
【2】 掛谷英紀先生とのコロナ人工ウイルス説論文に関する共同研究が決裂した経緯について

今回の記事は結果的に大変長い内容になってしまいましたが、分割するよりも一つにまとめた方が読む方にとっても良いかと考えました。
私は掛谷先生がnoteのアカウントをお持ちと認識していた事もあり、note記事のコメント欄での返答をお願いさせていただきました。また実際Twitter(X)上では書き込みが分散しがちであり、情報を追うのが難しいという理由もありました。けれども、残念ながら質問に対する明確な返答を頂く事はできませんでした。
しかしながら、今回の記事では極力掛谷先生のポストを引用しつつ、改めて私自身の主張の裏付けとさせて頂きたいと思います。またここでは掛谷先生に送った多数のメールの中から5つを私の主張の根拠として引用致します。

「ウソ」というのは非常に強い言葉です。
上が私のプレプリント (以降「荒川論文」とします) 、下が掛谷先生の論文 (以降「掛谷論文」とします) のリンクです。
掛谷先生と知り合った経緯及び共同研究を依頼された件

私は「この記事を見た筑波大学の掛谷英紀先生が2022年1月16日に私にコンタクトを取って来られました。」と書いており「最初のコンタクト」などとはそもそも言っておりません。
2021年の夏頃、とある匿名アカウントの人物がnote上で私を中傷する内容の記事を繰り返し書いていたのですが、知人から掛谷先生がその人物をご存知のようだと教えてもらい、その人物に関して聞くために私の方からメールを送ったのが掛谷先生と知り合った最初の経緯です。その件に絡み掛谷先生とは当時何度かメールのやり取りがありました。しかしながら、その事についてブログ内でお話する必要は無いと私は判断しましたので、最初の記事ではとくに触れてはいませんでした。

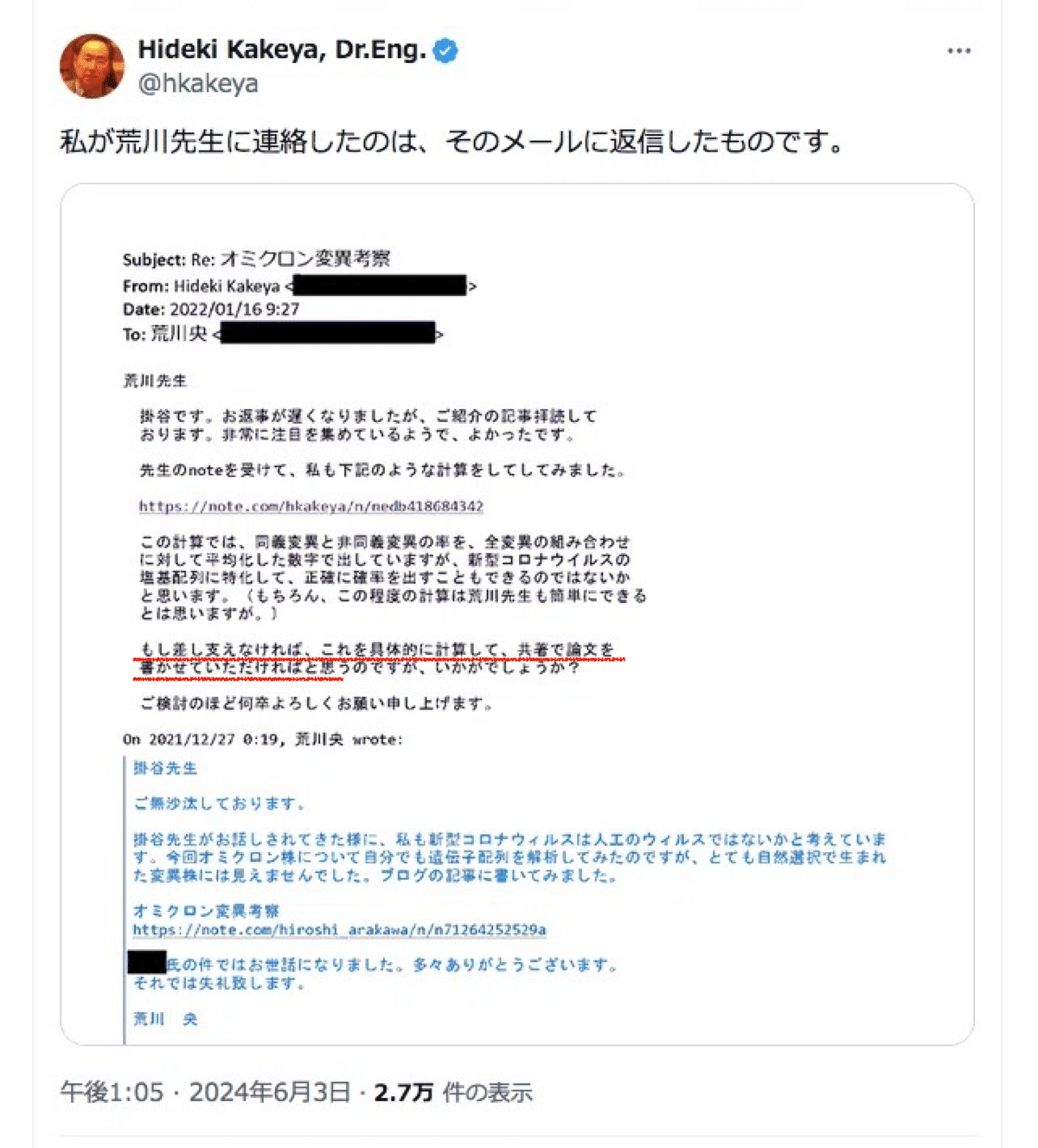
その後約半年後の2021年12月26日、私は自分自身で行った解析を元にしたコロナウイルスのオミクロン株の変異に関する考察についての記事をnoteに投稿しました。そしてその際、掛谷先生に改めて上記の件のお礼と挨拶をかねて記事のリンクを送りました。それが掛谷先生がTwitter(X)上で公開した私からのメールです。
ちなみに私が知人にメール等で記事を知らせる事自体はよくあり、このオミクロン変異考察の記事も同様に数名の知人に記事のリンクを送り、そのうちの一人に含まれていたのが掛谷先生でした。きっかけとしては、当時掛谷先生がコロナウイルス人工説に興味を持たれているのを知っていたという事もあります。

