
20240709: 腓腹筋内側・3次元構造・筋束構造・筋サイズ・uitarasonography

骨格筋は多様な構造プロファイルを示します。羽状筋では、筋束(筋原線維の束)が筋肉全体の作用線に対してある角度(羽状角)で配向しています。これにより、筋束の長さを犠牲にして、筋肉は所定の体積内により多くの筋束を詰め込むことができます。羽状構造は、筋肉の長さに沿って均一な筋束構造(長さと羽状角)を持つ平行四辺形としてモデル化されることがよくあります(例:Alexander & Vernon, 1975)。ただし、動物およびヒトの死体研究では、筋肉内の筋束の長さと羽状角に地域的なばらつきがあることが示されています(Huijing, 1985 ; Lee et al., 2015 ; Zuurbier & Huijing, 1993)。このような局所的な構造的多様性は、筋肉全体の力発揮能力に影響を及ぼすと考えられています (Ettema & Huijing, 1994 ; Higham & Biewener, 2011 ; Zuurbier & Huijing, 1993 )。たとえば、筋束の長さ自体の局所的多様性は、筋肉全体の逆 U 字型の力-長さ曲線を平坦化させる可能性があり (Ettema & Huijing, 1994 )、筋肉がより広範囲の長さで最大に近い力を発揮できるようになる可能性があります (Higham & Biewener, 2011 )。

これまでに、ヒトの体内で筋束構造の地域的変異を調べる試みがいくつか行われてきた。一部の研究では、内側腓腹筋(MG)内で筋束の長さは一定で羽根角は可変であることが明らかになった(Kawakami et al., 2000 ; Muramatsu et al., 2002)が、他の研究では筋束の長さは可変であることが明らかになった(Aeles et al., 2022)か、MG内でそのような構造の変異は見つからなかった(Bolsterlee et al., 2017 ; Maganaris et al., 1998 ; Narici et al., 1996)。特定の筋肉に関するこれらの矛盾する結果は、筋束構造を測定した関節構成や領域の定義の違いによるものである可能性があるが、筋束構造の地域的変異の大きさの個人差と関連している可能性がある。Aeles et al. (2022)は、束の長さの地域的変異の大きさは、性別や年齢に関係なく、個体間で大幅に異なることを実証しました。しかし、羽根の角度の変動における個体間の違いについては、あまり情報がありません。さらに、束の構造的変動における個体間の違いを決定するものは何なのかという疑問が残ります。
羽根角の個人差は、筋肉の厚さ (Brechue & Abe, 2002 ; Kawakami et al., 1993 , 2006 ) や体積 (Aagaard et al., 2001 ) などの筋肉の大きさの個人差と関連しています。以前の研究でも、運動トレーニングに反応して筋肉の大きさが増加すると、羽状角の増加が伴うことが示されています (Blazevich et al., 2007 ; Kawakami et al., 1995 )。トレーニングによって引き起こされる羽状角の増加は筋肉内で不均一であることが報告されており (Ema et al., 2013 )、大きな筋肉では筋束構造の地域差が大きいことも報告されています。筋束構造の地域差は筋肉の大きさに依存するという仮説が立てられています。これが事実であれば、以前の研究結果の矛盾を説明し、生体内の筋肉の機能と適応性に関する新たな洞察を提供できる可能性があります。私たちの仮説を検証するために、磁気共鳴拡散テンソル画像法(DTI)と、個々の筋束と筋肉全体を3Dで再構築できるトラクトグラフィーを使用して、ヒトの内側腓腹筋(MG)全体にわたる筋束構造を生体内で調べました。
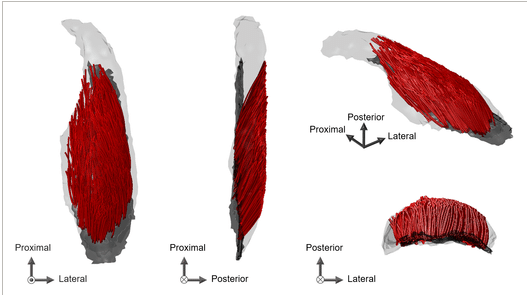

束構造の地域的変異
