乳児の腸内細菌叢はマウスの認知能力に寄与する

メインコンテンツへスキップ
細胞宿主微生物
ログイン
検索...
論文|第31巻第12号1974-1988.e4頁2023年12月13日
全号ダウンロード
乳児の腸内細菌叢はマウスの認知能力に寄与する
https://www.cell.com/cell-host-microbe/fulltext/S1931-3128(23)00454-7

トマス・セルド 10
アリシア・ルイス=ロドリゲス 10, 11
インマクラーダ・アクーニャ 10
マーティン・フォン・ベルゲン
クリスティーナ・カンポイ
アントニオ・スアレス
すべての著者を表示
脚注を表示発行:2023年12月04日DOI:https://doi.org/10.1016/j.chom.2023.11.004
ハイライト
腸内細菌叢組成とヒスチダーゼレベルは乳児の認知と関連する
糞便中および尿中のヒスチジン代謝物は乳児の認知と関連する
腸内細菌叢はマウスの物体認識記憶を向上させる。
Inf-aboveCCマウスのPRCでは、ヒスチジンおよびグルタミン酸に対するウロカニン酸の比率が低い。
まとめ
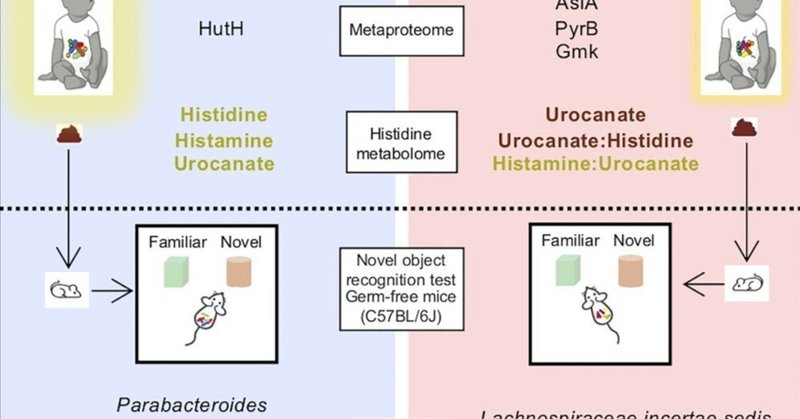
腸内細菌叢は乳児の神経発達に関連している。ここでは、乳児の複合認知と腸内細菌叢組成との関連が、早ければ6ヵ月で確立される。複合的認知力が中央値以上(Inf-aboveCC)の乳児と中央値未満(Inf-belowCC)の乳児では、より高い多様性と均等性が微生物群集を特徴付けた。メタプロテオミクスおよびメタボローム解析により、微生物のヒスチジンアンモニアリアーゼおよび乳児のヒスチジンメタボロームと認知との関連が確立された。Inf-aboveCCとInf-belowCCのドナーの糞便を無菌マウスに移植すると、新奇物体認識テストで評価される記憶力が伝達形質であることが示された。さらに、Inf-aboveCCマウスは、Phocaeicola属に属する種に富み、以前から認知に関連していたBacteroides属やBifidobacterium属にも富んでいた。最後に、Inf-aboveCCマウスは、Inf-belowCCマウスと比較して、糞便中のヒスチジン、ウロカニン酸:ヒスチジン、ウロカニン酸:グルタミン酸比が低い。全体として、これらの知見は、腸内細菌叢が乳児の認知に関与していることを明らかにし、ヒスチジン代謝産物レベルの調節が潜在的な基礎メカニズムであることを指摘している。
図解抄録
図サムネイルfx1
大きな画像を見るダウンロード 高解像度画像
キーワード
認知
腸
乳児
微生物叢
無菌マウス
ヒスチジンメタボローム
メタプロテオミクス
はじめに
生後1年間は、複雑な微生物生態系が新たに構築される重要な時期である1。腸内細菌叢によるコロニー形成は、グリアの分化、軸索の髄鞘形成、シナプス形成など、出生後の脳発達のダイナミックな段階や、神経発達機能の急速な出現と同時に起こる2。その後、腸内微生物の代謝経路の代謝ランドスケープや微生物叢由来分子のレパートリーだけでなく、あまり異質でないコミュニティ内の常在菌種の多様性も年齢とともに増加する3。この時期、常在細菌叢は免疫学的、内分泌学的、神経活性的、代謝的な経路を通じて脳とコミュニケーションし、気分や行動に影響を及ぼすことから、微生物叢-腸脳軸の存在が示唆されている4。この軸が生後早期の神経発達に果たす役割は、齧歯類モデルで示されており、腸内細菌叢の形成不全や無菌(GF)マウスにおけるその不在は、中枢神経系機能や認知・社会行動に影響を及ぼす神経伝達物質や脳容積を変化させる5,6,7。GFマウスの神経症状や行動の変化は、離乳後6週目または10週目に常在腸内細菌叢をコロニー形成した場合に部分的に回復したが、14週目では回復しなかったことから、脳回路の初期プログラミングがその後の行動に必要であることが示唆されている8,9,10。微生物組成の変化は、子どもの気質13,14,15、情動・注意行動、不安・抑うつ、身体症状16,17、恐怖行動を反映している18,18。小児集団にとって重要な自閉症における認知障害は、腸内細菌組成および代謝の変化と関連している19。
腸内細菌叢と神経発達の関係は存在するが、その根底にあるメカニズムはまだ特定されていない。本研究では、満期産の健康な生後6ヵ月の乳児の腸内細菌叢プロファイルを比較するフォワードアプローチを試験し、メタプロテオミクスプロファイルを作成し、ヒスチジン代謝産物に焦点を当てたメタボローム解析を行い、乳児の腸内細菌叢をGFマウスに移植することで因果関係を確立し、行動結果に影響を及ぼす可能性のある数種類に絞り込んだ。この戦略により、本明細書では、GFマウスに移植した場合に記憶機能に影響を与える腸内細菌群集と乳児の認知能力との機能的関連について報告する。行動と腸内細菌叢の関連性の動態を明らかにすることは、乳児の神経発達を促進する革新的な微生物叢ベースの治療法の開発につながる可能性があるため、極めて重要である22。
結果
研究集団の神経発達スコア
私たちの神経発達学的分析は、Bayley Scales of Infant Development, Third Edition(BSID-III)を中心に行われた。乳児は、BSID-IIIの3つの個別領域(複合認知(CC)、言語(受容性言語、表出性言語、複合言語)、運動(粗大運動、微細運動)の得点により、典型的な健康な神経発達の閾値を満たしていた。BSID-III得点の中央値(範囲)は以下の通りであった: CC(110[85-125])、受容言語(12[5-15])、表出言語(10[5-15])、複合言語(109[83-129])、粗大運動(9[2-17])、微細運動(12[5-17])であった(表S1)。乳児は、BSID-IIIの各スケールの中央値(50パーセンタイル)以上と未満の2群に二分された。
腸内微生物群集の多様性、構造、および組成は乳児の認知グループ間で異なる
我々は、腸内細菌叢が生後6ヵ月という早い時期に言語、運動、認知の各スコアと関連するかどうかを明らかにしようとした。健康な乳児から6ヵ月目に糞便サンプルを採取し、16S rRNA遺伝子のハイスループットアンプリコンシークエンシングにより腸内微生物組成の特徴を明らかにした。品質フィルタリング後、4,435,206のリード配列から、46科に属する102の別個の属に絞られた645の種レベルの細菌からなる腸内細菌プロファイルが得られた(図S1;表S2A)。以前の報告23,24と一致するように、リボソームデータベースプロジェクト(RDP-II)に対する分類学的分類では、Bacillota(479)に属する分類群にコミュニティメンバーが多く、Bacteroidota(101)、Pseudomonadota(30)、Actinomycetota(23)、未同定の細菌門(unclass_Bacteria)(8)、Fusobacteriota(4)が続いた。最も多かった属は、Lachnospiraceae属(unclass_Lachnospiraceae)、Enterobacteriaceae属(unclass_Enterobacteriaceae)、Bacteroides属、Lachnospirace incertae sedis属、Enterococcus属で、全リードの61.4%を占めた。乳児の腸内細菌叢では、サンプル間の変異が大きかった。unclass_Enterobacteriaceaeに分類された1種のみが全サンプルに存在し、総リードの4.84%を占めた。25種が高頻度(全配列リードの1%以上)であったが、そのうち6種のみが高頻度(サンプルの90%以上)であった。これらの高頻度・高存在種は、Bacillota属、Bacteroidota属、Pseudomonadota属に属していた。
まず、腸内微生物群集の多様性と構造の測定値が、7つのBSID-III尺度それぞれにおいて中央値以上または以下に分類された幼児間で異なるかどうかを検証した(表S3)。すべての種を考慮すると、腸内微生物群集構造と強い関連を示したユニークなBSID-III尺度はCCであった。微生物のα多様性指標では、CCスコアが中央値以上の乳児(Inf-aboveCC)の腸内細菌叢は、CCスコアが中央値未満の乳児(Inf-belowCC)と比較して、均等性、シャノン多様性、シンプソン多様性が高く、優性性が低いという特徴が示された(図1A)。分類群数(リッチネス)とフェイスの系統的多様性には、CC群間で差は認められなかった。種レベルでは、構成比を補正してβ多様性を評価するAitchison距離を用いた冗長性分析により、乳児CC群間の微生物群集構造に有意差が認められた(図1B)。属レベルでは、種の存在量の非類似性を考慮するBray-Curtis距離を用いて、乳児CC群の微生物プロファイル間の差が有意であった(図S1B)。最後に、距離ベースのPERMANOVA検定を用いて、全体的な微生物叢組成の個体間変動のうち、人体計測、周産期、母親、および栄養変数によって説明できる割合を求めた。種レベルでは、新生児の体重、性別、母親の年齢、母親の妊娠前BMI、母親のIQ、妊娠糖尿病(あり/なし)、分娩の種類(帝王切開、経膣)、妊娠中の飲酒(あり/なし)、妊娠中の喫煙(あり/なし)、3カ月目または6カ月目までの授乳日数、3カ月目または6カ月目までの母乳の種類(粉ミルク、混合、母乳のみ)との有意な関連は観察されなかった。属レベルでは、腸内細菌叢の変動の有意な説明効果が観察され、母親の年齢と妊娠糖尿病がそれぞれ全分散の8.53%と9.41%を占めた(図S1BおよびS1C)。共変量の影響は、必要に応じて下流の統計解析で考慮した。
図サムネイルgr1
図1Inf-aboveCC児(上)とInf-belowCC児(下)では、腸内微生物群集の多様性、構造、組成が異なる。
キャプション
大きな画像を見るダウンロード 高解像度画像
CCパフォーマンスの特徴的な分類群
腸内微生物群集の多様性と構造が乳児CCグループ間で異なっていたため、DESeq2を用いてInf-aboveCCグループとInf-belowCCグループ間で存在量に差のある分類群を同定した。Inf-aboveCCの乳児の腸内細菌叢はバクテロイーダに富み、バチロータに枯渇していた(図1C)。その結果、Inf-aboveCCの乳児では、Inf-belowCCの乳児と比較して、Bacillota:Bacteroidota比が有意に低かった(図1C)。属レベルでは、Inf-aboveCCの腸内細菌叢は、Inf-belowCCの乳児と比較して、バクテロイデス属、連鎖球菌属、パラバクテロイデス属、クロストリジウム属(Clostridium sensu stricto)、ホルデマネラ属(Holdemanella)、ルミノコッカス属(Ruminococcus)2が豊富であり、ラクノスピラセ属(Lachnospirace incertae sedis)、ヘモフィルス属(Haemophilus)、ラクトコッカス属(Lactococcus)が有意に減少していた(図1D、1E、S1D)。母親の共変量寄与を除いた一般化線形モデルを当てはめると、BacteroidesとLachnospirace incertae sedisの相対存在量と乳児の認知との間に有意な関連が確認された(図1F)。種レベルでは、12種が乳児のCC群間で有意に豊富であった。Inf-aboveCCの乳児では、10種がBacteroidiaとClostridiaクラスに濃縮され、ErysipelotrichiaとNegativicutesクラスからは1種が検出された。Inf-belowCCの乳児の腸内細菌叢では、BacilliクラスとClostridiaクラスの代表的な1つが濃縮されていた(図1GおよびS1E)。分類学データベースのrDNA配列の割り当て(表S4A)から、これらの種はPhocaeicola dorei、Phocaeicola vulgatus、Bacteroides caccae、Bacteroides stercorisの培養可能株と配列同一性(類似度96%以上)を共有していた、 Inf-aboveCC微生物叢にはParabacteroides bouchesdurhonensis、Streptococcus lutetiensis、Streptococcus gallolyticus、Holdemanella porci、Ruminococcus bromii、Veillonella ratti、Inf-belowCC微生物叢にはLactococcus lactisとSellimonas intestinalisの菌株を用いた。ラクトコッカスを除き、これらの菌株のメンバーは、ヒトや動物モデルにおける記憶や認知能力に関連している25,26,27,28,29,30。
