種類がある)。
基本転写因子の働きには「TATAボックスというプロモーターに共通して見られる配列を認識する」「RNAポリメラーゼを転写開始部位に正しく配置する」「転写開始部位のDNAをほどく」などがある。
基本転写因子とRNAポリメラーゼは、プロモーターの上で会合し、複合体を形成する。
また、多くの場合、介在因子(メディエーター)と呼ばれる大型のタンパク質複合体が、転写活性化因子とRNAポリメラーゼ・基本転写因子との適切な連携に役立っている(転写活性化因子は、DNAの特定の配列に結合し、転写を活性化させる因子。下図には1個しか描いていないが、ふつう複数の転写活性化因子が関わり、転写の速度やパターンを決定している。さらに、時には数千塩基離れた場所から作用して、RNAポリメラーゼ・介在因子・基本転写因子がプロモーターに集合するのを助ける)。
以上のように、たくさんのタンパク質同士の相互作用により、転写開始複合体(プロモーターを含むDNA上でRNAポリメラーゼと基本転写因子群が集合して形成される複合体)の配置と組み立てが制御されている(RNAポリメラーゼと基本転写因子が転写開始複合体を形成することで、転写開始反応が起こると考えられている)。

②RNAポリメラーゼが働き、鋳型鎖(アンチセンス鎖)と相補的な配列を持つRNAが合成される。
*2本鎖DNAのどちらの鎖が鋳型鎖になるかは、遺伝子によって異なる。
*A→U(ウラシル)、G→C、C→G、T→Aのように、鋳型鎖と相補的な配列を持つRNAが合成される。
③RNAポリメラーゼはRNAを合成する(RNAの3’方向が伸長していく)。
下図は転写のイメージ。

講義動画【転写】
雑談:上の動画に出てくるDNAヘリカーゼ(DNAアンワインディング酵素)という用語は発展。DNAヘリカーゼは、DNAの複製や、転写の過程で使用される、DNAを巻き戻す酵素である。真核生物の転写では、TFⅡHという基本転写因子がサブユニットの1つとして持っているDNAヘリカーゼによってDNAを開裂させる。TFⅡHのヘリカーゼ活性によりDNAの二本鎖が巻き戻され、RNAポリメラーゼがRNAを合成していく。下図はイメージ。
*原核生物においても、転写の大きな流れは変わらない(細かい点はかなり異なる。大学で学んでほしい)。DNAは部分的に開裂し、RNAポリメラーゼによってmRNAが合成されていく。

雑談:DNAポリメラーゼとは異なり、RNAポリメラーゼはプライマーを必要としない。
● 核内で合成されたmRNAは、核膜孔(かくまくこう)から細胞質へ移動する。核の外でmRNAはリボソームと結合する。リボソームはmRNAの塩基配列に基づいてタンパク質を合成する。



(2)翻訳の流れ
(mRNAの塩基配列情報をタンパク質のアミノ酸配列情報に『翻訳』する)
*スプライシングによって、イントロン部分が除去された(エキソン部分が連結された)mRNAのみが核の外へ出ることができる。スプライシングについては後述する。
① mRNAにリボソーム(タンパク質合成の場)が付着する。
*AUGが開始コドンである。開始コドンとなるAUGからアミノ酸の連結が開始される。
② mRNAのコドンに対応するアミノ酸と結合したtRNAが、アンチコドンの部分でmRNAのコドンと結合する。
③tRNAによって運ばれてきたアミノ酸が、伸長中のポリペプチドとペプチド結合する。
*ポリペプチド鎖のC末端が、tRNAに結合したアミノ酸のアミノ基とペプチド結合をつくる(合成されているポリペプチドは、リボソームから遠い方がN末端になっている)。


