
【高校生物】発生③「ショウジョウバエのボディープランはどのように形成されるのか?」

~プロローグ~
ショウジョウバエは、サイズが小さいため研究室のスペースを取らず、飼育が簡単である。また、丈夫で繁殖力が強く、幼生からは、遺伝学解析に有用な唾腺染色体を取り出すことができる。ショウジョウバエのボディープラン(多細胞動物のもつ基本的体制[外形や体内の器官の配置など、形態の基本的なパターン])に関する研究は、動物界全体に関わる重大な知見も提供してきた。遺伝子に関する話がたくさん出てきて大変だと思う(実際、ショウジョウバエの発生の研究が大きく発展したのは、遺伝子を扱う研究ができるようになってからである。ショウジョウバエの胚は小さくて複雑であり、顕微鏡観察や移植実験などにはあまり向かない)が、現代の分子発生生物学の雰囲気を感じてほしい。
「多くの結果がいきなり降ってくるその突如性が、われわれを誤らせる。だがそれは、われわれにとっての突如性でしかない。われわれから逃れ去るこの秒刻みの突如性のうちには、かぎりなく多くの過程がひそんでいる。」ニーチェ『愉しい学問』より
「いかに多くの種々な自然の要素が相互に関係し合っているか、ということを分析することが自然哲学の仕事の一つである。」ホワイトヘッド『科学的認識の基礎』より
★テストに出やすいワード
①母性因子
②分節遺伝子
③ホメオティック遺伝子
④ホメオティック突然変異
⑤表割
要点:ショウジョウバエの体節の形成は分節遺伝子により制御されている。
(1)ポイント
まずは「3つの分節遺伝子が体節構造の形成に関わり、ホメオティック遺伝子が各体節に個性を与える」ということだけ理解しておけばよい。
大雑把に言えば、まずたくさんの部屋(体節)をつくってから、それぞれの部屋に別々の模様替えを施す(それぞれの部屋に個性を持たせていく)イメージ。

(2)分節遺伝子
● ショウジョウバエの体節の形成は、①~③の分節遺伝子により制御されている。
①ギャップ遺伝子→②ペアルール遺伝子→③セグメントポラリティ遺伝子

*分節遺伝子:ショウジョウバエの胚発生において、前後軸に沿って、体節単位の繰り返しから成る分節構造へと転換する過程を制御する遺伝子群。
語呂「ギゅっとペアではグするホリデイ(ギャップ、ペアルール、セグメントポラリティの順番ではたらく)」
雑談:動物体の体軸方向に同じ構造が繰り返し並んでいるとき、その単位構造を体節(segment)という。ここで言う「体節(segment)」は、カエルの発生に出てきた「体節(somite:脊椎動物の発生において見られる中胚葉性の分節構造)」とは別の語である。
雑談:分節遺伝子のそれぞれの遺伝子群に変異が起きると、次のような表現型が見られる(それぞれの語源になっている)。
ギャップ遺伝子:「体節に大きな欠落(ギャップ)ができる」
ペアルール遺伝子:「2つに1つが欠落する(ペアルール)」
セグメントポラリティ遺伝子:「各体節のすべてに異常が出る。たとえば極性がおかしくなる(ポラリティが極性という意味)。それぞれの体節には明確な前後パターンがある[たとえば前方領域にのみ小歯状突起(おおきな棘)がある]が、セグメントポラリティ遺伝子の突然変異は体節内のパターンを変えてしまう。」
(3)ホメオティック遺伝子
● ホメオティック遺伝子と呼ばれる遺伝子群が、各体節に個性を与える。
*ホメオティック遺伝子の詳細については後述する。
(4)ショウジョウバエの初期発生の詳細な流れ
(あまりテストに出ない。まずはギャップ遺伝子→ペアルール遺伝子・セグメントポラリティ遺伝子の順番だけチェックしておけばよい。)
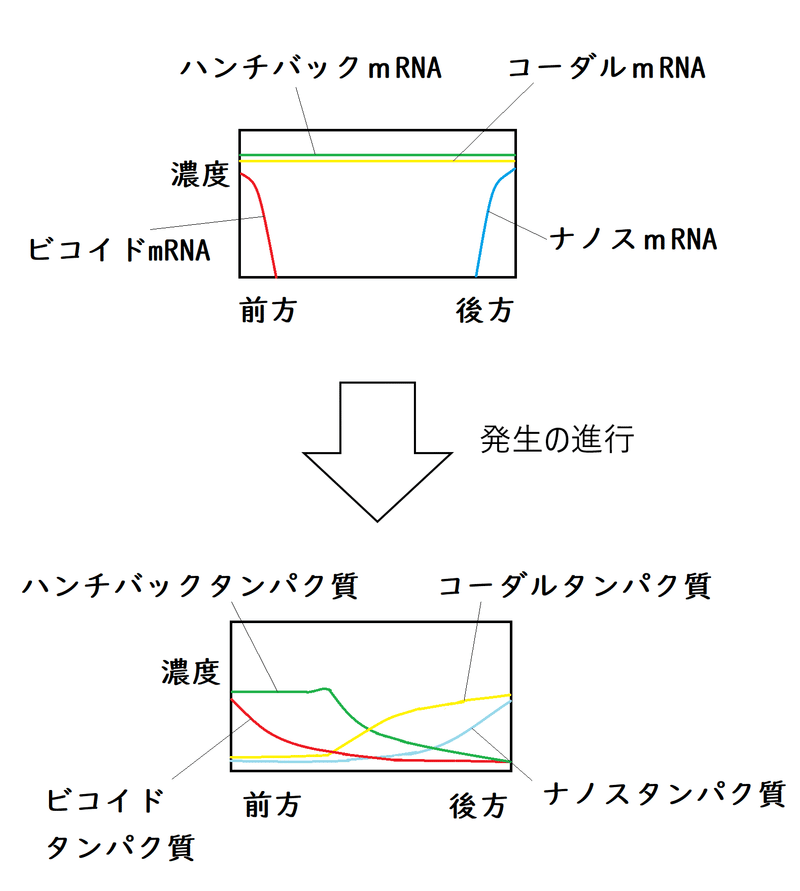
①未受精卵の前方にはビコイドmRNAが、後方にはナノスmRNAが局在している(どちらも有名な母性因子である)。これらのmRNAから翻訳によってビコイドタンパク質とナノスタンパク質が生じる。ビコイドタンパク質とナノスタンパク質の濃度勾配が前方と後方の方向性を決める。
・卵にあらかじめ入っているmRNAやタンパク質を母性因子という。
・卵形成の完了時には、ビコイドmRNAが前極に、ナノスmRNAが後極に局在するようになる。これら2種のmRNAは、翻訳されないよう休眠状態で貯蔵されており、排卵あるいは受精を機に翻訳される。ビコイドタンパク質は前極が高濃度となる濃度勾配を形成し、ナノスタンパク質は後極が高濃度になる勾配を形成する(核が分裂して増加すると[ショウジョウバエの発生では、まずはじめに核のみの増加が起こる。細胞の仕切りは後から出来る]、各々の核には、ビコイドタンパク質、ナノスタンパク質の存在比によって位置情報が与えられる)。

