
【高校生物】遺伝子④「遺伝子発現はどのように調節されているのか?」

~プロローグ~
ある遺伝子の発現を他の遺伝子の産物が制御する、すなわち、遺伝子の働きを調節する機能を持つ「調節遺伝子」が存在するという考え方は、分子生物学の初期に導入された重大な概念であった。この概念は、ジャコブとモノー(2人ともフランス生まれの分子生物学者)によって提唱された。
「遺伝のメッセージの読み取りは、喫茶店のジュークボックスが出す音楽に似ている。ボタンのひとつを押すことによって、その機械のレコードの中から聞きたいと思うレコードを選ぶことができる。」ジャコブ『生命の論理』より
★テストに出やすいワード
①オペロン
②プロモーター
③オペレーター
④構造遺伝子
⑤一遺伝子一酵素説
要点:真核細胞では転写は核内で、翻訳は細胞質で行われるが、原核細胞はどちらも細胞質で行われる。
● 真核細胞では、核膜が転写およびスプライシングの場(核)と、翻訳の場(細胞質基質)を分けている。
● 核膜のない原核細胞では、転写と翻訳は同時に同じ場所(細胞質基質)で行われる。また、原核細胞のDNAは環状であり、(一部例外を除いて)スプライシングは行われない。
● 原核細胞では、転写によって生じたmRNAに、すぐに(まだ転写が完了していないのに)リボソームが結合し、翻訳が始まる(1本のmRNAに、複数のリボソームが結合する。リボソームは、mRNAの5'末端側に結合し、ポリペプチドを合成しながら、mRNAの3'末端側に[RNAポリメラーゼに近い方に]に進んでいく)。
下図はイメージ。

講義動画【原核生物の転写・翻訳】
発展:真核生物と原核生物のmRNAの違い(モノシストロンとポリシストロン)
ふつう、真核生物のmRNAは、翻訳に際して、1種類のポリペプチド鎖のみをコードしている。覚えなくてよいが、このような真核生物のmRNAを「モノシストロン性mRNA」という(単一のオープンリーディングフレームをもつ)。
対して、原核生物の1本のmRNAは、複数の異なるポリペプチド鎖をコードしている(1本のmRNAが複数のオープンリーディングフレームをもつ)ことが少なくない。覚えなくてよいが、このようなmRNAを「ポリシストロン性mRNA」という。ポリシストロン性mRNAは一連の代謝経路に関連した酵素系をコードしていることが多く、これらの酵素系の発現を同一レベルに調節するのに役立っている。
*オープンリーディングフレーム(ORF):「開いた読み枠」などともいわれる。終止コドンに中断されずにアミノ酸を指定するコドンが続く遺伝暗号の読み枠。長いオープンリーディングフレーム(開始コドンからはじまり、ある程度の長さ[多くの場合数百bp以上]をもって終止コドンまで続くひとつながりの読み枠)は、タンパク質の遺伝情報をコードしている可能性が高い。高校生は覚えなくてよい。
*シストロン:遺伝子と同義と考えてさしつかえない。基本的に高校では使わない用語。
要点:遺伝子の発現は調節されている。
(1)オペロン説
● オペロン説 :ジャコブとモノーが提唱。オペロンとよばれる転写単位が、調節遺伝子によってつくられた調節タンパク質による調節を受けるとする説。
*オペロンとは、同一のオペレーターとプロモーターの制御下にある複数の遺伝子のセットである(が、オペロンの定義は厳密には決まっていない。構造遺伝子群の領域をオペロンと呼ぶイメージを持っていればよい)。単一の「オペ」レーターに制御される単位だから「オペ」ロンである。
①プロモーター :転写開始時にRNAポリメラーゼが結合する塩基配列。
②オペレーター :調節タンパク質が結合する塩基配列。
