水鳥の多様な水かき ~水かきの進化のひみつ~ 論文紹介

水鳥の多様な水かき ~水かきの進化のひみつ~
論文名 Developmental mechanisms underlying webbed foot morphological diversity in waterbirds
水鳥における蹼足形態の多様性の基礎となる発生機構
著者名 Masayoshi Tokita, Hiroya Matsushita and Yuya Asakura
掲載誌 Scientific Reports
掲載年 2020年
リンク https://www.nature.com/articles/s41598-020-64786-8
水鳥の水かきの形態がどのように進化したかを考察した2020年の論文です。
水鳥の水かきにはいろいろな形態をしています。水鳥を系統的に分類したときに、その系統と水かきに形態は一致しません。つまり、カモメはチドリ目ですが、チドリ目のすべての水鳥がカモメと同じ形態の水かきを持っているわけではないということです。このことは、同じような形態の水かきを持っていたとしても、その形態は独立に進化することで獲得されたと考えられます。進化を実験的に証明することはできるのであれば、実験によって、そのようなことが本当に起きたのかを調べることができますが、残念ながら現実的ではありません。そのため、別の方法で、検証する必要があります。
この論文では、Gremlin1遺伝子に注目しています。鳥の足の発生では、もともと趾(あしゆび)の間に趾間(しかん)組織と呼ばれる細胞群があり、趾がつながっています。発生が進むにつれて、この組織ではアポトーシスによる細胞死が起こり、その結果、趾が分離します(漫画「鳥の足のできかた」参照)。Gremlin1たんぱく質はアポトーシスを阻害しますので、趾間組織がそのまま残り水かきになります。同じ形態の水かきでも、発生段階でこの遺伝子の発現パターンが異なるのであれば、それは別々に進化したことを示す証拠となります。この方法を使って、うまく水かきの進化を説明しています。
しかし、ひとつの遺伝子の発現パターンの変化ですべてが説明できるほど進化は単純ではありません。この論文の考察にもありますが、その他の遺伝子が関係していることが考えられています。その他の要因も含めて、より多くの鳥類で調べることで、形態が独立にある一方向へと進化するなぞに迫れる可能性があります。この研究の今後の展開に期待したいと思います。
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
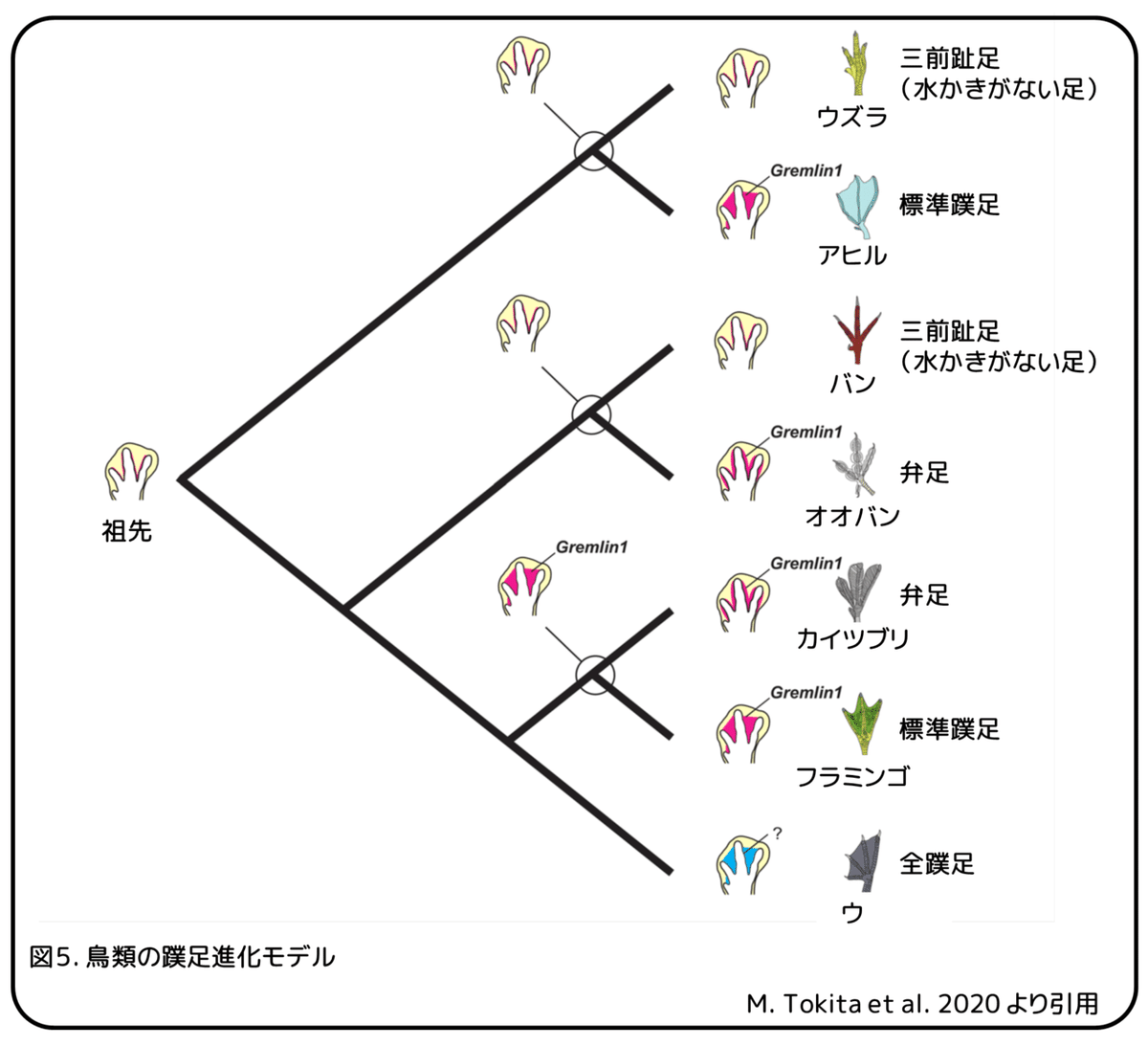
・バンとカイツブリの弁足は独立的に進化したと考えられる。
・バンでは三前趾足(水かきがない足)を持つ祖先からGremlin1遺伝子の発現領域が拡大することで弁足へ進化したと考えられる。
・カイツブリでは標準蹼足を持つ祖先からGremlin1遺伝子の発現領域が縮小することで弁足へ進化したと考えられる。
