食事と微生物相のクロストークと蠕虫感染症に対する免疫力

食事と微生物相のクロストークと蠕虫感染症に対する免疫力
ローラ・J・マイヒル、アンドリュー・R・ウィリアムズ
初出:2022年12月26日
https://doi.org/10.1111/pim.12965
この記事は出版が認められ、完全な査読を受けていますが、コピー編集、植字、ページ付け、校正のプロセスを経ていないため、このバージョンとVersion of Recordの間に相違が生じる可能性があります。この記事は、doi: 10.1111/pim.12965として引用してください。
Peer Review この論文の査読履歴は、https://publons.com/publon/10.1111/pim.12965 でご覧いただけます。
この記事について
共有する
概要
蠕虫は、ヒトや動物に広く存在する慢性疾患の原因となる大型の多細胞寄生虫である。腸内寄生虫は、宿主の腸内細菌叢や粘膜免疫ネットワークと密接な関係を持ち、その結果、相互の相互作用が感染症の経過に密接な影響を及ぼしている。食餌の構成は腸内細菌叢の構成と腸管免疫機能を強く制御していると考えられ、したがって、抗蠕虫免疫反応の調節に重要な役割を果たすと考えられる。食物繊維など様々な食物成分、免疫細胞、微生物叢の間に存在する多数の相互作用を明らかにすることは、蠕虫に特異的な免疫の制御に新たな光を当てることになるかもしれない。本総説では、蠕虫の感染時に食事成分の代謝がどのように免疫反応を形成するかについての我々の現在の知見に焦点を当て、寄生虫感染症および関連疾患を管理するための新しい治療法を設計するために、この情報をどのように利用できる可能性があるかに焦点を当てている。
はじめに 哺乳類の腸管微小環境は、常在する腸内細菌叢(GM)と宿主の免疫細胞および上皮細胞が密接に関連した複雑なシステムである。さらに、腸管は潜在的な病原性細菌、ウイルス、寄生虫によって絶えず脅かされている。この複雑な生態系が乱れると、慢性的な炎症を引き起こし、感染症への感受性が高まる可能性がある(1)。このように、腸内環境のバランスに影響を与える因子は、健康や疾病に大きな影響を与える可能性があります。腸内細菌の組成を形成する遺伝的および環境的要因の中で、食事は最も強い影響を及ぼす要因の一つであると考えられる(2, 3)。食事成分の違いによって、腸管組織の代謝能や腸管組織に存在する免疫細胞の活性が著しく変化し、腸管免疫の調子を劇的に変化させることがある(4)。これと同様に、現代社会では、食生活の変化(食物繊維の摂取量が減り、脂肪や単糖の摂取量が増える傾向など)により、肥満、大腸炎、アレルギーなど、炎症とディスバイオシスに関連した多くの疾病が生じやすくなっています(4, 5)。
食事の構成は、腸管感染症に対する免疫にも大きな影響を与える可能性がある。寄生虫は、ヒトに最も多く存在する病原体の一つであり、それらが腸内環境とどのように相互作用しているかが解明されつつある。蠕虫が宿主の免疫応答を調節することは古くから知られており、また、感染によって宿主の遺伝子組換え体組成が著しく変化することも現在では知られている(6)。また、宿主の遺伝子組換え状態が蠕虫の感染能力に大きく影響するという相互関係も、現在確立されつつある(7)。このように食事、遺伝子組換え、免疫機能の三者が密接に関連していることから、蠕虫の感染に対する免疫を促進する上で、食事成分の操作がますます重要な役割を果たすと考えられる。本総説では、食餌組成と蠕虫感染との相互作用について論じる。特に、特定の食事成分がどのように蠕虫疾患特異的免疫の発達を形成するか、この過程における遺伝子組み換え食品および遺伝子組み換え食品由来の代謝物の役割、そして抗蠕虫免疫を促進するための標的栄養介入(例えば、プレバイオティクスやプロバイオティクス)の設計の見込みに焦点をあてる。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用される2。蠕虫(ぜんちゅう)感染症-依然として大きな健康問題