語呂「ビコイドなの!(ビコイドが前、ナノスが後ろ)」
②母性因子によって、受精卵の中で色々なタンパク質の濃度勾配のパターンが実現する。それらは、ギャップ遺伝子の発現を制御している(たとえば、ビコイドは前方特異的なギャップ遺伝子を活性化し、後方特異的なギャップ遺伝子を抑制する)。
③ギャップ遺伝子が胚をおおまかに領域化する。
・ギャップ遺伝子にコードされている転写因子はペアルール遺伝子の発現を導く。
雑談:ギャップ遺伝子の突然変異体は、体節の一部区画が大きく欠失し、ギャップを生じる(『ギャップ』の語源)。ギャップ遺伝子は、一定の幅(約3体節分)で発現し、これら遺伝子の発現領域は一部で重なっている。
④ペアルール遺伝子がはたらく。
・ペアルール遺伝子は転写因子をコードし、セグメントポラリティ遺伝子を活性化する。
雑談:ペアルール遺伝子は7つの領域を決定する。各々のペアルール遺伝子は胚を約2体節の幅で分画化する。ペアルール遺伝子の突然変異体は、1体節おきに一部領域が欠失する(『ペアルール』の語源)。既知の8種類のペアルール遺伝子の発現領域は、すべて一致しているわけではない。また、発現するタイミングも同じではなく、第一次ペアルール遺伝子の発現は、第二次ペアルール遺伝子の発現やその抑制に影響する。異なった組み合わせのペアルール遺伝子産物が、セグメントポラリティ遺伝子を活性化する。
⑤セグメントポラリティ遺伝子がはたらく。
雑談:セグメントポラリティ遺伝子は転写因子やシグナル伝達に関与する分子をコードしている。
雑談:セグメントポラリティ遺伝子は7つの領域を14のストライプに分割する。
セグメントポラリティ遺伝子のmRNAやタンパク質は、胚を14の体節単位に区画化し、胚は体節からなる繰り返し構造を確立する。
また、セグメントポラリティ遺伝子は体節内のパターンの形成に関わる(それぞれの体節は前後が決まっている[前後パターンを持っている]。たとえば、各体節の前側には小歯状突起[色素の沈着した大きな棘]があるが、後ろ側にはない。セグメントポラリティ遺伝子はその体節内のパターン形成に関わる。セグメントポラリティ遺伝子に変異が起きた場合、体節の数は正常だが、体節の一部が欠失し、代わりに残りの部分が鏡像になって重複するようになる)。
ペアルール遺伝子とセグメントポラリティ遺伝子が作るパターンが重なり合って擬体節が区分化される(擬体節は、最終的に幼虫あるいは成虫の体の表面にあらわれる体節とは約半体節分位置がずれている)。
⑥ホメオティック遺伝子が各体節に個性を与える。

(5)分節遺伝子・ホメオティック遺伝子のはたらきのイメージ
*雰囲気だけつかめばよい。わかっていないことも多い。
①受精卵内にはいろいろなタンパク質(たとえばビコイドやナノス)が濃度勾配をつくっている。受精卵内のタンパク質は、ギャップ遺伝子の発現を制御する。
②ギャップ遺伝子、ペアルール遺伝子、セグメントポラリティ遺伝子は分節遺伝子であり、体節の形成を制御する。
それと同時に、ホメオティック遺伝子の発現領域も制御する。
③ホメオティック遺伝子は各体節に個性を与える。