・ウの全蹼足はGremlin1遺伝子による制御とは別のメカニズムにより進化したと考えられる。
[背景]
水鳥は通常、足を使った推進力を得るために水かきのある足(蹼足、ぼくそく)を持っています。蹼足は、形態学的に標準蹼足、半蹼足、全蹼足そして弁足の4つのタイプに分類することができます。標準蹼足は、水鳥に最もよく見られるタイプで、前を向く3本の趾(第2趾、第3趾、第4趾)が完全に蹼膜でつながっています(図1A)。半蹼足では,前側の3本の趾の基部に部分的に蹼膜を持っています(図1A)。全蹼足は前を向く第2趾と蹼膜でつながった第1趾と、第2,3,4趾が全体的に蹼膜でつながっています(図1A)。弁足の4本の趾は互いに分離していますが、それぞれの趾の縁には葉状の膜があり、水中での推進力を生み出しています。

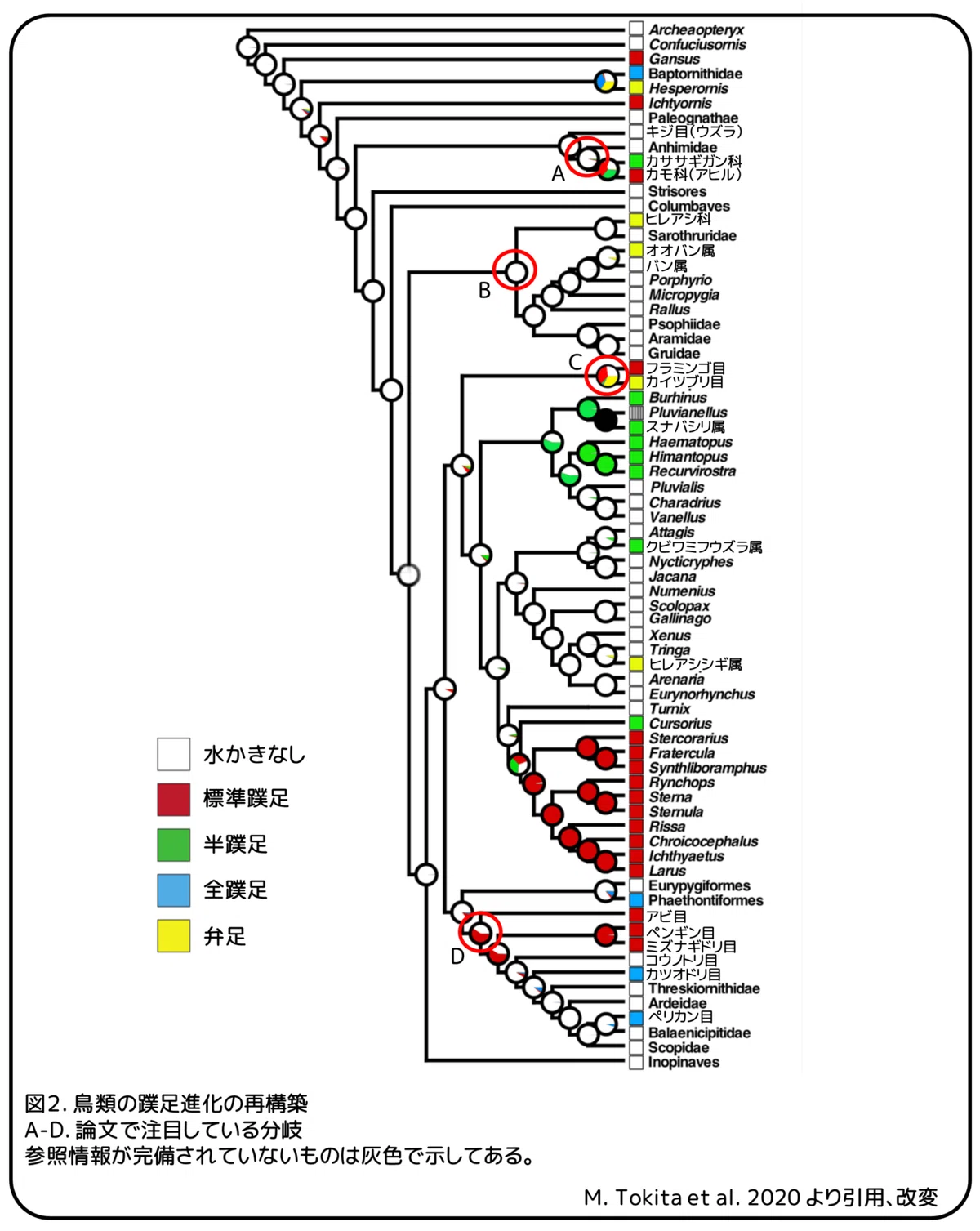
蹼足は現代の鳥類の5つの系統群で見られます。(1)標準蹼足を持つアヒルやガンと半蹼足を持つカササギガンを含むカモ目、(2)弁足を持つアメリカヒレアシやオオバンを含むツル目、(3)標準蹼足を持つフラミンゴを含むフラミンゴ目や弁足を持つカイツブリを含むカイツブリ目からなる系統群、(4)標準蹼足を持つカモメ、半蹼足を持つシギ・チドリ類、シギ科の弁足を持つヒレアシシギ属を含むチドリ目、(5)標準蹼足や全蹼足を持つジャノメドリ目、ネッタイチョウ目、アビ目、ペンギン目、ミズナギドリ目、コウノトリ目、カツオドリ目、ペリカン目を含む大系統群。蹼足は、現代の鳥類において独立的に少なくとも14回獲得されました(図1B)。鳥類の蹼足の起源と進化の歴史を十分に理解するためには、蹼足形成の発生メカニズムを明らかにすることが重要です。
鳥類の足は、発生的には胚の後肢芽(足板)の指板に由来します。足板は、将来の趾を生じさせる指列と、各指列をつなげる趾間組織からなります。ニワトリでは、発生段階表に基づく胚発生期(St)30〜35の間に、趾間組織細胞は、趾間細胞死(interdigital cell death, ICD)と呼ばれるアポトーシスを形態形成過程で起こします。(補足:アポトーシスは細胞の自然死のことで、多くは遺伝子によって制御されている。)ICDは、陸上の鳥類(例えば、ニワトリ、ハト、オウム、スズメなど)の大部分が持つ水かきの無い分離した趾の形成に寄与しています。胚発生におけるICDのタイミングは、陸生鳥類の種間で高度に保存されています。胚発生期32~34には、趾間組織が薄くなり、組織の退行が始まります。胚発生期35で、趾間組織は完全に退行します。趾間組織が消失した後に、各趾は分離し、爪と趾の膨らみが形成されます。