蠕虫(ぜんちゅう)感染は、低・中所得国において、ヒトの疾病率の主要な原因の1つである。
蠕虫疾患は、中低所得国の人々や世界中の家畜の疾病の主要な原因の一つです(8, 9)。蠕虫蠕虫の感染率は
蠕虫の有病率は、3つの主要な土壌伝染性蠕虫(回虫、鞭虫、鉤虫)については、ほぼ10億人であると推定されている(10)。
および鉤虫)(10)。蠕虫疾患は、低所得国、近年では中国やブラジルなどの先進国でもよく見られるようになった。
蠕虫疾患は、低所得国、中国やブラジルなどの先進国、そして米国などの高所得国でも、社会から疎外された人々の間でよく見られる疾患です。
米国などの高所得国では、Toxocara canis などの人獣共通感染症の感染が一部で蔓延している(11)。
のような人獣共通感染症の感染が広がっている地域もある(11)。家畜の場合、蠕虫は牧草を主食とする企業ではどこにでも存在する。
家畜の場合、蠕虫は牧草地で普遍的に見られるもので、一般に、世界中の羊やヤギの最も重要な健康上の懸念と考えられています。
ヘモンクシスなどの病気は、食糧安全保障や小型反芻動物飼育の持続可能性を脅かすものであり、世界的に羊やヤギの最も重要な健康上の問題と考えられています(12)。
ヘモンクシスなどの病気は、食糧安全保障と小型反芻動物農業の持続可能性を脅かしています(12)。また、牛、豚、鶏の生産においても、蠕虫は大きな関心事である。
この場合の影響は一般的に不顕性感染に限られるが、それでも効率と収益性に大きな影響を与える可能性がある(9)。
効率と収益性に大きな影響を与える可能性があります(9)。注目すべきは、蠕虫(ぜんちゅう)感染症が、以下のような温室効果ガスの排出に大きく寄与していることです。
このような効率の低下により、家畜からの温室効果ガス排出に大きく寄与していることが示されています(13)。
過去数十年にわたる多大な努力にもかかわらず、蠕虫寄生虫に対するワクチン接種は、ヒトと獣医の双方において依然として
過去数十年にわたる多大な努力にもかかわらず、ヒトと獣医学の双方において、蠕虫蠕虫に対するワクチン接種は依然として問題である。組換え鉤虫ワクチンは臨床試験中であるが
組換え鉤虫ワクチンは臨床試験中であるが、他の蠕虫に対するワクチンは、製薬会社のリソースと関心の不足により、いまだ停滞している。
また、適切なワクチン抗原とその防御免疫機構についての理解も不十分です。
と、ワクチン接種によって最もよく利用されるであろう防御免疫機構についての理解が不十分であるためである(10)。同様に
のワクチンは数カ国で利用可能ですが、他の動物用蠕虫に対するワクチンはまだ開発されていません。
動物用蠕虫類に対するワクチンも同様に供給不足で、ヒト用と同じハードルに直面しています(14)。
14)。従って、蠕虫の感染制御は、現在でもほぼ駆虫薬による治療のみに頼っている。
薬物療法にほぼ依存している。大量に薬剤を投与することは、健康面や衛生面でメリットがあることは間違いないが、その効果は限定的である。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文は、適用されるクリエイティブ・コモンズ・ライセンスが適用されます。
また、既存の薬剤の効果が低い場合もあります(10, 15)。鞭毛虫(Trichuris trichiura)
は、ベンズイミダゾール系などの一般的な薬剤では治療が困難であることが知られており、また、回虫(Ascaris trichiura)に対する治療失敗の報告もある。
