
【高校生物】植物生理①「植物はどのように刺激に反応するのか?」

~プロローグ~
植物は、人類の生活と深く関わっている。現在、多くの人々が、植物を栽培し、植物を食べ、植物に癒されている。また、古代エジプトでは、カミガヤツリの茎からパピルスが作られていた(パピルスは紙paperの語源)。人類は植物を利用して、自らの精神活動を後世に伝えたのである。
そして未来、食糧問題や環境問題が深刻化していく中で、植物の研究が果たす役割はますます大きくなっていくであろう。
下の写真はカミガヤツリ。

★テストに出やすいワード
①屈性
②極性移動
③頂芽優勢
④傾性
⑤根冠
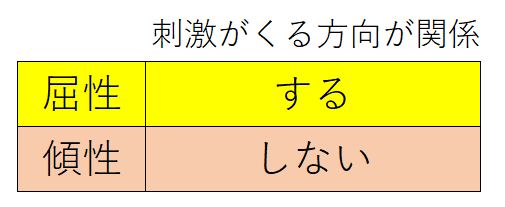
要点:植物が刺激に対して一定の方向に曲がる反応を屈性と言う。刺激の方向とは無関係に、ある一定の方向に屈曲する反応を傾性という。
(1)屈性
● 屈性(くっせい):植物が刺激に対して一定の方向に曲がる反応。基本的にすべて成長運動。刺激の種類によって、下表のように分けられる。刺激源の方向に向かう場合を正(+)、刺激源から遠ざかる場合を負(-)とする。

*下図は正の光屈性のイメージ。
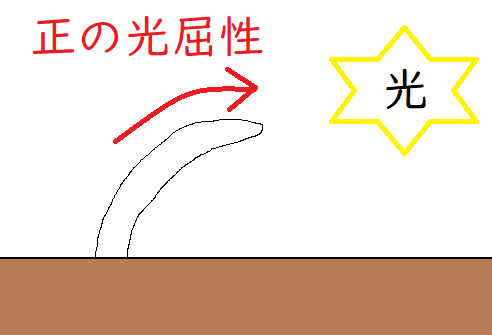
*巻きひげの接触屈性については、少し複雑である。後述する。
(2)傾性
● 傾性(けいせい):刺激の方向とは無関係に、ある一定の方向に屈曲する反応。成長運動と膨圧運動の両方で見られる。例 がよく問われる。
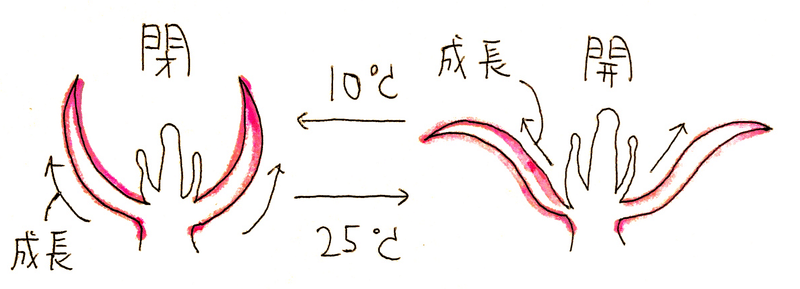
例1)チューリップの花の開閉(温度傾性)
*温度が高い(25℃)と花弁の内側が成長し、開く(逆に、低い温度では外側が成長する)。この反応には刺激の方向が関係しないので、これは傾性の例である(どこににストーブを置いて部屋を温めようが、温度が高ければ花は開く。右から温めたら右向きに開くなどと言うことはない)。
*チューリップの花の温度傾性は成長運動(成長速度の違いによって起こる運動)である。成長運動は一般に不可逆であるので、チューリップの花は、開閉を繰り返すたびに成長して大きくなっていく。
例2)オジギソウ(膨圧運動)の接触傾性
*どこから触れても(方向性なし)、葉がたれる。
*オジギソウの葉の開閉運動は、接触に関係なく、1日の変化によっても見られる。このような運動は「就眠運動(しゅうみんうんどう)」と呼ばれる。