② mRNAのコドンに対応するアミノ酸と結合したtRNAが、アンチコドンの部分でmRNAのコドンと結合する。
③運ばれてきたアミノ酸が、伸長中のポリペプチドとペプチド結合する。
*リボソームがmRNAの上を5’→3’方向(語呂「ゴミ方向」)に移動しながら②、③の過程が繰り返され、特定のタンパク質が合成されていく。
⓸リボソームが終止コドンのところまで来たら翻訳終了(リボソームはmRNAから離れる)。
*終止コドンが翻訳終了を指示している。
● 開始コドン(AUG[メチオニンを指定])はリボソームに向けた「アミノ酸の結合を開始せよ」という合図である。
語呂「恋は8月(AUGust)に始まる(AUG、開始コドン)」
● 終止コドン(UAA、UAG、UGA)はリボソームに向けた「アミノ酸の結合を終了せよ」という合図である。
語呂「うああ、うあぐ、うがあ(UAA、UAG、UGA)」
雑談:開始コドンの前、および終止コドンの後ろも転写される。つまり、mRNAには、翻訳される部分の前後に、翻訳されない、余白みたいなところ(非翻訳領域)がある。
雑談:「暗号は翻訳されないかぎり意味を持たない。」モノー(オペロン説の提唱者)
講義動画【翻訳】
雑談:tRNAは、自身の中で相補的な塩基同士が結合し、クローバーの葉のような形になっている(実際はさらに折りたたまれて複雑な形状になっている)。アミノ酸はtRNAの3'末端に結合する。下図はイメージ。水色の線は塩基対の形成を表す。

挿絵:トランペットのようなtRNA

要点:転写と翻訳には方向性がある。
● RNAポリメラーゼは、mRNAの3'末端を伸ばしながらDNA上を移動する(DNA鎖、RNA鎖に関わらず、すべてのヌクレオチド鎖は3'末端しか伸ばせない)。
● リボソームはmRNA上を3'方向に向かって移動する。
語呂「ゴミ方向(5'→3'方向にリボソームは進行)」
● リボソームからは合成途中のポリペプチドが伸びるが、リボソームから遠い方がポリペプチドのN末端になっている。
講義動画【転写と翻訳の方向性】
<Q.tRNAってどこからやって来るの?…tRNA遺伝子から転写されることでつくられる。細胞内に無数に存在する。>
<Q.センス鎖、アンチセンス鎖って何?…転写を考えたとき、非鋳型鎖をセンス鎖、鋳型鎖をアンチセンス鎖という。センスというのは「意味を持つ」といった意味。どうして非鋳型鎖をセンス鎖というのかというと、非鋳型鎖のほうがRNAの配列に似ているからである。下図を見よ。TをUに変えれば、RNAと同じ配列になっている。遺伝子配列を他人に示すときは、非鋳型鎖(センス鎖)を示すのが普通。問題文でも、何の説明もなく片方の鎖の遺伝子の配列が書かれていたら、それは非鋳型鎖(センス鎖)の配列と思ってよい。>

雑談:Amanita phalloides(タマゴテングダケ)のつくるアマニチンというペプチドは、RNAポリメラーゼと特異的に結合し、『転写』を阻害する。アマニチンの毒性は50%の致死率を持つ。また、ヒマの種からとれるリシン(アミノ酸のリシンとは別の物質)は、『翻訳』、つまりタンパク質合成を阻害する(冷戦中、リシンが暗殺に使われたこともあった)。これらの事実は、生命の営みが『転写と翻訳』に基づいていることを意味する。
発展:遺伝暗号の解読
人工RNAを合成することで、遺伝暗号表の解読が行われた。
講義動画【遺伝暗号の解読】
*オチョア、ニーレンバーグ、コラーナ(昔は名前が問われたが、最近はほぼ問われない。上の動画の原理を理解しておけばよい)によって遺伝暗号が解読され、遺伝暗号表(コドン表)が作成された。
雑談:ヌクレオチドポリマーを合成する研究がオチョアによって行われ、ポリヌクレオチド・ホスホリラーゼが発見された。この酵素を用いることで、人工mRNAの合成が可能になった。ニーレンバーグは、人工ポリウラシル(5'…UUUUUUUUU…3')mRNAを合成し、大腸菌由来の無細胞翻訳系に与えた。さらに、コラーナ(コラナ)は、2個の塩基(UC)が繰り返し並ぶポリUCmRNAや、4個の塩基(UAUC)が並ぶポリUAUCmRNAを合成し、遺伝暗号表の解読を行った(コラーナは、ウィスコンシン大学で、ヌクレオチドを2個、3個とつなげる化学反応を考案した)。
雑談:ニーレンバーグが歴史的な実験を行った時、まだ開始コドンAUGは発見されていなかった。しかし、幸運なことに、ニーレンバーグが用いた無細胞タンパク質合成系には高濃度のマグネシウムが含まれていたので、タンパク質合成が強制的に開始されたのだった。
雑談:ニーレンバーグは、人工ポリウラシル(5'…UUUUUUUUU…3')mRNAのRNAを用いて、UUUがフェニルアラニンをコードする暗号であることを発見した。そして、残りの暗号解読に向けた研究者同士の競争が始まった。コラーナは、RNAを人工合成することで遺伝暗号の解読を行った。不思議なことに、コラーナにより合成された反復配列をもったRNAの中には、ポリペプチドを長く合成できないものが含まれていた。ポリペプチド合成を止める配列が存在していたのである。これを今では終止コドンという。
雑談:三種類の終止コドンは、オーカーコドン(UAA)、アンバーコドン(UAG)、オパールコドン(UGA)と呼ばれている。発見者の1人の姓がバーンスタインで(ドイツ語で琥珀 amber を意味する)、オーカーとオパールは、アンバーと同様に準宝石に由来する。