ホメオティック遺伝子は各体節に個性を与える。
雑談:ショウジョウバエの発生における胚の動きについて(知らなくてよい)
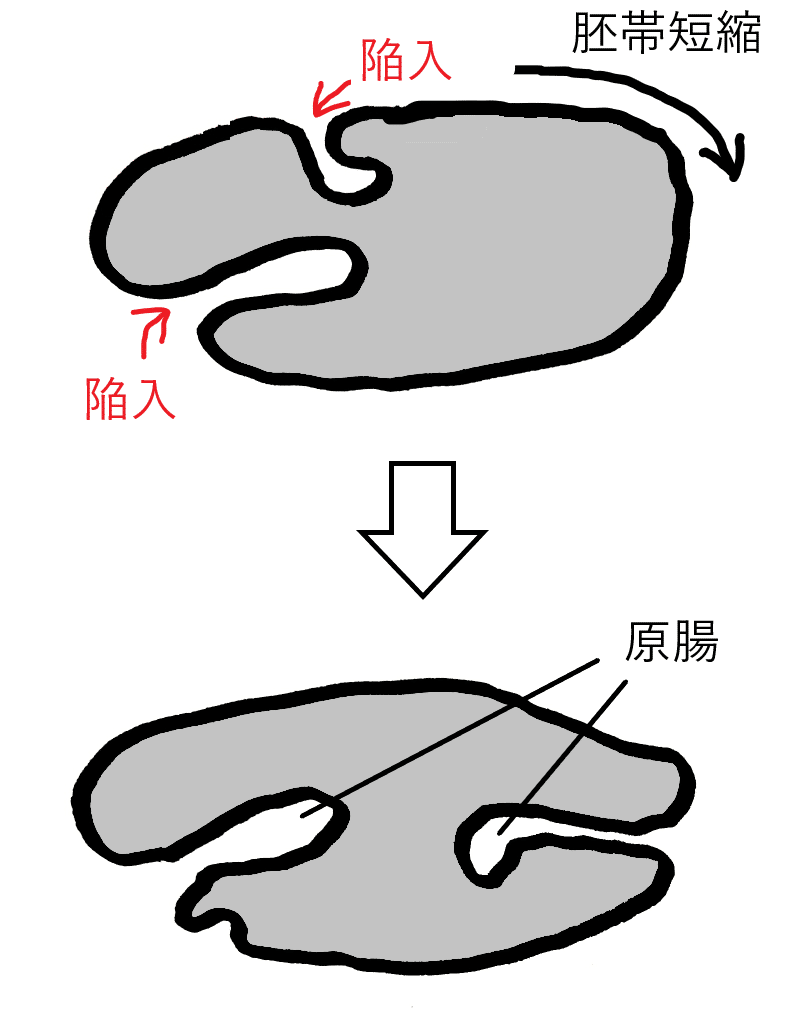
受精卵→胞胚→原腸胚となった後、14体節の体ができ、胚の後方が背側に折り返す(胚帯伸長[はいたいしんちょう]という。下図左)。やがて下図右のように、胚は縮んで元に戻る(胚帯短縮[はいたいたんしゅく]という)。その後、ふ化して幼虫となる。幼虫は脱皮を繰り返し、蛹となり、羽化して成虫となる。

雑談:原腸胚の初期、腹側に沿った胚帯は、後ろに伸び始め、胚帯伸長を見せる。その後、胚帯短縮が起き、消化管、気管、神経などへの分化が始まる。胚帯短縮の頃から、胚帯は左右両側から背側に向かって伸び、最終的にジッパーを閉じるように背部を閉鎖する。
雑談:ショウジョウバエの原腸形成は特殊であり、基本的に問われない(節足動物の原腸形成は多様である)。前方と後方から陥入する細胞群は腔所を作り、その両者が胚体内部に伸長する(下図はイメージ。あまり正確ではない)。図には描いていないが、この後、腔所は前後から出会って融合し、1本の管を形成する(前端と後端からの2つの陥入は中央で融合することで消化管が形成される)。
*ショウジョウバエは節足動物(旧口動物)であるので、原口は口になるのだが、実際は、消化管をつくるための予定内胚葉域の陥入は前後両側から起きる(下図のような内胚葉の陥入に先立って、中胚葉予定域細胞が胚内部へ落ち込み、中胚葉へと運命づけられる現象が起こる[腹側中胚葉の陥入]。このように、ショウジョウバエの胚の細胞の動きは少しややこしい。が、試験ではまず問われないから安心してよい)。
要点:ホメオティック突然変異の原因遺伝子として同定された遺伝子をホメオティック遺伝子という。
● ホメオティック突然変異の原因遺伝子として同定された遺伝子をホメオティック遺伝子という。

