
海鳥の帰巣能力

----------------------
※
月刊海洋 2016年9月号『大水薙鳥−外洋性海鳥の研究最前線−(上)』に寄稿した内容を転載しています(出版社の方からも許可をもらっています)
----------------------
繁殖期のオオミズナギドリは, 自分自身とヒナへのエネルギー供給のため, 営巣地から数 10〜数 100 km離れた海域まで餌を獲りにいっては巣に戻るという移動を繰り返している. 本章では, このような行動に関わる認知能力についてこれまでの研究でわかったこと, そしていまだ解明されていないことについて紹介する.
1. はじめに
オオミズナギドリ(Calonectris leucomelas)を含むほとんどの海鳥は, その繁殖期間中, 巣と外洋の餌場との往復を繰り返す(Gaston, 2004;「採餌トリップ」と呼ばれる). 親鳥は抱卵交代もしくは雛への給餌のため, 定期的に巣に戻らなければならず, このことが餌獲りに使える時間や餌場として利用できる海域を制限している(cf. Orians & Pearson, 1979).
それでも, 数100km, 時に1000 kmを超えるような採餌トリップを繰り返す種も珍しくない. このような長距離移動を, 限られた時間の中で, そして目印が乏しい海上で達成するためには, 優れた飛翔能力のみならず, 高い時間・空間認知能力が必要とされるはずである. 繁殖地から餌場までの移動, そして餌場から繁殖地までの帰巣を確実に遂行する能力は繁殖の成功に欠かせない要素であり, 行動生態学における重要な研究テーマと言えるだろう.
本稿ではまず, これまでの私たちの研究から明らかになった, オオミズナギドリの帰巣における時間調節行動について紹介する. そして, 海鳥のナビゲーションメカニズムに関する先行研究をレビューし, 今後の展望を述べる.
2. 時間認知能力(私たちの研究紹介)
時々刻々と変化する環境の中で生きる野生動物は, 繁殖, 採餌, 休息などそれぞれの目的に適した場所と時間を選び, 移動しなければならない.
ヒナを育てているオオミズナギドリは, 日出前に島を出発して海へと餌獲りに向かう. その日のうちに島へ戻ってくることもあれば, 数日間を海で過ごすこともある. しかし, 島へ戻ってくるのは必ず日没後, それも日没から数時間以内に到着することが多い. つまり, 昼間は島にいない. これは, 昼行性の捕食者である猛禽類などを避けるためだと考えられている(吉田, 1981). 島から一斉に出発し, そして再び一斉に帰島する様子は, 繁殖地で容易に観察できる. 一方, 海へ出ている間の個々の親鳥の行動については, 近年導入された動物装着型の記録計(データロガー)によって知見が集められている途上にある.
GPSデータロガーは, 緯度と経度を時刻とともに記録する. 2000年以降, 小型化・軽量化が進み, 飛翔性の鳥類や小型の陸生動物にも装着できるようになった(Steiner et al., 2000; von Hünerbein et al., 2000). これにより, 海鳥の移動経路を高精度に再現することが可能となった(Weimerskirch et al., 2002).
私たちは岩手県釜石市の三貫島および下閉伊郡山田町の船越大島という無人島で繁殖しているオオミズナギドリ 21羽にGPSデータロガー(Technosmart社製, イタリア)を装着し, 採餌トリップ中の移動経路を1分間隔で1週間以上記録した. その結果, 親鳥は島から数10km程度の比較的近い海域を日帰りで利用していることもあれば, 遠く数100kmも隔てた北海道沖にまで餌を獲りに行き, 1週間以上巣を離れていることもあるとわかった(図1).

図1: 親鳥の採餌トリップ中の移動経路
船越大島で繁殖する個体にGPSデータロガーを装着して得られたデータ
このように様々な距離のトリップを織り交ぜて行うことは他種の海鳥でも報告されている. ヒナへの給餌頻度を維持するために近場で繰り返し餌を獲り, そうしてヒナへの投資で衰えた自分自身のコンディションを回復するために, よりよい餌場を求めて遠出をすることが示されている(Weimerskirch et al., 2003; Ochi et al. 2010).
採餌トリップは, 島からの出発・餌場への移動(往路)・採餌・島への移動(復路)・到着, といったイベントがひと繋がりになったものとみなすことができる. 実際のオオミズナギドリの移動過程を見ると, 採餌トリップ中の往路・採餌・復路の3期間が比較的はっきりと分かれていた(図2).