3D再構成された束は、矢状面と冠状面の両方で様々な長さと角度で深部腱膜から発生しており、MG内の構造の複雑さを示しています。束の長さ、2Dと3Dでの羽状角、冠状面での束の角度には、領域による有意な主効果が見られました(すべてp < 0.001)。中内側領域の束の長さは、遠位内側(p < 0.001)および遠位外側(p < 0.001)領域の束の長さよりも有意に長く、中外側領域の束の長さは、遠位内側(p < 0.001)および遠位外側(p < 0.001)領域よりも有意に長かったです 。遠位内側領域の2D羽状角は他の領域のそれよりも有意に大きく(p ≤ 0.017)、遠位外側領域の2D羽状角は中内側(p = 0.003)および中外側(p < 0.001)領域よりも有意に大きかった。中内側の2D羽状角は中外側領域のそれよりも有意に大きかった(p = 0.005)。3D羽状角は遠位内側が中内側(p < 0.001)および中外側(p < 0.001)領域よりも有意に大きかった。遠位外側領域の3D羽状角は中内側(p < 0.001)および中外側(p < 0.001)領域よりも有意に大きかった。冠状面における束角度は、遠位外側領域では他の領域よりも有意に大きく(p ≤ 0.015)、遠位内側領域では中内側領域よりも有意に大きかった(p = 0.021)


正規化された筋肉量は、中外側領域の筋束長(r = 0.615、p = 0.015)および遠位内側(r = 0.774、p < 0.001、)および遠位外側(r = 0.530、p = 0.042)領域の3D羽状角度と有意に相関していたが、他の領域のアーキテクチャパラメータ(r = −0.217〜0.487、p = 0.066〜0.941)とは相関していなかった。一方、参加者間の領域間の筋束長のSDは4.0 mm〜10.7 mmの範囲であり、2D羽状角度と3D羽状角度はそれぞれ0.9°〜5.0°、3.0°〜8.8°の範囲であった。正規化された筋肉量は、筋束長(r = 0.765、p < 0.001)、2D 羽状角(r = 0.659、p = 0.008)、および 3D 羽状角(r = 0.828、p < 0.001)の SD と有意に相関していました。

ヒトMG内では、生体内で束の長さと羽状角の両方が地域的に変化することを示しました。さらに、構造的変異の大きさは個人間で大幅に異なり、筋肉量と正の相関関係にあり、これは私たちの仮説を裏付けています。MG内の束の長さの変動は性別や年齢とは無関係であることが報告されていますが(Aeles et al., 2022)、羽状角の変動についてはあまり知られていません。私たちの結果は、筋肉内の束の長さと羽状角の変動の両方に個人差があることを同時に示し、同時にそのような構造的変動が筋肉のサイズに依存していることを示す初めての証拠です。これは、以前の研究でグループレベルで束の構造の地域的変異に関する一貫性のない発見を部分的に説明できます。例えば、平均筋肉容積が195.1 ± 38.6 cm 3 ( n = 8) のMGでは、筋束の長さは領域間で同様(すなわち、変動が小さい)であると報告されている(Bolsterlee et al., 2017)。一方、MGでは筋容積が大きく、筋束の長さは100~400 cm 3の範囲で変動が大きいと報告されている(n = 32、Aeles et al., 2022)。さらに、私たちの生体内研究の結果は、 MG 内の筋束の長さが比較的一定であると報告した死体研究 (Friederich & Brand, 1990 ; Ward et al., 2009 ) の結果を説明することができます。これは、死体標本では通常、筋肉の体積が小さい (たとえば 107.5 cm 3 [筋肉量 (Ward et al., 2009 ) を筋肉密度で割ると 1.056 g/cm 3 (Ward & Lieber, 2005 )]) です。
多数の研究により、慢性的な機械的負荷によって引き起こされる筋肥大に伴う筋束長および/または羽状角の変化が実証されている (Aagaard et al., 2001 ; Blazevich et al., 2003 , 2007 ; Duclay et al., 2009 ; Ema et al., 2013 ; Franchi et al., 2014 ; Kawakami et al., 1995 ; Reeves et al., 2009 ; Seynnes et al., 2007 )。