このようにご自身で認められているように、共同研究の申し込みはその記事を読んだ掛谷先生側からの依頼によるものです。

にも関わらず、掛谷先生は以前は「荒川先生と知り合ったのは2022年の1月」と矛盾した事を言われていたのは一体なぜでしょうか。
荒川論文への掛谷先生の貢献について
当時私はちょうどコロナウイルス人工説についての論文 (荒川論文) を執筆中であったため、その依頼を受ける事にし、掛谷先生には共著者とするために論文を補強するアイデアやデータを加えて欲しいとお伝えしました。けれども掛谷先生は、武漢などにおける機能獲得実験を批判する政治的な文章を論文の末尾に加筆されたのみでした。
私信の公開および詐欺雑誌について


改めて確認させていただきますが、今回私は掛谷先生に私信の公開は許可しておりません。それ以前に、著者が論文をどの雑誌に投稿したかというのは研究における大切な内部情報の一つです。そのような情報を当人の許可なく公開する行為は研究者としてのモラルに対する疑問が持たれます。
さて、掛谷先生が私信である私のメールを公開された以上、この件についても私の方から改めて説明せざるを得ないと判断しましたので、ここで詳しく経緯を説明いたします。メール中で触れている「当時投稿中のHeliyon誌」には荒川論文はリジェクトされました。そしてそのリジェクトされた論文を、たまたまそのタイミングで投稿依頼が来た雑誌のThe International Journal of Engineering and Scienceに再投稿しました。それが記事内で触れた「詐欺雑誌」にあたります。実際、論文をリジェクトされた場合、その論文を他の雑誌に再投稿する事は論文投稿における通常のプロセスです。

そしてその雑誌社からのアクセプトのメールが来て一度は喜んだものの、その後論文は一向に掲載されませんでした。
私は雑誌THE IJES社に対して20通以上の抗議のメールを送りましたが、結局なしのつぶてで返信は一切ありませんでした。しかし私は論文の著作権フォームを投稿時に相手に渡してしまっていたため、権利を取り返す事もできず、結果他の雑誌への再投稿もできなくなり、そのまま論文はペンディング状態になり現在に至ります。
掛谷論文採用に対する「おめでとうございます」について


「おめでとうございます!」の一言については、最低限の礼儀からの社交辞令に過ぎません。私は誰かの論文を評価する場合はできるだけ具体的に賞賛します。例えば発見の重要性や洞察力、研究への情熱や努力などです。私はこの件に関してそのような表現をする事は到底できませんでした。
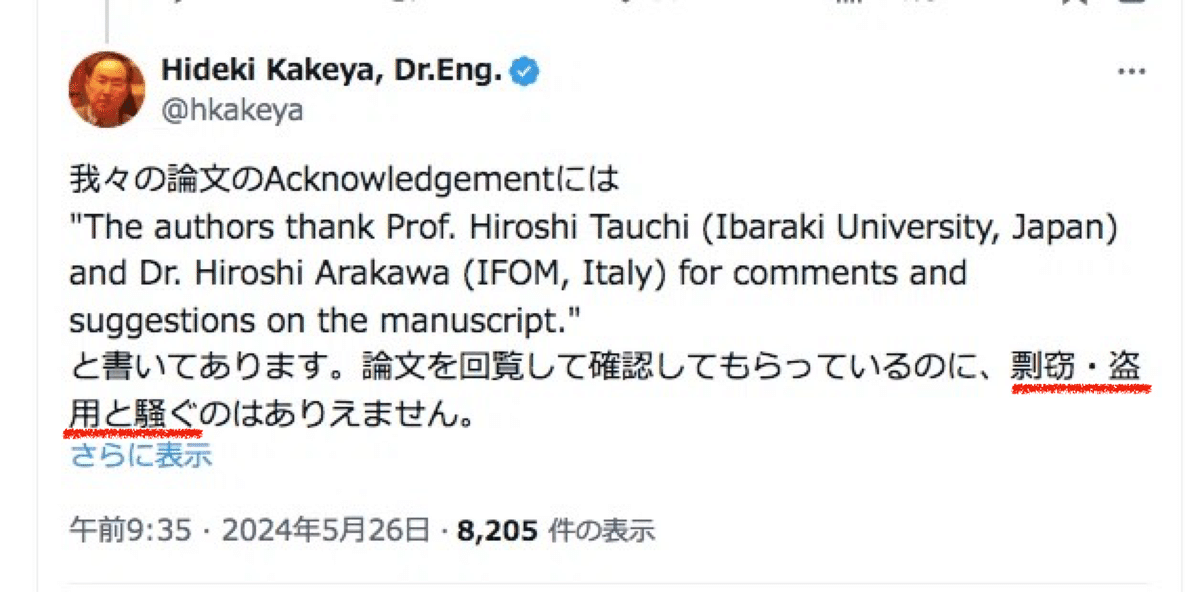
謝辞について
そもそも謝辞 (Acknowledgment) だけから研究者間の関係や背景までを測る事などはできません。それどころか自分に批判的な研究者に対して意図的に論文へのコメントを求めるような場合すらあります。というのも、謝辞に名前が入っている研究者は査読の審査員候補から外されるからです。例えばその相手から「これはナンセンスでどうしようもない論文だ!」といったコメントを貰った場合ですら、「○○氏には論文を批判的に読んでいただき感謝致します」のように謝辞に書く事もできるからです。反対に、査読の審査員として期待する研究者を謝辞に入れると審査員候補からは外れてしまうため、親しい研究者にはあえて論文のコメントを依頼しないというような手段もあります。
いずれにせよ、謝辞において例えば相手に対する失望や怒りなどを書く事はありませんし、実際外からはそうした感情までは判断する事はできません。逆に言うと、どれほど発想やデータで貢献があったとしても謝意を表明していなければ読者には「真の貢献」については分からないのです。