● 体のある部分が別の部分で置き換わってしまうような突然変異を、ホメオティック突然変異という。ホメオティック遺伝子ははじめ、ショウジョウバエで発見された(現在ではホメオティック遺伝子は動物、植物を問わず様々な生物で発見されている。植物におけるABCモデルに関する遺伝子などもホメオティック遺伝子である。A、B、Cのどれかの遺伝子が壊れたら、花の構造が変わってしまう[本来できるべき位置とは異なった位置に様々な構造ができてしまう])。
*ホメオティック突然変異体の例
①アンテナペディア突然変異体(触角が脚に置き換わった)
②ウルトラバイソラックス突然変異体(平均棍が翅に置き換わった)
雑談:アンテナペディア突然変異体では、触角(antenna)が脚(pedis)に転換されている。ウルトラバイソラックス突然変異体では、二重(bi)の胸部(thorax)と2対の翅が生じる(正常な双翅目のショウジョウバエは、その双翅目の名の通り、1対の翅しか有しない)。

雑談:平均棍(へいきんこん)・・・ショウジョウバエが飛ぶ時の安定性を保つ。平均棍は後翅が変形したものと考えられている。
雑談:ショウジョウバエは双翅目(そうしもく)であり、その名の通り、通常は翅は1対である。ハチなどは2対の翅をもつ。双翅目は4枚翅の昆虫から進化してきたと考えられているので、翅を4枚もつウルトラバイソラックス変異体は、先祖返りとも言える(ただし、進化に関わった遺伝子については確定していない)。
下図は正常な双翅目のイメージ。

● ホメオティック遺伝子には、ホメオボックス(180塩基対からなる)と呼ばれる共通の配列があることが多い。
● ホメオボックスにコードされるタンパク質の領域はホメオドメイン(60アミノ酸からなる)と呼ばれる。

雑談:ホメオティック遺伝子は転写因子をコードしている(ホメオドメインの領域がDNAとの結合に関わる。ホメオドメインは、ヘリックス・ターン・ヘリックスという、多くのDNA結合タンパクがもつ特徴的な構造をもつ。一般に、ホメオドメインをもつタンパク質は、DNAに結合し、転写因子としてはたらく)。
● ショウジョウバエのホメオティック遺伝子と相同な遺伝子は、ほとんどすべての動物に見出されており、ショウジョウバエのものも含めてHox遺伝子(ホックス遺伝子)と呼ばれている。
雑談:Hox遺伝子にコードされているタンパク質には、DNA結合領域であるホメオドメイン(ホメオボックスにコードされる領域)が含まれている。「Hox」は「ホメオボックス homeobox」の略に由来する。
雑談:はじめ、ホメオティック突然変異(体の構造の一部が別の構造に変化する形態形成異常変異)の原因遺伝子として、ホメオティック遺伝子が発見された。しかし、その後、同じような遺伝子が脊椎動物にも存在することがわかり、Hox遺伝子という総称がついた。
雑談:最初のホメオボックスがショウジョウバエで発見されるやいなや、生物学者たちは、そのホモログ(進化の過程で一つの共通祖先遺伝子に由来すると考えられる遺伝子)が脊椎動物にも存在するのではないかと予想した。そして実際にHox遺伝子が発見されたのである。
雑談:Hox遺伝子は、「ホメオボックスをもつ遺伝子の中で、ショウジョウバエのアンテナペディア遺伝子群とバイソラックス遺伝子群に帰属するホメオティック遺伝子群を足し合わせたものと進化的起源を共有すると考えられる遺伝子群」を指すことが多いが、定義には揺れがある。高校生は今使っている教科書に解釈を合わせればよい。
講義動画【ホメオティック遺伝子】
雑談:以下にホメオティック遺伝子に関わる用語についての雑談を記す。高校生はまったく気にしなくてよい。細かい定義には揺れがある。
● ホメオティック遺伝子=Hox遺伝子とすることもある。
● 生物界を見渡せば、ホメオボックスを持たないホメオティック遺伝子も多数ある(ホメオティック突然変異の原因となる遺伝子をホメオティック遺伝子と定義すれば)。
● 一般に、ホメオティック遺伝子は転写因子をコードするものが多い。動物では主として(ホメオボックスにコードされる)ホメオドメインを持つ転写因子、植物では(MADSボックスにコードされる)MADSドメインをもつ転写因子である。
● ホメオボックスを持つ遺伝子をホメオボックス遺伝子といい、その一部をHox遺伝子と呼ぶこともある(ホメオボックスをもつ遺伝子をホメオボックス遺伝子と総称し、ホメオボックス遺伝子を、ホメオドメインのアミノ酸配列・ショウジョウバエのホメオボックス遺伝子との類似性・ホメオドメイン以外の特徴などから、いくつかのグループ[Hox遺伝子、POUファミリー遺伝子、LIMファミリー遺伝子]に分類することも多い)。
● ホメオボックス配列を含むが、ホメオティック遺伝子としては機能しない遺伝子も発見されている。しかし、少なくとも動物では、ホメオボックス配列を含む遺伝子の大部分が発生に関与している。
● Hox遺伝子を持つことを、動物の共通の特徴とすることもある。
● ショウジョウバエに対してのみホメオティック遺伝子という名称を用いることもある。
● Hox遺伝子をhomeobox遺伝子の略とすることもある(たとえば『ギルバート発生生物学』メディカル・サイエンス・インターナショナル)。ただし、前述した通り、ホメオボックスをもつ遺伝子の中の1つのグループをHox遺伝子とすることも多い。
発展:ショウジョウバエのホメオティック遺伝子群と哺乳類のHox遺伝子群
● ショウジョウバエのホメオティック遺伝子は1個だけではない。ショウジョウバエは8個のホメオティック遺伝子をもつ。これらの遺伝子は、2つのブロック(アンテナペディア複合体とバイソラックス複合体)に分かれて1本の染色体上に存在している。これらを、ホメオティック遺伝子群と呼ぶことがある。下図はショウジョウバエのホメオティック遺伝子群のイメージ。
*ショウジョウバエのホメオティック遺伝子群もHox遺伝子群と呼ぶことが多い。