メタプロテオームの系統分類
ヒトにおける微生物叢と健康転帰との関係をよりよく理解するために、メタプロテオーム解析を用いて、乳児の腸内細菌叢内で発現したタンパク質の大規模な機能プロファイリングを行った31。ペプチドは6,063の細菌タンパク質群に分類され(表S2B)、50科111属に明確に分類された(表S2C)。腸管メタプロテオームへの群集組成の寄与は、乳児のCCグループとの強い関連性を示さず(図S2A)、細菌タンパク質グループの系統学的起源における個体間の高い類似性を示唆した。メタプロテオームの系統的割り当てを拡大すると、腸内メタプロテオームに対する放線菌門の中のEggerthellaの寄与は、CCと負の相関を示した(多重検定補正を加えたウィルコクソン順位和検定でp < 0.05)。
メタプロテオーム濃縮解析により、乳児の微生物叢における認知に特異的な機能シグネチャーが明らかになった。
メタプロテオームの機能解析では、cluster of orthologous groups (COGs)分類を用い、3つの階層に階層化され、各階層はより具体的な機能割り当て(メイン、セカンダリー、ファンクションカテゴリー)になっている。オーソログタンパク質の機能的冗長性により、6,063の細菌タンパク質グループが656の非冗長なCOG機能に絞り込まれた(表S2D)。サンプルあたりのCOG機能の平均数は176[73-279]であった。サンプル間のメインCOG層とセカンダリーCOG層の全体的な分布は、最も豊富なセカンダリーCOGが代謝メインカテゴリーに属するという、かなり類似したパターンを示した(図S3A)。24のCOG機能が乳児メタプロテオーム(コア)の90%に見られ、12種類のトランスポーターと膜タンパク質、および糖質、アミノ酸、エネルギー代謝に関与する12種類の酵素で構成されていた(表S2E)。これらのコア酵素のうち5つは、成人のメタプロテオームにも非常に多く存在することがわかった:ホスホケトラーゼ、グルタミン酸デヒドロゲナーゼ、エノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、フコースイソメラーゼ。これらの結果は、腸内細菌叢が発現するタンパク質群の機能プロファイルに関する過去の報告と一致している24,31,32。
次に、腸内細菌叢の機能的能力が乳児CC群間で異なるかどうかを調べた。19.8%と25.1%という高い割合のCOG機能が、それぞれInf-aboveCCとInf-belowCCのメタプロテオームでのみ発現していた(図S3B)。Inf-aboveCC児の腸内細菌叢は、細胞内輸送分泌および小胞輸送に関与するタンパク質に富んでいた(p < 0.027)が、Inf-aboveCCメタプロテオームでは糖質輸送および代謝に関与するタンパク質が高い傾向があった(p < 0.065)(図S3C)。sPLS-DAプロットは、乳児メタプロテオーム間の明確な識別を示し(図2A)、識別に寄与するタンパク質群を同定した(図2B)。有機代謝産物の硫黄修飾に関与するアリールスルファターゼA(AslA)、ピリミジン生合成の第一段階に関与するアスパラギン酸カルバモイル基転移酵素(PyrB)、グアノシンモノヌクレオチドのリサイクルに関与するグアニル酸キナーゼ(Gmk)、ヒスチジンの非酸化的脱アミノ化を触媒するゲート酵素であるヒスチジンアンモニアリアーゼ(HutH)に有意な存在量の差が観察された。ヒスチダーゼとしても知られるHutHは、Inf-aboveCCのメタプロテオームで有意に濃縮された唯一のタンパク質であった。これらの観察は、DESEq2差分存在量分析(図2C)および多重検定補正を用いたWilcoxon順位和検定(図S3D)を用いてさらに確認された。これらのタンパク質に関連する経路に関与する酵素をメタプロテオームで調べたところ、Inf-aboveCCメタプロテオームでは、ヒスチジン異化の2つの下流酵素、ウロカナートデヒドラターゼ(HutU)とイミダゾロンプロピオナーゼ(HutI)の存在量が高いことが観察されたが、幼児群間の差は統計的に有意ではなかった。この結果は、乳児における認知に関連したメタプロテオームシグネチャーを明らかにした。
図サムネイルgr2
図2メタプロテオーム濃縮解析により、乳児の微生物叢における認知に特異的な機能シグネチャーが明らかになった
キャプション
大きな画像を見るダウンロード 高解像度画像
乳児-aboveCC微生物叢における高い腸内細菌HutH活性
HutH、HutI、およびHutUタンパク質は、神経伝達物質ヒスタミンの前駆体として機能するヒスチジンとの関連から興味をそそられた。33,34,35,36。タンパク質の存在量が実際に酵素活性を反映しているかどうかを評価するために、糞便中の細菌から抽出したタンパク質を用いて、HutHによるヒスチジンのウロカナートへの変換を測定した。その結果、Inf-aboveCCの腸内細菌叢では、Inf-belowCCの腸内細菌叢と比較して、HutH活性が1.3倍高いことが確認されたが、その差は統計学的に有意ではなかった(図S3E)。
ヒスチジンのメタボロームと乳児のCCとの関連
HutHに関するメタプロテオミクスおよび生化学的結果から、乳幼児におけるCCがヒスチジンのメタボロームの変化とどの程度関連しているかを調べることになった。我々は標的メタボローム解析法を開発し、乳児の腸内微生物と糞便から抽出したヒスチジン、ヒスタミン、ウロカネート、酢酸イミダゾール、プロピオン酸イミダゾール、N-アセチルヒスタミン、グルタミン酸のレベルを定量した(図3A)37。ヒスタミンとN-アセチルヒスタミンは細胞内ではほとんど検出されなかった。ヒスチジン、イミダゾールプロピオン酸、グルタミン酸、ウロカナートの細胞内レベルは、糞便中のレベルと有意に相関していた(図S4A)。糞便中のウロカネートレベルは、Inf-aboveCC児ではInf-belowCC児に比べて有意に低かったが(図3B)、その他のヒスチジン代謝物については差が認められなかった(図S4B)。腸からの代謝産物は循環に吸収され、最終的に宿主によって共代謝され尿とともに排泄されることから、腸内微生物の代謝がCCに関連するヒスチジン代謝産物を調節するという仮説を立て、乳児の尿サンプル中のヒスチジン代謝産物レベルを調べた。その結果、ヒスタミン、ヒスチジン、ウロカネートの尿中濃度は、Inf-aboveCCの乳児ではInf-belowCCの乳児に比べて有意に高かった(図3CおよびS4C)。また、2つの代謝産物間の比も計算した。その理由は、変化が、生物学的システムまたは神経発達障害に関連する経路の摂動を示唆している可能性があるからである。さらに、CCを識別する分類群について検討したところ、糞便中のウロカネートはunclass_Lachnospiraceae_41と正の相関を示し、糞便中のヒスチジンはRuminococcus_53と正の相関を示し、Lachnospirace incertae sedis属と負の相関を示した。糞便中のウロカネート/ヒスチジン比は、unclass_Lachnospiraceae_41およびLachnospirace incertae sedis属と負の相関を示し、Bacteroides_3と正の相関を示した(図3F)。尿中では、ヒスチジンのレベルはStreptococcus属_42およびBacteroides属と正の相関を示し、ウロカネートのレベルはHoldemanella属_16と負の相関を示し、Streptococcus属と正の相関を示した(図3G)。最後に、尿中ヒスタミン/ウロカネート比はStreptococcus属と負の相関を示した。
図サムネイルgr3
図3ヒスチジンメタボロームと乳児の複合認知機能との関連
キャプション
大きな画像を見るダウンロード 高解像度画像
Inf-aboveCCドナーの糞便をGFマウスに移植すると、Inf-belowCCドナーのものより優れた記憶機能が促進される。
微生物叢とヒトの健康との関連性に関する研究は強力であるが、乳児の神経発達における腸内細菌叢の関与をさらに立証するには、原因究明のための実験が必要である42。腸内細菌叢が認知に寄与しているかどうかを調べるため、糞便微生物叢移植(FMT)実験を行い、(1)記憶機能の伝達性を調べ、(2)マウスの腸内細菌プロフィールと代謝を比較解析し、(3)記憶機能と関連する種を絞り込み、(4)記憶反応とヒスチジン代謝産物の変化を関連付けた。Inf-aboveCC(n=8)およびInf-belowCC(n=8)ドナー由来の16の糞便サンプルをGFマウス(n=32)に移植した。ドナーの糞便サンプルは、乳児CCの記録とCCを識別する特徴(α多様性値、特徴的分類群、プロテオーム量、ヒスチジン代謝物レベル)に基づいて、各乳児ドナーをランク付けすることにより、合理的に選択した。
GFマウスに乳児腸内細菌叢をコロニー形成させた結果、行動に違いが生じたかどうかを調べるため、FMT後2週目に、オープンフィールドテスト(OFT)と新規物体認識テスト(NORT)という2つの確立された行動テストを実施した。OFTでアリーナを探索する慣れは、通常、マウスがアリーナの周辺部を探索する時間(通常、壁に接触して探索する時間(thigmotaxis))が、無防備な中央部よりも有意に長いため、探索行動と運動能力を評価することができる。OFT後、NORTは、訓練セッションで探索された2つの同じ物体のうち1つを置き換えた新しい物体の視覚的探索をテストセッションで比較することにより、記憶学習を評価した。OFTに関して、Inf-belowCC微生物叢を摂取したマウスは、Inf-aboveCC微生物叢を摂取したマウスと比較して、中心部での不動時間が増加した(図4A)。アリーナの中心部と周辺部では同じ時間を過ごし、平均速度に差はなく同程度の距離を移動したため、これらの差は運動障害の結果ではなかった(表S5)。NORTでは、Inf-aboveCC微生物叢を摂取したマウスは、Inf-belowCC微生物叢を摂取したマウスよりも有意に優れた識別指数を示した(図4B)。マウスの識別指数と乳児ドナーの体格、周産期、母体および栄養変数との関連は観察されなかった。このヒト化行動モデルにおいて、我々の結果は、記憶機能が乳児の腸内細菌叢を介して伝達されることを示し、因果関係を示唆した。
図サムネイルgr4
図4Inf-aboveCCドナーの無菌マウスへの糞便移植は、Inf-belowCCドナーからの移植よりも優れた記憶機能を促進する。
キャプション
大きな画像を見るダウンロード 高解像度画像
乳児腸内細菌叢をコロニー形成したマウスでは、記憶機能と関連する細菌分類が異なる
微生物叢移植の妥当性とCCとの関連を評価するために、FMT実験の各マウスから新鮮な糞便サンプルを採取した。ドナーの乳児とマウスのサンプルからは、398の種レベルの細菌を含む1,071,999のリード配列が得られ、高信頼性の系統的注釈の結果、93科に属する196の別個の属に絞り込まれた(表S6)。予想されたように、ドナー微生物叢とレシピエント微生物叢の間でα-およびβ-多様性のシフトが観察された。マウスに移植した場合、細菌種の減少が観察されたが、これはおそらくサンプル処理および/または種/宿主不適合の結果であろう(図S5A)。平均して、ドナー微生物叢の細菌種の69%が、対応するヒト化マウスのペアに存在した。BacteroidotaとVerrucomicrobiotaの分類群は増殖したが、ActinomycetotaとPseudomonadotaの分類群はマウス腸内で増殖しなかった。ドナーのCCスコアでビン分けすると、Inf-aboveCCの微生物相は、Inf-belowCCの微生物相を持つ微生物相と比較して、均一性が高く、シャノン多様性指数とシンプソン多様性指数が高い傾向を示した(図S5B)。Aitchison距離を用いたβ-多様性解析では、ドナー微生物叢とレシピエントマウスサンプルは異なる群集構造を有し(図S5C)、CCによって有意にクラスタ化された(図5A)。マウス群間の微生物群集構造の違いは、属レベルのBray-Curtis距離でも説明された(図S5D)。Inf-aboveCCマウスの門レベル組成は、Inf-belowCCマウスと比較して放線菌門が有意に濃縮され、バチロタが枯渇していた(図5B)。機械学習のノンパラメトリック分類・回帰ツールであるランダムフォレスト(RF)を用いて、マウスグループを区別する属を探索した。RFを用いて80%のサンプルでモデルを学習させたところ、76.2%の精度が得られ、残りの20%のサンプルでテストしたところ、マシューズ相関係数(MCC)は83%、曲線下面積(AUC)は84.6%であった(図S6A)。