
【高校生物】動物生理②「神経細胞はどのような働きをもつのか?」

~プロローグ~
「わたしは考える、ゆえに、わたしは存在する。」デカルト『方法序説』より
脳の認知機能は、いったいどのように生み出されてるのだろうか。情報は、どのように知覚・統合され、どのような内的表現となり、どのように行動として出力されるのであろうか。複雑な記憶はどのように形成され、思考・感情は、どのように言語化されるのであろうか。神経科学は、そのような科学の究極の疑問に挑んでいると言えよう。
★テストに出やすいワード
①静止電位
②ナトリウムポンプ
③活動電位
④全か無かの法則
⑤閾値
要点:活動電位はナトリウムイオンの流入により生じる。
(1)静止電位(せいしでんい)
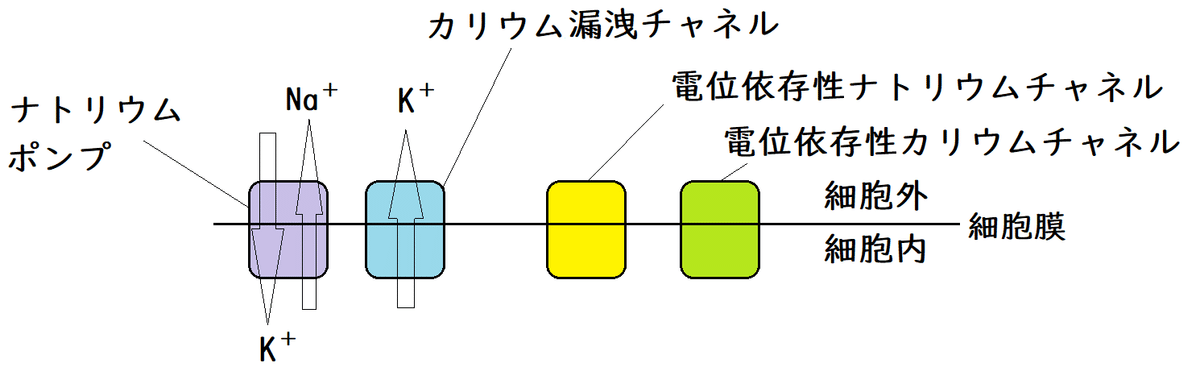
● ナトリウムポンプは、ATPを分解して得たエネルギーを用いて、細胞外にNa+を、細胞内にK+を能動輸送する(ATPのエネルギーを用いて3つのNa+を細胞外へ排出し、2つのK+を細胞内に入れる)。
*特に問題文で指定が無ければナトリウムポンプとナトリウム-カリウムATPアーゼは同義としてよい。
● 常時、カリウムチャネル(カリウム漏洩[ろうえい]チャネル[カリウムリークチャネル]とよばれる種類のカリウムチャネル)がK+(正に帯電している)を細胞外に漏らしている。
よって、静止時、細胞外は正(+)に、細胞内は(細胞外に比べて相対的に)負(-)に帯電している。
このときの膜内外の電位差を静止電位(せいしでんい)という。