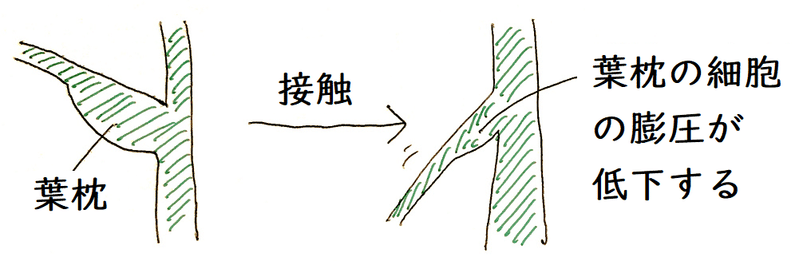
雑談:オジギソウなどのマメ科植物は、葉に葉枕(ようちん)をもつ。葉枕は、葉柄(葉の柄の部分)の下端が関節状に肥厚したものである。葉枕の細胞には、昼、カリウムイオンとともに水が入る→膨らむ。夜は、逆に水が出ていくことで膨圧が減る。就眠運動は、1日の周期を持つ概日リズムである場合が多い。就眠運動の正確な仕組みは未解明であるが、葉を閉じさせる活性を持つ物質(就眠物質)と、葉を開かせる物質(覚醒物質)のバランスによって制御されているのではないかと考えられている。
(3)成長運動と膨圧運動
● 成長運動:成長速度の違いによって起こる運動。一般に、すべての屈性と、一部の傾性(チューリップの温度傾性)に関わる。
● 膨圧運動:細胞の膨圧の変化によって起こる植物の運動。多くの傾性に関わる。
膨圧運動の例)
気孔の開閉、オジギソウの葉の就眠運動、オジギソウの接触傾性(カリウムイオンの透過による。イオンと、それにつられた水が細胞間隙へ移動する[膨圧が上昇する]。非常にすばやく反応が起きる。しくみは未解明)
発展:成長運動と膨圧運動
一般に、成長運動は不可逆であるが、膨圧運動は可逆的である。

雑談:他の物に巻きついて自分の体を保持・安定させるように変態した植物体の部分を、巻きひげと言う。つる植物に多いが、変態する器官はさまざまである(枝 [ブドウ]、葉([エンドウ]などの例がある)。巻きつきには、一般に、接触によって生ずる急速な成長運動が関わる。高校ではこの反応は接触した面を凹とするように屈曲する接触屈性とすることが多いが、接触刺激を受けた面と無関係に、決められた方向に屈曲する接触傾性を示すものもある(いずれの機構でも、棒などに巻き付くことになる)。エンドウの巻きひげはその中間型とも言え、接触刺激により接触面を凹とするような屈曲を示す面と、接触刺激に反応しない面とをもつと考えられている。下図はイメージ。
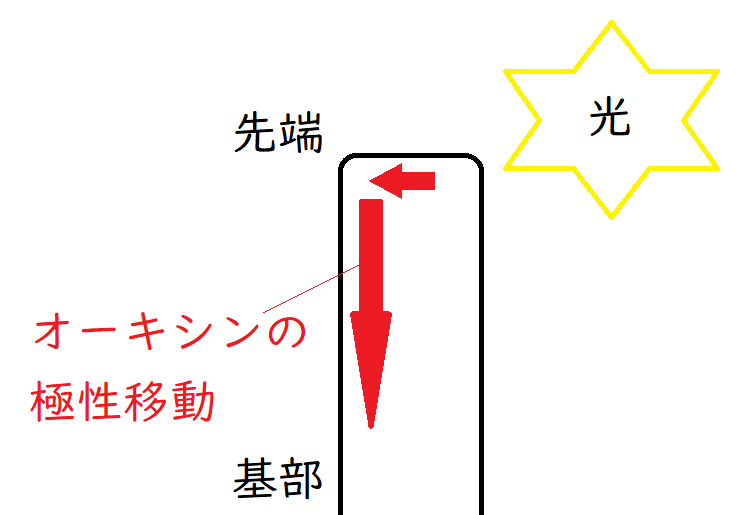
要点:オーキシンは光の受けていない側に移動し、基部方向へ移動し、伸長成長を促進させる。
(1)オーキシンの実体
● 幼葉鞘(ようようしょう。イネ科植物の子葉に相当すると考えられている器官)で見られる光屈性には、オーキシン(auxieはギリシャ語で成長。inは物質という意味)とよばれる植物ホルモンが関与している。