繰り返しますが、謝辞を含め論文をいくら読み込んでも、本来は共著のはずだった論文が決裂した事情など外部からは窺い知る事はできません。実際今回の荒川論文の謝辞に名前がある掛谷先生も、掛谷論文の謝辞に名前がある荒川も、「著者から謝辞への格下げ」という異例の事態が背景に存在しています。両論文の謝辞に名前がある部分について誤解を受ける恐れもあると思いましたので、ここで改めて私の意見を記しておきました。
パリグループ (Evidence-based investigation of SARS-CoV-2 proximal origin) について

2022年2月17日に私が参加した際のパリグループ学会のZoomによるオンライン発表時には、非常に多くの参加者の顔が見え、それが何画面にも渡っていました。ざっと見渡したところ、200名ほどは居たように私には見受けられました。
いずれにせよ、ギフトオーサーシップ自体は研究倫理に抵触する行為ですが、掛谷先生はその学会で「自分の論文に賛同する場合は誰でも著者に加えても良い」とお話しされた件については否定されていません。
掛谷先生から要求されたデータを提供した件について
遺伝子の変異を解析するためにはまずは遺伝子配列が必要となります。そして、複数の遺伝子配列を比較し、その違いとして変異が同定されます。コロナ変異株の祖先型の配列は本来荒川論文のために私が同定したものです。これらが変異解析の元データとなり、荒川論文、共同研究決裂前の掛谷論文の両方に使われました。
私が掛谷先生から要求されたのはこうした遺伝子配列そのものや変異のデータです。データの要求は繰り返し何度もあり、私はその都度要求された通りのデータを提供しました。以下は私からの返信のうちの3つです。
2022年2月13日
掛谷先生
変異株に特異的な変異と、複数の変異株に重複している変異を数えてみました。excelの表にまとめています。わかりにくかったらまた聞いてください。
例えば、オミクロンの非同義変異29のうち、7つは他の変異株でもみられます。オミクロンに特異的な非同義変異は22 (not sharedと表記)、7つは他の変異株のどれかと共通 (sharedと表記、どの変異株に見られるかも記しています)。1つだけある同義変異はオミクロン特異的です。他の変異株でも同様に表から読み取ってください。
7つの変異株の合計で重複無しの非同義変異が52、重複している非同義変異が8です。重複している変異をそれぞれ1回だけ数えると、7つの変異株に独立に入った非同義変異は合計60です。合計2つの同義変異は重複無しです。中立進化及び自然選択が無かったとしてもランダムな変異がこのような非同義変異、同義変異の配分で固定される確率はかなり低くなります。
間違いはないと思うのですが、もし後で数え違いがわかったらまたご連絡します。ありがとうございます。よろしくお願いします。
荒川 央
2022年3月6日
掛谷先生
突然変異を整理して数えてみました。興味深いデータです。
ORF1abは21 kbの大きな遺伝子なのですが、C -> Tに顕著な偏りがあります (DNA解析ソフトを使っているのでUではなくTと表記しています)。スパイクタンパクを除いて他のORFでもC -> Tに偏りがありますが、スパイクタンパクではこの偏りが見られないか小さいです。変異株ごとに見てみると、この傾向はアルファ、ベータ、デルタ、ミューで顕著です。
抗体遺伝子の体細胞突然変異でもC -> T、G -> Aに偏りが見られるものですが、SARS-CoV-2ではG -> Aへの偏りは見られません。これはSARS-CoV-2ゲノムが一本鎖RNAだからかと思われます。コロナウイルスのゲノムは正鎖のRNA (タンパク翻訳用のRNA) です。負鎖はコロナウイルスゲノム合成のための鋳型です。複製には自前のRNA依存性RNA replicaseを用い、正鎖を鋳型にしてその相補鎖である負鎖を合成し、さらに負鎖を鋳型にして正鎖を合成します。おそらく正鎖から負鎖を合成する量は少ないのに対し、負鎖を鋳型にして正鎖を合成する量が多く、RNA複製に伴うC->T変異も増えるはずです。そのため、G -> Aへのバイアスは目立たないのでしょう。
全ての変異株のスパイクタンパクは自然な進化も人工進化も経ていないデザインされた遺伝子ではないでしょうか。その仮説のサポートになると思います。
荒川 央
2022年3月8日
掛谷先生
一昨日送ったexcelのデータのindependent mutationsが「各変異株で共通する変異をダブルカウント」せずに集計したデータに当たります。それぞれの変異 (A→C、A→G、A→T、、、など) の合計も表の右に記載されているので、そこからC→Tの数を引いてください。わかりにくかったらまた聞いてください。
このデータでよろしいでしょうか?よろしくお願い致します。
荒川 央
このように私は何度も掛谷先生からの要求に応じてデータを提供しています。