語呂「静かでうまい(静止時は う ちがわが マイ ナスになっている)」
*静止電位:静止状態にある神経細胞の膜電位(膜をはさんだ電位『差』)。細胞の種類によって異なるが、一般的にー40~ー80mV(ミリボルト)の範囲にある(アホらしいが、数値が問われることもある。今使っている教科書の静止電位の値を一応チェックしておこう)。
<Q.どうして陽イオンしかないのに「負」なんて言葉が登場するの?…まず、細胞には陰イオン(Cl-)もある。授業で「静止時、細胞内の電位は負(マイナス)になる」と説明されたかもしれないが、その意味は、静止時、カリウムイオン流出の分だけ細胞内の陽イオンが少なくなり(そして、細胞外の陽イオンが多くなり)、「細胞外と比べれば」細胞内の電位が負になる(細胞内より細胞外の方が陽イオンがいっぱいある)ということである(電位という概念は、どこを基準にとるかで値が異なる。細胞外を基準(0mV)とすれば、細胞内の電位は負になる[細胞内を基準とすれば、細胞外は正になる])。このように、静止時においては、膜の外側で陽イオンが過剰になっている(高校生は気にしなくてよいが、膜の内側では陰イオン[Cl−]が過剰になっている)。この細胞膜を挟んだ電荷の分離(膜の外側で正電荷が過剰、膜の内側で負電荷が過剰)が、膜内外の電位差(膜電位)を生じさせている(近接し、相殺されていた正の電荷と負の電荷を引き離すと、それらの電荷の間には、電位差が生じる)。>
雑談:微小な電極を使うことで、静止電位を記録することができる。基準となる電極と、もう一方の電極を、両方とも細胞の外に置いた場合は、電位差は記録されない。しかし、基準となる電極を細胞の外に置き、もう一方の電極を細胞に挿入すると、オシロスコープに静止電位が表示される(下図はイメージ)。多くの神経細胞の静止電位はー65[mV]程度である。


雑談:空間内のある点の電位は、基準点(通常は無限遠にとる)から単位点電荷(1C)を考えている点までゆっくりと運ぶ時に電場がなす仕事と定義される。まあ、高校生物では、電位の正確な定義は気にしなくてよい。電位は「高さ」のようなものと考えておけばよい。細胞外を基準にとり、測定電極を細胞内に挿入すると、静止時、細胞内の電位は負に測定される(細胞外を基準とすると、測定電極が存在する細胞内の電位は負)。これは、観察する人が細胞外にいて、細胞内を見ている状況に似ている。たとえるなら、細胞外は2階で、細胞内は1階である。2階から1階を見れば「低い」ので、細胞外を基準(自分がいる地面のようなもの)とすると、細胞内の電位は負(地面よりも下)に測定される。もし、測定電極と、基準となる電極を、どちらも細胞外に置けば、電位差は記録されない(2階から見て2階は高くも低くもない)。
(2)活動電位(かつどうでんい)
● 細胞内の電位が正の値側に変化する現象を脱分極(だつぶんきょく)という。逆に、細胞内の電位が負の値側に変化する現象を過分極(かぶんきょく)という。
● ニューロンが刺激を受けると電位依存性(でんいいぞんせい)ナトリウムチャネルが開いてNa+が細胞内に流入し、細胞内の電位が、一時的に、マイナスの静止電位から+40mVに上昇する。
この電位『変化』を活動電位(かつどうでんい)という。

*この状態は1/1000秒(1ミリ秒)程度の非常に短い時間で終わる。

● 電位依存性ナトリウムチャネルはすぐに閉じ、不応期と呼ばれる時期に入り、しばらく反応しない。
● 続いて、遅れて開く電位依存性カリウムチャネルが開いてK+が細胞外へ流出し、電位がもとにもどる(どうして電位依存性カリウムチャネル遅れて開くかはよくわかっていない)。

そして電位依存性カリウムチャネルが閉じて、元の状態(静止状態)に戻る。
*ナトリウムポンプはずっと働いている(ATPのエネルギーを用いて3つのNa+を細胞外へ排出し、2つのK+を細胞内に入れ続ける)。
*カリウム漏洩チャネルはずっと開きっぱなしである(実際はランダムに開閉を繰り返している)。このカリウム漏洩チャネルからのK+の流出が静止電位を実現している。
*すべてのチャネルは受動輸送しか行わないことに注意。たとえば、興奮時、電位依存性ナトリウムチャネルを通って細胞内にNa+が流入するのは、細胞外より細胞内の方がNa+の濃度が低いからである(濃度勾配に従ってNa+が移動する)。
雑談:ナトリウムポンプが輸送する+イオンの数の差(Na+が3個、K+
+が2個)が静止電位をつくる原因であると勘違いしている人もいるが、誤りである。ポンプによって輸送される+イオンの数の差は静止電位の形成にはほぼ寄与しないことが知られている(ナトリウムポンプは3Na+に対して2K+を輸送する。よって、確かに、このポンプが働くと電流が流れることになる。しかし、ナトリウムポンプを流れる電流はイオンチャネルを流れる電流に比べるとわずかであり、静止膜電位をたかだか数mVほど変化させる程度の大きさである)。K+漏洩チャネルによるK+の細胞外への流出こそが静止電位形成の主な原因である(細胞膜のもつ、K+チャネルの選択的透過性が、静止電位を作り出すカギとなる決定的な要素であることが明らかになっている)。
下図はイメージ。