雑談:幼葉鞘は子葉鞘(しようしょう)とも呼ばれる。幼葉鞘は単子葉類のイネ科などに独特の器官で、発芽時に最初に地上に出る部分。イネ科植物の成熟種子には、胚乳を吸収するための器官である胚盤(はいばん。これもイネ科特有の器官)が形成される。胚盤は子葉の一部と考えることができるが、発芽の際、胚盤は地下に残り、子葉の残りの部分が地上に出る。この地上に出る部分は、葉というより、鞘のように筒状になって茎を包んでいるので、幼葉鞘と呼ばれる。
● オーキシンの化学的な実体はインドール酢酸(IAA)とよばれる化学物質である(インドール酢酸はオーキシンの化学物質名。天然に存在するオーキシンはほとんどがインドール酢酸である)。2,4-D(2,4-dichlorophenoxyacetic acid)や、ナフタレン酢酸にもオーキシンとしてのはたらきがある(2,4-Dとナフタレン酢酸は合成オーキシン)。
雑談:高濃度のオーキシンは植物の成長を抑制する。除草剤の中には、合成オーキシンである2,4-Dが含まれているものがある。一般に、2,4-Dは、イネ科植物に影響が小さく、安価なコムギ、イネなどの栽培における除草剤として広く使用されている。
雑談:天然に存在するオーキシンはほとんどがインドール酢酸なので、逆にインドール酢酸と同じ生理作用を有するものをオーキシンと定義している。
(2)オーキシンの性質
①茎の先端でつくられて、光の当たらない側に分布する。
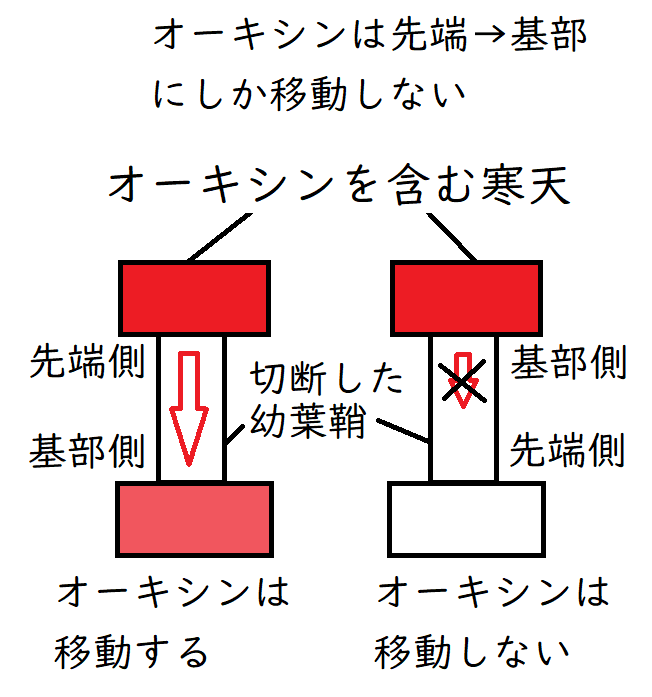
②基部(植物体の中心部)方向へ移動する。逆方向には移動しない(極性移動という)。
*オーキシンの極性移動は、重力に依存せず、エネルギーを用いて行われる。決してオーキシンは重力で落ちていくわけではない。幼葉鞘を切断し、逆さまにして置いても、先端部から基部へ移動する(重力逆らっても移動する)。
③下方に(基部方向に)移動したオーキシンは、細胞の成長を促進させる。
*オーキシンは細胞壁の主成分であるセルロース繊維の結びつきを弱める→細胞は吸水して成長する。
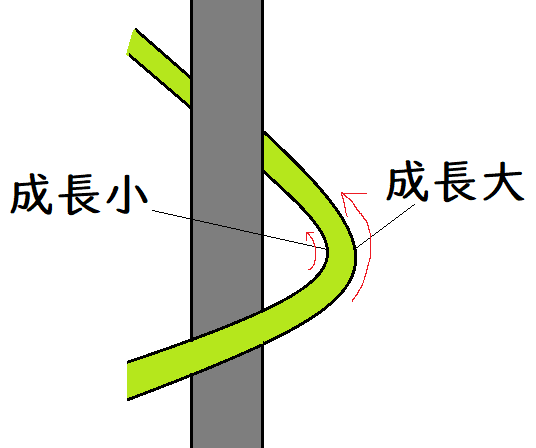
*オーキシンは、光の当たらない側で高濃度になっている。高濃度のオーキシンは茎を大きく成長させる。光の当たる側と、その反対側で、成長の大きさが異なる(光の当たらない方がよく成長する)ので、光のくる方向に屈曲する(茎が正の光屈性を示す)。