脊椎動物では、成長因子である骨形態形成たんぱく質(bone morphogenetic protein, BMP)がICDを促進します。しかし、指列の周辺組織は、ICDを回避し、腱や他のタイプの結合組織になります。足板のこの領域では、2つのBMPアンタゴニスト、GremlinとNogginがBMPシグナルを阻害し、ICDを阻害します。アヒル胚では、Gremlinの発現領域はすべての趾間組織に拡大しています。線維芽細胞増殖因子(fibroblast growth factor, FGF)もまた、ICDを防ぐことが知られています。例えば、FGF8はコウモリやイルカの胚の前肢の趾間組織に発現し、ICDを阻害することが知られています。
アヒルの足の発生に関する研究により、標準蹼足の進化の基礎となる発生メカニズムが部分的に明らかにされました。しかし、水鳥の蹼足の形態学的多様性の基礎となる発生メカニズムは、完全には解明されていません。特に、弁足の形成を制御する分子および細胞のメカニズムについての報告はほとんどありません。
本研究では、水鳥における蹼足の形態の多様性を理解するために、一連の比較解析を行いました。第一に、鳥類における蹼足の進化の過程を理解するために、最新のゲノム情報に基づく鳥類系統図に基づいて祖先的な特徴の再構成を行いました。第二に、蹼足のある種とない種の代表的な鳥類の数種の足形成パターンを比較し、鳥類の蹼足形成の一般性と多様性を明らかにしました。第三に、蹼足の形態学的多様化の基礎となる細胞機構を理解するために、Gremlin1遺伝子の発現パターンと趾間組織内でのアポトーシス細胞の局在を明らかにしました。最後に、弁足の進化の基礎となる細胞機構を理解するために、趾組織内での増殖細胞の分布を、弁足を持つオオバンとカイツブリ、および弁足を持たないコバンの3種の鳥で比較しました。
[結果]
蹼足の進化の歴史
鳥類系統の祖先的な特徴の再構築から、蹼足は次のように進化したと考えられます(図2)。カモ目の共通祖先は水かきのない足を持っていました(図2A)。カモ科(アヒル)はカササギガン科(カササギガン)と分岐した後で標準蹼足を獲得しました。共通祖先が水かきのない足(三前趾足)を持っていたツル目では、ヒレアシ科(ヒレアシ)とクイナ科のオオバン属(オオバン)において、おそらく並行的に水生環境への二次的適応のために獲得されました(図2B)。標準蹼足を持ったフラミンゴ目(フラミンゴ)と弁足を持ったカイツブリ目(カイツブリ)の共通祖先は標準蹼足を持っていたように見えます(標準蹼足:36%、弁足:33%、水かきなし:27%、半蹼足:2%、全蹼足:2%、図2C)。チドリ目の共通祖先は水かきのない三前趾足を持っていましたが、この科では蹼足進化の複雑性がみてとれました。標準蹼足はカモメ亜目の根本で一度進化しました。半蹼足は少なくとも3回進化していました。1回目はチドリ亜目の共通祖先で、2回目はクビワミフウズラ属(クビワミフウズラ)で、3回目はカモメ亜目のスナバシリ属(スナバシリ)です。弁足はヒレアシシギ属(ヒレアシシギ)で祖先の三前趾足を変化させることで一度進化しました。アビ目、ペンギン目、ミズナギドリ目、コウノトリ目、カツオドリ目、ペリカン目の共通祖先は標準蹼足を持っていました(図2D)。コウノトリ目、カツオドリ目、ペリカン目の共通祖先は標準蹼足を失い、水かきのない足を再獲得しました。カツオドリ目(カツオドリ、グンカンドリ、ウ)とペリカン科(ペリカン)の全蹼足は並行して進化した可能性があります。