語呂「かつ丼に納豆流入(活動電位、ナトリウムイオンの流入)」
雑談:フグの毒の成分であるテトロドトキシンは、電位依存性ナトリウムチャネルの強力な阻害剤である。中毒死は呼吸麻痺による(テトロドトキシンは当初はフグの代謝生産物と考えられていたが、細菌の生産する毒素をフグが餌を通じて蓄えているものと判明した)。
雑談:K+は、開きっぱなしのカリウム漏洩チャネルを通って外に出る(K+の拡散)。そのために細胞外は細胞内より+に偏り静止電位が生じる。しかし、K+がどこまでも流出してしまうということはない。なぜならば、K+の流出は、細胞外を+に(細胞内をーに)偏らせ、+の電荷をもつK+の細胞外への流出を阻止しようとする方向に働くからである(K+は電気的な力によって細胞内に引き戻されようとする)。それで、丁度良い電位差になって変化が止まる(K+の濃度勾配はK+を細胞外に流出させようとする[拡散]が、膜内外の電位差は逆にK+を細胞内に流入させようとする。これらの2つの勢いが等しくなって変化が止まる。すなわち、濃度勾配によって駆動されるK+の細胞外への流出が、膜内外の電位差によって駆動されるK+の細胞内への流入と等しくなる。なお、Clーも同様にして膜内外で平衡状態に達するが、負電荷をもつClーはほとんどが膜電位によって細胞外に保持される)。
このような、細胞膜を横切る正味のイオンの流れがない平衡状態を想定し、細胞の静止電位と定義している。静止電位を主として決定するのはK+の勾配であることがわかっている(動物細胞の静止電位[細胞外を基準に細胞内の電位を測定した場合]はー20~ー120mVであり、生物や細胞の種類によって異なる)。

雑談:カリウムチャネルには様々な種類が発見されている。サソリの毒の中には、ある種のカリウムチャネルの小さな穴の所に結合し、ブロックしてしまうものがある(その結果、犠牲者は死に至る)。
雑談:我々が正常に歯医者の治療を受けることができるのは、局所麻酔薬のおかげである(歯医者が歯に穴をあけても、我々はほぼ何も感じない)。局所麻酔薬は、電位依存性ナトリウムチャネルに結合することによって、活動電位の発生を阻止している。
(3)伝導と不応期
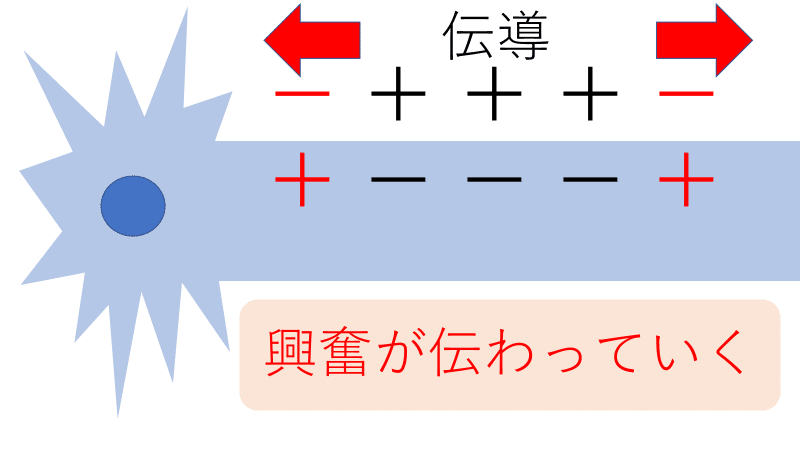
● 軸索の途中に刺激を与え、興奮させたとする。刺激を受けた部分でナトリウムイオンが流入し、活動電位が発生する。すると、その電位の変化がきっかけとなって、興奮部の両隣でもナトリウムイオンが流入し、活動電位が発生する(興奮した部分は、自身の両隣も興奮させようとする)。
下図はそのイメージ(髄鞘や跳躍伝導については考えていない。跳躍伝導については次の「動物生理③」で解説する)。
● 一度活動電位が発生したところは、不応期(ふおうき)に入り、しばらく活動電位が発生しなくなる。このようにして、刺激を受けた場所から両方向に興奮が伝わっていく。これを興奮の伝導(でんどう)という。