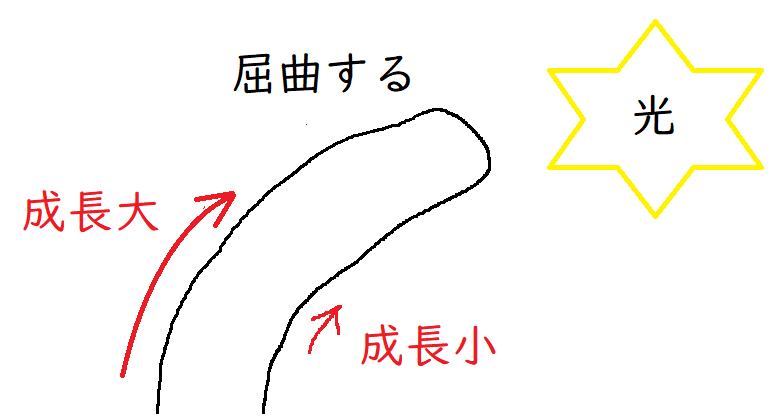
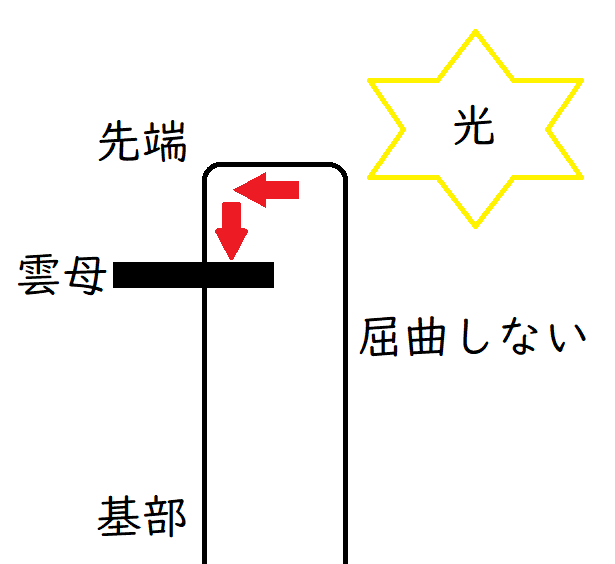
(3)オーキシンの移動に関する実験
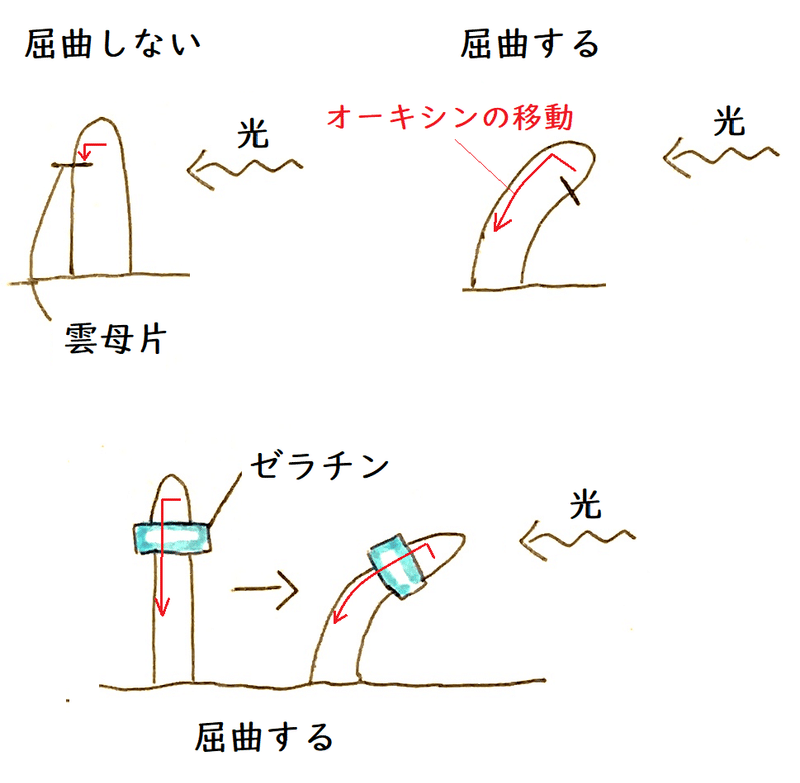
● たとえば、下図のように雲母片(オーキシンを通さない)をはさみ、右から光を当てるとする。
すると、オーキシンが下方に(オーキシンが成長を促進する部位に)移動できず、屈曲しない。
*雲母はうすく剥がれるので、オーキシンの移動を阻害する実験によく使われる。
(4)オーキシンの濃度
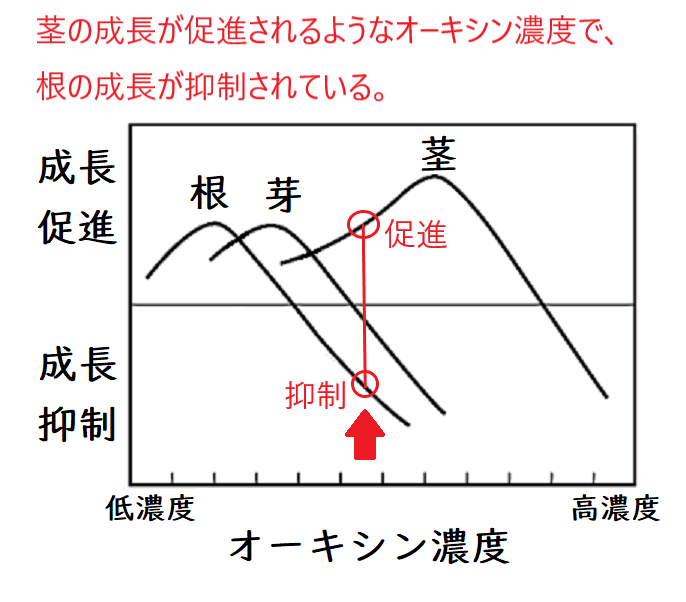
● オーキシンは細胞の成長を促進するが、その最適濃度は器官によって異なる。濃すぎると成長は抑制される。
*「成長が抑制される」とは、ちょっとしか成長しないということであり、縮むわけではない。
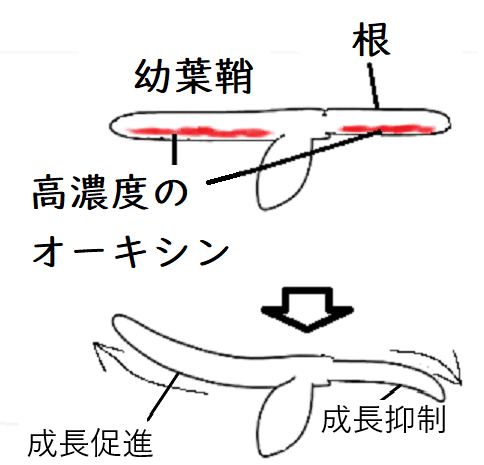
● 暗所に水平におかれた幼葉鞘では、オーキシンが下方に移動して、茎の下側の濃度が高まる。そのため茎の下側の成長が相対的に促進され、茎は負の重力屈性を示す。
● 根でも同様に下側のオーキシン濃度が高まるが、高濃度のオーキシンは根の成長を抑制するため、下側の成長が抑制されて正の重力屈性を示す。
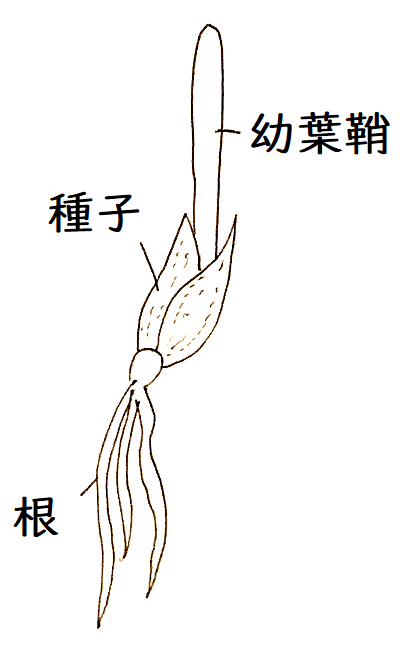
これを水平に寝かせる(下図)。暗所に水平におかれた幼葉鞘では、オーキシンが下方に移動することが知られている。
(5)根冠
● 重力方向は、根の先端にある根冠(こんかん)によって感知されている。
雑談:根冠を切除すると、根冠が再生されるまで根はランダムな方向に伸長する。

発展:根冠による重力の感知
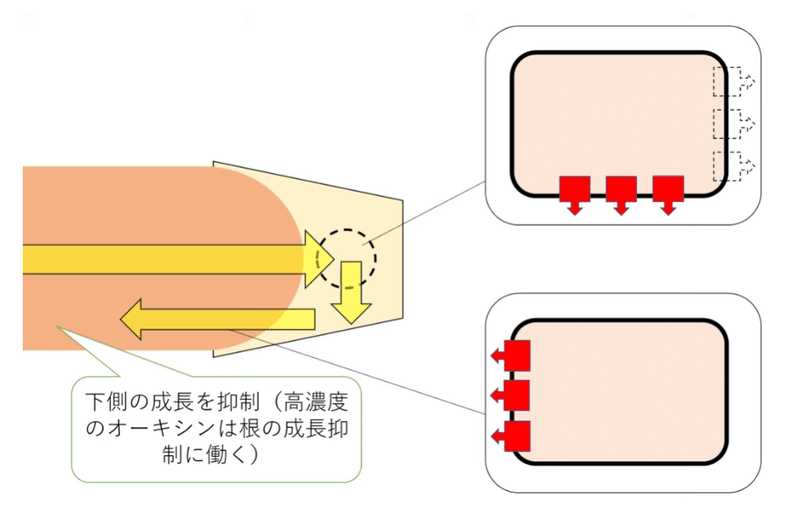
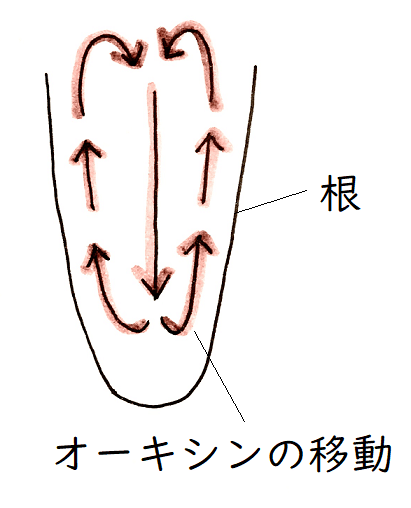