趾形成パターンと趾間組織退縮の比較
足の形態形成において、いつどのようにICDがみられるのかをウズラ、マガモ、カイツブリ、カワウ、バン、オオバンの胚で確かめました。

ウズラ:三前趾足(水かきがない足) 発生段階33-34の間に、趾間組織は退縮をはじめ、それぞれの趾は指先側に突き出しはじめました(図3A)。発生段階35には、趾間組織は完全に消失し、発生段階36には、爪や趾の膨らみが形成されました。
マガモ:標準蹼足 発生段階33-34の間に、趾間組織の指先側の縁がわずかに退縮しました(図3B)。発生段階35には、第2趾と第3趾、および第3趾と第4趾の間の趾間組織は残ったままでしたが、第1趾は第2趾から完全に分離しました。爪が形成された発生段階36以降では、趾間組織は水かきとして残っていました(図3B)。
カイツブリ:弁足 発生段階33-34の間に、趾間組織の指先側の縁にくさび状の裂け目ができました(図3C)。発生段階35では、第2,3,4趾に沿った趾間組織の広い領域が残っていました。発生段階36以降は、趾間組織はより指先側に広がり、第2,3,4趾はオール状の突出部を伴っていました(図3C)。
カワウ:全蹼足 発生段階33-34で、趾間組織の指先側の縁がわずかにくぼみました(図3D)。発生段階35では、ほとんどの趾間組織は第1趾と第2趾、第2趾と第3趾、および第3趾と第4趾の間で残ったままでした。全ての趾で爪が形成された発生段階36以降は、趾間組織は残ったままでした。
バン:三前趾足(水かきがない足) 陸生のウズラ胚に見られるように、発生段階33-34において、趾間組織はすでに退縮していました(図3E)。発生段階35では、趾間組織は完全に消失し、それぞれの趾が完全に分離した後は、趾がウズラよりも比較的長くなりました(図3A、E)。
オオバン:弁足 発生段階33-34の間に、趾間組織は退縮をはじめ、それぞれの趾が成長する様子が観察されました(図3F)。発生段階35では、趾間組織が消失したように見えました。しかし、第2,3、4趾の幅はバンよりわずかに広くなっています(図3E、F)。発生段階36以降は、第2,3、4趾の幅が更に広がることで、趾に沿った突出部を形成しました。発生段階37では、突出組織のくびれが趾骨関節部で観察されました。
Gremlin1遺伝子の空間的発現パターンと趾間組織のアポトーシス