*下図は伝導の大雑把なイメージ(軸索の中央に刺激を与えた場合のイメージ)。

● 伝導のイメージ
①静止時は細胞の内側がー、細胞の外側が+。閾値以上の強さの刺激を軸索の途中に加えると、興奮が発生する(膜内外の+、ーが逆転する)。
*閾値:興奮を発生させる最小の刺激の強さ。


②興奮は両方向に伝わっていく(伝導)。

発展:興奮が伝わる方向
上の図では軸索の途中を人為的に刺激して興奮させているが、通常、はじめに興奮が伝わり活動電位が発生するのは、細胞体のすぐ近くの軸索の領域である。不応期があることで、細胞体側で発生した活動電位が、軸索末端の方向に一方向的に伝わることになる。下図はそのイメージ(直前に興奮していた場所は不応期に入っているから、細胞体→軸索末端の方向のみに興奮が伝わっていく)。

雑談:不応期があるため、たとえば、軸索の端の2か所を人工的に刺激した場合に、2つの興奮が軸索上ですれ違うことは無い。2つの興奮が出会った場所で、興奮は2つとも消失する(一方の興奮が進もうとする場所は、もう一つの興奮が終わって不応期に入っているから)。
<Q.伝導では興奮は両方向に伝わるって習ったのに、どうして上の図では一方向に伝わっているの?…伝導において興奮が両方向に伝わるのは、軸索の途中を"人為的に"刺激した場合である。通常、活動電位は、軸索の途中ではなく、まず、細胞体のすぐ近くの軸索の領域(軸索初節という領域)で発生する。そして、活動電位はそこから軸索末端方向へ一方向的に伝わっていく。>
雑談:多くのニューロンでは、通常、活動電位は軸索初節という、細胞体に最も近い軸索の領域から発生する(下図左の矢印の領域。樹状突起や細胞体が脱分極すると、この領域で活動電位が発生する)。感覚ニューロンでは、感覚神経終末で発生する(下図右の矢印の領域。感覚刺激によってまずこの領域で脱分極が生じ、感覚神経を伝導する活動電位が発生する)。赤く色が付いている領域は、電位依存性ナトリウムチャネルの密度が高い領域。

発展:活動電流
活動電位が生じる結果、活動電流が流れる(電荷が運ばれる)。
*活動電流:神経あるいは筋細胞において、活動電位が生じる結果流れる電流。活動電流が流れる理由は、興奮部が静止部に対して膜内部が正(+)の方向に分極していることによるものである。
活動電流は、細胞の外側では静止部(+)から興奮部(ー)へ、細胞の内側では興奮部(+)から静止部(ー)へ流れる(+からーへ流れる)。このとき、電荷を運んでいるのは、電荷を帯びたイオンである(金属のように自由電子e-が電荷を運ぶわけではない。陽イオンがプラスの電荷を運んでいる。陽イオンの流れがすなわち電流となる)。下図は軸索に流れる活動電流のイメージ(興奮部では、細胞内へのNa+の流入が起こっている。すなわち、興奮部では細胞内へNa+が吸い込まれている。①の電流は、その吸い込み口へ陽イオンを供給していると見なすことができる。一方、②の電流は、興奮部ー静止部の電位差によって生じる陽イオンの移動である)。