● アンテナペディア複合体には5つの遺伝子が、バイソラックス複合体には3つの遺伝子がある(ショウジョウバエは合計8個のホメオティック遺伝子をもつ)。アンテナペディア突然変異体は、アンテナペディア複合体にあるAntp遺伝子が異常発現していると考えられている。ウルトラバイソラックス突然変異体では、バイソラックス複合体にあるUbx遺伝子が正常に発現していないと考えられている。
● ショウジョウバエのホメオティック遺伝子群は1つであるが、マウスとヒトではそれに類似した遺伝子群を4組、すなわち4組のHox遺伝子群(HoxA、HoxB、HoxC、HoxD)をもつ(それぞれ別の染色体上にある)。下図は哺乳類のHox遺伝子群のイメージ。各遺伝子群に含まれる遺伝子には、3'から5'方向に、1番から13番までの番号が付けられている。
各Hox遺伝子群内での遺伝子の並びは基本的に昆虫のホメオティック遺伝子群と同じである(これら4つのHox遺伝子群は、すべて、原型となる1個の遺伝子群から重複によって生じたらしいと考えられている)。

ホメオティック遺伝子群、Hox遺伝子群では、遺伝子が一定の順序に並んで集団を形成している。これまでの研究で、以下のように、その遺伝子の並ぶ順序に重要な意味があるらしいことが分かっている(ただし、どうしてそのようになっているのか、どのような発現調節が行われているのか等、完全にはわかっていないことも多い)。
・発生の過程で発現する遺伝子の順番が、3'から5'方向に並んでいる順番と一致する傾向がある(3'の方にある遺伝子の方が、5'の方にある遺伝子より、発生の早い時期に発現する傾向がある)。
・3'から5'方向に並んで分布する遺伝子の順番と、動物の頭→尾方向に沿って発現する遺伝子の順番が一致する傾向がある(3'の方にある遺伝子は前方[頭がある方]で、5'の方にあるHox遺伝子は後部で発現する傾向がある[ただし、前部領域で発現する遺伝子は、後部領域でも発現し、他のHox遺伝子の領域と重なり合うことが多い。また、より後方で発現しているHox遺伝子は、より前方で発現しているHox遺伝子の機能を無効にする傾向がある。この現象は後方優位として知られている])。
*下図は一般的な傾向のイメージ(あくまでもイメージ。正確ではない)。

3'から5'方向に並んで分布する遺伝子の順番と、動物の頭→尾方向に沿って発現する遺伝子の順番が一致する傾向がある。
講義動画【ショウジョウバエの発生(分節遺伝子など)、哺乳類のHox遺伝子群】
要点:ショウジョウバエの卵は心黄卵であり、卵割様式は表割である。
(1)哺育細胞
雑談:ショウジョウバエの卵原細胞は、不完全な細胞質分裂を伴った4回の核分裂を行い、16個の繋がった細胞群を形成する(それらはリングキャナルと呼ばれる通路で連結されている)。このうち15個が哺育細胞(ほいくさいぼう)となり、1つのみが卵母細胞になる。これらの細胞は、何百もの濾胞細胞(ろほうさいぼう)という体細胞で囲まれた卵室に収納されている。下図はイメージ(細胞数などは正確ではない)。
● 哺育細胞や濾胞細胞が卵母細胞にmRNAやタンパク質を供給する。これらのmRNAやタンパク質が母性因子(ぼせいいんし。卵にあらかじめ入っているmRNAやタンパク質)として働き、軸形成に関与する。