本研究は横断的観察であり、結果の因果関係は推測の域を出ないが、本研究で観察されたように、より大きな筋肉で筋束長および羽状角の地域的変動が大きいことは、機械的負荷に対する構造的変化が単一の筋肉内で地域的に異なることを示唆している。構造適応の地域的変動は、運動中に筋線維にかかる機械的ストレスに起因する可能性があり、これは、筋電図検査 (Gallina et al., 2011 ) または機能的 MR 画像 (Kinugasa et al., 2011 ) 研究で観察されるように、局所的な神経/代謝活動と関連している可能性があります。より頻繁かつ高度に活性化された筋肉は、全体的な筋肉サイズがより大きくなる一方で、構造のより大きな地域的変動につながる可能性のある不均一な機械的ストレスにさらされる機会が多くなると推測されます。これは、より大きな筋肉で観察される筋束構造のより大きな地域的変動を説明できる可能性があります。この推測を明らかにするには、さらなる縦断的研究が必要です。
死体研究 (Huijing, 1985 ) では、筋束の長さの地域的変異が、ヒトの腓腹筋内の直列サルコメア数の地域的変異と関連していた。この研究によると、より長い筋束はより多くのサルコメアを直列に持つことができ、それによって、与えられた筋束の長さの変化に対するサルコメアの長さの変化量はより少なくなる (Ettema & Huijing, 1994 )。その結果、筋束の長さの地域的変異は、個々の筋束が異なる全体の筋肉の長さで最適な長さに到達し、全体の筋肉の逆 U 字型の力-長さ曲線を平坦化すると考えられる (Ettema & Huijing, 1994 ; Zuurbier & Huijing, 1993 )。これにより、筋肉は広範囲の全体の筋肉の長さで力を発揮することができる (Higham & Biewener, 2011 )。一方、本研究では、MG内では、より長い束はより小さな羽状角(2Dおよび3Dの両方で)を持ち、より短い束はより大きな羽状角(2Dおよび3Dの両方で)を持つ傾向があった。収縮の初期状態でより大きな羽状角は、全体の筋肉の長さの与えられた変化に対して、より大きな束の回転量とより小さな束の長さの変化につながる可能性があり(Azizi&Deslauriers、 2014;Brainerd&Azizi、 2005)、これはまた、可変の束の最適長さに寄与する。したがって、全体の筋肉の能動的な力-長さ曲線に対する束の長さと羽状角の変化の影響は、互いに相殺される可能性がある。より大きな筋肉におけるより大きな構造的変動性の機能的結果を解明するためには、束/サルコメア挙動の地域的変動とそれが力の発生に及ぼす影響についてのさらなる調査が必要である。
筋束と筋長に沿った軸の角度(筋の作用線の推定値)として計算された 3D 羽状角は、筋束と筋表面の角度として計算された 2D 羽状角よりもいくらか大きかった。さらに、3D 羽状角と 2D 羽状角では、地域による変動のパターンが異なっていた。これらの不一致は、筋束が冠状面でゼロ以外の角度を持ち、そのような筋束の冠状面の角度が地域によって異なっていたためである。これらの知見は、文献で通常使用されている 2D 羽状角が、筋長に沿った軸に対する筋束の 3D 方向またはその地域による変動を反映していないことを示唆している。したがって、3D 羽状角は筋機能に関連しているはずである。さらに、私たちの研究結果は、死体ヒト筋肉の筋束の3D配向に関するこれまでの研究結果(Chiu et al., 2020)と一致しており、筋線維の力が、筋肉の長さと厚さ(腱膜に垂直)に沿った方向だけでなく、幅(厚さに垂直)の方向にも領域特異的に伝達される可能性を裏付けています。
DTI と 3DUS ベースの構造の間には強い類似性が見られ、これは、 ラット外側腓腹筋の DTI ベースの羽状角と直接解剖学的検査で測定されたものとの間に 強い相関 ( r = 0.89) を示した以前の検証研究 (Damon et al., 2002 ) と一致しています。さらに、私たちの結果は、DTI と 2 次元 US の間でヒト MG 束の長さや遠位端の羽状角に有意差がないことを示す以前の研究結果 (Bolsterlee et al., 2015 ) と一致しています。したがって、私たちの結果は、in vivo での束構造の定量化における DTI の収束妥当性を検証しました。 