雑談:詳しくは大学で学んでほしいが、興奮部と静止部において、下図1のような回路が形成されると考えられている(上の図の右半分だけを見ている。Aが上の図の①で、Cが上の図の②である)。
A:Bへの陽イオンの供給。
B:細胞外から細胞内へのNa+の流入。
C:興奮部ー静止部の電位差によって生じる陽イオンの移動。
D:下図2を参照。細胞内側の膜に陽イオンが移動し、電気的に中和する。その結果、細胞外側の膜に引き付けられていた陽イオンが解き放たれる(静止時では、細胞膜の内側に陰イオン、外側に陽イオンがあり、引き付け合っている)。全体としてみれば、Dの向きに陽イオンが流れた(電流が流れた)と見なすことができる。


雑談:軸索は、低温下で興奮をゆっくり伝えるので、痛みは氷によって軽減されるとされる。
講義動画【伝導・伝達】
講義動画【膜電位の測定】
講義動画【静止電位】
講義動画【活動電位】
雑談:以下は静止状態のイオン分布(イカの巨大軸索の場合)。細胞内にK+が多いこと、細胞外にNa+が多いことをチェックせよ。クロライドイオン(Clー)は気にしなくてよい(これだけ見ると、細胞内の電荷が非常にアンバランスに見えるが、細胞内にはClーの他にも陰イオン[主にタンパク質などの高分子]がたくさんあるため、+とーの合計はほぼ等しくなる。細胞外についても、この表に書かれていないイオンを勘定すれば、+とーの合計はほぼ等しくなる)。

<Q.なんで静止時に細胞内が負になってるの?…ナトリウムポンプがNa+を外に、K+を内に輸送し、細胞内のK+の一部がカリウム漏洩チャネルを通って外へ抜けるので、外が内に比べて正に(内が外に比べて負に)なるからである。静止状態を作り出さないと、興奮することができない(適切に刺激に反応するためには、静止と興奮を区別する必要がある)ため、静止時にこのような電位差を生み出していると考えられる(実際、ナトリウムポンプの駆動のために大量のATPを使用している)。>
<Q.どうして細胞外にNa+が多いの?…仕組み的には、ナトリウムポンプがNa+を外に、K+を内に輸送しているから。進化的な意義は不明。Na+は毒性が強いという説もある。>
発展:静止電位・活動電位の大きさ
*静止電位の大きさは、おおよそK+の濃度勾配のみに左右され(K+が細胞外に漏洩する勢いに左右される)、活動電位の大きさはおおよそNa+の濃度勾配のみに左右される(Na+が流入する時の勢いに左右される)としてよい(実際はこれほど単純ではない。細胞内外の様々なイオンや電場に影響される)。
この知識は難問を解く時に必要になる。
下図はイメージ。勾配(膜内外の差)が大きいほど、イオンが移動する勢いが大きくなる。

発展:細胞内のK⁺の半分をNa⁺におきかえて膜電位を測定するとどうなるか?
細胞内のK+の半分をNa+におきかえて膜電位を測定すると・・・
①静止電位は上昇する(0mVに近づく)。
(細胞内のK+濃度が減少し、膜内外のK+の濃度差が小さくなる。K+が細胞外へ漏れ出る勢いによって静止電位が生じるので、静止電位の大きさは小さくなる)
②活動電位の大きさは小さくなる。
(細胞内にNa+が増えるので、Na+の流入する勢いが減る。活動電位の大きさは流入するNa+の量で決まるとしてよい)
下図はイメージ。