③調節遺伝子 :調節タンパク質をコードする遺伝子(調節タンパク質=他の遺伝子の発現を促進したり抑制したりするタンパク質)。転写を抑制するような調節タンパク質をリプレッサー(repress=抑制)という。
④構造遺伝子 :酵素などのタンパク質をコードしている遺伝子(「構造」遺伝子は、酵素などのタンパク質の一次「構造」を決定する情報をもっている)。
*調節遺伝子が舞台裏で指示する監督、構造遺伝子が主演のようなイメージ(もちろん調節遺伝子が重要でないという意味ではない。遺伝子発現の調節は非常に重要で、正確に行われる必要がある)。

(2)ラクトースオペロン
● ラクトースオペロン(ラクトースがあり、グルコースがない場合のみ、ラクトース分解にかかわる酵素群が発現される):グルコースがなくラクトースがあるとき、リプレッサーがはずれる(ラクトースが変化したアロラクトースがリプレッサーに結合する。すると、リプレッサーの立体構造が変化し、リプレッサーのオペレーターに対する親和性が下がる)。その結果、構造遺伝子の転写が活性化される。
下図はイメージ(DNAが線状に描かれているが、大腸菌のDNAは環状である。念のため)。
RNAポ:RNAポリメラーゼ
リプ:リプレッサー
プロ:プロモーター
オペ:オペレーター
構造:構造遺伝子

発展:ラクトースオペロンのCAPによる調節
細胞内にグルコースが豊富にある場合、別な仕組みが働き、ラクトース分解に関わる酵素群の転写は抑制されることが知られている(細胞内にグルコースがないとき、大腸菌はcAMPを生産する。cAMPがCAP[環状AMP受容タンパク質]に結合すると、CAPはDNAに結合できるようになり、ラクトース分解に関わる酵素群の転写を活性化させる[オペロンをONにする])。
つまり、①グルコースがなく(CAPが活性化し)、②ラクトースがある(リプレッサーがオペレーターからはずれる)時にのみ、オペロンがONになる。

雑談:ジャコブとモノーが提示したモデルは「グルコースとラクトースが両方存在している培地で生育している大腸菌がラクトース分解酵素を発現しないのはなぜか」という疑問に触れなかった。実際は、上図のように、CAPが関係していた。
(3)トリプトファンオペロン
● トリプトファンオペロン(宿主がトリプトファンを多く食事を含む食事をしている時など、トリプトファンを大腸菌が自分で合成する必要がない場合は、トリプトファン合成にかかわる酵素の発現は抑制されている):リプレッサーがトリプトファンと結合し、活性化される(オペレーターとの親和性が上がる)。活性化したリプレッサーによって、トリプトファン合成酵素(トリプトファンの合成にかかわる数種類の酵素の遺伝子群)の遺伝子の転写が抑制される。
下図はイメージ。

<Q.ラクトースとかトリプトファンって何?…ラクトースの別名は乳糖。哺乳類の乳にも豊富に含まれる糖。トリプトファンはアミノ酸の一種。>
雑談:オペロン説が発表された当時、リプレッサーは単離されておらず、それがRNAなのかタンパク質なのかさえわかっていなかった。
講義動画【オペロン説の概要】
講義動画【オペロン説の詳細】
雑談:ラクトースオペロンの構造遺伝子には、lacZ,lacY,lacAがある。lacZはラクトース分解酵素の遺伝子、lacYはラクトースを細胞内に取り込む酵素の遺伝子である。lacAはガラクトシドアセチル基転移酵素だが、そのはたらきの生理的意義についてはまだよくわかっていない。
