遺伝(ノーベル賞特別企画)-CRISPR/Cas9-No.5

前回までで、CRISPR/Casシステムには、Adaptationフェーズ、Ecpressionフェーズ、Interferenceフェーズがあることを紹介してきました。
引用文献:新海暁男 CRISPR-Casシステムの構造と機能 生物物理54(5);2014:247-252
今回のノーベル賞で話題になった、CRISPR/Cas9は、2013年にCas9-sgRNAの系が人工ヌクレアーゼとして利用可能であることが報告されたことが発端でした。

図1 Cas9を用いたゲノム編集
まず、sgRNAについて記載する。
sgRNAは今までのCRISPR/Casシステムの中では出てきていない。
これは、single guide RNA(sgRNA)というもので、標的DNAに対して相補的な配列を持つcrRNAの3’末端にtracrRNAを連結させたものである。
引用文献:遠藤亮ほか CRISPR/Cas9によるゲノム編集技術の進展と作物への応用発展し続けるCRISPR/Cas9を用いたゲノム編集技術 化学と生物 2018;56(11):759-768
Cas9タンパク質がCRISPR/Cas9システムの中心的役割を担っている。
通常Streptococcus pyogenes(化膿レンサ球菌)由来のCas9(SpCas9)が用いられる。
ちなみに、SpCas9以外にもさまざまな細菌由来のCas9やCas9類似のRNA依存性DNAエンドヌクレアーゼCpf1(CRISPR from Prevotella and Francisella 1)も利用され始めている。
SpCas9は様的配列に隣接したNGG(N:A or C or G or T)という配列をPAMとして認識する。
SpCas9は2つのローブ(RECローブとNUCローブ)によって構成され、RECローブはgRNAと標的DNA鎖の認識に機能し、NUCローブは標的DNA鎖の相補鎖とPAMの認識および二本鎖DNA切断に働くことがわかっている。

図1 SpCas9ポリペプチドの二次構造

図2 SpCas9とsgRNAおよびDNAの複合体 三次元構造イメージ
Protospacerとは、ゲノム上の任意の狙いたい領域という意味で用いられる。
SpCas9はTypeⅡ-Aに分類され、ゲノム編集ツールとして使用される。

図3 CRISPR-Cas9によるDNA切断メカニズム
引用:CRISPR-Cas9がDNAを切断する瞬間の撮影に成功 https://www.jst.go.jp/pr/announce/20171110-2/index.html
図3の形となるために、今まではCas9-RNA複合体がDNA上をスライドして、標的配列を探していくとされていました。
しかし、引用先の報告では、3次元方向への拡散を利用したDNあへの衝突により標的配列を探索することが直接的に示された。
さらにハサミの一つであるHNHドメインが大きく由来だ構造を取ることも示された。
切断反応にはマグネシウムイオンが必要であり、HNHドメインがDNAの切断部位に移動した後、切断されたDNAがCas9から離れていく過程が判明している。
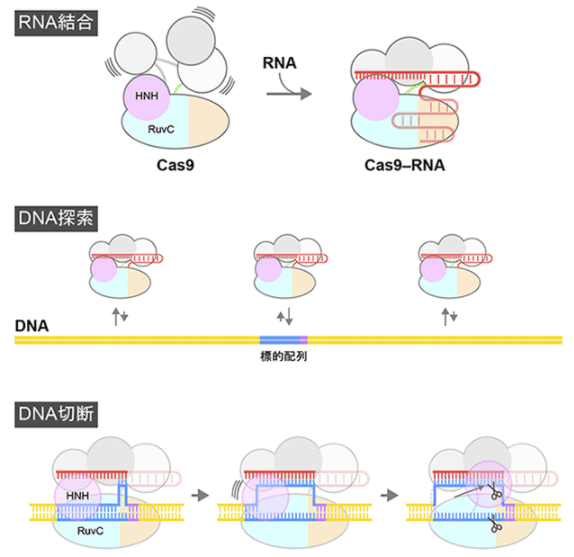
図4 CRISPR-Cas9もよるDNA切断のダイナミクス
Cas9-RNA複合体が標的配列に結合すると、DNAの巻き戻しが起こり、HNHドメインはDNAの近傍に移動し、相補鎖DNAを切断する。
一方、RuvCドメインは一本鎖となった非相補鎖DNAを切断する。
ここで、
切断された2本鎖DNAの修復の際に、様々な長さの挿入や欠失変異などを導入することができる。
その修復が、非相同末端再結合または相同組換え修復である。
CRISPR/Cas9によるゲノム編集は、これらの修復過程までを含めることで、様々な編集を行うことができるというわけである
この記事が気に入ったらサポートをしてみませんか?