発展:膜電位の測定に関する難問対策(基準電極と測定電極の位置を変える)
膜電位を測定する問題では、「どちらの電極が基準となっているか」に注意する必要がある。下の動画を見よ。
講義動画【膜電位の測定(難問対策)】
雑談:上の動画中で「+ーは高さの概念に似ている」と言っているが、当然、電荷と電位を区別する必要がある。本質は、「重力もクーロン力も保存力であり、位置エネルギー(ポテンシャルエネルギー)は常に基準点を決めた上で定められる」ということである。
雑談:ヤリイカは巨大軸索(径約500μm)をもつ。巨大軸索を用いることによって、直接ガラス毛細管電極を挿入して静止電位と活動電位を測定できるようになり、神経生理学が大きく前進した。
雑談:膜電位に関する変化は、細胞膜に極めて近い領域のみで起きている(膜電位を生じさせるイオンは、細胞膜に近い1nm以下の範囲で層をつくっている)。
下図1が「膜電位=0」の状態のイメージ(+とーはそれぞれ陽イオンと陰イオンを表す)。

そして、下図2が「膜電位が生じている時」のイメージ(膜電位を生じさせるイオンは、膜の反対側にある対イオン[反対の電荷をもつイオン]に電気的に引き付けられている)。膜電位は、膜の両側のイオンの数に僅かな差を生じるが、細胞内外の「濃度」にはほとんど影響しない程度の電荷の移動(つまり、膜の近くにおける、ほんのわずかな量の移動)によって形成される(イオンチャネルを通るわずかなイオンの流れが、膜電位に大きな変化をもたらす。たとえば、直径10μmの細胞で膜電位を100mV変えるには、細胞質にあるK+イオン総量の約10万分の1のK+が細胞外に流出するだけでよい。この場合、細胞内外のK+濃度は、実質的に、ほぼ変化しない)。実際、図2のように、静止時の神経細胞の細胞膜の外側表面および内側表面には、それぞれ、陽イオン、陰イオンが広がった薄い雲が形成されていると考えられている。細胞膜の脂質二重層は、イオンの拡散に対する障壁となるため、この電荷の分離が維持されている。この電荷の分離は、膜内外の電位差(膜電位)を生じさせる。