そして掛谷先生も上記のように私からデータを受け取った事を認めておられます。
しかしながら、掛谷先生のそれらのデータに対する扱いは以下のようなものでした。
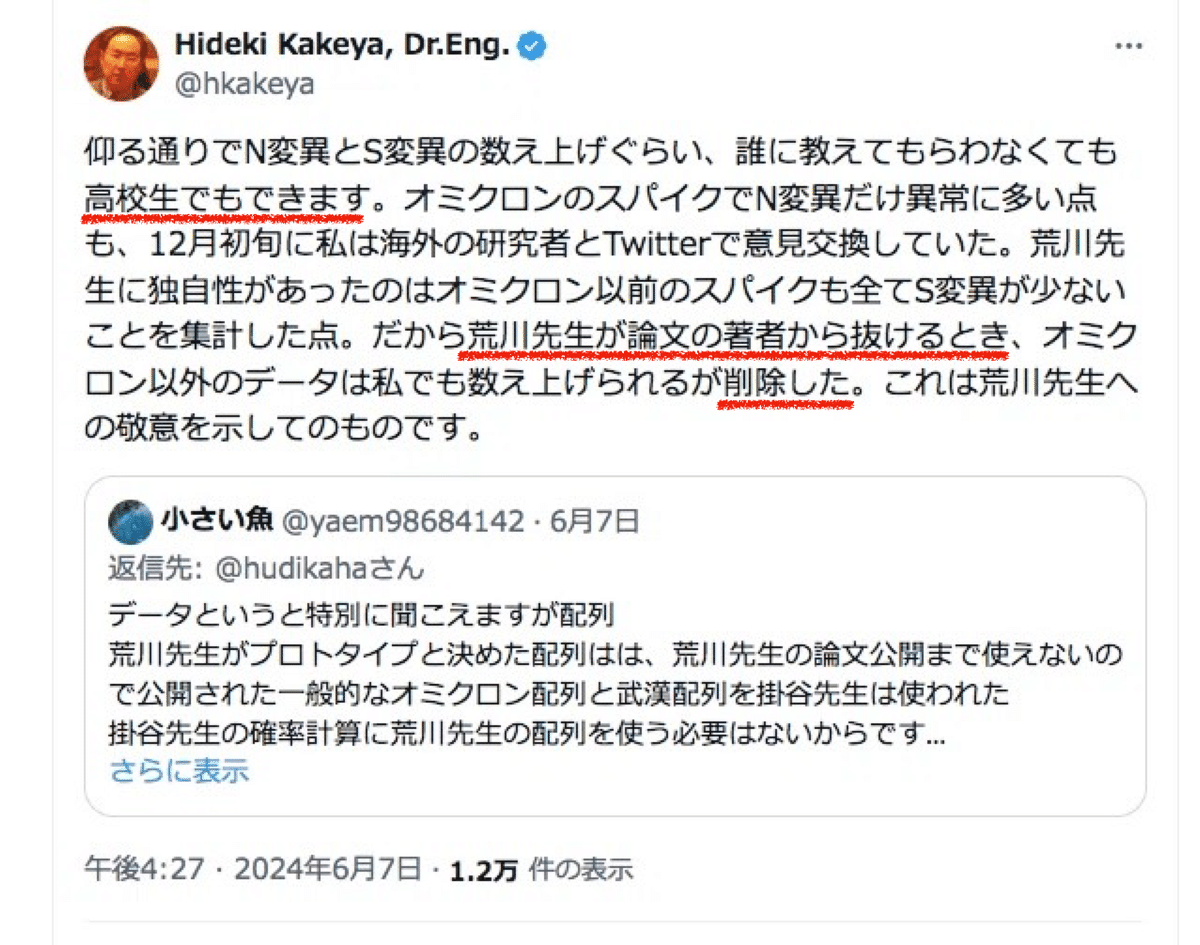
掛谷先生はデータを私に繰り返し要求したにも関わらず、データにも私自身に対してもリスペクトが致命的に欠落していました。研究者にとっては自分の知恵を絞って出したアイデアもデータもいわば宝石の原石のようなものです。その提出されたデータを見るだけ見た後は事実上ゴミ箱に捨てて無かったものにし、それどころか今度はそこから類似のデータを探し出してきて自前のデータだと主張し始めるなど、研究者として相手を蔑ろにした態度どころか侮辱行為とまで受け取られても仕方ありません。