雑談:ラクトースの非存在下でも、リプレッサーはたまにオペレーターからはずれる(生体分子の相互作用は、ふつう0か1かではない。ほとんどすべては親和性の問題である。たまにリプレッサーがオペレーター配列からはずれるからこそ、ラクトースを細胞内に取り入れる酵素が発現し、周囲にラクトースが添加された場合に、それを効率よく取り込めるのである。それに、アロラクトースは、ラクトース分解酵素によってラクトースから作られる)。だから、正確には、ラクトースの有無にかかわらず(ラクトースがない場合も、少しは)構造遺伝子が発現している。
雑談:大学では、リプレッサーのように、他のDNA鎖に影響を与えることができる因子をトランスに作用する因子(トランス作用因子=転写因子)とよぶ。対して、CAP結合部位(CAPが結合するDNA領域)が転写開始能力に影響を与えるように、『同一の』鎖に影響を与える配列を、シス調節配列と呼ぶ(トランス作用因子という考え方は、ジャコブとモノーのオペロン説の基盤となった)。大腸菌はふつう単相である。しかし、遺伝子導入(DNAを導入する)によって複相にすることができる。複相の研究によって、鎖を超えてトランスに作用するリプレッサー(トランス作用因子)の存在が明らかになり、また、オペレーターがシスの状態に連鎖した遺伝子座にのみ作用する[オペレーターは、自身と同じ鎖の上にある遺伝子の発現調節に関わる]ことがわかった。これらの事実は、ジャコブとモノーがオペロン説を立てる根拠になった。
雑談:実際は、ラクトースオペロンの場合、プロモーターとオペレーターは重なっている(厳密には、オペレーターの1つはプロモーターと隣り合っており、もう一つのオペレーターはlacZのコード領域内にある。四量体であるリプレッサーは、2か所のオペレーターに同時に相互作用し(下図)、その結果、DNAのループが形成される。すると、RNAポリメラーゼとプロモーターの結合が抑えられる[しかし、RNAポリメラーゼとリプレッサーの両方がDNAに結合している場合があることも知られている。リプレッサーは、転写の開始に必要なDNAとRNAポリメラーゼの構造の変化を抑制することによっても、転写を抑制している可能性がある])。トリプトファンオペロンの場合は、オペレーターはプロモーターの内部にある。
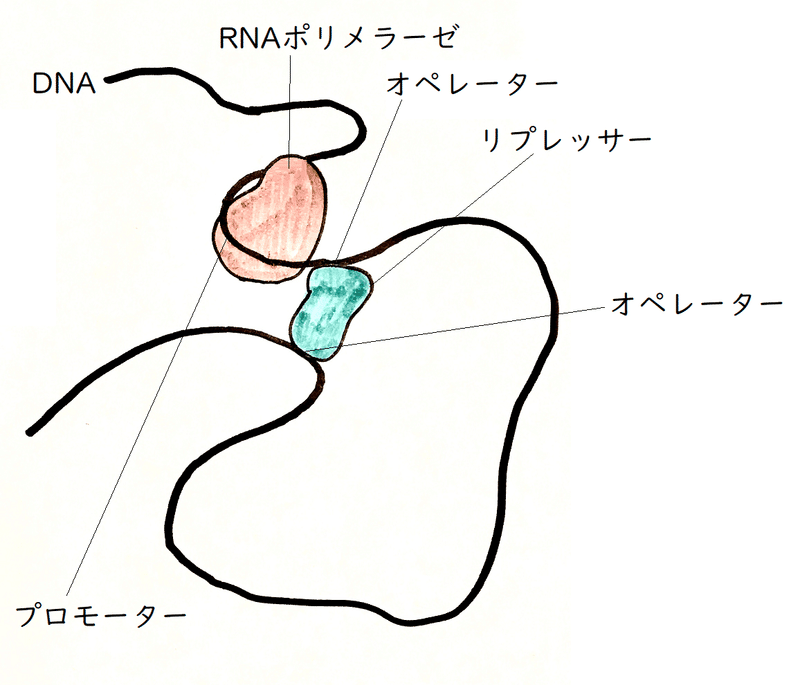
雑談:その働きを考えれば明らかだが、調節遺伝子は、ラクトースオペロンのように、必ずしもプロモーターの近くにある必要はない。実際、トリプトファンオペロンでは調節遺伝子はプロモーターから遠く離れたところにある。
雑談:遺伝子を書く場合は斜体にするのが普通である。同じアルファベットが遺伝子を表しているのか、そこから発現するタンパク質を表しているのかを区別するため、遺伝子の方を斜体で書く。
発展:アラビノースオペロン
アラビノースオペロン(araオペロンともいう)の発現調節には、AraCと呼ばれるタンパク質が関わる(アラビノースは糖の一種)。
アラビノースがAraCに結合すると、下図1のような配置になり(隣り合うDNA上の2つの領域①②に、二量体の形で結合できる立体構造になり)、転写が活性化される(アラビノースの代謝に必要な酵素群をコードしている遺伝子の発現が活性化される[知らなくてよいが、大腸菌のアラビノースオペロンでは、araB, araA, araDの3つの遺伝子が、それぞれ、リブロキナーゼ、アラビノースイソメラーゼ、リブロース-5-リン酸エピメラーゼというアラビノースの代謝に関わる酵素をコードしている])。
領域②にAraCが結合することが、転写の活性化に重要であることがわかっている。