次に、特徴選択プロセスを用いて、関連する属を同定した。放線菌門、バクテロイデス門、およびバチロタ門に属する9属は、予測精度を96.9%に、AUCを100%に向上させた。記憶機能を識別する属は、モデル精度への寄与度が高い順に、unclass_Erysipelotrichaceae、Parabacteroides、unclass_Lachnospiraceae、Anaerostipes、Lacrimispora、Bifidobacterium、Hungatella、Eubacterium、Lachnospirace incertae sedisであった(図5C)。DESEq2を用いた存在量の差分析では、Inf-aboveCCマウスとInf-belowCCマウスの間で、unclass_Erysipelotrichaceae、Parabacteroides、Bifidobacterium、Anaerostipes、Hungatella、Lacrimispora、Lachnospirace incertae sedisに有意差があることが確認された(図5D)。RFをマウスグループの識別に使用した場合、テストデータセットとして確保された20%のサンプルで実行された予測は、88.5%の精度を示し、MCCは93.3%、AUCは91.5%であった(図S6B)。トップダウン・クロスバリデーションテストでは、予測精度96.1%、MCC100%、AUC99.4%で、モデルを12の関連する識別種に絞り込んだ。判別微生物のほとんどはバクテロイディア(Bacteroidia)クラスとクロストリジア(Clostridia)クラスに属し、アクチノバクテリア(Actinobacteria)クラスの代表は2種、うどんこ病(Erysipelotrichia)クラスの代表は1種であった(図5E)。これらの観察結果は、DESEq2解析によってさらに裏付けられた(図5F)。Lachnospiraceae_incertae_sedis_7とAnaerostipes_42を除き、Inf-aboveCCマウスではInf-belowCCマウスと比較して判別可能な微生物が濃縮されていた。最も近い培養可能な近縁種(類似度96%以上)は、BacillotaのClostridium innocuum株、Ruminococcus gnavus株、Lacrimispora xylanolytica株、Anaerostipes caccae株、Phocaeicola dorei株、Phocaeicola vulgatus株である、 BacteroidotaではBacteroides xylanisolvens、Bacteroides luhongzhouii、Bacteroides faecichinchillae、Parabacteroides distasonis、ActinomycetotaではBifidobacterium pseudocatenulatumとBifidobacterium longumであった(表S4B)。総合すると、Inf-aboveCCおよびInf-belowCCの腸内細菌叢を受容したGFマウスは、マウスの記憶機能を識別する異なる微生物プロファイルを保有していた。
図サムネイルgr5
図5乳児腸内細菌叢をコロニー形成したマウスは、記憶機能に関連する異なる細菌分類群を保有している
キャプション
大きな画像を見るダウンロード 高解像度画像
微生物叢はヒスチジン代謝物レベルを調節する
ヒスチジン代謝物と乳児のCC転帰との関連に基づき、腸内細菌叢がヒスチジン代謝物を調節する可能性があるかどうかを検証した。我々は、FMT実験から得られたヒト化マウスの糞便、血漿、尿、および脊髄周囲皮質(PRC)サンプル中のヒスチジン代謝物の標的メタボロームプロファイリングを利用した。マウスのマトリックスからすべての代謝物を検出した。N-アセチルヒスタミンを除き、マウスのヒスチジン代謝物の細胞内レベルと糞便レベルには有意な相関が認められた(図S6C)。次にRFを用いて、ヒスチジンのメタボロームがマウスの記憶機能の予測因子であるかどうかを評価した。マウスのヒスチジン代謝物のメタボロームプロファイルは、予測精度52%、MCC50%、AUC62.3%で記憶機能を識別した(図S6D)。特徴選択プロセスを実行した後、2つの代謝物、糞便中のヒスチジンレベルとPRC中のウロカネートレベルが引き続き関連し、マウスグループの識別に大きく寄与し、モデルの予測精度を68%、MCCを66.7%、AUCを69%に高めた(図6A)。糞便中のヒスチジンレベルは有意に低かったが(図6B)、PRC中のウロカネートレベルはInf-aboveCCマウスとInf-belowCCマウスで差がなかった。ヒスチジンのメタボローム比の場合、予測精度は50.0%、MCCは50%、AUCは51.8%であった(図S6E)。特徴選択プロセスでは、モデル精度をAUC 81.1%まで高める2つの比率、すなわち、Inf-aboveCCマウスではInf-belowCCマウスに比べて有意に低いPRCのウロカナート/ヒスチジン比およびウロカナート/グルタミン酸比が関連因子として同定された(図6C)。最後に、記憶パフォーマンスを識別するヒスチジン代謝物のレベルが細菌分類と相関するかどうかを検証した。スピアマンの相関から、糞便ヒスチジンはPhocaeicola_13、unclass_Erysipelotrichaceae_22、unclass_Bacteroidaceae_150、およびunclass_Erysipelotrichaceae属と正の相関があり、Anaerostipes_42およびAnaerostipes属と負の相関があることが示された(図6D)。PRCのウロカナート/ヒスチジン比とウロカナート/グルタミン酸比には、バクテロイデス属に属する種との間に負の相関が観察された。また、HutH活性を測定したところ、Inf-aboveCCの腸内細菌叢を受け取ったマウスでは、Inf-belowCCの腸内細菌叢を受け取ったマウスに比べて高い傾向が見られた(図S6F)。これらの結果から、腸内細菌叢がマウスのヒスチジン代謝産物を調節している可能性が明らかになった。
図サムネイルgr6
図6微生物叢はヒスチジン代謝物レベルを調節する
キャプション
大きな画像を見るダウンロード 高解像度画像
考察
本研究では、腸内細菌叢の組成のばらつきが、生後6ヶ月の乳児の認知能力と有意に関連することを示した。さらに、腸内メタプロテオームの比較から、ヒスチジン異化のゲートキーパー酵素であるHutHを含む認知関連シグネチャーが同定された。HutHレベルの違い、および糞便中と尿中のヒスチジン代謝物の違いは、乳児の認知機能と関連していた。最も重要なことは、乳児ドナーの腸内細菌叢の糞便移植による記憶機能がGFマウスに伝達されたことである。認知機能の高い乳児の腸内細菌叢を持つマウスは、認知機能の低い乳児の腸内細菌叢を持つマウスと比較して、高い記憶機能を示し、Phocaeicola属、Bacteroides属、Bifidobacterium属などの分類群に富み、ヒスチジンのメタボロームプロファイルも異なっていた。これらの結果は、腸内細菌叢が、おそらくヒスチジン代謝の調節を介して、乳児の認知能力に影響を及ぼす可能性を示唆している。
乳児の腸内細菌叢と認知能力との関連については、これまでにいくつかの報告がある。Carlsonら43およびTamanaら44は、Bacteroidesが優勢な腸内細菌型にクラスター形成された1歳および2歳の乳児は、FaecalibacteriumやRuminococcaceaeが優勢な腸内細菌型にクラスター形成された乳児よりも、学習複合スコアが良好であったと報告している。一方、Rothenbergら(45)およびSordilloら(46)は、Faecalibacterium属、Sutterella属、ClostridiumクラスターXIVa属の存在量と乳児の認知力には正の相関がある一方、Lachnospiraceae属の存在量には逆の相関があることを示した。これらの研究において、α-多様性の指標は認知と逆相関しているか、あるいは相関していなかった。今回明らかになった乳児の認知と関連する特徴的な腸内細菌叢は、これらの先行研究を部分的に再現するものである。我々のデータでは、認知に関連する腸内微生物の生態系がより多様で均一であることが示されたが、認知能力の高い乳児の腸内細菌叢はバクテロイデスの濃縮を示す一方で、Lachnospirace incertae sedisでは枯渇を示し、Bacillota:Bacteoroidota比が低かった。研究間の格差は、民族、乳児の年齢、乳児の神経発達評価、心理テスト、サンプルサイズに対する微生物叢のサンプリングの違い、および識別分類群を同定する統計的アプローチの違いに起因している可能性がある。それでもなお、我々の知見とこれまでに報告された結果は一貫して、成長初期の重要な時期における乳児の認知能力と腸内細菌叢との関連を示している。腸内細菌プロフィールと認知能力との関連は、乳児や母親の共変量とは無関係であった。
常在腸内細菌叢が神経学的発達に影響を及ぼすことの原因証明は、gnotobiotic動物モデルで確立されている7。微生物叢のないマウスや抗生物質誘発性ディスバイオージスのマウスでは、神経学的および行動学的異常の複数のシグナルが認められた47。周産期は、腸内微生物のコロニー形成と神経細胞組織との共進化によって特徴づけられる重要な発達段階である。関連性から因果関係へとスケールアップするために、生後6ヵ月の満期健康児の腸内細菌叢をGFマウスに移植した。その結果、腸内細菌叢が記憶を促進する役割を果たすことが示された。さらに、これらのマウスは馴化課題中にアリーナ中央での不動が減少しており、これは探索行動が優れていることの表れであると解釈できる。多くの菌種が失われたにもかかわらず、記憶機能は乳児からマウスへと受け継がれ、非常に多様な組成の中で、代謝の可塑性と冗長性により、腸内細菌叢の機能的安定性と回復力が高いという仮説がさらに支持された51。それでもなお、乳児の認知機能に対するLachnospirace incertae sedisとBacteroidesの関連は、ヒト化マウスでも再現された。Lachnospirace incertae sedisは、系統学的に異種混合の未培養Candidatus属であり、大うつ病性障害のヒト被験者から糞便を移植したマウスにおいて、うつ病様行動と関連している52,53。この知見は、R. gnavus ATCC 29149をGFマウスに単独接種しても記憶機能が改善しなかったというColettoらの報告からも支持される54。一方、Bacteroides属およびBacteroides属とPhocaeicola属に属する種は、認知機能の改善と関連していた。幼児マウスとレシピエントマウスにおいて、認知機能を向上させたのは、Phocaeicola doreiとPhocaeicola vulgatus(旧Bacteroides doreiとBacteroides vulgatus)であった。認知機能障害を特徴とするアルツハイマー病と自閉症のモデルマウスでは、バクテロイデス・オバタス(Bacteroides ovatus)またはバクテロイデス・フラギリス(Bacteroides fragilis)の補給が認知障害を救うことが示されている。GFマウスおよびアルツハイマー病マウスにおける機能的記憶に対するビフィズス菌の有益な効果は、それぞれLukら57およびAbdelhamidら58によって報告されている。今回の実験により、微生物による影響に因果関係があることが証明されたため、本研究で同定されたPhocaeicola株とBifidobacterium株を分離し、記憶機能に対する有効性を検証したくなる。認知は、腸内細菌叢の影響を受ける可能性のあるユニークな神経発達反応ではない59。我々のFMTの結果や、腸内細菌プロファイルを早期の行動転帰と関連付ける研究60,61,62,63,64,65に基づき、4つの心理学的領域、すなわち社会的相互作用、ストレスと不安、学習と記憶、運動制御の発達に寄与する特定の腸内細菌叢種または表現型的に選択されたコンソーシアムを同定するためのさらなる研究が必要である。このような微生物は、乳幼児の神経心理学的欠損を改善する治療法として役立つ可能性がある。