どのようなデータベースに由来する情報であっても解析次第では発見につながる可能性のあるものです。また、論文に発表された情報のメタデータ解析も貴重で重みのある情報です。そうしたものは未発表のうちは公知の情報とは言えません。にも関わらず、それを「公知」であると主張されるのは、公的情報の解析による他者の未発表データは自由に利用しても良いという発言と同義にも受け取れます。
祖先型解析の意味と視点

また、荒川の着眼には「先駆性がない」と掛谷先生はおっしゃいますが、ではなぜわざわざそのような私に共同研究を申し込まれたのでしょうか。
どのような実験手法や解析手法を用いるべきかは目的次第です。荒川論文の目的は、コロナ変異株が人工ウイルスではないかという仮説の検証です。
遺伝子の変異の解析をするためには遺伝子配列が必要となります。新型コロナウイルスの遺伝子配列は既に数百万もの数がデータベースに登録されています。今までいくつものコロナ変異株が登場しましたが、ウイルスは感染や増殖を繰り返す過程で変異を繰り返すために、配列によっては変異がより多く蓄積されます。これは、本来RNAウイルスは変異率が高い上、そもそも変異を100%防ぎながらゲノムを複製する手段が無いからです。また、配列によってはシークエンシングエラーも問題となります。ナノポアなどにおけるディープシークエンシングのエラー率は高く、さらには実験のエラーと変異を区別するのは至難の技です。実際、任意のウイルスゲノム配列を解析に用いる場合、シークエンシングエラーは変異と見分けがつかないために深刻な問題ともなり得ます。
遺伝子の変異を解析するためには「どの配列とどの配列を比較するのか」が重要になります。変異やシークエンスエラーのために、選択する配列によって解析結果が違ってしまうからです。コロナ変異株が誕生した後の変異やシークエンシングエラーなどの「ノイズ」を取り除くために、私は荒川論文のためにそれぞれのコロナ変異株の祖先型を同定し、祖先型が武漢型からどのように進化したかを検証しました。
実際、新型コロナの数百万個のゲノム配列の分子進化に関しても既にいくつもの論文が発表されています。しかし、大規模な解析では逆に見えにくい重要な現象もあります。研究において大切なものは目的と視点です。ノイズを排除した祖先型の配列の解析は人工ウイルス説検証に適していると私は考えました。配列の選定、祖先型の同定、計算手法など、シンプルな解析においてこそ一つ一つのステップが大きな意味を持つのです。
dN/dS法について


この指摘は正しくはありません。そもそもdN/dS比の計算法には長い歴史があり、そのプログラムやアプリケーションも以前から公開されているからです。
また、データを提供した私からの共著の「辞退」は痛烈な抗議の証です。

私からの「素人」という言葉については、相手に対する謙遜に過ぎません。
遺伝学では非同義部位あたりの非同義置換数 (dNまたはKa) と同期間における同義部位あたりの同義置換数 (dSまたはKs) の比が中立進化のバランスを推定するために使われます。dN/dS比の計算法にはLWL法 (1985年)、Nei-Gojobori法 (1986年) を始めとして10通り以上の計算法があります。dN/dS比は目的に応じて改良され、それぞれに長所や特徴があります。シンプルである、広く使用されている、標準化されている、短い配列に適している、複数の置換を処理できる、精度が高い、特定の変異のバイアスを補正できる、部位ごとの置換を推定できるなどです。初期に開発されたLWL法が現在でも使われているように、必ずしもどの解析方法がベストというわけでもなく、目的に応じて使い分けられます。このように、dN/dS比の計算法のために新しい計算式を作成する必要が特にあるわけではありません。
私は荒川論文のための研究の際に、予備解析として10通りの計算法を試しています。その結果、例えば、ベータ株スパイク遺伝子のdNの数値は0.0023892 (NG (= Nei-Gojobori法))、0.0023468 (LWL法)、0.00242272 (MLWL法)、0.0025465 (LPB法)、0.0025465 (MLPB法)、0.0023626 (GY-HKY法)、0.00235134 (YN法)、0.00226391 (MYN法)、0.0023626 (MS法)、0.00231212 (MA法) など (括弧内はdN/dS比の計算法) でした。

一般的なウイルス進化と比べて、変異株におけるdN/dSの差は実際には非常に大きく、それどころか些細な差を考慮する必要が無いレベルでした。大きな差をデータとして明示するためには新規の方法を開発する必要も無く、むしろ長い時間をかけて検証されている既知の手法を用いる方がより信頼性や説得力が生まれます。私は今回の研究にはdN/dS法の計算法による大きな差は無いと考え、シンプルなNG法を採用して計算を続けました。荒川論文のデータをLWL法でも再解析をしましたが、結果は同様でした。つまり、他の計算法を用いても結果や結論には大きな差は出ないと考えられます。
そもそも生物学的にはdN/dS法という既に確立された手法が存在します。そして、オミクロンのスパイク遺伝子はN変異とS変異の差が大きいため、異なるdN/dS法を用いても今回の目的では結果は大きく変わりません。にも関わらずあえてプログラムを組もうとするならば、研究倫理としては先行する論文の出版と競合する事態を避けるべきなのです。二重投稿は投稿規定に違反するものであり、事実上この事が共同研究決裂の最大の原因となりました。