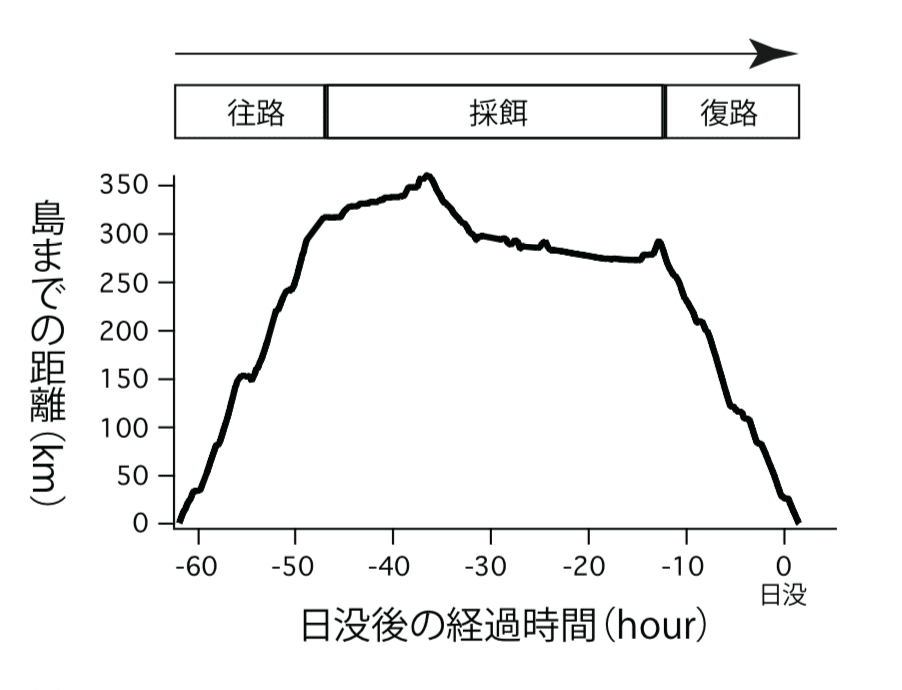
図2: 採餌トリップ中の各地点から島までの距離を時系列で表したグラフ
島から離れていく期間(往路), 距離の変化が小さくなる期間(採餌), 島へと近づいていく期間(復路)に分けることができる
オオミズナギドリの復路開始時刻に注目すると, 日没1時間前のときもあれば, それより20時間近く早いこともあり, 採餌トリップごとに大きく異なっていた. そしてこのばらつきは, 復路の開始地点から島までの距離で説明できることが明らかになった. 餌獲りに出かけた親鳥は, 遠くから戻ってくるときほど, 早い時間帯に帰り始めていたのである(図3a).
復路開始時刻の平均値は, 島までの距離が 1 km長くなると 0.036時間早くなっていた. 一方, 復路における平均移動速度は時速 27.8 kmであり, この値から 1 kmの移動所要時間を算出すると平均 0.036(= 1/27.8)時間となる. つまり, オオミズナギドリは移動に必要な時間が増えた分だけ帰り始める時刻を早めるという調節をしていたのである(図3b). その結果, 復路開始地点から島までの距離によらず, 日没後数時間以内に島に到着することができていた(Shiomi et al., 2012).


図3:(a)採餌トリップの復路開始地点での 島までの距離と時刻の関係(Shiomi et al., 2012の図を改変). 時刻は日没時刻をゼロとした相対時刻に換算している. (b)復路開始時刻の調節について表した概念図.
オオミズナギドリは, 決まった時刻に島に到着しなければならない一方で, なるべく長く餌場に留まろうとしていたように見える. 島への移動を開始するタイミングを場所ごとに調節することによって, 採餌と帰巣, そして捕食者回避という複数の目的を同時に達成していたようだ.
このように柔軟な時間調節は人間以外の動物では初めて報告された行動で, 野生動物の時間・空間認知能力の高さを示唆する結果と言える. 島までの直線距離から算出した所要時間分だけ復路の開始時刻を早めていたという事実は, 少なくとも数 100 km 程度であれば, どれだけ離れようとも帰り道を見つける容易さは変わらないということを示唆しているのではないだろうか.
3. 空間認知能力(レビュー)
それでは, オオミズナギドリは何を手がかりに自身の現在地を知り, 帰るべき方角を決定しているのだろう. 率直に言って, オオミズナギドリのナビゲーションメカニズムについては現時点ではほとんど何もわかっていない. 上述の実験で明らかになった洋上での移動パターンをヒントに, 今後研究を進めていく予定である. そのため本節では, 他種の海鳥のナビゲーションメカニズムについて先行研究で明らかになっていることをレビューする.