私たちの研究では、乳児の認知のメタプロテオミクス的特徴から、多くの識別タンパク質、主にトランスポーターと酵素が明らかになったが、中でもHutHは、ヒスチジンのウロカナート(His-Uro-Glu)を介したグルタミン酸への異化を制御することから、際立っていた。ヒスチジンの異化は神経伝達物質ヒスタミンを産生し、グルタミン酸で終わるが、これらはすべて記憶と認知能力に関連すると報告されている。さらに、アルツハイマー病、自閉症、うつ病の被験者は、ヒスチジン代謝産物の変化を伴う認知機能障害を経験した69,70,71,72。メタボローム解析を用いて、我々は、ヒスチジン、ヒスタミン、ウロカネートの濃度が、認知能力の高い乳児と低い乳児の間で差があることを観察した。Matsumotoらによる以前の研究では、特定の病原体を含まない腸内細菌叢でGF腸内細菌叢を回復させると、糞便中のヒスチジンレベルが有意に低下し、ウロカネートレベルが回復することが示され、微生物由来であることが示された73。この報告と一致して、GFマウスに乳児用腸内細菌叢をコロニー形成させると、糞便中のヒスチジン代謝産物レベルが変化し、記憶機能に関連する嗅覚野周囲のウロカネートおよびHis-Uro-Glu比が変化することがわかった。最近、運動皮質と海馬におけるHis-Uro-Gluの活性化が、運動学習や認識記憶などの行動を促進することが報告された74。これらの結果を総合すると、腸内細菌叢が、宿主の神経発達に生理的な影響を及ぼすヒスチジン代謝系の制御因子であることが明らかになった。しかし、腸内細菌叢がこれらの化合物を通じて認知能力を調節する正確なメカニズムについては、まだ明らかにされていない。
研究の限界
我々の研究に限界がないわけではない。強力なDNA配列決定技術を用いた横断的研究は、微生物の組成シグネチャーと宿主の表現型を結びつける仮説を確認する出発点となる。GF動物は、高度に制御された環境で腸内細菌叢の機能を調べるための優れたツールであるが、例えばヒトの疾患における認知機能障害を調節するなどの応用医学への応用は、大胆ではあるがエキサイティングなことである。本研究は、小児コホートからの16サンプルに限定されているが、腸内細菌叢が記憶機能に影響を及ぼすことは、ここに示した結果から明らかである。ヒスチジン代謝産物に加えて、微生物相が認知能力に影響を与える可能性のある他の免疫学的・代謝学的メカニズムを排除することはできない。実際、我々の研究では、認知における潜在的な役割について調査が必要な微生物タンパク質がいくつか同定された。本研究で得られたヒスチジン代謝産物や特定の細菌株と認知機能との関連については、受胎時からマウスに投与し、行動評価と脳機能検査を組み合わせてさらに検証する必要があると考える。生涯を通じて腸内細菌叢と脳機能の共進化に関するエビデンスが蓄積していることを考えると、動物モデルやヒトを含む他の種における長期的影響についても研究すべきである。乳児ドナーはオスとメスであったが、FMTのレシピエントマウスはオスであった。従って、性別による影響は十分に評価されておらず、さらなる調査が必要である。加えて、ドナーとなった乳児はアンダルシア地方に住むスペイン系白人の集団に属しており、場所や民族によるバイアスの可能性を排除する必要がある。
STAR★方法
主要資源表
試薬またはリソースソース識別子
生物学的サンプル
ドナー由来の乳児便サンプル PREOBE プロジェクト Campoy et al.75
化学物質、ペプチド、組換えタンパク質
ジルコニア/シリカビーズ Biospec 11079110z
Trizma Sigma-Aldrich T1503
HCl VWR 20.252.420
塩化ナトリウム Sigma-Aldrich S3014
分子生物学用水 Sigma-Aldrich 95284
MilliQ - 水 Millipore N/A
フェノール緩衝液 AppliChem A1153
クロロホルム/イソマイルアルコール Sigma-Aldrich C0549
エタノール Sigma-Aldrich E7148
酢酸ナトリウム Sigma-Aldrich S2889
アガロース Sigma-Aldrich A5093
トリス-酢酸-EDTA バッファー Sigma-Aldrich T9650
1 kb+ DNA ラダー Thermo Scientific SM1331
100 bp DNA ラダー Thermo Scientific SM0241
ドデシル硫酸ナトリウム Scharlab SD00100500
アクリルアミド Sigma-Aldrich A3574
リン酸緩衝生理食塩水 Sigma-Aldrich P3813
バグバスター Millipore 70584
フェニルメチルスルホニルフルオリド Sigma-Aldrich P7626
トリプシン Sigma-Aldrich T6567
dNTPs KapaBiosystems KN1009
SYBR Green I ロシュ 11759078001
酢酸 Panreac 1.310.081.211
L-システイン Sigma-Aldrich 168149
L-ヒスチジン VWR 1B1164
ヒスタミン Acros Organics 411710010
ウロカン酸 Acros Organics 228960050
L-グルタミン酸 Sigma-Aldrich G1251
N-アクテイルヒスタミン Sigma-Aldrich 858897
イミダゾール酢酸 Sigma-Aldrich 219991
イミダゾールプロピオン酸 Sigma-Aldrich 72951
アセトニトリル VWR 20.060.290
グルタチオン シグマアルドリッチ G4251
ジシルグリシン Alfa Aesar A10523
ジチオスレイトール Sigma-Aldrich D9779
Teklad Global 18%タンパク質げっ歯類用飼料 Envigo 2018S
重要な市販アッセイ
MySeq Reagengt Kit V2(500サイクル)Illumina MS-102-2003
Nextera XT DNAライブラリー調製キットIllumina FC-131-1096
バイオアナライザー RNA 6000 Nano キット Agilent 5067-1511
Quant-iT™ PicoGreen™ dsDNAアッセイキット Invitrogen P11496
Expand High Fidelity PCR System Roche 11759078001
寄託データ
質量分析プロテオミクスデータ ProteomeXchange Consortium PXD009056
実験モデル 生物/株
マウス WT: C57BL/6J雄マウス グラナダ大学 N/A
オリゴヌクレオチド
16S V1フォワードSigma-Aldrich AGAGTTTGATCNTGGCTCAG
16S V2 reverse Sigma-Aldrich TGCTGCCTCCCGTAGGAGT
16S V3フォワードSigma-Aldrich CCTACGGGGNGGCWGCAG
16S V4リバース シグマ・アルドリッチ GACTACHVGGGTATCTAATCC
ソフトウェアとアルゴリズム
R 統計プログラミング環境 http://www.R-project.org バージョン 4.0.3
Proteome Discoverer ソフトウェア Thermo Fisher Scientific バージョン 3.1
Metaboanalyst https://www.metaboanalyst.ca/ バージョン 5.0
Ribosomal Database Project (RDP-II) http://rdp.cme.msu.edu/ version 2.14
16S baed-ID https://www.ezbiocloud.net/ 更新 2023/08/23
SPSS IBM バージョン 22.0
その他
フレキシブルフィルム Gnotobiotic Isolators クラス バイオロジカルクリーン N/A
アイソケージP マイクロアイソレーター テクニプラスト SO72P
ミニビーズビーター24 Biospec N/A
ナノHPLCシステム NanoMate Advion Triversa NanoMate
Orbitrap Fusion質量分析計 Thermo Fisher IQLAAEGAAPFADBMBHQ
新しいタブで表を開く
リソースの有無
リードの連絡先
詳細は、リードコンタクトのAlicia Ruiz-Rodriguez 博士 (aliruizrodriguez@ugr.es) までお問い合わせください。
材料の入手可能性
本試験では新たな試薬は得られていない。
データおよびコードの利用可能性
質量分析プロテオミクスデータは、PRIDE76パートナーリポジトリを通じてProteomeXchange Consortiumに寄託され、データセット識別子PXD009056で公開されている。
本論文ではオリジナルコードは報告していない。
本論文で報告されたデータを再解析するために必要な追加情報は、要請があれば主任連絡先から入手可能である。
実験モデルと被験者の詳細
本試験では、PREOBE試験コホート(ClinicalTrials.gov NCT01634464)に属する乳幼児パネルから、腸障害を認めず抗生物質を服用していない生後6ヵ月の健康な満期乳幼児を選択した75。このプロジェクトでは、2007年から2012年の間に、スペイン、グラナダのサン・セシリオ大学病院と母乳大学病院で妊婦を募集した。母親の研究除外基準は、他の研究への同時参加、あらゆる種類の薬物治療、診断された疾患(例えば、妊娠前糖尿病、高血圧または子癇前症、子宮内発育遅延、母体の感染症、甲状腺機能低下症/甲状腺機能亢進症、肝疾患、腎疾患)、菜食であった。新鮮な便は分娩6ヵ月後に採取され、処理されるまで直ちに-80℃で保存された。この研究では、人体測定、健康アンケート、子どもの医学的評価などが行われた。このプロジェクトは、ヘルシンキ宣言(1989年に香港で、2000年にエディンバラで見直された)およびEECのGood Clinical Practice勧告(文書111/3976/88 1990)で認められた倫理基準と、ヒトを対象とした臨床研究を規制する現行のスペインの法律(勅令561/1993)に従った。この研究は開始前に参加者に説明され、両親はインフォームド・コンセントに署名した。無菌のC57BL/6J雄マウスを飼育し、スペイン、グラナダ大学のGnotobiotic Facilityのフレキシブルフィルムアイソレーターで維持した。マウスは通常の12時間明暗サイクル(朝7時点灯)で飼育され、オートクレーブ滅菌水とEnvigo社(ウィスコンシン州マディソン)製の滅菌可能なTeklad Global 18% Protein Rodent Dietを自由摂取させた。衛生状態は、gnotobioticマウスの16S配列決定を含む、培養依存および非依存的方法により日常的にモニターした。すべてのマウス実験はJunta de Andalucía Animal Committeeの規定に従って行われた。
方法の詳細
乳児の神経発達の評価
生後6ヶ月の乳児の神経発達の評価にはBSID-IIIを使用した(n=225)。BSID-IIIによる乳幼児評価は、認知能力、言語、運動発達の3つの主要な領域にわたって行われる。言語尺度では、受容言語と表出言語という2つの発達領域が調査される。運動尺度では、微細運動と粗大運動の両方の発達を調べることができる。言語領域と運動領域の複合スコアが得られる。BSID-IIIの各神経発達尺度の得点により、乳児は中央値(50パーセンタイル)以上と未満の2群に二分された。
便サンプルからのDNA抽出
生後6ヵ月の乳児(n = 69)の便検体からゲノムDNAを抽出した(31)。簡単に説明すると、便検体は1mlのTN150緩衝液(10mM Tris-HCl pH8.0、150mM NaCl)に懸濁した。ジルコニウムガラスビーズ(0.3 g)と150 mlの緩衝フェノールを加え、4℃で5000 rpmに設定したミニビーズビーター(Biospec Products社、米国)で15秒間、細菌を破砕した。遠心分離後、上清からゲノムDNAをフェノール-クロロホルム抽出法で精製し(明晰な溶液が得られるまで)、次いでDNAをエタノール沈殿させた。得られたペレットを70%(v/v)エタノールで洗浄し、4℃で一晩インキュベートして100mlのDNAフリーのヌクレアーゼフリーの水に溶解した。DNAサンプルはさらに処理するまで-20℃で保存した。品質はアガロースゲル電気泳動でチェックし、Quant-iT PicoGreen dsDNA assay kit (Invitrogen, Darmstadt, Germany)で定量した。
16S rRNA遺伝子の塩基配列決定とデータ処理