雑談:電気的な信号を用いることで、非常に速い速度で興奮を伝えることができるようになっている。たとえば、足の親指を曲げようと意識してから、"一瞬で"足の親指を曲げ、「あ、曲がった」と認識できる。もし、「何らかの化学物質を脳が分泌して、その化学物質が拡散によって足の親指まで運ばれて・・・」などという仕組みだったら、これほど素早い反応は起こせない。
発展:ネルンストの式・ゴールドマンの式
*ネルンストの式やゴールドマンの式は覚えなくてよい。ほぼ入試にも登場しない。もし式を使う場合は問題文で示されるので、ただ式に値を代入して解けばよい。
● 細胞内外のイオンについての電気化学ポテンシャルが0になる時の膜電位を平衡電位といい、平衡電位はネルンストの式という式で与えられる。
● 膜のチャネルを通るイオンの流れは、そのイオンの電気化学的勾配によって駆動される。
● その勾配には、「膜内外の電位差」と、「膜内外のそのイオンの濃度差」という2つの要素がある(電気的に安定になろうとする勢いと、濃度が均一になろうとする勢いの2つの勢いを考えなければならない)。この2つの要素が釣り合っている場合、電気化学的勾配が0になる(そのイオンがチャネルを通る流れも、見かけ上は0になる)。その時の平衡状態をもたらす電位差(膜電位)が平衡電位である。
● 平衡電位(内側の電位ー外側の電位)をEとすると、Eは以下の式(ネルンストの式)で与えられる。
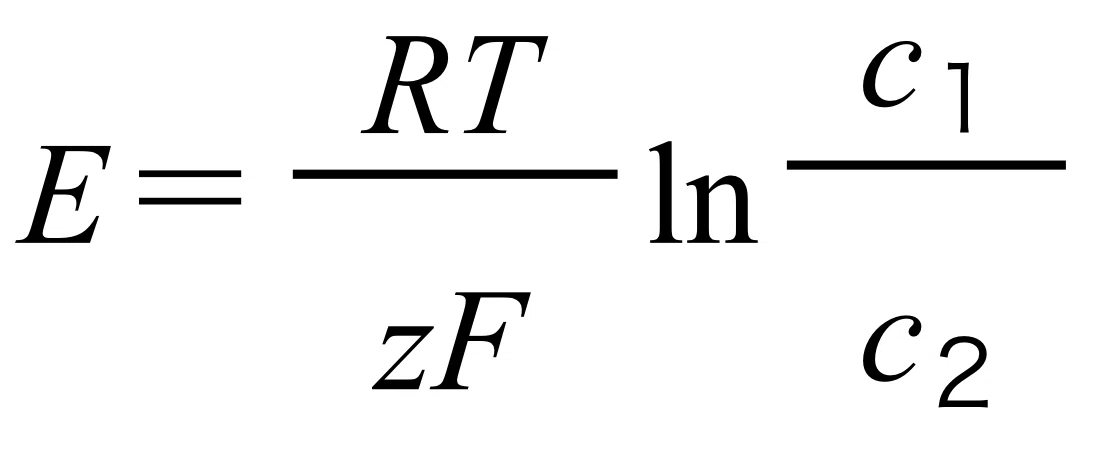
R:気体定数、T:絶対温度、F:ファラデー定数、z:イオン電荷数、
c1:細胞外部のイオン濃度、c2:細胞内部のイオン濃度、ln:eを底とした自然対数
● 上のネルンストの式では、1種類のイオンの平衡電位を考えた。次に、多種のイオンがつくる膜電位について考えよう。
K+、Na+、Clーのみが膜を貫いて移動しているとする。
K+についての平衡電位をEK、
Na+についての平衡電位をENa、
Clーについての平衡電位をEClとすると、
膜電位は、EK、ENa、EClの中間の値になる。
一般に、膜電位が2種以上のイオンによって決定される場合、1種類のイオンの膜電位に対する寄与は、そのイオンの膜に対する「透過性」も関係する。
膜電位Vmとイオンに対する透過性およびイオン濃度との関係は、以下のゴールドマンの式で表される(ゴールドマンの式は、電気化学ポテンシャル勾配に従うイオンの移動によって運ばれる電流の総和が0の時の膜電位の値を示している)。
[ ]oは[ ]iはそれぞれ細胞外、細胞内のイオン濃度を表す。PK、PNa、PClはそれぞれK+、Na+、Clーに対する膜の透過性を表す(単位は速度cm/s)。Rは気体定数、Tは絶対温度、Fはファラデー定数である。

要点:ひとつの神経細胞やひとつの筋細胞は『全か無かの法則』に従う。
● 興奮を引き起こす最小の刺激の強さを閾値(いきち)という。興奮は閾値より弱い刺激ではまったく起こらず、それ以上では刺激の強さに関係なく同じ大きさの興奮(活動電位)が生じる。これを全か無かの法則(ぜんかむかのほうそく)という。
雑談:教室の電気も全か無かの法則に従っていると考えることができる。やさ~しく電気のスイッチを触っても電気はつかない。だんだんスイッチを押す力を強くしていくと(刺激を強くしていくと)、ある強さ(閾値)に達した時、スイッチがカチッと付いて、教室が明るくなる(興奮が生じる)。たとえ閾値以上の強さで、ぎゅ~~っとスイッチを押しても、同じように電気が付くだけで、別に教室がめちゃめちゃ明るくなるわけではない(半分冗談だが、全か無かの法則は、この電気のスイッチの法則のように、「刺激の強弱によって反応が起こるか起こらないかの2通りの反応しかあり得ず、刺激を加減することによって反応の度合いに大小を生ずることはできないという法則」である)。下図はイメージ。