*Phe(フェニルアラニン)、Leu(ロイシン)、Ile(イソロイシン)、Met(メチオニン)、Val(バリン)、Ser(セリン)、Pro(プロリン)、Thr(トレオニン)、Ala(アラニン)、Try(チロシン)、His(ヒスチジン)、Gln(グルタミン)、Asn(アスパラギン)、Lys(リシン)、Asp(アスパラギン酸)、Glu(グルタミン酸)、Cys(システイン)、Trp(トリプトファン)、Arg(アルギニン)、Gly(グリシン)はアミノ酸。
雑談:「人間からバクテリアまで、すべての生体は、いかなる遺伝のメッセージであろうと正確に解釈することができるようにみえる。遺伝の暗号表は普遍的であって、それを解く鍵は生物界すべてに知られているようである。」ジャコブ『生命の論理』より
要点:真核生物の核内では、スプライシングが起こる。
● 真核生物ではスプライシングが起こり、mRNA前駆体(単にRNAと記されることもある)が成熟mRNA(あるいは単にmRNAとよばれる)になる(スプライシングを議論しない場合は、転写産物をそのままmRNAと記すことも多い)。
● 真核生物では転写されたRNAはまだ未熟である。真核生物では、核内でイントロン部分が取り除かれ、エキソンの配列がつなぎ合わされる。これをスプライシングという。基本的にスプライシングは真核生物のみでみられる。

● エキソン領域のみが核膜孔から細胞質へ出ていく。
雑談:核の「外」に出る領域だから「エキソ」ンと言う。エキソは外を表す。エキソサイトーシスも同じ語源。
雑談:核膜の機能の一つは、スプライシングの場を確保することである。
● スプライシングのとき、同じ遺伝子から転写されたRNAでも、異なるエキソンがつなぎ合わされて、異なるmRNAがつくられることがある。これを選択的スプライシングといい、この仕組みにより、1つの遺伝子から複数のタンパク質を合成することができる(選択的スプライシングの利点としてよく問われる)。
*人の遺伝子は20000程度しかないのにもかかわらず、遺伝子数以上のタンパク質が合成されているのは、選択的スプライシングが原因の一つであると考えられている。
発展:真核生物では、転写によって生じたRNAの5'にキャップ構造が、3'にポリAが付加される。
● 真核生物の行うRNAの加工には、スプライシングの他にも、5’末端へのキャップ構造の付加、3’末端へのポリAテールの付加などが知られている。
①キャップ構造の付加(キャップ構造はリボソームを呼び寄せるらしい)、②スプライシング、③ポリAテールの付加(ポリAテールはRNAの寿命にかかわるらしい)は、RNAプロセシングと呼ばれている。
下図はイメージ。

雑談:真核生物の多くのmRNAの5' 末端にある修飾構造をキャップという。 必ずしも構造上の画一性はないが、5' 末端は必ず7-メチルグアノシンであり、その5' 部位と次のヌクレオシドの5' 部位が、三リン酸を介して結合している。転写開始直後(約25ヌクレオチドRNAが合成されると)すぐにキャップが付加される。
雑談:真核生物のmRNAの3' 末端に普遍的に存在するアデニル酸の連続した配列をポリAテール、もしくはポリA配列という。 その長さは数十から200ヌクレオチド程度であり、一つのmRNA分子種についても長さは一定でない。 ポリAテールは遺伝子によってコードされておらず、後から付加される(下図はイメージ)。一般に、mRNA前駆体中には、ポリAが付加される10~30ヌクレオチド上流に、AAUAAAまたは類似の配列(ポリA配列付加シグナル)がある。この配列が認識されると、mRNA前駆体が切断され、できた3' 末端にポリAが付加されていく(下図では省略しているが、様々なタンパク質が関わる)。この切断およびポリA付加反応(ポリアデニル化)は、核内で起こる。ポリA配列の機能は確定的でないが、mRNAの安定化、翻訳、核から細胞質への輸送、mRNAスプライシングの際の3'末端にあるエキソンのシグナルとしての役割などが示唆されている。