また、回虫(Ascaris lumbricoides)に対する治療失敗の報告もある(16)。動物用蠕虫では
レバミゾールやベンズイミダゾールなどの旧来の薬剤に対する耐性が、ほぼすべての地域で高濃度に存在し、状況は危機的である。
レバミソールやベンズイミダゾールなどの旧来の薬剤に対する耐性が、ほぼすべての小反芻動物生産地域で高いレベルで存在し、状況は危機的です(15)。イベルメクチンに対する耐性もまた
イベルメクチンに対する耐性も広く存在し、最近発売された薬剤であるアミノアセトニトリル誘導体に対する耐性も増加している (9, 17)。
9, 17)。
このように、蠕虫の防除は今後も大きな課題となっている。蠕虫と腸内環境の相互作用の理解
蠕虫と腸内環境との相互作用、および腸内環境が宿主の食事やGMなどの要因によってどのように調節されるかを理解することは、2つの点で重要である。
宿主の食事やGMなどの要因によってどのように調節されるかを理解することは、2つの重要な意味を持つと思われる。第一に、これらの外来因子がどのように免疫応答を制御しているのかについて理解が深まることである。
外来因子がどのように免疫応答を制御しているのかを理解することで、防御免疫の基本的なメカニズムに光を当てることができ、将来のワクチン開発にも役立つ可能性があります。
その結果、将来のワクチン開発に役立つ可能性があります。さらに、このような知見により
プロバイオティクス、プレバイオティクス、あるいは免疫系や上皮系の特定の受容体を標的とした新薬など、新規治療薬の開発に道を開くかもしれません。
腸管内の免疫細胞や上皮細胞上の特定の受容体を標的として、免疫反応を促進したり、腸管寄生虫に直接拮抗したりする新薬が開発される可能性もあります。
腸内寄生虫に直接拮抗する新薬の開発への道を開くかもしれない。粘膜免疫機能の形成における食事の役割
腸管バリアは、複雑な配列の吸収性上皮細胞を覆う保護粘液の層で構成されている。
吸収性上皮細胞の複雑な配列を覆う保護粘液層で構成され、その両脇には水とイオンの細胞外輸送を制御するタイトジャンクションがある。
とイオンの細胞間輸送を制御するタイトジャンクションによって支えられている。大腸では、外側の粘液層は緩く付着しており、常在菌を含んでいる。
一方、内側の粘液層は上皮にしっかりと付着しており、腸内細菌と腸管細胞の間に不浸透性のバリアーを形成している。
のバリアを形成している(18)。吸収性の腸管細胞に加えて、上皮には貪食細胞も存在する。
上皮には、吸収を担う腸細胞のほかに、杯細胞、パネス細胞、腸内分泌細胞、房細胞などが存在する。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文は、該当するクリエイティブ・コモンズ・ライセンスが適用されます。粘液分泌、抗菌ペプチド産生、自然免疫反応の調整における役割
を担っている。さらに
常駐するセンチネル免疫細胞(樹状細胞やマクロファージ)は、内腔内容物を継続的に採取し
さらに、常駐するセンチネル免疫細胞(樹状細胞とマクロファージ)は、腸管内容物と腸内細菌抗原を継続的にサンプリングし、腸内細菌の細胞との免疫応答を調整する。
さらに、常駐するセンチネル免疫細胞(樹状細胞とマクロファージ)は、腸管内容物と腸内細菌抗原を絶えずサンプリングし、腸管関連リンパ組織の細胞との間で免疫反応を調整し、腸の恒常性を維持している。
このように、腸管上皮は微生物と免疫細胞の相互作用のための肥沃な土地であり、微生物(病原体も含む)による免疫調節が行われています。
微生物(病原性および常在性の両方)、環境および食事性抗原の刺激による免疫調節のための肥沃な土壌である。
刺激による免疫調節が行われる。