雑談:ショウジョウバエの卵の成長は哺育細胞からの細胞質の流入による。
雑談:ショウジョウバエの哺育細胞は卵形成時にできる。
(2)表割の詳細
(『心黄卵』と『表割』以外ほぼテストに出ない)
● ショウジョウバエは、ほとんどの昆虫と同様、表割(ひょうかつ)を行う(その卵は中心に卵黄が多い心黄卵[しんおうらん]である)。
● 卵割期には、細胞質分裂を伴わない核分裂が繰り返され、多核細胞が生じる。
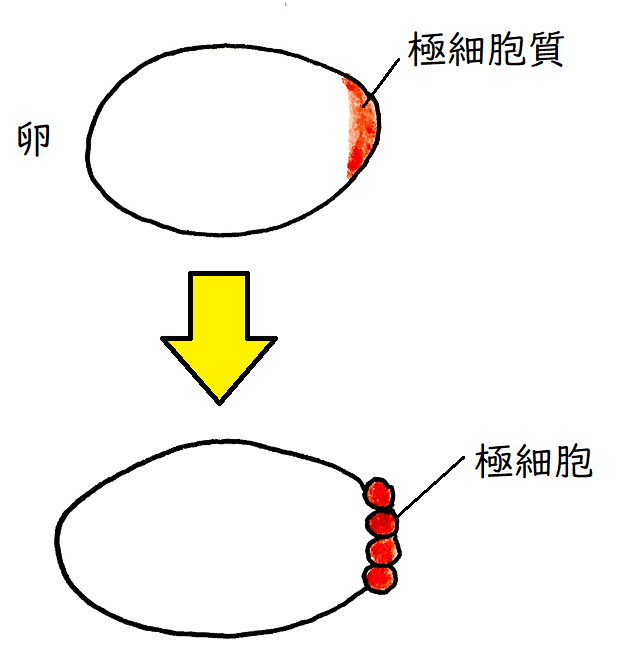
● 9回目の核分裂の時に、4~5個程度の核が胚後極の表層へと到達する。この5個程度の核は、やがて細胞膜に覆われると、将来成虫において配偶子を作り出す極細胞(きょくさいぼう)となる。
雑談:双翅目などの多くの昆虫の受精卵の後極には、極細胞質という特殊な細胞質が存在する。極細胞質には、生殖細胞(と腹部)の形成に関わる母性因子が含まれている。極細胞質を含んだ細胞は極細胞となり、生殖細胞に分化する(極細胞質を人為的に卵の後極以外の場所に移植すると、その場所に極細胞が形成され、この極細胞は生殖細胞に分化する能力をもつことが知られている)。
雑談:極細胞質を他の胚の前方に移植すると、極細胞質によって取り囲まれた核は、生殖細胞の核として指定されることが分かっている。
さらに、もしその細胞を第3の胚の後極に移植すると、その細胞は機能を持つ生殖細胞として分化する。
雑談:卵の後極に紫外線を当てると極細胞の形成が阻害されるが、この極細胞の形成阻害を回復させる分子として、ミトコンドリアの large ribosomal RNA(mtlrRNA)が見出されている。mtlrRNAの他にも、極細胞の形成には多数の物質が関わることが明らかになっているが、そのしくみについては完全には解明されていない。
● 核分裂の後、細胞膜がくびれこみ、細胞化が行われる。
雑談:細胞質分裂によって多核性が失われた胚を細胞性胞胚という(なお、胚全体の細胞化が完了するのは14回の核分裂時である)。
雑談:受精は、精子が卵門と呼ばれる開口部から侵入することで起こる(ショウジョウバエでは、精子が卵内に入ることのできる場所が1か所に決められている。その場所を卵門という)。

要点:ショウジョウバエの未受精卵には、母性因子として、前端にビコイドmRNA、後端にナノスmRNAが局在している。
(1)ポイント
まずは
①「母性因子」「母性効果遺伝子」の用語
②ビコイドmRNA、ナノスmRNAが母性因子であること
③ビコイドmRNAが前端に、ナノスmRNAが後端に局在していること
④様々な生物の発生には、モルフォゲンとして働くタンパク質が関わること
だけチェックすればよい。
(2)母性因子
● 母性因子(ぼせいいんし)は、卵にあらかじめ入っているmRNAやタンパク質のことである。
● 母性因子をコードしている遺伝子を母性効果遺伝子(ぼせいこうかいでんし)という。
講義動画【母性因子が関わる遺伝の問題】
(3)モルフォゲン
● 発生の過程では、いろいろなタンパク質(たとえばビコイド)の前後軸に沿った濃度勾配が形成される。
● 濃度差によって異なる細胞運命を特定化する因子をモルフォゲンという(「形作るもの」の意味)。様々な生物の発生において、モルフォゲンのはたらきは重要である。
発展:ショウジョウバエの母性因子
以下、ショウジョウバエにおける母性因子のはたらきに関する詳細。覚えなくてよい。
・ビコイドタンパク質は拡散により、前端が濃く後端が薄い濃度勾配を形成する。そのため、発生が進んで細胞性胞胚になる(細胞の仕切りができる)と、胚の前方にある細胞ほどビコイドタンパク質の濃度が高くなる。
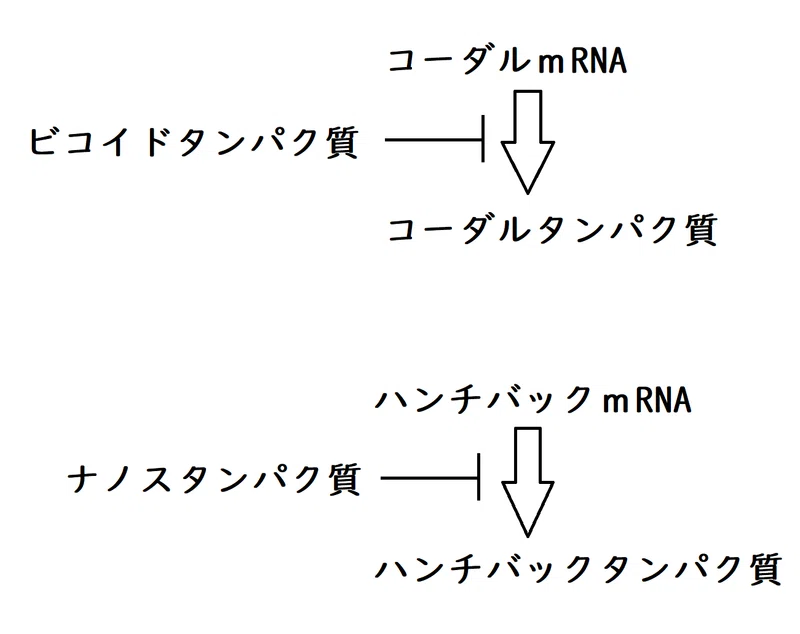
・コーダルmRNAは母性因子として卵全体に分布している。受精後、コーダルmRNAの翻訳が始まるが、その翻訳はビコイドタンパク質に阻害される。ビコイドタンパク質はコーダルmRNAに結合し、その翻訳を阻害する。
・一方、未受精卵の後端には、母性因子の一つであるナノスmRNAが局在している。
・ナノスmRNAが翻訳されてできるナノスタンパク質は、標的となるmRNAに結合する。ナノスの標的の1つがハンチバックmRNAである。ナノスタンパク質はハンチバックmRNAに結合して翻訳を阻害する。
母性因子の一つであるハンチバックmRNAは、卵全体に分布している。
ハンチバックタンパク質は転写因子であり、ギャップ遺伝子に影響を与えるモルフォゲンとして働く(ハンチバック遺伝子自体もギャップ遺伝子の一つである)。
下図はイメージ。
ナノスタンパク質はハンチバックmRNAに結合し、その翻訳を阻害する。
発展:四肢動物の肢芽とモルフォゲン
● 将来、四肢(しし。一般にヒトで言う両手足と考えてよい)を形成する隆起を肢芽(しが)という。
● 肢の形成には、ソニックヘッジホッグと呼ばれるモルフォゲンの濃度勾配が関わっていることがわかっている。
雑談:肢芽は、側板中胚葉の予定領域(肢の骨の前駆細胞になる)と、体節中胚葉(肢の筋肉の前駆細胞)から細胞が移動・増殖することで形成される。