また、コドン表のdN/dS法への応用は40年ほど前に既に確立している手法ですので、掛谷先生がなぜここに来てコドン表の分析から始めようとしているのか私には奇妙に映りました。コドン表は分子生物学教科書の最初に習うごく初歩的な知識であり、一体それをどう応用しようというのかと訝ってはいたのです。掛谷先生の手法の考え方のベースはむしろ原始的なdN/dS法に近いものです。dN/dS法はN変異やS変異をコドンによって補正 (コドン毎のN変異やS変異になりやすさで補正) する手法で、さらにはdN/dS法によっては特定の変異のバイアスを補正する事もできます。ではなぜその部分で今更「新しい」計算法を開発する必要性や必然性があったのかはなはだ疑問であり、むしろ掛合先生がdN/dS法自体をよく理解されていなかった証拠ではないかとすら感じました。実際なぜわざわざ「自前のプログラムを持ち出してまで、先行する荒川論文と類似の解析をしたのか?」理解に苦しみました。いずれにせよ、2つの論文の元となった遺伝子配列のデータは私から提供されており、どちらの論文も変異の不自然さからオミクロンが自然発生したものでないという仮説を論じています。そのため私は自分が提供したデータを掛谷先生は私と競合する目的に使用したと認識しました。
2つの共同研究の同時決裂について
以下は共同研究の決裂を決めた際の私からのメールです。
2022年3月13日
掛谷先生
論文をよく読んでからコメントさせていただきます。
その前にいくつか気になった事です。
1) 変異の元になったデータの扱いについて
変異の元になったデータは変異株の祖先型のalignmentから来ており、それはもう1つの論文のFIgure s1、s2に由来するものです。変異株の変異については論文も出ていますが、問題はどのアクセション番号の配列のデータを使うかであり、それをそれぞれの変異株の代表として扱っていいかという問題がありました。そのため、私はそれぞれの変異株のうち初期に採取されたもの同士の比較から祖先型を推定し、仮想的な祖先型から変異を同定しています。もう1つの論文のFIgure s1、s2のデータを使えば一義的に変異は決まり、今回のデータもそれに基づいています。逆に言うと、そうしたものがないとどうやって変異の数を同定したかがわからないという事になります。
変異の数を論じる際にもう1つの論文の引用という形にしていただきたいです。そうすればデータの扱いに問題が出ません。
2) それぞれの著者のこの論文の貢献についてはどうお考えでしょうか?一度先生のお考えをお聞かせください。
私は元になった論文の祖先型の同定、alignment、変異の同定に2ヶ月ほど費やし、また先生が必要とするデータを取るのにそれぞれ数日費やしています。KaKs caluculator以外には自動で計算できる便利なアプリを持っているわけではなく、alignmentデータの整理、カウント、目視での再確認など時間のかかる地道な作業です。
3) preprintについて
もう1つの論文の引用という形であれば問題ありません。私は私の論文を来週投稿を始めようかと思います。どこかの雑誌にin pressになってから引用という形ではどうでしょうか。
4) 私の書いた論文について
数人からコメントをもらいほぼ完成しました。研究所の所長の○○からタイトルを変え、Discussionの最後の2章を省いてほしい、表現を穏やかに変えてほしい。研究所を攻撃されないためだ、というコメントをもらい従いました。全体的な主張は変わりません。KaKsについて説明を加え、コントロールのデータを加えました。来週投稿を開始する予定です。最後の修正後、先生に原稿を送ります。
引き続きよろしくお願い致します。
荒川 央
質問 2)に対する掛谷先生からの返答は「荒川論文の方は、私の貢献度は1%かそれ以下ではないかと思っています」でした。また、あわせて「望むならば荒川論文から私の名前は外しても構わない」との言葉をいただきましたので、私は以下のように返信しました。
2022年3月13日
掛谷先生
返信をありがとうございます。諸々の事情を踏まえた結果、やはり私の執筆した論文に関しては単著で発表した方が良さそうに思いましたので、そのようにさせていただきます。投稿は来週始める予定です。掲載が決まってから掛谷先生の論文で引用されてください。塩基配列データもFigure s1、s2で公開されます。今回の経緯として、掛谷先生から共著で論文を書く誘いをいただいた際、それなら単著で書いている私の論文に合流させてはと考えて私からも共著の論文の提案をさせていただきました。しかし、結果的にこちらの論文はほぼ私の解析と執筆で出来上がる事になりました。
また、前回のメールにも書きましたが、研究所の現所長の○○からタイトルを変え、Discussionの最後の2段落を省いてほしい、表現を穏やかに変えてほしい。研究所を攻撃されないためだ、という事になり、同意しました。結果、大変申し訳ないのですが、掛谷先生の加筆も削らざるを得なくなりました。critical readingを除いて、アイデア、データ取得、解析、執筆を基本的には自分一人で担当した事になりました。
掛谷先生にはパリのミーティングもご紹介いただき、情報なども教えていただいて大変感謝しております。
気を悪くされないでいただけたらありがたく思います。掛谷先生の論文については引き続き協力させていただきます。引き続き宜しく御願い致します。
それでは失礼致します。
荒川 央
事実上、掛谷先生の貢献は何ら認める事ができなかったために荒川論文からは掛谷先生の名前を外す旨をお伝えしました。このような経緯を経て私の内心としては怒り心頭です。しかしながら、そのような事態であっても最後まで礼節は尽くすべきであると考え、「気を悪くされないでいただけたらありがたく思います」とも記しました。そして同時に「荒川論文の掲載が決まってからその論文を引用する形にしていただきたい」とお伝えしました。実際、配列などの元データは私から提供したものであり、それに由来する同様の内容を無断で先に出版されるわけにはいかなかったのです。そのため、こちらとしてはその時点では無駄に関係がこじれないように丁寧にお願いするしかありませんでした。そして、掛谷先生とその後の関係を断つか否かはこのメールに対する掛谷先生の返答次第とも思いました。