アラビノースが存在しない場合、図2のような配置でAraCはDNAに結合する(一方の単量体が、図1に比べて、遠くのDNA領域[③]に結合する。それぞれの単量体が結合する領域[①と③]は194塩基対離れているので、AraCが結合すると、両部位間のDNAは湾曲し、ループを形成する)。この配置の時は、(領域②にAraCが結合していないので)転写は活性化されない(なお、アラビノースオペロンはCAPによる制御を受けるが、図には描いていない。グルコースが存在しない場合、CAPが転写の活性化を助ける)。
すなわち、AraCは、単独ではリプレッサーとして、アラビノースに結合した状態ではアクチベーター(転写活性化因子)として機能していることになる。
(なお、知らなくてよいが、下図のDNA領域①、②、③は、正確には、それぞれaraI₁、araI₂、araO₂と呼ばれている)
図1

図2

要点:ビードルとテータムは一遺伝子一酵素説を提唱した。
● 「一つの遺伝子が一つの酵素合成を支配している」という考え方を一遺伝子一酵素説といいビードルとテータムがアカパンカビ(子のう菌類:真核生物)を用いた実験で提唱した。
語呂「ビードル!そのパンカビ生えてるってー!」「いちいちうるさいなあ」(ビードルとテータムは、アカパンカビを用いた実験により、一遺伝子一酵素説を提唱した。)
● 野生型の(変異のない)アカパンカビは、糖、無機塩類、ビオチン(ビタミンの一種)という単純な栄養だけで生育できる。これらの栄養しか含まない培地を最少培地という。
● 野生型のアカパンカビは、最少培地に含まれている糖から、アルギニンというアミノ酸を合成する。この反応系では、一つ一つの反応に別々の酵素が働くと考えられている。
代謝経路:前駆物質→オルニチン→シトルリン→アルギニン
語呂「おっしゃる(オルニチン、シトルリン、アルギニン)」
● X線などでアカパンカビの突然変異株をつくることで一遺伝子一酵素説の研究が行われた。
● X線を照射し、アルギニンをつくる代謝経路で働く酵素をコードする遺伝子を、1つ壊す(どの酵素をコードする遺伝子が壊れたかは、培地にどの栄養を加えれば生育できるかを調べることで確かめられる)。
● 最少培地では生育しないが、最少培地に特定の栄養を加えて培養すると生育可能になる突然変異株を、栄養要求株という。
● 代謝経路:前駆物質→①→オルニチン→②→シトルリン→③→アルギニン
において
*①に働く酵素をα、②に働く酵素をβ、③に働く酵素をγとする。
*酵素αがつくれない変異株Ⅰ、酵素βがつくれない変異株Ⅱ、および酵素γがつくれない変異株Ⅲがいたとする。
培地にどの栄養を加えれば、変異株Ⅰは生育できるだろうか?
たとえば、変異株Ⅰ(酵素αをもたない。酵素β、γはもつ)は、前駆物質を与えられても、(酵素αがないため)オルニチンに変えることができない。したがって最終産物であるアルギニンの材料であるオルニチンが得られず、死んでしまう。しかし、オルニチンやシトルリンが与えられると、アルギニンがつくられるので生育できる。
それぞれの変異体についてまとめると以下の図のようになる。
生育できる場合+、できない場合-を記す(変異はただ一か所のみに起きたとしてよい)。
前駆物質→①→オルニチン→②→シトルリン→③→アルギニン
*①に働く酵素をα、②に働く酵素をβ、③に働く酵素をγとする。
変異株Ⅰ:酵素αがつくれない
変異株Ⅱ:酵素βがつくれない
変異株Ⅲ:酵素γがつくれない


*一遺伝子一酵素説の例外もよくテストに出る。
タンパク質の四次構造を考えた場合、サブユニットは別々の遺伝子によって支配されている場合がある(一遺伝子一酵素ではなく、複数の遺伝子が1つの酵素のそれぞれのサブユニットをコードしていることがある)。
また、酵素以外をコードする遺伝子もある。したがって一遺伝子一酵素説は常に正しいとは言えない。
では一遺伝子一ポリペプチド説とすれば正しいか?