雑談:肢芽において見られる上皮の肥厚構造をAER(外胚葉性頂堤)という。AERからは、肢の伸長に関わる因子など、様々な因子が分泌されることがわかっている。
AER直下にある肢芽の細胞を進行帯(PZ:progress zone)という。肢芽の伸長と分化は、AERと進行帯の相互作用によって引き起こされることがわかっている(進行帯では、中胚葉の盛んな細胞増殖が引き起こされ、結果として肢芽が伸長する。下図ではAERから進行帯への働きかけしか描いていないが、実際は進行帯側からもタンパク質が分泌されている)。

雑談:初期の肢芽と体壁後方境界に近い中胚葉性の領域を極性化活性帯(ZPA:zone of polarizing activity )という。極性化活性帯はモルフォゲンとして機能するShh(ソニックヘッジホッグ)というタンパク質を分泌し、発生中の肢の前後軸を決めている。肢の前後軸には、Shhの濃度勾配だけでなく、Shhに曝されている時間も影響するらしいことがわかっている。

発展:ニワトリの表皮と真皮(しんぴ)
● ニワトリの羽毛やうろこは表皮から分化するが、どちらに分化するかは真皮の誘導によって決まることが知られている。
以下は詳細(あまり気にしなくてよい)。
*皮膚は主に表皮(外胚葉由来)と真皮(中胚葉由来)からできている。

*ニワトリにおいて、背中には羽毛が形成される。肢にはうろこが形成される。
*真皮が皮膚構造をつくる司令塔となることが知られている(真皮からタンパク質[誘導物質]が分泌される)。
*真皮が由来する領域に対応した皮膚構造ができる。
①背中の真皮→表皮は羽毛を形成
②肢の真皮→表皮はうろこ等を形成

*たとえば、背中の表皮と肢の真皮からは、うろこが形成される(真皮が由来する領域に対応した皮膚構造ができる)。

雑談:羽毛は、保温、防水、外傷からの保護の他、時に装飾として発達し、同種の認識や他者に対する威嚇、異性の勧誘に機能する。
雑談:まだ形態形成が起きていない胚の皮膚をコラーゲン分解酵素で処理することで、表皮と真皮を分離することができる。
雑談:様々な動物の、様々な部位において、表皮は、毛や爪、ウロコ、ひづめになる。ただし、硬骨魚類のウロコは真皮由来である。
講義動画【表皮・真皮・肢芽・モルフォゲン等について】
雑談:上の動画についての雑談を以下に記す。
・羽毛の無い翼はイメージしにくいかもしれないが、手羽先をイメージすればよい。
・鳥の指はマウスやヒトの第2,3,4指に相当すると考えられてきたが、正しくは第1,2,3指に相当するらしいことがわかっている。
・誘導を受ける能力を「応答能(おうとうのう)」と言うが、発生のステージが進むにつれて表皮の応答能はなくなっていく(当然である。たとえば、分化の進んだ表皮に、後から真皮をくっつけたところで、何も起きるはずもないことはイメージできるだろう)。
・表皮は、真皮を集合させるシグナルを発する。
・肢を作るのに必要な形態形成の原理は、基本的にあらゆる四肢動物で共通である。哺乳類の肢芽由来の移植片は、ニワトリの肢の形成をコントロールできる。
・マウスでは、Hox遺伝子のHoxb9、Hoxc9、Hoxd9の発現パターンによってTbx5(前肢)とTbx4(後肢)が側板領域で発現する。ニワトリでは、前肢のところ(Tbx5発現領域)には翼が形成される。