● 1つの神経細胞(や、1つの筋細胞)は全か無かの法則に従う。下図はイメージ。


● 一方、神経細胞(や筋細胞)の集合体である神経(や筋肉)では、各細胞の閾値がそれぞれ異なるため、この法則は成り立たない。また、個々の神経細胞の活動電位発生の頻度は、刺激が強いほど高くなる。
● 刺激の強さは、活動電位の大きさではなく、神経細胞1つ1つの『活動電位発生の頻度』および『興奮する神経細胞の数』で脳へ伝えられる。(非常によく問われる)
*「神経」は多数の神経細胞の集合体(筋肉は多数の筋細胞の集合体)。細胞ごとに閾値が異なる。下図はイメージ。

*1つの神経細胞を刺激した場合、刺激が強い方が活動電位の発生頻度が大きくなる。下図はイメージ(1つの神経細胞について見ている)。


問題:ニューロンにおける全か無化の法則について解説した以下の文章の空欄を埋めよ。なお、すべての空欄には同じ語が入る。
「( )未満の刺激では興奮が生じず、( )以上の刺激で初めて興奮が生じるが、( )以上であれば、刺激の大きさに関わらず、生じる興奮の大きさは一定であるという法則。」
答え:閾値
講義動画【ポンプやチャネルの復習】
雑談:イタリアの医学者ガルヴァーニ(ガルバーニ)は、解剖したカエルの足の筋肉が、金属に触れてけいれんするのを偶然発見した(ガルヴァ―二は、鉄製の手すりにかけた真ちゅうのかぎ針にカエルをつりさげていた。カエルが手すりに接すると、カエルの筋肉が収縮した)。ガルヴァーニは、観察したこの現象の原因を「筋肉中の動物電気のため(すなわち、電気は、筋肉自体が発生したものである)」と考えた(この考えは誤りであった)。対してボルタ(イタリアの物理学者。彼の名に由来する「ボルト」は現在の電位差[電圧]の単位となっている)は、この現象を、生物体内の電気の放電ではなく、「2種の金属をつなぐことによって発生する電流による」ものと考えた(この考えは正しかった。実際、2種の金属を塩水に浸すと、これらの間に電流が流れた)。ボルタはその後、ボルタの電堆(電池の原型)を発明した(ガルヴァーニは誤った結論を導いたが、彼の研究は、生体の電気現象の研究や、電気学の発展、電池の発明に大きく貢献した。彼がカエルの筋肉のけいれんを発見していなかったら、電池の発明はもっとずっと遅れていただろう)。
まだわかっていないこと
● どのようなシグナルによって生体内における神経細胞の配置が決定されるか。
● 電位依存性ナトリウムチャネルや電位依存性カリウムチャネルの構造・機構・進化の道筋について、完全には明らかになっていない。
● カリウムチャネルにたくさんの種類があるのはなぜか(細かく見ていくと、非常に多くの種類がある)。
● 神経細胞1つ1つの閾値の違いは、何が決めているのか。
● 電気的に興奮しない細胞が電位依存性のイオンチャネルをもっているのはなぜか。
● ヒトの学習、運動、思考、記憶の細胞学的な知見と、あなたの経験を結び付けるにはどのような研究が必要か?脳→神経→神経細胞→チャネルやポンプと、小さい世界に降りれば降りるほど、神経科学に関する知見が深まっていくように思える。
しかし、神経細胞におけるイオンの移動から、あなたがいつ、誰に恋するかが説明できるだろうか?あなたがどんな映画が好きかを説明できるだろうか?マクロな精神活動とミクロな細胞学的活動を、どのような研究によって繋げることができるのか。
● どの程度高等な生物が、複雑な精神をもっているのか(精神活動の本質は何であろうか)。アメーバに心はあるか。