いやいや、選択的スプライシングが起きる場合、一つの遺伝子から複数種のポリペプチドができる。
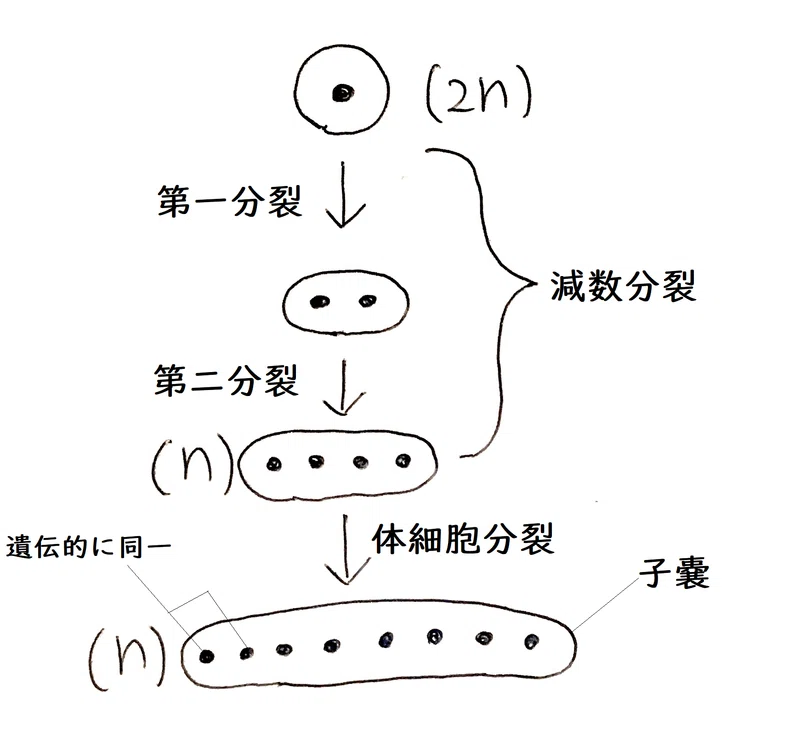
<Q.どうしてアカパンカビを用いたの?…アカパンカビはふつう単相(n)で生活するので、遺伝子型がそのまま表現型として現れるので分析しやすい。また、減数分裂から生じる4個の細胞が「子嚢(しのう)」の中に並んで作られるので研究がしやすい。>
雑談:アカパンカビでは、接合によって生じた接合子の核(2n)は、すぐに減数分裂を行い、4個の単相の核(n)になる。その後、1回、核だけが分裂する体細胞分裂を行い、8個の核(n)になる。この核の周りに細胞膜と細胞壁ができて、子嚢胞子となる。アカパンカビの子嚢では、分裂した細胞が移動しないので、分析が容易い。
講義動画【一遺伝子一酵素説】
雑談:フェニルケトン尿症について
フェニルケトン尿症は尿中にフェニルケトンが大量に排出される病気で、放置すれば脳の発育に障害が起こる。この病気の原因は、フェニルアラニンをチロシンに変換する酵素の遺伝子の塩基が変化したことによって、正常な酵素がつくられなくなったことである。この酵素がないと、本来ならばチロシンヘと変換されるフェニルアラニンが余ってしまう。すると、フェニルアラニンからフェニルケトンが大量につくられ、体内のフェニルケトン量が多くなって障害が発生する。乳幼児に神経系の障害を引き起こすため、フェニルアラニンを除いたミルクで育てる必要がある。
雑談:アルカプトン尿症について
アルカプトンを酸化する酵素が欠けているために起こるアルカプトン尿症という遺伝病も知られている。アルカプトン尿症は、ホモゲンチジン酸(アルカプトン)オキシダーゼの欠損が原因であり、単一の酵素の遺伝的伝達と関連づけられた初めての病気であった。尿中にどんな物質が含まれているかはっきりわかっていなかった頃、アルカプトンという名称が充てられたが、現在ではその実体はホモゲンチジン酸であることがわかっている。1902年(ビードルとテータムの一遺伝子一酵素説の発表よりも前のことである)、ギャロッドは、一つの遺伝単位(後に遺伝子と呼ばれる)のせいでアルカプトン尿症患者の尿か黒く変化することを提唱した(欠損した酵素の基質であるホモゲンチジン酸が、多量に尿中に排出される。ホモゲンチジン酸は、尿が空気にさらされると同時に酸化され、黒く変色する)。一般に、アルカプトン尿症の患者は関節炎症状を示す。

雑談:ビードルは、ノーベル賞の受賞講演で次のように述べた。「私たちはこの長い回り道を経て、ギャロッドが何十年も前にすでに明らかにしていた事実を再発見したに過ぎない。私たちは彼の研究を知っていたし、本質的な新しいものを何か付け加えたわけではない。私たちは、ヒトのいくつかの遺伝子といくつかの化学反応についてギャロッドがすでに明らかにしていたことが、アカパンカビの多くの遺伝子と多くの化学反応にも該当することを証明しただけである。」
雑談:アルビノについて
動物で、遺伝的な要因でメラニンの形成が行われないために、皮膚、毛髪、眼などに色素が生じない個体のいことをアルビノという。原因の多くはチロシナーゼの欠損である。メラニンは皮膚のメラニン細胞など特定の細胞でチロシンからつくられる。アルビノ(色素が欠如している)は太陽光に対して極端に敏感であることが多い。また、ひどい日焼けや皮膚がんが多いだけでなく、視力が弱いことが多い。
発展:真核生物の転写調節
● 真核生物の転写の開始には基本転写因子が必要であることは重要(真核生物では、プロモーターに正しくRNAポリメラーゼが結合するために基本転写因子が必要)。基本転写因子については、遺伝子③の資料も参照せよ。
● 真核生物では、同じ機能に関わる遺伝子が、同じ塩基配列の転写調節領域をもつことで、同じ調節タンパク質に調節され、協調的に発現することがある。
講義動画【真核生物の転写調節】
発展:TATAボックス
真核生物でも、原核生物と同様に、プロモーター領域にRNAポリメラーゼが結合してmRNAの合成が開始される。転写開始部位からおよそ25塩基離れたところに、TATAボックス(プロモーターのほとんどすべてに共通する重要な配列)という塩基配列が存在する。