等モル比のアンプリコンをプールしてライブラリーを調製し、Illumina MiSeqプラットフォーム(Genetic Service, University of Granada)を用いて塩基配列を決定した。リードをデマルチプレックスしてソートし、ペアエンドをマッチングして240 ntリードを得た。データセットにフィルターをかけ、MOTHURプログラムunique.seqsとpre.clusterを用いて99%の類似度で分類単位を定義した。種の分類は、RDP-IIのナイーブベイズアルゴリズムCLASSIFIERを用いて割り当てた80。信頼値スコアが0.8未満の種は未割り当てとみなし、上位の分類学的ランクを用いてアノテーションを行った。EzBioCloud81 Identify と RDP-II Sequence match ツールを使用して、興味深い種に最も近い培養株を同定した。
タンパク質の抽出、分離、同定、データ処理
タンパク質抽出は、糞便中の細菌(n = 14)から、以前に記載された方法で行った。電気泳動後、トリプシン消化によりタンパク質バンドからペプチドライセートを生成し、ナノHPLCシステムAdvion NanoMateとOrbitrap Fusion質量分析計(Thermo Fisher Scientific)で分析した。Proteome Discoverer (v1.4, Thermo Fisher Scientific)は、16S rRNA遺伝子配列決定により選択された細菌分類群のタンパク質コードエントリーを含むデータベースに対してSequestHTアルゴリズムを使用し、FDR < 1% の閾値でランク1のペプチドのみを取得した。質量分析プロテオミクスデータは、PRIDE76パートナーリポジトリを介してProteomeXchange Consortiumにデータセット識別子PXD009056で寄託された。タンパク質量は、異なるサンプル間のタンパク質量の相対比較を可能にする正規化されたスペクトル量に基づいて計算された。実験グループを比較するために、補正されたメタプロテオーム行列をsPLS-DAに使用した。sPLS-DAは、実験グループ間のばらつきを明らかにし、可能な限り低い誤差率で識別を可能にする特徴を決定する教師ありモデルである。
ヒスチジンのメタボロームとヒスチジンアンモニアリアーゼ活性の測定
ヒスチジンのメタボロームとヒスチジンアンモニアリアーゼ活性の測定は、Acuñaらの記述に従って行った37。簡単に述べると、糞便、微生物、血漿、尿および臍帯周囲皮質87のマトリックスを抽出し、UHPLC-MSグレードのアセトニトリルと混合し、ろ過し、Waters Acquity UPLC™ System I-Class (Waters, UK)を用いてクロマトグラフィーで分離し、Watersトリプル四重極質量分析計 (XevoTQ-XS, Waters, UK)で分析した。ヒスチジンアンモニアリアーゼ活性は、グルタチオンとグリシルグリシンの存在下、1 mg mL-1ヒスチジンを基質として、タンパク質抽出液中37℃で測定した。尿サンプルについては、尿分析物の定量を調整するためにクレアチニン値を同時に分析した。
微生物叢の移植
便サンプル(n = 16)の糞便微生物叢移植は、無酸素グローブチャンバー内の嫌気条件下で行った。約0.3 gの便を、還元剤として2 mM DTTを含む5 mLの嫌気性PBSに懸濁した。各便サンプルを1分間ボルテックスし、500xgで1.5分間遠心した。懸濁液を嫌気チャンバーから取り出し、直ちに使用した。各糞便懸濁液200μLを、無作為に選んだ9週齢のC57BL/6J雄マウス2匹に経口投与で接種した。同じ幼児ドナー移植のマウスをISOCAGE Nアイソレーターケージに一緒に収容し、オートクレーブした水と滅菌可能なTeklad Global 18%タンパク質齧歯類用飼料を自由摂取させた。新鮮なマウスの便と尿を採取し、直ちに-80℃に保存した。行動試験後、マウスの尾から血液を採取した。マウスは頸椎脱臼し、脳を摘出し、脊髄周囲皮質をスライスして-80℃で保存した。
新規物体認識
記憶機能は、移植2週間後に新奇物体認識テスト(NORT)で評価した。NORTは、マウスの学習と記憶に関する神経心理学的変化を調べるのに、比較的ストレスが少なく効率的な方法として広く用いられている88。NORTは、データを混乱させる可能性のある正の強化も負の強化も必要とせず、マウスの新奇なものに対する自然な好奇心、すなわち新奇な対象を探索する傾向に基づいている。NORTは2日間にわたって実施され、馴化日と訓練・試験日が含まれた。馴化はオープンフィールドテストで行われ、50×50×30cm(縦×横×高さ)のオープンフィールドボックスの中央にマウスを1匹ずつ置いた。マウスの行動はビデオ・コンピュータ追跡システムで5分間記録された。フィールドの中心部と周辺部に滞在した時間、中心部に入った回数、移動距離、速度が算出された。馴化から24時間後、マウスは同じアリーナの中央部に置かれ、アリーナの中央対称位置にある2つの同じ物体から等距離に置かれ、物体を探索するために5分間与えられた。2時間後、マウスは慣れ親しんだ物体と新しい物体に5分間暴露された。各試験後、物体は70%エタノールで洗浄した。すべてのマウスは5秒以上探索した。弁別指標はd1/e2として計算され、d1は新しい物体の探索時間から見慣れた物体の探索時間を引いたもの、E2は全探索時間である。t検定は、架空群(ランダム探索、弁別能力なし、実験群で観察された標準偏差あり)と各マウス群の弁別値を独立に比較するため、およびマウス群間の弁別指標を比較するために用いられた。
定量化と統計解析
統計解析および可視化は、SPSSバージョンv22.0およびRと複数の解析パッケージ(ggplot2、ggpubr、ggstatsplot、stats、corrplot)を用いて行った。すべての決定について、有意性のカットオフはp≤0.05、または多重検定補正を適用した場合はFDR(False Discovery Rate)≤0.05とした。すべてのデータは、統計検定の前に正規分布と分散の等質性を検定した。多重検定補正にはBenjamini-Hochberg手順を用いた。適切なマウスのサンプルサイズを事前に決定するための検出力計算は行わなかった。腸内細菌叢組成に対する研究変数の効果量および有意性は、veganパッケージのenvfit関数を用いて決定した。組成データのβ多様性の順序プロットは、ユニフラック(Gunifrac)距離およびアイチソン距離(Aitchison distances)90(この距離は、スケール不変性、摂動不変性、並べ替え不変性およびサブ組成優位性を有する)、およびブレイ・カーティス非類似度測定基準(Phyloseq)を用いて算出し、続いてPERMANOVA検定(Vegan)を用いて実験群間の差異を検出した。 α多様性は、種レベルで解析した。ヒトの研究では、DESeq2パッケージを用いて共変量で調整し、多重比較補正を行った両側Waldの検定を用いて、いくつかの異なる分類レベルでの有意差のある系統型存在量を非正規化生カウント表から構築した。微生物群の共変量の影響を除外した後の分類群とCCの間の関連は、glm R関数を用いて一般化線形モデルをフィットさせることで評価した。マウスの研究では、Random Forest回帰モデル(randomForest、caret、tuneRanger、Boruta、pROCパッケージ)を用いて、予測因子のセットからその決定因子を取り除くことによって生じる誤差の増加を推定することによって、各決定因子に重要度スコアを割り当てる主要な識別特徴を同定した。トレーニング(サンプルの80%)とテスト(サンプルの20%)のデータセットをランダムに作成し、識別特徴の予測モデルを構築した。このモデルは、元の属性の重要度を、それらの並べ替えコピーを用いて推定されるランダムに達成可能な重要度と比較し、そのテストを安定させるために無関係な特徴を徐々に除去することによって、関連する特徴のトップダウン探索を実行するBorutaパッケージを用いてさらに改良された。重要であると確認されたすべての特徴は、関連するものとして扱われ、モデルの予測力を調べるために使用された。メタボロームデータと分類群データの相関は、スピアマンの順位相関を用い、多重比較補正のためにcorr.test (Psych)を用いて求めた。
謝辞
Ana NietoとMaría José García Chicanoの協力、Inmaculada TerceroとAna Lópezの技術者のサポートに感謝する。著者の研究室での研究は、スペイン保健科学省(PREOBEプロジェクトP06-CTS-02341、BFU2012-40254-C03-01、PID2020-114269GB-I00)、Junta de Andalucía、UFZ-Germanyから提供されたリソースに基づいて行われた。T.C.はUGRの生物医学博士課程に在籍し、スペイン経済・競争力省によるFPIプログラム(BES-2013-065133)のフェローシップを受け、現在はカルロス3世研究所の「サラ・ボレル」プログラム(CD21/00187)の支援を受けている。I.A.、S.M.-M.、A.R.-R.はスペイン経済競争力省より博士号奨学金(FPU)を授与された。A.R.-R.は現在、UGRのマリア・サンブラノ次世代EU人材再導入プログラム(María Zambrano Next Generation EU-Talent Reincorporation)を受けている。
著者の貢献
構想、C.C.およびA.S.、方法論、A.R.-R.、T.C.、I.A.およびA.S.、形式分析、T.C.、A.R.-R.、I.A.およびA.S.、調査、T.C.、A.R.、I.A.、S.M.-M.、F.G.、M.G.、N.J.、S.-B.H.およびA.S.、リソース、A.S.、 原案執筆:A.R.-R.およびA.S.、査読および編集:全著者、可視化:T.C.、A.R.-R.およびA.S.、資金獲得:A.S.、M.v.B.およびC.C.、監督:A.S.
利益申告
著者らは、競合する利益はないと宣言している。
補足情報
pdfをダウンロード (4.12 MB)
pdfファイルのヘルプ
ドキュメントS1. 図S1-S6
ダウンロード .xlsx (.02 MB)
xlsxファイルのヘルプ
表S1. PREOBE研究に登録された乳児のBayley Scales of Infant Development, Third Edition(BSID-III)テストのスコアとSTAR Methodsとの関連
ダウンロード.xlsx (.26 MB)
xlsxファイルのヘルプ
表S2. 図1および図2に関連する、乳幼児における操作的分類単位とCOG機能
ダウンロード .docx (.02 MB)
docxファイルのヘルプ
表S3. 図1に関連する、生後6ヵ月の乳児における腸内微生物のα多様性指標、β多様性距離、BSID-III尺度間の関連性に関する有意なp値
ダウンロード.docx (.02 MB)
docxファイルのヘルプ
表S4. 図1および図5に関連した、リボソームデータベースプロジェクト(RDP-II)およびEzBioCloudデータベースにおける、乳児コホートおよびマウス群の培養可能な株への異なる存在量の種の割り当て
ダウンロード.docx (.02 MB)
docxファイルのヘルプ
表S5. Inf-aboveCC腸内細菌叢およびInf-belowCC腸内細菌叢を投与したマウスのオープンフィールド馴化課題の結果(図4関連
ダウンロード.xlsx (.03 MB)
xlsxファイルのヘルプ
表S6. 図5に関連した、乳児ドナーの腸内細菌叢を受け取った無菌マウスで特徴付けられた、異なる存在量の種のrDNAの操作上の分類単位と割り当て結果
参考文献
シュミット T.S.B.
Raes J.
ボーク P.
ヒト腸内細菌叢:関連から調節へ。
Cell. 2018; 172: 1198-1215
論文で見る
スコープス (457)
PubMed
要旨
全文
全文PDF
グーグル奨学生
スタイルズJ.
ジャーニガン T.L.
脳の発達の基本
Neuropsychol. 2010; 20: 327-348
論文で見る
スコープス (1114)
PubMed
クロスフィルム
グーグル奨学生
セルドT.
ルイスA.
スアレス A.
カンポイC.
プロバイオティクス、プレバイオティクス、脳の発達。
栄養素。2017; 9: 1247-1265
論文で見る
スコープス (59)
PubMed
クロス
グーグル奨学生
クライアンJ.F.