根冠を構成する平衡(コルメラ)細胞の中にはアミロプラストという色素体(しきそたい)があり、その中に重いデンプン粒が入っている。アミロプラストが重力方向に移動するため、根冠は重力の方向を感知することができる。我々も平衡石(耳石)をもっているが、維管束植物の平衡石は、重いデンプン粒を持つアミロプラストなのである(図1)。植物が重力を感知するしくみについては、まだ完全には解明されていないが、アミロプラストが移動することにより、PIN(オーキシンを細胞外に排出する輸送体)の配置が変化すると考えられている(下図には示していないが、垂直に根を置いた場合は、PINは細胞表面に均一に分布する。根を水平に置いた場合、PINは下側に優先的に配置されるようになる)。根を横に倒した場合は、図2のような経路でオーキシンが輸送されると考えられている。結果、根の下側で成長が抑制され、根は正の重力屈性を示す。
*図1、2にはAUX(オーキシン取り込み輸送体)は描いていない。
*PINが移動する(再配置される)仕組みは解明されていないが、PINは小胞膜上と細胞膜上を行き来しているという説がある。
*実際は、PINには様々な種類があるが、高校生は気にしなくてよい。
図1
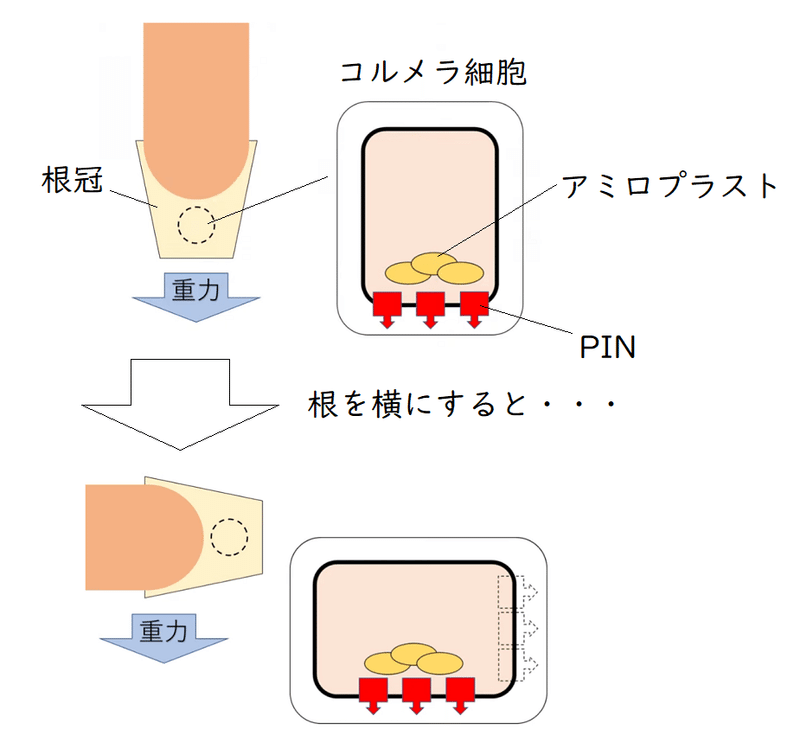
図2
雑談:植物細胞の細胞質に含まれる葉緑体、アミロプラスト、有色体(トマトの果皮、ニンジンの根などにある)、白色体(茎や胚乳、白い花弁にある)などの細胞小器官は、色素体と呼ばれている。
雑談:茎でも重力を感知している。茎では重力屈性の刺激の受容は皮層の内側に存在する内皮細胞層で行われると考えられている。内皮細胞には、根のコルメラ細胞と同様にアミロプラストが含まれている。内皮細胞欠失変異体では、根では重力屈性を示すが茎では重力屈性を全く示さない。
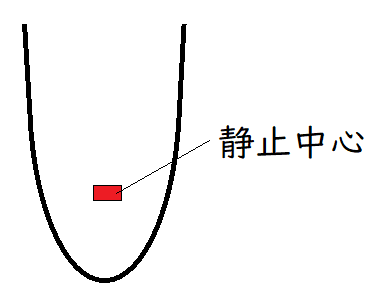
発展:静止中心(せいしちゅうしん)
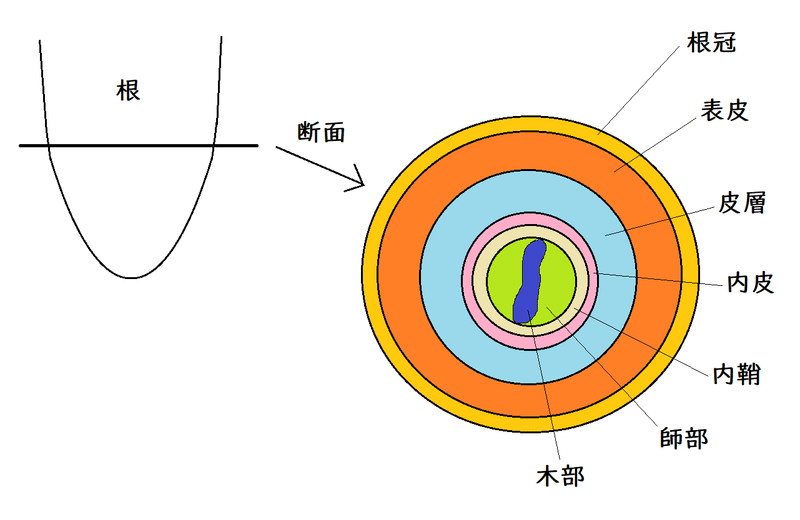
維管束植物の根の断面を見ると、外側から、表皮・皮層・内皮などの構造が、同心円状に配置されている。このような根の放射軸に沿ったパターンは、根端分裂組織が生み出す。
根の根端分裂組織の中心部には、ほとんど細胞分裂を行わない領域(静止中心:周りの組織と比較して細胞分裂の頻度が比較的低いことからその名が付いた)が存在する。近年の分子生物学的研究から、静止中心の機能が明らかになりつつある。
シロイヌナズナでは、静止中心は4個の細胞から構成されている。また、静止中心は、様々な組織を生み出す始原細胞に囲まれている(下図には描いていない)。静止中心をレーザーで破壊すると、静止中心に隣接した始原細胞が(異常な分裂を行い)早期に分化したことから、静止中心には「始原細胞の分化を抑制し、分裂能力を続けさせる」働きがあることが明らかになった。