さらに、ずっと離れた数kbp上流に転写調節配列があって、ここに調節タンパク質(転写因子)が結合し、転写の速度を制御していることもある(転写因子によって、転写を促進する場合も抑制する場合もある)。
*kbp:キロベースペアー。1000塩基対を表す。
発展:RNA干渉・ゲノム編集
高校では、タンパク質がDNAに結合して遺伝子発現を調節することを学ぶが、実は、RNAも遺伝子発現を調節している。

*RNA干渉(RNAiと略記):二本鎖RNAによって配列特異的にmRNAが分解される、あるいは翻訳が抑制される結果、遺伝子発現が抑制される現象。
*CRISPR(くりすぱー)系:細菌が短い非翻訳RNA分子を用いて行う防御機構。外来ウイルスゲノムを探し、破壊する(ウイルスが細菌に感染すると、細菌は、ウイルスDNA断片を自身のゲノムに組み込む。組み込まれたDNAを鋳型として作られたRNAが、再度侵入してきたウイルスの破壊に働く)。細菌のCRISPR系を用いた免疫についてはまだわかっていないことが多いが、この現象を利用して、DNA上の好きな配列を切断する技術が発明された(ゲノム編集)。
講義動画【RNA干渉、CRISPR系】
以下全て上の動画についての雑談。
・RNAi(RNA干渉)系は真核生物に広範囲に存在する。
・ヒトでは、タンパク質をコードする遺伝子の50%近くが、RNAによる発現調節を受けているとも言われている。
・上の動画のようなRNA干渉で用いられる短いRNA鎖をmiRNA(マイクロRNA)という。このほかにも、siRNA(低分子干渉RNA)というRNAがガイド役として使われることもある。 siRNAはmRNAの選択的分解の誘導により翻訳を阻害したり、クロマチン構造の凝集によって転写を抑制したりする(後者のような転写抑制の場合、siRNAとアルゴノートなどのタンパク質との複合体はRITSと呼ばれる。RITSはヒストンのメチル化、DNAのメチル化を介して転写を抑制する)。
・miRNAとsiRNAは、その前駆体により区別される。miRNAが前駆体RNAのヘアピン構造の1つから生成されるのに対して、siRNAはもっと長い二本鎖RNAから多数生じることがわかっている(たまに、miRNAとsiRNAの違いを「自然に細胞で生じるのがmiRNAで、ウイルスに由来する外来二本鎖RNAから生じるのがsiRNA」とする説明があるが、正確ではなく、siRNAには内在性のものもある[2本鎖RNAには、自然には、RNAウイルスやトランスポゾン由来のものがある。DNAのある領域で逆方向にRNAが転写され、生じたRNAが結合して2本鎖になることもある]。細胞内に2本鎖RNAがあると、ダイサーにより切断され、siRNAが生じる)。
・生殖系列の細胞で特異的に作られるpiRNAもある(トランスポゾン遺伝子を転写レベルで抑制することで、生殖系列の細胞を守っていると考えられている)。
・多くの植物では、miRNAと標的mRNAは広域で結合し、アルゴノートがmRNAを切断することで急速なmRNA分解を促している。
・多くの哺乳類では、miRNAは標的mRNAと狭い領域のみで結合し、急速に翻訳を抑制させている。この場合も、mRNAはやがて分解される(mRNAは不安定になり、P-体[細胞質にある]に移行して、やがて分解される)。
・ゼブラフィッシュでは、母性mRNAの排除にmiRNAが関わっていることがわかっている(母性因子の一斉処分)。
・RNA干渉は、ウイルスなどの外来DNAや、トランスポゾンに対する防御機構だったようである。今では、正常な発生を含め、様々な生命現象に関わっている。
・miRNAやsiRNAなど、タンパク質をコードしていないRNAをncRNA(ノンコーディングRNA)という。
・RISCは、RNA-induced silencing complex(RNA誘導サイレンシング複合体)の略。
・miRNAなどによるRNA干渉とCRISPR系は似ているが、決定的に異なる点が2つある。 ①RNA干渉では主に1本鎖RNAが標的だが、CRISPR系は2本鎖DNAが標的である。 ②RNA干渉とは異なり、CRISPR系では、遺伝情報のCRISPR座位への取り込みが起こる。