マズマニアン S.K.
微生物叢-脳軸:文脈と因果関係。
サイエンス。2022; 376: 938-939
論文で見る
スコープス (31)
PubMed
クロス
グーグル奨学生
デボネL.
クラークG.
トラプリンA.
オサリバンO.
クリスピーF.
モロニー R.D.
コッター P.D.
ディナン T.G.
Cryan J.F.
マウスにおける青年期早期からの腸内細菌叢の減少:脳と行動への影響。
脳行動。Immun. 2015; 48: 165-173
論文で見る
スコープス (499)
PubMed
クロス
グーグル奨学生
フレーリッヒ E.E.
ファルジ A.
マイヤーホーファーR.
ライヒマン F.
ヤーチャン A.
ワグナー B.
ジンザー E.
ボルダーグ N.
マグネス C.
フレーリッヒ E.
他。
抗生物質誘発性腸内細菌異常症による認知障害:腸内細菌叢-脳コミュニケーションの解析。
脳行動。Immun. 2016; 56: 140-155
論文で見る
PubMed
クロスフィルム
グーグル奨学生
ルチンスキー P.
マクヴェイ・ノイフェルドK.A.
オリアックC.S.
クラーク G.
ディナン T.G.
Cryan J.F.
バブルの中で育つ:無菌動物を用いて腸内細菌叢が脳と行動に及ぼす影響を評価する。
Int. J. Neuropsychopharmacol. 2016; 19pyw020
論文で見る
スコープス (369)
PubMed
クロス
グーグル奨学生
ノイフェルド K.A.
カン N.
ビエネンストックJ.
フォスターJ.A.
不安様行動に対する腸内細菌叢の影響。
コミュニケーション。Integr. 2011; 4: 492-494
論文で見る
PubMed
クロスフィルム
グーグル奨学生
ノイフェルド K.M.
カン N.
ビエネンストックJ.
フォスター J.A.
無菌マウスにおける不安様行動の減少と中枢神経化学的変化。
Neurogastroenterol. Motil. 2011; 23: 255-264
論文で見る
(1046件)
PubMed
クロス
グーグル奨学生
ディアス・ヘイツR.
ワン・S.
Anuar F.
Qian Y.
Björkholm B.
サミュエルソン A.
ヒバード M.L.
フォルスベリ H.
ペッターソンS.
正常な腸内細菌叢は脳の発達と行動を調節する。
Proc. Natl. Sci. USA. 2011; 108: 3047-3052
論文で見る
スコープス(2305)
PubMed
クロス
グーグル奨学生
ボレ Y.E.
オキーフG.W.
クラークG.
スタントン C.
ディナン T.G.
クライアンJ.F.
微生物叢と神経発達の窓:脳疾患への影響。
トレンドMol。Med. 2014; 20: 509-518
論文で見る
スコープス (749)
PubMed
概要
全文
全文PDF
グーグル奨学生
リウ・Z.
Wei S.
チェン X.
Liu L.
Wei Z.
Liao Z.
Wu J.
Li Z.
Zhou H.
Wang D.
小児および青年期における抗生物質の長期または反復使用が中高年成人の認知障害に及ぼす影響: コホート研究。
Front. Aging Neurosci. 2022; 14833365
論文で見る
グーグル・スカラー
クリスチャン L.M.
ギャレーJ.D.
ヘイド E.M.
ショッペ=サリバンS.
カンプ・ドゥシュ C.
ベイリーM.T.
腸内細菌叢の組成は幼児期の気質と関連している。
脳行動。Immun. 2015; 45: 118-127
論文で見る
スコープス (129)
PubMed
クロス
グーグル奨学生
フォックス M.
リー S.M.
ワイリーK.S.
ラギシェッティV.
サンドマン C.A.
ジェイコブズ J.P.
グリンL.M.
乳児腸内細菌叢の発達は生後1年間の気質を予測する。
Dev. Psychopathol. 2021; 10: 1-12
論文で見る
Google Scholar
クエルダシ F.R.
エンダース C.
カルナニ N.
ブロークマンB.
ヤップ・センC.
グラックマン P.D.
メアリー・ダニエル・L.
ヤップ F.
エリクソン J.G.
Cai S.
他。
多世代にわたる逆境が、幼児期におけるヒト腸内細菌叢の組成と社会情動機能に及ぼす影響。
Proc. Natl. Sci. USA. 2023; 120e2213768120
論文で見る
日本学術振興会特別研究員 (0)
パブコメ
クロス
グーグル奨学生
ニエト-ルイスA.
セルドT.
ジョルダーノB.
トーレス-エスピノーラF.J.
エスクデロ-マリーン M.
ガルシア-リコバラザM.
ベルムデス M.G.
ガルシア・サントス J.A.
スアレス A.
カンポイC.
母親の体重、腸内細菌叢、幼児期の行動との関連:PREOBEフォローアップ研究。
児童思春期。精神医学。健康。2023; 1741
論文で見る
スコープス(2)
PubMed
クロス
グーグル奨学生
Ou Y.
ベルザー C.
スミットH.
デ・ウィースC.
生後14年間の腸内細菌叢の発達と思春期の内面化・外面化困難および社会不安との関係。
ユーロ。児童思春期。精神医学。2023;
論文で見る
スコープス (3)
PubMed
クロス
グーグル奨学生
カールソン A.L.
シャ・ケイ
アズカラートペリルM.A.
ロジン S.P.
ファイン J.P.
ムー W.
ゾップ J.B.
キンメル M.C.
スタイナー M.A.
トンプソン A.L.
他。
乳児の腸内細菌叢組成は、パイロット研究における非社会的恐怖行動と関連している。
Nat. Commun. 2021; 123294
論文で見る
スコープス (23)
クロスリファレンス
グーグル奨学生
シャロン G.
クルーズ N.J.
カン D.W.
ガンダル M.J.
ワン・ビー
キム Y.M.
ジンク E.M.
ケイシー C.P.
テイラー B.C.
レーン C.J.
他
自閉症スペクトラム障害のヒト腸内細菌叢はマウスの行動症状を促進する。
Cell. 2019; 177: 1600-1618.e17
論文で見る
スコープス (561)
PubMed
要旨
全文
全文PDF
グーグル奨学生
シャオ E.Y.
マクブライド S.W.
シェン S.
シャロンG.
ハイド E.R.
マキュー T.
コデリ J.A.
チャウ J.
ライズマン S.E.
ペトロシーノ・J.F.
他
微生物叢は神経発達障害に伴う行動的・生理的異常を調節する。
Cell. 2013; 155: 1451-1463
論文で見る
筑波大学
パブコメ
概要
全文
全文PDF
グーグル奨学生
パラシオス N.
リー J.S.
スコット T.
ケリー R.S.
ブパティラジュ S.N.
ビゴニア S.J.
タッカーK.L.
プエルトリコ人コホートにおける循環血漿中代謝産物と認知機能。
J. Alzheimers Dis. 2020; 76: 1267-1280
論文で見る
スコープス (7)
PubMed
クロス
グーグル奨学生
チェン L.H.
リウ Y.W.
Wu C.C.
Wang S.
ツァイ Y.C.
精神保健、神経変性および神経発達障害におけるサイコバイオティクス。
J. Food Drug Anal. 2019; 27: 632-648
論文で見る
スコープス(115)
PubMed
クロス
グーグル奨学生
ルイス A.
セルドT.
Jáuregui R.
Pieper D.H.
マルコス A.
クレメンテ A.
ガルシア F.
マルゴレス A.
フェレール M.
カンポイ C.
スアレスA.
1年間のカロリー制限は、肥満の青少年における代謝性能ではなく、腸内細菌組成に影響を与える。
環境。Microbiol. 2017; 19: 1536-1551
論文で見る
スコープス (48)
PubMed
クロス
グーグル奨学生
セルドT.
ルイズ A.
アクーニャ I.
Jáuregui R.
イェムリッヒ N.
ハーンゲ S.B.
フォン・ベルゲン M.
スアレス A.
カンポイC.
ヒトの初期生活における腸内微生物の機能的成熟と継承。
環境。Microbiol. 2018; 20: 2160-2177
論文で見る
スコープス (27)
PubMed
クロス
グーグル奨学生
Kim H.
Jeon S.
キム J.
Seol D.
Jo J.
Cho S.
Kim H.
マウスにおけるStreptococcus thermophilus EG007の記憶増強作用の検討とナノポアシーケンスを用いた分子およびメタゲノム特性の解明。
Sci. Rep. 2022; 1213274
論文で見る
Google Scholar
Park S.
ウーX.
副交感神経系の抑制を介した記憶障害とアルツハイマー病における腸内細菌叢の調節。
Int. J. Mol. Sci. 2022; 2313574
論文で見る
グーグル学者
サングイネッティ E.
グッツァルディ M.A.
トリポディ M.
パネッタ D.
セルマ-ロヨM.
ゼガ A.
テレスキ M.
コラド M.C.
イオッツォ P.
マウスモデルにおける過体重の母親の子孫における記憶と探索行動の低下に関連する微生物叢シグネチャー。
Sci. Rep. 2019; 912609
論文で見る
スコープス(30)
クロスリファレンス
グーグル奨学生
カイン W.W.T.
ヴォン・M.L.
ン T.K.S.
Feng L.
レーン G.A.
クマール A.P.
クア E.H.
マヘンドラン R.
マヘンドラン R.
リーY.K.
メンタルアウェアネスは軽度認知障害を改善し、腸内細菌叢を調節した。
Aging. 2020; 12: 24371-24393
論文で見る
スコープス (23)
パブコメ
クロス
グーグル奨学生
Lu S.
Shao L.
Zhang Y.
Yang Y.
Wang Z.
Zhang B.
Yu J.
Xu Q.
王 S.
Chen X.
et al.
高血圧患者における認知機能障害に対する腸内細菌叢の予測値。
Dis. Markers. 2021; 2021: 1-9
論文で見る
スコープス (3)
クロス
グーグル奨学生
ラジャブザデ D.
ボッシュJ.A.
ウイターリンデンA.G.
ズウィンダーマン A.H.
イクラム M.A.
ファン・ムースJ.B.J.
ルイク A.I.
ニーウドープ M.
ロク A.
ファン・ドゥインC.M.
他
うつ症状の腸内細菌ワイド関連研究。
Nat. Commun. 2022; 137128
論文で見る
スコープス (41)
クロスリファレンス
グーグル奨学生
フェレール M.
ルイズ A.
ランザ F.
ハーンゲ S.B.
オーバーバッハ A.
ティル H.
バルジェラ R.
カンポイ C.
セグラ M.T.
リヒター M.
他。
痩せている青年と肥満の青年の遠位腸からの微生物叢は、コミュニティ構造の明確な違いのほかに、部分的な機能的冗長性を示す。
環境。Microbiol. 2013; 15: 211-226
論文で見る
スコパス (187)
パブコメ
クロス
グーグル奨学生
Xiong W.
エイブラハム P.E.
Li Z.
Pan C.
ヘティヒ R.L.
ヒト腸内細菌叢の代謝機能と活性の範囲を特徴付けるための微生物メタプロテオミクス。
Proteomics. 2015; 15: 3424-3438
論文で見る
スコープス (82)
PubMed
クロス
グーグル奨学生
タラッカー-マーサーA.E.
ゲラーM.E.
ヒスチジンサプリメントの利点と副作用。
J. 栄養2020; 150: 2588S-2592S
論文で見る
スコープス(23)
PubMed
要旨
全文
全文PDF
グーグル奨学生
笹原一郎
藤村直樹
野沢洋子
古畑靖之
佐藤博之
疲労・睡眠障害スコアの高い被験者におけるヒスチジンの精神疲労および認知能力への影響。
Physiol. Behav. 2015; 147: 238-244
論文で見る
スコープス (48)
PubMed
クロス
グーグル奨学生
中村哲也
長沼文彦
工藤美U.
Roh S.