雑談:以下は維管束植物の根の断面のイメージ。「放射状に色々な構造が配置されているなあ」と思っておけばOK。現在、このような放射パターンの形成にどのような遺伝子が関係しているのかについての研究が進んでいる(未だ完全には解明されていない)。
要点:オーキシンによって側芽の成長が抑制される(頂芽優勢)。
● オーキシンは、屈性に関わる(細胞伸長を促進する)だけでなく、以下のようなはたらきもあることがわかっている(まずは①頂芽優勢だけしっかりチェックすればよい)。
①頂芽優勢(ちょうがゆうせい):頂芽で合成されたオーキシンによって、側芽の成長が抑制される現象。頂芽で合成されたオーキシンが、茎を下降し、側芽でのサイトカイニン(側芽の成長を促進する植物ホルモン。次の資料で学ぶ)の合成を抑制する→側芽の成長が抑えられる。
頂芽を除くと、側芽が発育を始めることが多い。
*頂芽優勢:茎に頂芽と側芽とが共存する場合、頂芽はよく発育するが、側芽は発育しにくくなる現象。

② 不定根の形成:根の組織以外の細胞から形成される根を不定根といい、オーキシンは不定根の形成を促進する(カルスを高濃度オーキシン条件の培地で培養すると、根が形成される。カルスについては次の資料で学ぶ)。
③ 落葉・落果の防止:落葉や落果は、離層とよばれる特別な細胞層ができて起こる。オーキシンは離層の形成を抑制する。
④ 子房・果実の成長促進
講義動画【オーキシンと光屈性】
発展:オーキシンの極性移動について
細胞壁(細胞外)のpHは5付近に保たれており、インドール酢酸(IAAと略される。植物界に広く存在する天然のオーキシン)は、 Hとくっ付いたIAAHと負にイオン化したIAAーの形のものが約半々に存在している。IAAHはそのままリン脂質二重層を横断する。負にイオン化した IAAーは、取り込み共輸送体により細胞内に入る (細胞壁はH+-ATPaseの働きによって酸性に保たれている。そして、オーキシン取り込み輸送体[AUX]は、H+が細胞内に流れ込む勢いを使ってオーキシンを細胞内に輸送する)。