3-1. マップ & コンパス仮説
ある場所から目的地へとたどり着くためには, まず現在地と目的地の位置関係を把握し, 次に, 進むべき方角に定位して移動する, という2つのステップを繰り返さなければならない. 私たち人間が, 地図を見て現在地を知り, 方位磁石などで方角を定めるやり方になぞらえ, これらはそれぞれ,「マップステップ」と「コンパスステップ」と 呼ばれている(Kramer, 1953).
また, この2つのステップに加えて, 定位した方角に進行方向を維持するための「第二のコンパスステップ」も必要であると考えられている(Penny & Emlen, 1967; Guilford & Taylor, 2014). リファレンスがない状態で移動方向を一定に保ち続けることは難しく,「第二のコンパス」がなければ, 目指していた方角から徐々にずれていくことが予想されるためである.
それでは, それぞれのステップにおいて海鳥たちはどのような環境情報を手がかりに使っているのか. 実はどの種においても, その問いに明確な答えは得られてはいないのだが, ここではいくつかの有力とされている説について述べる.
3-2. マップ仮説
鳥類のナビゲーションメカニズムに関しては, コンパスステップよりもマップステップの方が物議を醸しているようである. しかし関連論文を概観すると, 海鳥のナビゲーションでは匂い情報が関わっている可能性が最も高く, 地磁気は重要ではない, という考え方が現時点では優勢であるようだ(cf. Bonadonna et al., 2003; Gagliardo, 2013). 海鳥の中でも特にミズナギドリ目では,嗅覚に関わる脳領域(嗅球)が発達していることが知られており(Bang & Cobb, 1968), 匂い情報に頼って帰巣するという説は現実的であると思われる(cf. Bonadonna et al., 2003).
ナビゲーションにどの感覚器が関与しているかを明らかにするため, 特定の器官を機能しないようにする, もしくは感覚を狂わせるような操作をしてから, 巣から離れた場所へ人為的に運び, 放たれた後の帰巣行動を調べるという方法がある. 近年, 嗅神経に麻酔をかけたり, 頭に磁石を取り付けたりした海鳥の放鳥後の帰巣経路を, GPSデータロガーで記録した研究が報告され始めた(e.g. Gagliardo et al., 2013; Polonara et al., 2015). 以前は,放鳥地点から飛び去った方角や巣に戻るまでにかかった時間を測って帰巣能力の指標にするしかなかったため, 高精度な移動経路を連続的に記録できるようになったことは, ナビゲーション研究の分野においても画期的な出来事と言える.
オオミズナギドリと同属のオニミズナギドリ(C. borealis)や Scopoli’s shearwater(C. diomedea)を巣から数100 km離れた場所から放鳥した実験では, 嗅覚を阻害すると帰巣経路が大きく変化した(Gagliardo et al., 2013; Pollonara et al., 2015). それに対して, 磁石をつけた個体の帰巣経路には, 何も感覚操作がされていない対照個体の経路との有意な違いは見出されなかった. この結果は, GPSデータロガーが登場する以前に報告された実験結果とも矛盾しない(e.g. Massa et al., 1991). 以上のことから,嗅覚がナビゲーションに貢献している可能性が高く, 逆に地磁気情報の重要度は低いと考えられている.
海鳥が何の匂いを利用しているかについても研究が進められている. 可能性のある匂い地図としては 2パターンが考えられており, 1つは濃度勾配地図, もう1つはパッチ地図である. 前者では連続的に分布した化学物質の濃度勾配によって位置情報を得る場合, 後者ではパッチ状に分布した匂いを利用する場合を想定している. パッチ地図を形成しうる要素としては, 例えばパッチ状に分布したプランクトン由来の匂いなどが挙げられている(Nevitt, 2008; Reynolds et al., 2015).
オニミズナギドリ(C. borealis), Scopoli’s shearwater(C. diomedea),Cape Verde shearwater(C. edwardsi)の3種では, パッチ状に分布した匂いを探索しながら移動を繰り返して島にたどり着くようなメカニズムを仮定した場合に予想される帰巣経路の性質と, 実際にGPSデータロガーで記録された移動経路の特徴が一致していたことから, パッチ地図を帰巣の手がかりにしている可能性が高いと考えられている(Balkovsky & Shraiman, 2002; Reynolds et al., 2015).
ただし, 匂い情報が使えなくなったら帰巣できなくなるわけではない. 上述のミズナギドリ類では, 嗅覚が阻害された場合には地形などの視覚情報を使っていた可能性が示唆された(Pollonara et al., 2015). 自然界でも常に最良の環境で移動できるとは限らないため, 1種類の情報だけに頼るのではなく, このように「保険」を備えることは合理的と言えるだろう(cf. Walcot, 2005).