柳井一男
吉川敏明
ヒスチジンの経口摂取はマウスにおいてヒスタミン作動性神経系の活性化を介してワーキングメモリを改善する。
Biochem. Biophys. Res. Commun. 2022; 609: 141-148
論文で見る
スコープス (1)
PubMed
クロス
グーグル奨学生
アルバレスE.O.
認知におけるヒスタミンの役割。
行動。脳研究 2009; 199: 183-189
論文で見る
スコパス (51)
PubMed
クロス
グーグル奨学生
アクーニャ I.
ルイス A.
セルドT.
カンタレロ S.
ロペス-モレノA.
アギレラ M.
カンポイ C.
スアレス A.
ヒトおよびマウスの微生物叢および生体試料中のヒスチジン代謝中間体の迅速同時定量。
BioFactors. 2022; 48: 315-328
論文で見る
スコパス (7)
PubMed
クロス
グーグル奨学生
ソビオ U.
グールディングN.
マクブライドN.
クック E.
ガッチョーリ F.
チャノック・ジョーンズ D.S.
ローラー D.A.
スミスG.C.S.
母体の血清代謝産物比は、用語で胎児の成長制限を予測する。
ナット。医学。2020; 26: 348-353
論文で見る
スコープス (74)
パブコメ
クロス
グーグル奨学生
D'Eufemia P.
フィノッキアーロR.
チェッリ M.
ヴィオッツィ L.
モンテレオーネ D.
ジャルディーニO.
特発性小児自閉症における低血清トリプトファン対大型中性アミノ酸比。
Biomed。Pharmacother. 1995; 49: 288-292
論文で見る
スコパス (43)
PubMed
クロス
グーグル奨学生
バーンズW.
チャウドハリ B.P.
ハフナーD.N.
一般的な新生児神経疾患の神経遺伝学的および代謝学的模倣。
Semin. Pediatr. Neurol. 2022; 42100972
論文で見る
スコパス(2)
PubMed
クロス
グーグル奨学生
クレイトン T.A.
ベイカー D.
リンドンJ.C.
エベレットJ.R.
ニコルソン J.K.
ヒトの薬物代謝に影響を及ぼす重要な宿主-微生物間代謝相互作用のファーマコメタボノミックによる同定。
Proc. Natl. Sci. USA. 2009; 106: 14728-14733
論文で見る
スコープス (606)
PubMed
クロス
グーグル奨学生
フィッシュバッハM.A.
マイクロバイオーム:因果関係とメカニズムに焦点を当てる。
細胞。2018; 174: 785-790
論文で見る
スコープス (151)
PubMed
要旨
全文
全文PDF
グーグル奨学生
カールソン A.L.
Xia K.
アズカラートペリルM.A.
ゴールドマンB.D.
アーン M.
スタイナーM.A.
トンプソン A.L.
ゲン X.
ギルモア J.H.
ニックマイヤーR.C.
認知発達に関連する乳児腸内細菌叢。
Biol. Psychiatry. 2018; 83: 148-159
論文で見る
スコープス (303)
PubMed
要旨
全文
全文PDF
グーグル奨学生
タマナ S.K.
トゥン H.M.
コンヤ T.
チャリ R.S.
フィールド C.J.
グットマン D.S.
ベッカー A.B.
モラエス T.J.
ターベイ S.E.
スバラオ P.
他
乳児期後期のバクテロイデス優勢腸内細菌叢は神経発達の亢進と関連する。
腸内細菌。2021; 13: 1-17
論文で見る
スコープス (53)
PubMed
クロス
グーグル奨学生
ローテンバーグ S.E.
チェン Q.
シェン J.
Nong Y.
Nong H.
トリン E.P.
Biasini F.J.
Liu J.
Zeng X.
Zou Y.
他。
中国農村部の3歳児を対象とした横断的解析における神経発達と腸内細菌叢の相関。
Sci. Rep. 2021; 117384
論文で見る
スコープス (16)
PubMed
クロス
グーグル奨学生
ソルディロ J.E.
コリック S.
ラランホN.
キャリーV.
ワインストックG.M.
ゴールド D.R.
オコナー G.
サンデル M.
バカリエL.B.
ベイジェルマンA.
他
乳児の腸内マイクロバイオームと幼児期の神経発達アウトカムとの関連:VDAART無作為化臨床試験の補助研究。
JAMA Netw. Open. 2019; 2e190905
論文で見る
スコープス(61)
PubMed
クロス
グーグル奨学生
クッソット S.
アウイゼレートB.
カプロンL.
腸から脳へ:因果関係を探るツールとしてのマウスにおける微生物叢の減少。
脳行動。Immun. 2021; 94: 4-5
論文で見る
スコパス(2)
PubMed
クロス
グーグル奨学生
ワーナー B.B.
神経発達と神経精神疾患に対する腸内細菌叢の寄与。
Pediatr. Res. 2019; 85: 216-224
論文で見る
スコープス(78)
PubMed
クロス
グーグル奨学生
Lu J.
Synowiec S.
Lu L.
Yu Y.
Bretherick T.
タカダ S.
ヤルニク V.
キャプラン J.
キャプラン M.
クラウド E.C.
et al.
微生物叢はC57BL/6Jマウスの脳と行動の発達に影響する。
PLoS One. 2018; 13e0201829
論文で見る
スコープス(95)
クロスリファレンス
グーグル奨学生
ウォルター J.
アーメットA.M.
フィンレイ B.B.
シャナハン F.
腸内マイクロバイオームの因果関係の確立または誇張:ヒト微生物関連げっ歯類からの教訓。
Cell. 2020; 180: 221-232
論文で見る
スコープス(258)
PubMed
要旨
全文
全文PDF
グーグル奨学生
モヤ A.
フェレール M.
攪乱を受ける腸内細菌叢の機能的冗長性が誘発する安定性。
Trends Microbiol. 2016; 24: 402-413
論文で見る
スコープス (337)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ヴァッカ M.
チェラーノG.
カラブレーゼ F.M.
ポルティンカサ P.
ゴベッティ M.
デアンジェリスM.
ヒト腸Lachnospiraceaeの論争の役割。
微生物。2020; 8573
記事で見る
スコープ (589)
PubMed
クロス
グーグル奨学生
鄭 P.
Zeng B.
Zhou C.
Liu M.
Fang Z.
Xu X.
Zeng L.
Chen J.
Fan S.
Du X.
et al.
腸内細菌叢のリモデリングは、宿主の代謝を介する経路を介してうつ様行動を誘導する。
Mol. Psychiatry. 2016; 21: 786-796
論文で見る
PubMed
クロスフィルム
グーグル奨学生
コレット E.
ラトゥサキス D.
ポンティフェックス M.G.
クロスト E.H.
ヴォー L.
ペレス・サンタマリーナ E.
ゴールドソン A.
ブリオン A.
ハジホッセイニ M.K.
Vauzour D.
他
腸と脳のコミュニケーションにおけるムチン-糖鎖採食性ルミノコッカス・グナバスの役割。
腸内微生物。2022; 142073784
論文で見る
スコパス (10)
PubMed
クロス
グーグル奨学生
バケロ F.
マルティネスJ.L.
F Lanza V.
ロドリゲス-ベルトランJ.
ガラン J.C.
サン・ミラン A.
カントン R.
Coque T.M.
抗生物質耐性における進化の経路と軌跡。
Clin. Microbiol. Rev. 2021; 34e0005019
論文で見る
スコープス (45)
PubMed
クロス
グーグル奨学生
Li Y.
Sun H.
Huang Y.
Yin A.
Zhang L.
Han J.
Lyu Y.
Xu X.
Zhai Y.
Sun H.
et al.
ADHDの腸内メタゲノム特性から、バクテロイデス属オバツスに関連する宿主認知障害が少ないことが明らかになった。
Gut Microbes. 2022; 142125747
論文で見る
スコパス (7)
クロスフィルム
グーグル奨学生
Luk B.
ヴィーララガヴァンS.
エンゲヴィク M.
バルデラス M.
メジャー A.
ルンゲ J.
ルナ R.A.
ヴェルサロヴィッチJ.
ヒト "乳児型 "ビフィドバクテリウム種による生後コロニー形成が、グノトビオティック成体マウスの行動を変化させる。
PLoS One. 2018; 13e0196510
論文で見る
スコープス (54)
クロス
グーグル奨学生
アブデルハミド M.
Zhou C.
大野和彦
久原 毅
タスリマ F.
アブドラ M.
ユング C.G.
道川正明
プロバイオティクスBifidobacterium breveは、APPノックインマウスにおいてアミロイドβ産生とミクログリア活性化の両方を減少させることにより記憶障害を予防する。
J. Alzheimers Dis. 2022; 85: 1555-1571
論文で見る
スコープス (15)
PubMed
クロス
グーグル奨学生
モライス L.H.
シュライバーH.L.t.
マズマニアンSK。
行動と脳疾患における腸内細菌叢-脳軸。
Nat。Rev. Microbiol. 2021; 19: 241-255
論文で見る
スコパス (657)
PubMed
クロス
グーグル奨学生
セルドT.
ニエト-ルイスA.
ガルシア-サントスJ.A.
ロドリゲス-ペーンラインA.
ガルシア-リコバラザM.
スアレス A.
ベルムデス M.G.
カンポイC.
微生物叢、腸脳軸の発達に母親と乳児の栄養の影響に関する現在の知識。
アヌ。Rev.栄養2023; 43:251-278
記事で見る
スコープ(1)
PubMed
クロス
グーグル奨学生
サヴィニャック H.M.
Kiely B.
ディナン T.G.
Cryan J.F.
ビフィズス菌はBALB/cマウスのストレス関連行動および生理機能に株特異的な影響を及ぼす。
Neurogastroenterol. Motil. 2014; 26: 1615-1627
論文で見る
スコープス (312)
PubMed
クロス
グーグル奨学生
アクーニャI.
セルドT.
ルイス A.
トーレス-エスピノラF.J.
ロペス-モレノA.
アギレラ M.
スアレス A.
カンポイC.
細かい運動能力に関連する乳児腸内細菌叢。
栄養素。2021; 131673
記事で見る
スコープス(12)
クロス
グーグル奨学生
モライス L.H.
ゴルベバA.V.
モロニーG.M.
モヤ-ペレスA.
ヴェントゥーラ・シルヴァ A.P.
アルボレヤ S.
バスティアンスセンT.F.S.
オサリバン O.
レア K.
ボレ Y.
他。
マウスの帝王切開による出産によって誘発される永続的な行動効果。
Curr. Biol. 2020; 30: 3761-3774.e6
論文で見る
スコープス (51)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ゲーリッグJ.L.
ヴェンカテッシュ S.
チャン・H.W.
ヒバード M.C.
カン V.L.
チェン J.
チェン・R.Y.
スブラマニアン S.
カワーディン・C.A.
マイヤー M.F.
他。
gnotobiotic動物および低栄養児における微生物指向性食品の効果。
Science. 2019; 365eaau4732
論文で見る
スコープス(240)
Crossref
グーグル奨学生
セルドー T.
ルイス A.
アクーニャ I.
ニエト-ルイスA.
ディエゲス E.
セパルベダ-ヴァルブエナN.
エスクデロ-マリーン M.
ガルシア・サントス J.A.
ガルシア-リコバラザM.
ヘルマンF.
他
シンバイオティクス、長鎖多価不飽和脂肪酸、および乳脂肪球膜を補充した粉ミルクは、微生物叢の成熟と神経発達を調節する。
Clin. 2022; 41: 1697-1711
論文で見る
スコパス (4)
PubMed
要旨
全文
全文PDF
グーグル奨学生
チェン Y.
Xu J.
チェン Y.
腸内細菌叢による神経伝達物質の制御と神経疾患における認知への影響。
栄養素。2021; 132099
論文で見る
グーグル学者
プロベンシG.
コスタ A.
イズキエルド I.
ブランディーナ P.
パッサーニM.B.
脳ヒスタミンは、認識記憶を調節する:主要な認知障害における可能な含意。
Br。J. Pharmacol. 2020; 177: 539-556
論文で見る
スコープス (28)
PubMed
クロス
グーグル奨学生
テイラー R.G.