・中胚葉と外胚葉の相互作用(お互いがタンパク質を分泌しあう)によって肢芽が形成される。

・極性化活性帯(ZPA)と進行帯(PZ)は一部領域が重複しているとされるが、研究中である。
・肢芽の伸長においては、中胚葉と外胚葉がお互いにタンパク質を分泌し、刺激しあっている。中胚葉と外胚葉から分泌される成長因子が、互いに細胞増殖を誘導しあっている(正のフィードバック)。
・ソニックはセガのキャラクターである。ソニックヘッジホッグの発見者がそのゲームのファンであった。
・外胚葉性頂堤=apical ectodermal ridge(AER):肢芽の先端に形成される。AERの下の間充織細胞を未分化な増殖できる状態に保ち、肢を伸長させるなどの機能をもつ。
・極性化活性帯=zone of polarizing activity(ZPA):初期の肢芽の後方部と体壁との境界に近い中胚葉性の組織の領域。ソニックヘッジホッグの作用を介して発生中の肢の前後軸を特定化する。
・進行帯=progress zone(PZ) :AERの直下にある肢芽の間充織細胞。肢芽の伸長と分化は、AERとPZの一連の相互作用によって引き起こされる。

雑談:ラットの眼の形成不全の原因遺伝子として、Pax6という遺伝子が知られていた。このPax6は、ショウジョウバエにおける眼が発生しない突然変異の原因遺伝子eyelessと非常に相同性が高いことが明らかになった。そこで、ヒトのPax6遺伝子をショウジョウバエの触角や脚の原基に導入したところ、それらの器官に眼が生じたのである。このことは、昆虫の複眼と、脊椎動物のカメラ眼(この2種の眼は、一見、似ても似つかない)が、同じ遺伝子の作用により形成されるということを表している。
発展:昆虫の脚の再生
ゴキブリやコオロギなどの昆虫は、脚を再生できる。
ゴキブリの脚はたくさんの節からつくられており、基部―先端部軸がある。そして、基部―先端部軸方向に「位置価」のようなものを持っていると考えられている。位置価が連続していない細胞同士を隣り合わせると、位置価を連続したものにするために、欠けている位置価が成長によって挿入されることがわかっている。
ゴキブリの脚を先端側で切断し、より基部側で切断した脚に移植すると、(欠けている位置価を回復するように)移植片と宿主の接合部で局所的な成長が起こり、欠けていた中央部分が挿入される(下図はイメージ。基部―先端方向に、123245の位置価を想定する。4と5の間で切断し、移植片をつくる。それを、1と2の間で切断した宿主に移植する。すると、再生が起こり、欠けている部分が挿入される。なお、ゴキブリのような昆虫では、この再生に寄与するのは主に先端側の断片である)。

もし、基部側で切った脚を、より先端側の切断部に移植して、ふつうより長い脚をつくると、欠けている位置価が再生により回復することで、脚はいっそう長くなる(下図はイメージ。脚を4と5の間で切断し、宿主とする。そして、1と2の間で切断した移植片を、宿主に移植する。すると、再生が起こり、欠けている部分が挿入される)。なお、この時、再生で生じた部分は、残りの脚とは逆向きとなっている(体表の剛毛の向きを見てみると、水色の部分だけ逆になっている)。この逆向きの再生は、位置価の勾配が逆であるために起こると考えられる。

まだわかっていないこと
● ホメオティック遺伝子の進化的な起源は何か(すべての多細胞生物[動物、植物、菌類]がHox様の遺伝子をもっている)。
雑談:「制御の理解というのは、分子生物学と進化生物学の和解に必要な中心的問題だと信じている。これら2つの生物学分野の統合が起こるとすれば、それは発生学という共通基盤の上においてであろう。」グールド(古生物学者)
● 発生の速度を調節しているものは何か。
● ギャップ遺伝子、ペアルール遺伝子、セグメントポラリティ遺伝子、ホメオティック遺伝子が、どのように制御されているかについては、わかていないことが多い。
● 発生の過程で、転写因子同士はどのように関わりあっているのか(膨大な種類の転写因子が正常な発生の進行に関わる)。これまで動物の発生について多くのことを紹介してきたが、そのほとんどについて、遺伝子の働き合いというレベルでは、完全には明らかになっていない。
雑談:以下は、ある遺伝子Aを過剰発現させた場合の各遺伝子の発現量の変化(左)と、遺伝子Aの発現を阻害した場合の各遺伝子の発現量の変化(右)である(僕の大学時代の研究データの一部)。このように、1つ1つ遺伝子間の相互作用を解析していく。(RT-PCRの産物を電気泳動に流している。バンドが濃くなったということは発現量が増加したということを表し、バンドがうすくなったということは発現量が減少したということを表している)