「真理の探究などは誰か別の人にでもやってもらえば良い。自分の論文発表はある意味政治的行為として正義のために行なっている」といった旨の掛谷先生の発言を聞いたのはZoomでお話した際のものであり、そもそも私は記事内で「メール」とは書いておりません。
しかしながら、「掛谷先生が論文を発表される際には荒川論文の引用という形にして欲しい」という私の要望は拒否されました。それどころか掛谷先生は、私が渡したデータを元に類似の変異パターンを持つ配列を探し出し、似たようなデータを再現しました。実際、解析すべき配列さえ分かれば類似の配列をデータベースから検索する事自体は容易なのです。また、たとえ既に公開されている配列を使ったとしても、問題が無いわけではありません。実際その配列が祖先型と同様の配列かどうかについては、共同研究者の立場により知り得た未公表の情報が無ければ判断できなかったはずだからです。そして掛谷先生は、「これで荒川には迷惑はかからないだろう」と主張されながら掛谷論文を修正しました。事実上これらの行為が私と掛谷先生との亀裂の決定的な原因となりました。そのため私は掛谷論文に名前を含める事を拒否しました。
二重投稿の禁止は研究者としてのモラルだけの問題ではありません。雑誌投稿の投稿規定にも明記されているものです。その規定に違反し同一著者が同様の研究を他の雑誌に投稿すれば、学術雑誌ひいては学術界に対する「詐欺」行為とも受け取られかねないのです。
論文の内容が被る場合は、先行の論文 (この場合荒川論文) と統合する、または被らない場合は2つの論文に分ける事もできます。しかし掛谷先生は荒川論文には自身のデータを提供せず、その後に内容が大きく被る掛谷論文を書き (その内容は私が提供した配列 (又は類似の配列) を利用し、変異解析によりオミクロンが人工ウイルスだと主張するもの)、私に先んじて投稿しようとされました。こうした行為は研究者としてはいわば「お前の研究を叩き潰す」「お前の論文を台無しにしてやる」という意味の宣戦布告と受け取られても致し方ありません。

オミクロン人工ウイルス説の検証には「新しい」手法の開発は特に必要ないはずでした。そして、「解析手法の開発」が掛谷論文の主な論旨であるならば、新型コロナの遺伝子配列を使う必要すらありません。掛谷先生が荒川論文に対して「先駆性が無い」「新規性が無い」「重要ではない」とおっしゃるならば、そもそもなぜ掛谷先生はご自身の論文に私が提供したデータを含めようとしたのでしょうか。そして私の抗議を受けた後、類似の配列を探し出してまでそのデータを含めた理由は何でしょうか?
改めて経緯を振り返りますが、最初、掛谷先生からの共同研究の依頼に応じて私は「現在自分は単著で論文 (荒川論文) を執筆中であり、先生が希望されるなら共著で発表しても良い」と返信しました。
つまり私は掛谷先生に「アイデアとデータをお互いに出し合って一緒に論文を書きましょう」と返信したという事です。それに対して掛谷先生は、ご自分ではアイデアもデータも出す事は無く、それどころか私の草稿を見た後に突如として別途に自分でも独立した論文 (掛谷論文) を書きたいと言い始めました。そしてその結果として出てきた掛谷論文は、荒川論文と被らないようにする配慮自体を怠ったものであり、事実上二重投稿にも当たるものでした。それを見た私は驚き呆れつつも「協力しますのでどうか順番を守って2番目に出してください」とお願いしました。けれどもその要請を断り、私のデータを別の似たデータに置き換えてまで先に投稿しようとしたのが掛谷先生のされた事なのです。そして掛谷先生はご自身の論文 (掛谷論文) の発表をことさら急ぐ理由を、危険な機能獲得実験を一刻も早く止めるためであり、社会のためなのだとZoomでお話しした際にも主張されましたが、私にはそれは「荒川を出し抜いてでも一刻も早く自分が先に論文を出す」ためのエクスキューズにしか聞こえませんでした。
実際、2つの論文の共同研究がほぼ同時に決裂したという事実が存在します。荒川論文から掛谷先生の名前を除いたのは掛谷先生の実質的な貢献が無かったからです。そしてデータの提供者である私が掛谷論文に荒川の名前を含めるのを拒否したのは私からの痛烈な抗議の証です。
学会発表や査読済み論文について





学会発表や査読済みの論文の出版は研究者にとっては日常です。それをここまで自慢するような「プロの研究者」を私はこれまで見た事がありませんでした。
掛谷先生が言わなかった事