雑談:RNAポリメラーゼが遺伝子の端に来ると、そこに特別な塩基配列が待ち受けている。その配列の転写でRNAが出来ることがきっかけとなって、様々な酵素群がそこに移動し、ポリAの付加や転写の終結などの反応が引き起こされる。
雑談:スプライシングは転写と組み合わさって起きる。多くの場合、スプライシングに関する反応は、mRNA前駆体の合成途中で開始されることが知られている(ただし、エキソンとイントロンの境界の目印付けは転写と同時に行われるが、実際のイントロン除去は後で起こる)。
雑談:RNA内の塩基配列がスプライシングの起こる場所を決めている(イントロンとエキソンの境界にmRNA前駆体中でスプライシングが起きるべき正確な位置を示す塩基配列が存在する)。
雑談:選択的スプライシングは活性化因子と抑制因子(いずれもタンパク質)によって調節されている。これらのタンパク質がRNAに結合することで、特定の位置でのスプライシングが活性化されたり抑制されたりする。
問題:以下のような遺伝子から、選択的スプライシングによって多様なmRNAがつくられるとする。ただし、エキソン1は必ずmRNAに残るものとする。理論上、選択的スプライシングによって何種類のmRNAが生じ得るか。

答え:4種類。
各エキソンが、選ばれるか、選ばれないかの2択なので、2×2=4。
(エキソン1のみ、エキソン1と2、エキソン1と3、エキソン1と2と3)
エキソン2と3の順番は逆転しないことに注意。
下図はイントロンが除去される時のイメージ。
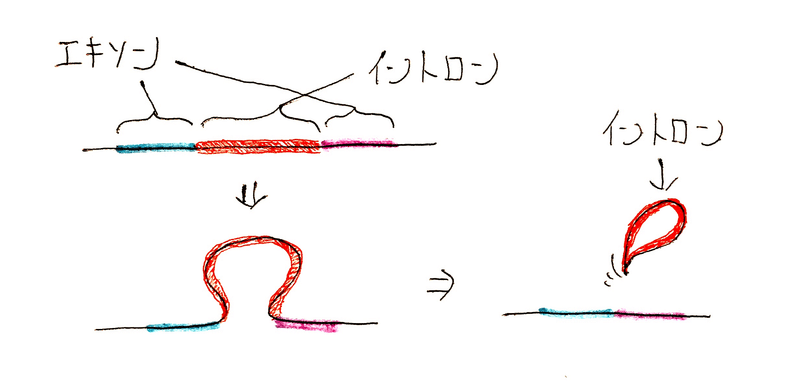
発展:スプライシング
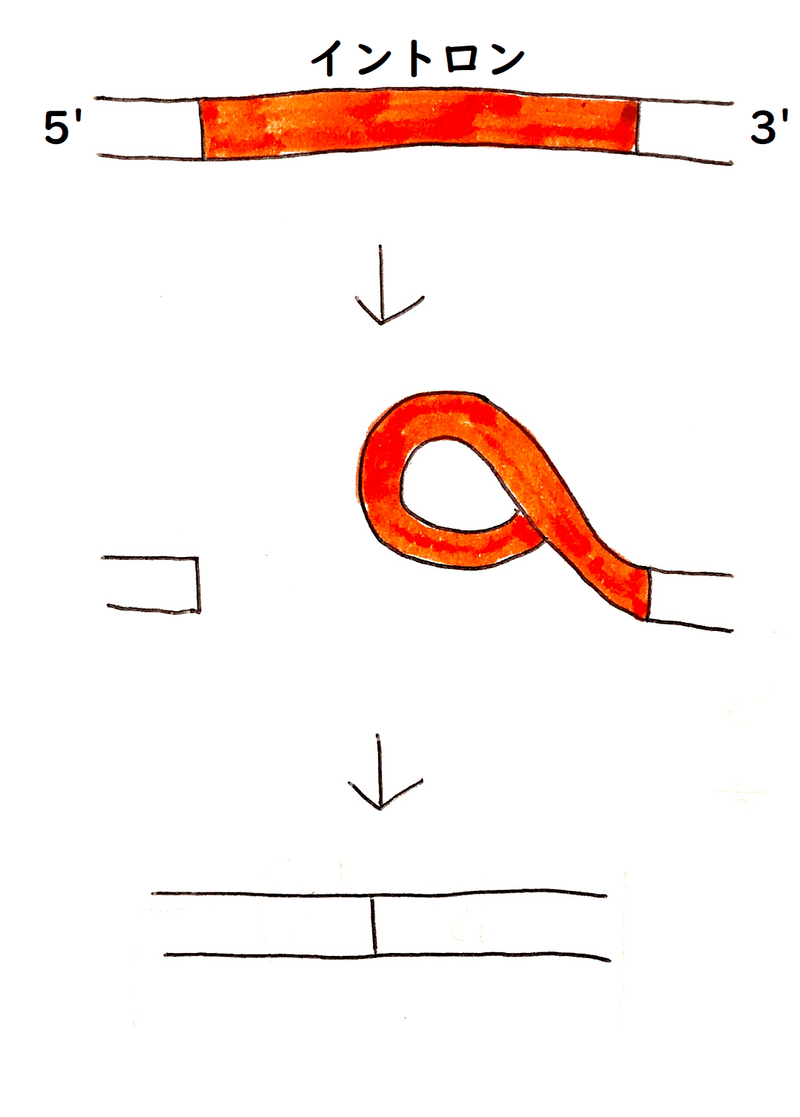
鋳型となったDNA鎖と、スプライシングを受けて完成したmRNAを実験的に結合させると、DNAのイントロン領域がループ状に突き出るのが観察される(DNAの鋳型鎖の配列と、mRNAの配列は、相補的になっているので、実験的に結合させることができる)。
下図はイメージ。
①転写で生じたmRNA前駆体は、スプライシングによってイントロンが除かれ、エキソン同士が連結し、成熟したmRNAになる。(DNAの非鋳型鎖は描いていない。)