3DUS では、最適な束面を特定し、束の経路を長さに沿って正確に追跡することが難しいため、計測できる束の数は限られていました (つまり、単一の超音波ボクセルアレイにつき 1 つの束)。しかし、DTI 法では、広い関心領域にわたって何千もの MG 束を自動的に生成しました。この方法間の違いは、束が複数の平面を通過することが多く、そのような束を単一の超音波平面で捉えることができないという事実によるものと考えられます。DTI ベースの束では、ほとんどの領域で測定の再現性が高く、束の長さ (CV: 4.0 ± 1.0%) と 2D 羽根角 (CV: 3.9 ± 0.7%) の平均値の再テスト差は、領域間の変動 (束の長さの CV: 16.0 ± 3.8%、2D 羽状角の CV: 12.1 ± 4.1%) よりも低かったです。一方、Aeles et al. ( 2022 )は、羽状角の信頼性が十分であることを確認できなかった。本研究では、端点が筋表面近くにあり、中点が筋表面から遠い線維のみを選択することで、誤った線維を自動的に除外した。これが、本研究の比較的高い再現性に寄与した可能性がある。しかし、DTIベースの線維束構造の再現性は、MGの近位領域では低かった。再現性の低さの原因は不明であるが、この領域に大量の神経血管路とその隣接する脂肪組織があることと関連している可能性がある。これらの組織は、拡散関連信号を減衰させ、結果として得られる線維追跡の品質に影響を与える可能性があるためである。この制限にもかかわらず、本研究の結果は、少なくともMGの腹部(中部および遠位領域)内では、ヒトの筋肉の線維束構造を生体内で定量化するためにDTIを使用することを正当化するものである。
本研究では、ヒトの MG 内では、束の長さと羽根の角度の両方が体内で変化し、3D での構造の複雑さが強調されることが実証されました。一方、束の構造の地域的変化の程度は個人間で大きく異なり、筋肉のサイズと正の相関関係がありました。これらの結果は、筋肉のサイズが束の構造の変動性に依存していることを明確に示しています。
まとめ
筋束構造(長さおよび羽状角)は、筋肉内で地域によって異なることがある。ヒトの筋肉の構造の多様性は生体内で評価されているが、個人間の差異およびその決定要因は不明である。筋内の羽状角の不均一な変化が慢性的な機械的負荷による筋肉の大きさの変化と関連していることを考慮して、我々は筋束構造の地域的差異が筋肉の大きさの個人間差異に依存しているという仮説を立てた。この仮説を検証するために、我々は磁気共鳴拡散テンソル画像法およびトラクトグラフィーを用いて、膝を完全に伸展した中立足関節位で、15人の健康な成人(男性10人および女性5人、23.7 ± 3.3歳、165.8 ± 8.3 cm、61.9 ± 11.4 kg、平均±標準偏差)の右下腿の内側腓腹筋全体に沿っておよび横切って筋束を3次元的に再構成した。 3D 再構成された束は、矢状面と冠状面の両方で長さと角度が可変で深部腱膜から発生していました。束の長さは、中間部 (中間内側: 52.4 ± 6.1 mm、中間外側: 52.0 ± 5.1 mm) の方が遠位部 (遠位内側: 41.0 ± 5.0 mm、遠位外側: 38.9 ± 3.6 mm、p < 0.001) よりも有意に長かったです。 2D 羽状角(筋肉表面に対する角度)は、中間領域よりも遠位領域で、外側領域よりも内側領域で有意に大きかった(中間内側:26.6 ± 3.1°、中間外側:24.1 ± 2.3°、遠位内側:31.2 ± 3.6°、遠位外側:29.2 ± 3.0°、p ≤ 0.017)。一方、3D 羽状角(筋肉の作用線に対する角度)については、近位遠位差のみが有意であった(p < 0.001)。これらの結果は、3D における筋束の構造的変動を明確に示している。領域間の標準偏差として評価された地域変動の大きさは、個人間で大きく異なり(筋束の長さは4.0〜10.7 mm、2D羽状角は0.9〜5.0°、3D羽状角は3.0〜8.8°)、これは体重に対して標準化された筋肉量と正の相関関係にあった(r = 0.659〜0.828、p ≤ 0.008)。これらの結果は、筋束構造の変動性が筋肉サイズに依存していることを示しています。
この記事が気に入ったらサポートをしてみませんか?