ウズラ胚では、発生段階31において、Gremlin1遺伝子は指根本側の趾間組織のほとんどで発現していましたが(図4A)、ニワトリ胚で報告されたように、発生段階33になると、その発現領域は比較的小さくなりました(図4B)。アヒル胚の発生段階31-33では、以前に報告されたように、趾間組織の指先側の縁を除いたほとんどの領域でGremlin1遺伝子は発現していました(図4D、E)。ウ胚の発生段階31では、Gremlin1遺伝子は全ての趾間組織に発現していましたが(図4G)、発生段階33になると、趾に隣接した領域を除いて発現は消失しました(図4H)。発生段階31のカイツブリ胚では、Gremlin1遺伝子は全ての趾間組織に発現していましたが、Gremlin1遺伝子が発現していないくさび状の領域が第3趾と第4趾の間にある趾間組織の指先から中間点にかけての領域で観察されました(図4J、赤い矢じり)。発生段階33では、この領域は根本方向へ広がり、裂け目状になりました(図4K)。同時に、Gremlin1遺伝子の発現していないくさび状の領域は第1趾と第2趾、および第2趾と第3趾の間にもみられました(図4K)。発生段階31のバン胚では、第1趾と第2趾、第2趾と第3趾、および第3趾と第4趾の間にある趾間組織の指の根本領域でV字型のGremlin1遺伝子の発現領域が観察されました(図4M)。発生段階33では、Gremlin1遺伝子の発現領域の境界が明確になり、指先に沿った発現になりました(図4N)。弁足を持ち、バンと近縁のオオバン胚の発生段階31では、第1趾と第2趾、第2趾と第3趾、および第3趾と第4趾の間にある趾間組織の幅広い領域で扇形のGremlin1遺伝子の発現が観察されました(図4P)。発生段階33では、バンのように、Gremlin1遺伝子は趾に沿って強く発現していました(図4Q)。しかし、オオバン胚の趾に沿ったGremlin1遺伝子発現領域の幅は、同じ発生段階のバン胚よりも広くなっていました(図4N、Q)。

胚の足の趾間組織におけるアポトーシス細胞の分布パターンは、調べた鳥種の間で大きな違いがありました。水かきを持たないウズラとバンでは、弁足を持つカイツブリとオオバンと同じように、アポトーシス細胞は趾間組織のGremlin1遺伝子の発現していない領域に分布していました(図4C、L、O、R)。標準蹼足を持つアヒルと全蹼足を持つウでは、わずかなアポトーシス細胞が趾間組織で見られました(図4F、I)。
[考察]
弁足の発生と進化
これまでの顕微鏡による観察と組織化学的な解析から、オオバンの弁足はICDによりそれぞれの趾が分離されたあとに、趾周辺から2次的に形成されることが分かっています。本研究の胚発生の種間比較によって、このことは確かめられました(図3F)。オオバンの水かき形成の過程は、ICDが起こらない趾間組織が最終的に水かきになるアヒルの過程とは異なるものです(図4D、E、F)。本研究では、弁足を持つオオバン胚で趾に沿ったGremlin1遺伝子の発現領域の幅が、弁足ではないバン胚の同じ発生段階と比べて広いことが分かりました(図4N、Q)。このことから、オオバンの趾の突出部は2段階で形成されると予測されます。はじめに、第2、3、4趾周辺のGremlin1遺伝子発現領域が突出部の原基を形成し、続いてICD終了後に、細部増殖率を上昇または維持することで突出部原基を大きくします。