IAAHは細胞内(pH7付近)ではIAAーになり、細胞膜を通過することができなくなる(一般に電荷をもつものは、何らかの仕組み無しでは細胞膜を自由には通れない)。つまり、IAAHは細胞内には自由に入れるが、入ってイオン化したら、もう細胞から勝手に出ることができない。そこで、細胞膜上にはオーキシン排出輸送体(PIN)が設置されている。そのオーキシン排出輸送体が、細胞膜の特定の方向の面にしかないので、極性移動が可能となる。
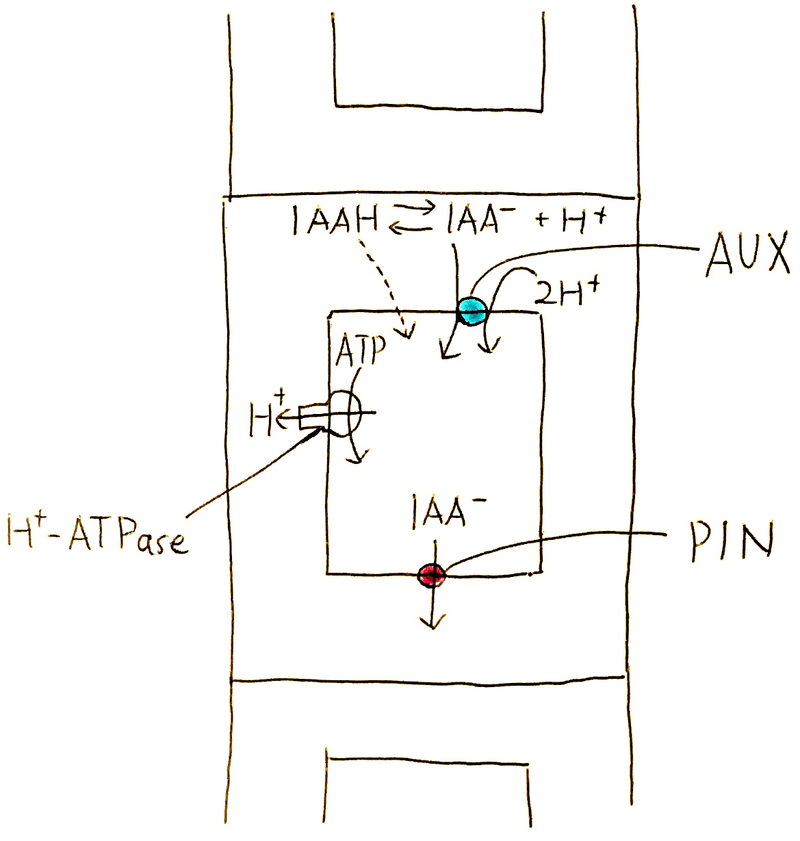
講義動画【オーキシンの極性移動】
雑談:植物体内には、水や養分を輸送する道管、師管のほかに、オーキシンを輸送する通路が張り巡らされていることになる(オーキシンは主に木部柔細胞を通ると考えられている)。
雑談:根の先端まで移動したオーキシンは、折り返す。そして、根の先端から基部方向へ移動したあと、ふたたびUターンすると考えられている(つまりぐるぐる循環していることになる)。このオーキシンの移動は、PIN1~7といった、複数のオーキシン排出輸送体が、決まった側の細胞膜へ設置されていることによって起こっていると考えられている。このような説をオーキシン還流ループモデルという(まだ完全には解明されていない)。
雑談:オーキシンと光屈性に関する説は、ここまで説明したような説が非常に有力であるが、光屈性は成長『阻害』物質の不均等分布によるとする説(ブルインスマ・長谷川説)も提唱されている。いずれにしろ、オーキシンの生合成、光感受、オーキシンの輸送(特に横輸送)については、まだわかっていないことも多い。
発展:オーキシンについての研究
(マカラスムギの幼葉鞘を用いた研究によって、オーキシンについての研究が発展していった)
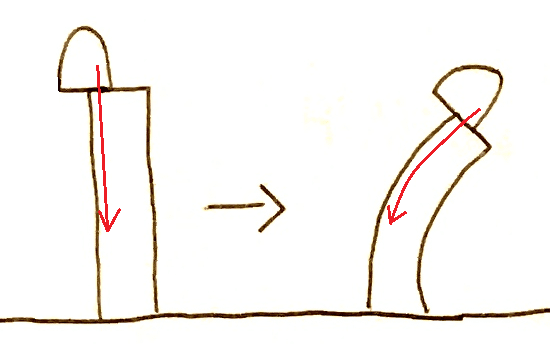
①ダーウィン父子(進化論で有名なダーウィンと、その息子)は、「光を感知するのは幼葉鞘の先端である」、「先端で感知された情報は下方に伝えられて屈曲する」ことを示した。
*先端を切り落としたり、光を通さないキャップを先端にかぶせると、屈曲しない。

②ボイセン・イエンセンは、「幼葉鞘の先端でつくられる成長を促進する物質は光の当たらない側に移動すること」「その物質はゼラチンを通過するが(鉱物の)雲母のような水を通さない障害は通過しない」ことを示した。
③パールは「幼葉鞘の先端でつくられる成長を促進する物質は化学物質である」ということを示した。屈曲は光の刺激がなくても起こった(成長を促進する化学物質は光によって作られるのではない)。
語呂「パルイイイイン(パリーン[パール]、と割れる音で、幼葉鞘が割れている上の図をイメージする)」