コロナウイルス人工説を提唱する人達の中には、人工であるからこそ新型コロナは恐ろしい疫病であり、その恐ろしい疫病の救世主がコロナワクチンであると考える人も少なくありません。そしてその中にはコロナワクチンの危険性に対する警鐘を鳴らす者を陰謀論者として扱う人達がいるのも私はこれまで見てきました。
また、掛谷先生はご自分の論文の成果を頻繁にSNS等でアピールされますが、掛谷論文の背景に荒川との共同研究の決裂があった事実については触れてこられませんでした。「何を言ったか」ではなく「何を言わなかったか」も大切なのです。
盗用、剽窃という言葉について



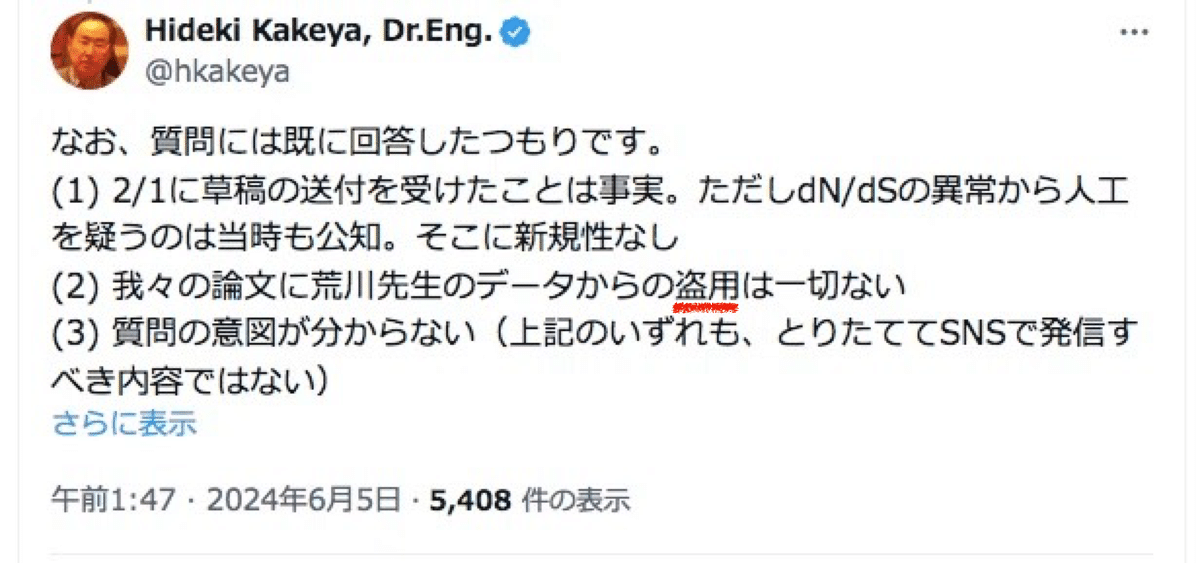
そもそも私は掛谷先生と私の共同研究決裂の経緯を説明させていただいたに過ぎず、記事内でも「盗用」や「剽窃」といった言葉は一切使っておりません。にも関わらず「盗んだ」「盗用」「剽窃」という言葉を繰り返し持ち出されているのは掛谷先生ご自身に他なりません。
真にLNP/mRNA製剤に反対しているのは誰なのか






改めて今回の経緯を例えるならば、「協力関係を結びましょう!」と言われて握手をしようとしたところ反対の手で殴りかかられたようなものです。しかも、抗議をしてもご本人にはまるで伝わる様子もありません。その上この期に及んでも掛谷先生は、私やコロナワクチンへのDNA汚染問題およびレプリコンワクチン問題に警鐘を鳴らす人達を揶揄し続けています。こちらは両手でガードを続けて来ましたが、延々と殴り続けられている状況なのです。正直私にはまさに理解不能です。
以下は前回の記事の繰り返しになりますが、最後にもう一度記しておきます。
思い返してもこの経緯は私の研究人生における「恥」だと感じています。この一連の話を私がこれまで話してこなかった理由は、これがあまりにも醜悪で「痛い」体験であり、いわゆる反ワクチン界隈の内輪的な揉め事や醜態を晒す事にもなるからです。そして、そもそも私はこういった相手への「正しい対処法」を知るわけでもなく、この件は実質的には泣き寝入りせざるを得ないと思っておりました。しかしながら、少し前からSNS上で直接的または婉曲的に私を揶揄するなどの掛谷先生の非礼な発言が散見され、もはや当人として看過する事はできなくなってきました。また掛谷先生は、mRNAワクチン反対運動やコロナワクチンへのDNA汚染問題、さらにはレプリコンワクチン問題の矮小化にも加担し始め、それどころか東京理科大学名誉教授の村上康文教授を含む問題を提起されている方達に対してまでも非礼な言動を始めました。もしこのまま私が沈黙を続けるならば、掛谷先生の行動を容認しているとも受け取られかねないと考え、ここで改めて掛谷先生の科学者としての姿勢を問うべきと思った次第です。そのような理由でこのタイミングになりましたが、今回ここで事の経緯を公表する事にしました。
#コロナワクチン
#ワクチン
#コロナ
*記事は個人の見解であり、所属組織を代表するものではありません。