オオバンとカイツブリはそれぞれ独立的に弁足を獲得したにもかかわらず、両種の胚の趾間組織においてGremlin1遺伝子は発現していました(図4J、K)。カイツブリの弁足で見られた関節部におけるくびれがない幅広い突出部の形成はGremlin1遺伝子の幅広い発現領域と関係している可能性があります。このことから、カイツブリ胚のGremlin1遺伝子を発現している趾間組織は、オオバンで指摘したような突出部原基の2次的な成長による相対的成長ではなく、足全体の一律な成長によって将来的な突出部になると予測されます。
祖先型の再構築から、オオバンの共通祖先は三前趾足を持っていましたが、カイツブリの共通祖先は標準蹼足を持っていたと考えられます(図2)。これらの結果から、この2つのタイプの弁足は別々の発生過程によって形成されると考えられます(図5)。オオバンの弁足は、たぶん三前趾足を持つ祖先の趾に沿ってGremlin1遺伝子発現する趾間組織の細胞増殖を介して進化しました。一方で、カイツブリの弁足は、標準蹼足を持つ祖先では趾間組織全体にみられたGremlin1遺伝子の発現がくびれることで進化した可能性があります。
全蹼足の発生と進化
全蹼足を持つウの発生段階31の胚では、Gremlin1遺伝子は後肢の全ての趾間組織で発現していました(図4G)。しかし、その発現は、発生段階33の胚の趾間組織中央部で消失しました(図4H)。このことから、全蹼足の水かきはアヒルの標準蹼足とは異なる発生機構によってもたらされたと考えられます(図4D、E、図5)。これまでに、FGFを染み込ませたビーズを鳥類の趾間組織に移植することでICDを阻害することが分かっています。また、翼とひれあしをそれぞれ持つコウモリとクジラでは、FGF8遺伝子が趾間組織に発現することが報告されています。FGFシグナルの制御の変化は、全蹼足を持つウにおける趾間組織での細胞死を回避することが可能なメカニズムでしょう。
祖先型の再構築から、アビ目、ペンギン目、ミズナギドリ目、コウノトリ目、カツオドリ目、ペリカン目の共通祖先は標準蹼足を持っていましたが、コウノトリ目、カツオドリ目、ペリカン目の共通祖先は二次的に水かきのない足を再獲得したと考えられます(図2)。また、カワウとその近縁種を含むカツオドリ目、ペリカンとその近縁種を含むペリカン目、およびネッタイチョウとその近縁種を含むネッタイチョウ目の全蹼足は独立的に獲得され、標準蹼足ではなく、水かきのない三前趾足から生じたことが分かりました。複合的な形態構造が非常に複雑な発生過程を経て作られた後に失われたとき、その特徴の再獲得は一般的にまれなことです。しかし、新しい発生メカニズムによって再獲得されたいくつかの例があります。もし、標準蹼足と全蹼足が異なる分子メカニズムによって形成されるのであれば、標準蹼足から全蹼足への、またその逆においても、連続的な進化は必ずしも必要ではない可能性があります。全蹼足を持つ鳥類が足発生メカニズムを共有しているかどうかを明らかにするために、ペリカン目とネッタイチョウ目の胚発生を調べることが重要でしょう。
よろしくお願いします。