レヴィ H.L.
マクインズR.R.
ヒスチダーゼとヒスチジン血症。臨床的および分子学的考察。
Mol. Biol. Med. 1991; 8: 101-116
論文で見る
パブコメ
グーグル奨学生
チャン C.H.
リンC.H.
レーンH.Y.
アルツハイマー病におけるd-グルタミン酸と腸内細菌叢。
Int. J. Mol. サイエンス 2020; 212676
論文で見る
Google Scholar
デレイ J.B.
パタン F.
ラグリュー E.
ル・ティリーO.
Bruno C.
ヴィヨーム M.L.
レイノー M.
ベンツ・デ・ブルターニュ I.
ラウモニエ F.
ヴルクP.
他
血漿アミノ酸プロファイルのポストホック分析:自閉症スペクトラム障害と知的障害における特異的パターンに向けて。
Ann. Clin. Biochem. 2018; 55: 543-552
論文で見る
スコープス (16)
PubMed
クロス
グーグル奨学生
マロッタ R.
リゾレオM.C.
メッシーナG.
パリシL.
カロテヌート M.
ヴェトリ L.
ロッケラM.
自閉症の神経化学。
脳科学 2020; 10163
論文で見る
スコープス(120)
PubMed
クロス
グーグル奨学生
メイネリス-ペルシャックスJ.
カステルス-ノバウA.
アルノリアガ-ロドリゲスM.
マーティン M.
デ・ラ・ベガ-コレアL.
サパタ C.
ブロカス A.
ブラスコ G.
コル C.
エスクリッチA.
他。
プロリン代謝における微生物叢の変化はうつ病に影響を与える。
Cell Metab. 2022; 34: 681-701.e10
論文で見る
PubMed
要旨
全文
全文PDF
グーグル奨学生
松本光弘
大賀 崇
木部 亮
相羽康夫
古賀祐子
辨野義人
低分子代謝産物の大腸吸収は腸内細菌叢に影響される: パイロット研究。
PLoS One. 2017; 12e0169207
論文で見る
スコープス(48)
クロス
グーグル奨学生
Zhu H.
Wang N.
Yao L.
Chen Q.
Zhang R.
Qian J.
Hou Y.
Guo W.
Fan S.
Liu S.
et al.
適度な紫外線曝露は、脳における新規グルタミン酸生合成経路を促進することにより、学習と記憶を増強する。
Cell. 2018; 173: 1716-1727.e17
論文で見る
スコープス (103)
PubMed
要旨
全文
全文PDF
グーグル奨学生
カンポイ C.
マルティン・バウティスタ E.
ガルシア-バルデスL.
フロリード J.
アジル A.
ロレンテ J.A.
マルコス A.
ロペス・サバター M.C.
ミランダ・レオン T.
サンツ Y.
他
Estudio de la influencia de la nutrición y genética maternas sobre la programación del desarrollo del tejido adiposo fetal (Estudio PREOBE) [Study of maternal nutrition and genetic on the foetal adiposity programming (The PREOBE study).
栄養ホスピ2008; 23: 584-590
記事で見る
パブコメ
Googleの学者
ビスカイノJ.A.
Csordas A。
デルトロN.
ディアネスJ.A.
グリスJ.
ラビダス I.
メイヤー G.
ペレス・リベロル Y.
ライジンガー F.
ターネントT.
et al.
PRIDEデータベースとその関連ツールの2016年アップデート。
Nucleic Acids Res. 2016; 44: D447-D456
論文で見る
スコープス (2847)
PubMed
Crossref
グーグル奨学生
ベイリーN.
ベイリー乳幼児発達尺度。
第3版。ハーコート・アセスメント社、2006年
記事で見る
グーグル・スカラー
アルバース C.A.
グリーヴ A.J.
テストレビュー Bayley, N.(2006). Bayley Scales of Infant and Toddler Development-Third Edition (Harcourt Assessment). San Antonio, TX: Harcourt assessment.
J. Psychoeduc. Assess. 2007; 25: 180-190
記事で見る
クロス
グーグル・スカラー
シュロス P.D.
ウェストコット S.L.
リャビン T.
ホール J.R.
ハルトマン M.
ホリスター E.B.
レスニウスキー R.A.
オークリーB.B.
パークス D.H.
ロビンソンC.J.
他
Mothurの紹介:微生物群集を記述し比較するための、オープンソース、プラットフォーム非依存、コミュニティ支援型ソフトウェア。
Appl. Microbiol. 2009; 75: 7537-7541
論文で見る
筑波大学
PubMed
クロス
グーグル奨学生
王 Q.
ガリティG.M.
タイジェJ.M.
コール J.R.
新細菌分類法へのrRNA配列の迅速な割り当てのためのナイーブベイズ分類法。
Appl. Environ. Microbiol. 2007; 73: 5261-5267
論文で見る
筑波大学
PubMed
クロス
グーグル奨学生
ユン S.H.
ハ S.M.
Kwon S.
イム J.
Kim Y.
Seo H.
Chun J.
EzBioCloudの紹介:16S rRNA遺伝子配列と全ゲノムアセンブリーの分類学的に統一されたデータベース。
Int. J. Syst. Evol. Microbiol. 2017; 67: 1613-1617
論文で見る
スコープス (5008)
PubMed
クロス
グーグル奨学生
バンチェフ M.
ルメールS.
サヴィツキーM.M.
Kuster B.
プロテオミクスにおける定量的質量分析:2007年から現在までの最新レビュー。
Anal. Bioanal. Chem. 2012; 404: 939-965
論文で見る
スコープス (614)
PubMed
クロス
グーグル奨学生
グアッツァローニ M.-E.
Herbst F.-A.
ロレス I.
タマメス J.
ペラエス A.I.
ロペス-コルテスN.
アルカイデ M.
デル・ポゾ M.V.
ヴィエイテス J.M.
フォン・ベルゲン M.
ナフタレン暴露と生物刺激に対する細菌の応答を超えたメタプロテオゲノムの洞察。
2013; 7: 122-136
論文で見る
スコープス (95)
PubMed
クロスフィルム
グーグル奨学生
シュナイダーT.
シュミットE.
デ・カストロJr.
カルディナーレ M.
エーベルL.
グルーベ M.
ベルク G.
リーデルK.
肺地衣類(Lobaria Pulmonaria L. Hoffm.)の共生パートナーの構造と機能をメタプロテオミクスで解析。
Proteomics. 2011; 11: 2752-2756
論文で見る
スコープス (100)
PubMed
クロス
グーグル奨学生
タツォフ R.L.
クーニンE.V.
リップマン D.J.
タンパク質ファミリーに関するゲノムの視点。
Science. 1997; 278: 631-637
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
Pang Z.
Zhou G.
Ewald J.
チャン L.
ハカリス O.
Basu N.
Xia J.
グローバルメタボロミクスデータのLC-HRMSスペクトル処理、マルチオミクス統合、共変量調整にMetaboAnalyst 5.0を使用。
Nat. Protoc. 2022; 17: 1735-1761
論文で見る
スコープス (328)
パブコメ
クロス
グーグル奨学生
デシュムク S.S.
ジョンソンJ.L.
クニエリムJ.J.
物体の存在下で採食を行うラットにおいて、海馬周囲皮質は空間情報ではなく非空間情報を表現する:外側嗅内皮質との比較。
Hippocampus. 2012; 22: 2045-2058
論文で見る
スコパス (96)
パブコメ
クロス
グーグル奨学生
ヘルター S.M.
ギャレットL.
アイニッケJ.
Sperling B.
ディルシャール P.
ツィンプリヒ A.
フックス H.
ガイルス・デュルナー V.
フラビェ・デ・アンジェリス M.
ヴルスト W.
マウスにおける認知機能の評価。
Curr. Protoc. Mouse Biol.
論文で見る
スコープス (46)
PubMed
クロス
グーグル奨学生
アッカーマン S.
プリッカーツJ.
スタインブッシュH.W.
Blokland A.
物体認識テスト:統計的考察。
Behav. 2012; 232: 317-322
論文で見る
スコープス (84)
パブコメ
クロス
グーグル奨学生
アイチソン J.
バルセロ-ビダルC.
マルティン-フェルナンデスJ.A.
パウロフスキー・グラーン V.
対数比分析と構成距離。
数学。Geol. 2000; 32: 271-275
論文で見る
スコープス (334)
クロスリファレンス
グーグル・スカラー
論文情報
出版履歴
発行 2023年12月4日
受理 受理:2023年11月7日
改訂版受理 2023年9月26日
受理:2023年9月26日 受理:2023年5月24日
識別
DOI: https://doi.org/10.1016/j.chom.2023.11.004
著作権
© 2023 Elsevier Inc.
サイエンスダイレクト
ScienceDirectでこの記事にアクセスする
図
図サムネイルfx1
グラフィカルアブストラクト
サムネイルgr1
図1Inf-aboveCC(上)とInf-belowCC(下)では、腸内微生物群集の多様性、構造、組成が異なる。
図サムネイルgr2
図2メタプロテオミクス濃縮解析により、乳児の微生物叢における認知に特異的な機能シグネチャーが明らかになった。
図のサムネイルgr3
図3ヒスチジンのメタボロームと乳児の複合認知との関連
図1.
図4Inf-aboveCCドナーの無菌マウスへの糞便移植はInf-belowCCドナーのものより優れた記憶機能を促進する
図1gr5
図5乳児腸内細菌叢をコロニー形成したマウスでは、記憶機能に関連する細菌分類が異なる
図5腸内細菌叢をコロニー形成したマウスでは、記憶機能に関連する細菌分類が異なる。
図6微生物叢はヒスチジン代謝物レベルを調節する
関連記事
広告
研究ジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノム
細胞宿主と微生物
細胞代謝
細胞レポート
セルレポーツ医学
セルレポーツ・メソッド
セルレポート 物理科学
細胞幹細胞
細胞システム
化学
化学触媒
カレントバイオロジー
発生細胞
ヘリオン
免疫
アイサイエンス
ジュール
物質
医学
分子細胞
ニューロン
一つの地球
パターン
STARプロトコル
構造
トレンドレビュージャーナル
生化学
バイオテクノロジー
癌
細胞生物学
化学
認知科学
生態学・進化学
内分泌学・代謝学
遺伝学
免疫学
微生物学
分子医学
神経科学
寄生虫学
薬理学
植物科学
パートナージャーナル
AJHG
生物物理ジャーナル
生物物理学レポート
EBioMedicine
HGGアドバンス
分子植物
分子療法ファミリー
植物通信
幹細胞レポート
イノベーション
著者
論文投稿
複数ジャーナルへの投稿
STARメソッド
プレビュー - プレプリント
査読者
査読者向け情報
ニュース&イベント
ニュースルーム
細胞シンポジウム
コンソーシアムハブ
ウェビナー
ラボリンク
マルチメディア
セルプレスポッドキャスト
セルプレスビデオ
カラーリングとコミック
フィギュア360
セル画ショー
研究篇
セルプレスについて
セルプレスについて
オープンアクセス
COVIDハブ
持続可能性
インクルージョンと多様性
コンタクト
お問い合わせ
ヘルプ&サポート
採用情報
セルプレス採用情報
サイエンティフィックジョブボード
アクセス
登録する
請求
今すぐ読む
司書に推薦する
出版アラート
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
Nucleusコレクション
スナップショット・アーカイブ
インフォメーション
広告主の皆様へ
リクルーターの方へ
図書館員の方へ
プライバシーポリシー
ご利用条件
アクセシビリティ
本サイトのコンテンツは、あらゆる分野の医療従事者および研究者を対象としています。
当サイトでは、サービスの提供・向上やコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を更新するには、このサイトのクッキー設定をご覧ください。
このサイトのすべてのコンテンツ: 著作権 © 2023 Elsevier Inc.
テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビア社に帰属します。
すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条件が適用されます。
プライバシーポリシー 利用規約 アクセシビリティ ヘルプ&サポート お問い合わせ
RELX
この記事が気に入ったらサポートをしてみませんか?