②人工的に1本鎖にしたDNAの鋳型鎖と、成熟したmRNAを結合させるとする。DNAには、mRNAにはないイントロン領域の部分があるので、その部分が突き出る。

このような実験により、(追い出された)ループ状のDNA鎖が観察されたことは、スプライシングが起こっている根拠のひとつになった。
*上図では、ポリAテールは省略している。実際は、mRNAの3'末端では、ポリAテールが"しっぽ"のように突き出ている(ポリAテールはmRNAに後から付加される鎖なので、ポリAテールと相補的な配列は鋳型のDNA鎖に存在しない)。
雑談:イントロンの除去は、せっかくつくったRNAを捨てるということであり、資源の無駄になる。また、イントロン領域がある分、DNA合成が面倒になる。もともと全ての生物がイントロンをもっていたが、原核生物は、イントロンという無駄を省き、素早く増殖する方に進化の戦略の舵を切ったのだという説がある。また、もしかしたら、イントロンは、寄生性の動く遺伝因子が由来なのかもしれない。イントロンの由来については、結論が出ていない。
雑談:以下は少しだけ正確なスプライシングのイメージ(図には描いていないが、切り出されたイントロンは、拡散していき、分解される運命にある)。
まだわかっていないこと
● tRNAはどのように適切なアミノ酸をリボソームまで運搬するのか、その仕組みについては完全には解明されていない。
● 生物種ごとに特有なコドンの使用の偏りがあるが、その理由については完全には明らかになっていない。
● ミトコンドリアゲノムや、少数の生物種で(標準のコドン表に従っていない)変則的なコドンも見出されているが、その意義・進化的な起源については完全には明らかになっていない。
● どのような遺伝子が(どのような形質を支配する遺伝子が)、生物界に広まりやすいのだろうか。ドーキンスは次のような問いかけを行った。
「そうしたいときにはいつでもまともな用語に直せるという自信があるなら、不正確を承知の上で、遺伝子が意識的な目的を持っているかのように語ることができよう。そうしたら、私たちは次のように問うてみることができる。では個々の利己的な遺伝子の目的はいったい何なのか?」ドーキンス『利己的な遺伝子』より