3-3. コンパス仮説
いまだ論争が続くマップステップのメカニズムとは対照的に, 鳥類のコンパスステップについては, 基本的には太陽を使って進行方位を定め, 雲の厚い日など太陽の位置がわからない時には地磁気をバックアップ情報として利用しているというメカニズムがほぼ定説として受け入れられている印象を受ける.
時刻情報があれば, 太陽の位置を参照して方位を知ることができる. 人間のように時計という道具を持っているわけではない野生動物は, 体内時計によって知覚される時刻を利用していると考えられている.
この仮説を検証するため, 人為的に明暗リズムを調節した部屋に動物を数日間置き, 体内時計を実際の時刻から遅らせたり早めたりした後, 進行方向がどのように変化するかを調べる実験(クロックシフト実験)がおこなわれてきた. 巣箱から離れた場所で放たれた鳩の飛行方位が, 体内時計のシフトから予想される方角にずれた例が報告されており, 太陽コンパスを使っていることを示す最も強力な証拠とみなされている(e.g. Schmidt-Koenig, 1960). ただし, これらは晴れた日に得られた結果であり, 曇って太陽が隠れている日にはクロックシフト後の飛行方位に変化は見られなかった. このことから, 太陽コンパスが使えない状況では別の環境情報を利用していることがわかる.
しかし, すべての種でこのような検証がされたわけではもちろんなく, 海鳥での実証例はほとんどない. クロックシフト実験による太陽コンパス仮説の検証がおこなわれたのは, 私が調べた限りでは, マンクスミズナギドリ(Pufinus pufinus)とアデリーペンギン(Pygoscelis adeliae)の2種のみである(Matthews, 1955; Emlen & Penney, 1964). そして, 上述のような太陽コンパスのメカニズムを実証する結果は報告されていない.
また, 鳥類が太陽コンパスに加えて地磁気コンパスも使っているという証拠は鳴禽類で数多く報告されている. 実験室で人為的に磁場を様々に変化させた時に鳥がどちらへ向かおうとするかを調べた結果, 磁気ベクトルの向き(磁極)ではなく, 傾き(伏角)によって方位を知覚していることが示された(Wiltschko & Wiltschko, 1972). それを実現する分子メカニズムも提唱されており, 特に右目の網膜上の光受容体が重要な役割を担っていると考えられている(Ritz et al., 2000; Wiltschko et al., 2002).
海鳥においては, 地磁気コンパス仮説をそのような方法で検証した実験や, 仮説を支持する結果は報告されていない. 3-2.に記したように, 磁石を頭に乗せても帰巣行動は大きく変わらなかったことを考えると, コンパスステップにおいても地磁気情報が不可欠というわけではなさそうである.
こうしてレビューしてみると, オオミズナギドリに限らず, 海鳥のナビゲーションメカニズムについてはっきりと解明されていることは多くはないことがわかる. たくさんの研究者が長いあいだ取り組んできたにもかかわらず, いまだに解明されていない謎について, 今後どのようなことが明らかにされていくのか楽しみである.
4. おわりに
私たちのこれまでの研究では, 島を出たオオミズナギドリがどのような経路をたどって再び島へと戻ってくるのか, その空間パターン・時間パターンを, GPSデータロガーを用いて明らかにすることができた. 次なる目標は, 前節でも述べたようにそのメカニズムを明らかにすることである.
野生動物がいつ・どこを通って移動するかは, 現在地や目的地の方角を知るためのナビゲーション能力に加えて, 風向きや地形といった物理環境や同種・他種個体の行動, 自身の生理状態など, 未知の要因が多数関与して決定されているはずである. したがって, それぞれの要素を個別に切り離して調べるだけでなく, 全てが綯い交ぜになった自然環境下での振る舞いから, 彼らにとってどの要素が重要であるのかをあぶり出すステップが不可欠だろう.
しかし海鳥の帰巣行動に関する過去の研究では, 仮説先行型の感覚操作実験が精力的に行われてきた一方で, 自然環境下での行動の分析はほとんどなされてこなかった(Mattern et al., 2013・ Reynolds et al., 2015 などは後者のアプローチ). オオミズナギドリの帰巣行動については, 自然環境下での移動経路の分析がここで紹介した内容以外にも着々と進められている. そのため, 海鳥の帰巣メカニズムについて, これまでにない知見が日本のオオミズナギドリグループから発信される日も近いのではないかと期待している.
この記事が気に入ったらサポートをしてみませんか?