・ゲノム編集には、よくCRISPR/Cas9系(クリスパーキャスナインけい)が使われる。この系を用いて決まった位置でDNAの2本鎖を切断させる。それで遺伝子を壊すこともできるし、少し難しいが、遺伝子をノックインすることもできる。DNAを切断された細胞は、その修復に、われわれが導入した遺伝子断片を使うことが多い。細胞は、壊れた(切られた)領域の修復に、我々が注入したDNA断片を参照するのである。結果、狙った位置に、我々が導入した遺伝子が組み込まれることになる。この組み込みの仕組みを、相同組換えという。
・RNA干渉とCRISPR系は似ている所があるが、進化的なつながりは完全には明らかになっていない。真核生物は、多細胞化に伴って、CRISPR系のような「細胞を守る」免疫システムから、インターフェロン(ウイルス感染などにより誘発されて宿主細胞が生産するウイルス増殖抑制タンパク質)などを用いた「個体を守る」免疫システムに切り替えていった可能性がある。
・RNA干渉は、もともと、実験的に二本鎖RNAを導入し、相補的mRNAの配列特異的な分解を引き起こす機構を指す語であった。しかし、近年では、RNA干渉の語は、細胞自身のゲノムにコードされているmiRNAなどによる遺伝子発現阻害を含めた意味でよく使われている。
・アルゴノートとは、もともとは、ギリシャ神話において黄金の羊毛を探しに出航したアルゴ船の船員のこと。
発展:トランスポゾン
(高校生はDNA上を動く配列が存在するということだけ知っておけばよい)
ゲノム内のある位置から他の位置へと移動することができるDNA領域をトランスポゾン(転移因子)という。トランスポゾンが他の遺伝子の間に入り込んでしまうと、その遺伝子の働きが失われ、深刻な影響が出る場合もある。
雑談:以下、トランスポゾンについての雑談。
・ある種のトランスポゾンには、トランスポザーゼという酵素の遺伝子が含まれ、転移の際、この酵素がトランスポゾンの両端に作用して、トランスポゾンを新たな部位に挿入することが知られている。
*トランスポザーゼ(トランスポゼース):あるDNA領域から他のDNA領域へとトランスポゾンが転移する際のDNA組換え反応(DNAの切断と再結合)を触媒する酵素。
・トランスポゾンには大きく2種類がある。
①DNA型トランスポゾン:転移にはトランスポザーゼが使われる。移動の間もDNAとして存在する。
②レトロトランスポゾン:転移には逆転写酵素などが使われる。転写により一度RNAがつくられ、そこからDNAが逆転写によってつくられる。最終的に二本鎖DNAがゲノムに挿入される。レトロウイルスはレトロトランスポゾンから進化したとも考えられている。
・トランスポゾンはゲノムのさまざまな位置に入り込むことができるが、ほとんどのトランスポゾンはめったに動かない(高頻度の転移は、宿主のゲノムを壊してしまうだろう。また、一般に、宿主となる生物はトランスポゾンの転移を抑制する様々な機構を持ち、自身のゲノムを保護している)。
・どの生物でも数多くのトランスポゾンが確認されている。
・トランスポゾンは進化の歴史において重要な役割を演じてきたと考えられている。
・たとえばヒトゲノムでは、トランスポゾンに関連する配列は(トランスポゾンの断片や変異で不活性化した[ある意味"死んだ"]因子も含めれば)ゲノムの50%以上を占めると考えられている。
トランスポゾンのアイデアははじめはまったく受け入れられなかった(君たちも一度聞いただけでは信じられないだろう)。1931年、トランスポゾンの発見者マクリントック(アメリカの遺伝学者)は、紫色と白色の斑入りを示す穀粒を付けるトウモロコシの変異体から、トランスポゾンの着想を得た(マクリントックは、ゲノム中のある領域から、穀粒の色に関する遺伝子の内部に移動する遺伝因子が存在し、穀粒の色を変化させていると考えた。実際、トランスポゾンの転移の時期と頻度などによって、斑入りの程度が粒ごとに異なることがわかっている)。トランスポゾンのアイデアは、当時、ほとんど無視された。
トランスポゾンの正体が分子レベルで明らかになり、マクリントックがノーベル生理学・医学賞を受賞したのは、1983年のことである。その時マクリントックは81才だった。
受賞の知らせを聞いた彼女は一言「おやまあ( Oh my dear! )」とつぶやいた。