④ウェントは「幼葉鞘の先端部には成長促進物質が存在する」ということを示した。
発展:ウェントは活性な成長促進物質が寒天のブロック中に拡散していくことを示した。彼はまた、オーキシンを定量するために、幼葉鞘屈曲テスト(アベナテスト)を考案した(アベナはマカラスムギの学名に由来する)。オーキシンの量が多いほど、よく屈曲する(オーキシンを含ませた寒天片をのせ、どの程度屈曲するか[屈曲する角度]を測定する)。
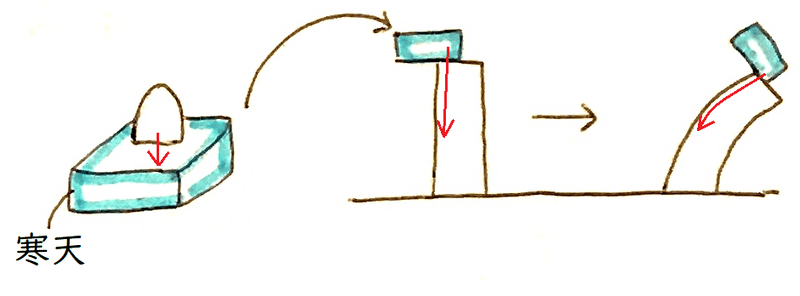
雑談:多くの植物学者は、幼葉鞘の先端をすりつぶして成長促進物質を単離・同定しようとしていた。しかし、組織をすりつぶすことで、細胞内に存在する様々な阻害物質も抽出されてしまい、どの研究もうまくいかなかった。ウェントが成功したのは、幼葉鞘の先端をすりつぶすことなく、寒天に成長促進物質を拡散させたからであった。この寒天に成長促進物質を入れてみせた実験は歴史的な実験であり、当時、ウェントはまだ大学院生であった。
雑談:ウェントはまた、オーキシンを集めた寒天を加熱しても、有効性が失われないことも確かめた。これにより、オーキシンがタンパク質でないことが確認できる。
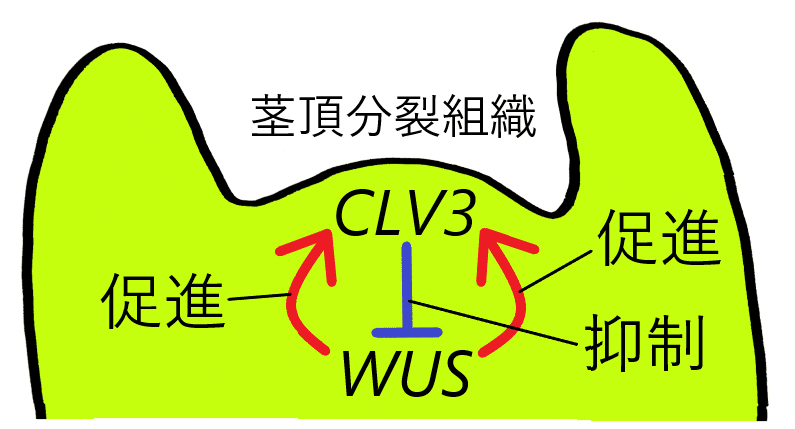
雑談:茎頂分裂組織の大きさを維持するしくみ
WUSという遺伝子は、茎頂分裂組織において、幹細胞の維持に働いている。実際、WUSを欠損した変異体では、茎頂分裂組織が委縮する。また、WUSは、CLV3という遺伝子の発現を促進している(下図赤)。CLV3は、WUSの発現を抑制する(下図青)。実際、CLV3を過剰発現させると、WUSの転写が抑制され、分裂組織が失われる。WUS自身が、CLV3を介した抑制の標的なので、「何やっとんねん」と思うかもしれないが、このフィードバックのしくみ(WUSによるCLV3の活性化、CLV3によるWUSの抑制)によって、WUSタンパク質レベルが安定化し、分裂組織のサイズが安定する(茎頂分裂組織のサイズが、一生を通して維持される)と考えられている。
まだわかっていないこと
● 光屈性誘導機構は、植物種を超えてよく保存されているようである。その進化的な起源はどこにあるか。
● オーキシン生合成・光感受・オーキシンの横輸送(オーキシンの横方向の再分配)の仕組みについて、詳細が明らかになっていない。