RAY SPANGENBURG 、 DIANE KIT MOSER 著『ノーベル賞学者 バーバラ・マクリントックの生涯 ―動く遺伝子の発見― 』によれば、「評価されるまで長い時間がかかりましたが、大変でしたか?」という記者の質問に対して、彼女は、次のように答えたという。
「ノー、ノー、ノー」「ずっと楽しく過ごしてきたし、公の評価なんて必要ないのよ」
非常に美しい言葉である。
発展:ヒストンのアセチル化による転写活性化
ヒストンに修飾が加わることで遺伝子発現が制御されていることが明らかになっている。まだそのメッセージの意味は完全には確定できていない。たとえば、一般に、ヒストンのアセチル化は、転写活性化に働く(クロマチン構造をゆるませることによる[アセチル化によって電荷が変化したヌクレオソームは、クロマチン繊維を形成しにくくなる])。
雑談:ヒストンは、アセチル化以外に、メチル化もされ得る。ヒストンのメチル化は、転写活性化と転写抑制の両方に関与する(修飾される部位によって効果が異なる)。
発展:エピジェネティクス
● DNAの塩基がメチル基などで修飾されることで、遺伝子発現が調節されていることが明らかになった(メチル化された塩基配列は、転写装置や活性化因子の結合を妨げることがある)。
● 塩基配列に変化を与えずに、後から行う修飾により、遺伝子の発現を制御する機構の総称を「エピジェネティクス」という。
● 心臓では心臓に必要な遺伝子が発現し、肝臓には肝臓に必要な遺伝子が発現する(選択的遺伝子発現)。その特異性および可塑性は発生段階でエピジェネティクスによって確立され、決定される(受精卵が体細胞分裂で数を増やしても、基本的に塩基配列は変わらない。細胞が個性を持つためには、どこかで、塩基配列によらない調節をする必要があるだろう)。
● DNAメチル化、ヒストン修飾、非コードRNAなどによるエピジェネティックな遺伝子発現調節機構の解明が進んでいる。
● 「エピ」はギリシャ語で「後で、上に」を表す。エピジェネティクスは、「遺伝子の上に修飾が加わった」ということを表す(語源は後成説epigenesisに由来する)。
● ヒストンは、メチル化、アセチル化、リン酸化などのさまざまな修飾を受ける(これらはクロマチン構造のダイナミックな変化に関与している)。
● DNAのメチル化は、哺乳類ゲノムを直接的に修飾する唯一の仕組みであり、このメチル基を付加したりはずしたりすることによって遺伝子発現調節を行う。
● エピジェネティクスは、ゲノムインプリンティング(ゲノム刷り込み。後述する)や、X染色体不活性化、細胞分化、がん化などに関わっている。
発展:ゲノムインプリンティング(ゲノム刷り込み)
● ゲノムインプリンティング:遺伝子発現が、親の由来によって異なる調節を受ける現象(その意義については謎が多い)。雄親由来の遺伝子が休止し、雌親由来の遺伝子が活性をもつ場合もあれば、その逆の場合もある。
雑談:様々な生物種でゲノムインプリンティングは発見されている。ヒトでは約300個の遺伝子がゲノムインプリンティングを受けている。
● たとえば、下図の場合、雄親由来の遺伝子Aは、(塩基が)メチル化されていて、発現しない。生殖細胞がつくられる過程で、一度メチル化は解除される。そして、生殖細胞の発達に伴い、(今回の例では)精子内の遺伝子Aがメチル化されることになる。
下図には描いていないが、実際はDNAは2本鎖である。メチル化を受ける場合、2本の鎖は両方メチル化される。

まだわかっていないこと
● 真核生物はどのように遺伝子を調節しているのか、完全には明らかになっていない。真核生物は原核生物と異なり、同じことに関わる遺伝子が、互いに離れた位置にあることがある。このことは真核生物の遺伝子発現調節の解析を困難にしている。
● 真核生物の場合は、原核生物のオペロンのような構造を取らない。つまり、一つの遺伝子は一つのポリペプチド鎖しかコードしていない。原核生物では、複数の関係する酵素が連続して転写され、1つのRNAに複数のタンパク質の情報が入る。この事実に、どのような進化的・適応的な意味があるのか。
● エピジェネティックな現象については、未解明な部分が多い。
● ゲノムインプリンティングがなぜ存在するのかは謎である。ゲノムインプリンティングの仕組みの破綻は、疾患の原因になることが知られている。