粘膜リンパ球は、粘膜病原体に対する個別の機能を持つ細胞の緊密な組織化されたネットワークである。
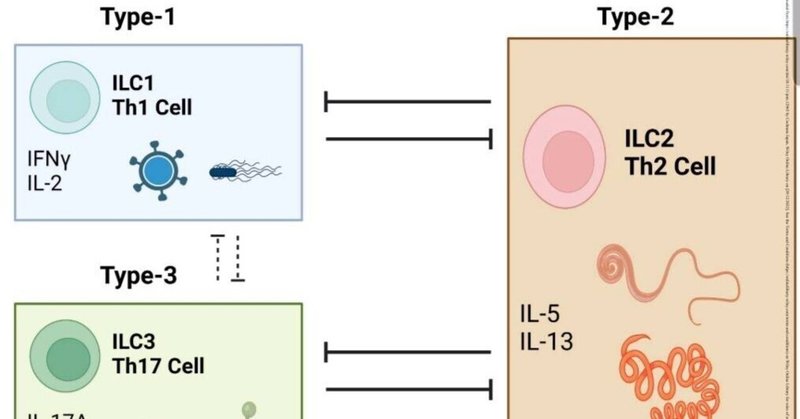
粘膜リンパ球は、粘膜病原体に対する個別の機能を持つ細胞ネットワークである。タイプ1およびタイプ17の反応は、T-ヘルパー(Th1)-1細胞およびTh17細胞によって媒介される。
タイプ1およびタイプ17反応は、Tヘルパー(Th1)-1およびTh17細胞と、タイプ1およびタイプ3の自然免疫系細胞(ILC1およびILC3)によって担われ、主にウイルスや細菌の感染に対する防御を行う。
主にインターフェロンや炎症性サイトカイン(IL-6やIL-17など)を産生することで、ウイルスや細菌感染から身を守ります。
IL-6やIL-17などの炎症性サイトカインを産生することでウイルスや細菌感染を防御する(19)。逆に、Th2細胞や2型自然免疫リンパ球(ILC2)は、蠕虫に対する免疫に関与している。
のような2型サイトカインを放出することによって特徴づけられる、異なるエフェクター機構を引き起こすことによって、蠕虫に対する免疫に関与している。
IL-5やIL-13などの2型サイトカインが放出され、粘液分泌や上皮細胞のターンオーバーを促進することが特徴である。
のような2型サイトカインを放出することを特徴とするエフェクター機構を引き起こす。これらの免疫細胞の反応に異常が生じると、炎症性腸疾患のような様々な自己免疫疾患につながる
これらの免疫細胞の反応の異常は、炎症性腸疾患やアレルギーなどの様々な自己免疫疾患や、様々な病原体に対する逆調節作用につながる(図1)。
図1)。
自然免疫細胞および適応免疫細胞の活性は、栄養素(例えば、アミノ酸やビタミンなど)に対して高い反応性を示す。
自然免疫細胞も適応免疫細胞も、アミノ酸やビタミンなどの栄養素や遺伝子組み換え食品由来の代謝産物に高い反応性を示し、食事による免疫調節のメカニズムが明らかにされている(3)。
を介した免疫調節の基礎となるものである(3)。食事による宿主免疫の調節は、食事由来の代謝物を介して行われることがある。
食餌性免疫調節は、食餌性成分の微生物分解産物である食餌性代謝産物(GM依存性、または「プレGM」)を介して行われる。
(GM依存性、または「プレバイオティック」)、または食事成分そのものによる上皮細胞や免疫細胞の直接的な調節(GM非依存性)を介して、宿主の免疫を調節することができる。
GM非依存的)。哺乳類のGMは、膨大な数の微生物のコンソーシアムをホストしています。
の利用を含むいくつかのメカニズムを通じて、宿主の健康維持に貢献している。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規約はWiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は、適用されるクリエイティブ・コモンズ・ライセンスに準拠し、アクセス可能な食事成分、必須栄養素の供給、腸管上皮バリア機能と統合性の維持が規定される。
を提供し、腸管上皮のバリア機能および完全性を維持する。大腸内細菌叢による食物繊維の分解は、例えば、以下のような結果をもたらします。
例えば、大腸菌による食物繊維の分解は、酢酸、プロピオン酸、酪酸などの短鎖脂肪酸(SCFA)の産生につながる。
酪酸などの短鎖脂肪酸が生成される。これらの代謝産物は、腸管上皮細胞の豊富なエネルギー源となり、さらに
また、上皮のGタンパク質共役型受容体(GPCR - GPCR42, GPCR43)に直接結合し、腸管バリア機能を変化させることもある(23)。
バリア機能を変化させる(23)。SCFAおよびその他のGM由来代謝物もまた、強力な免疫調節作用を有している
を持つ。SCFAはヒストン脱アセチル化酵素の活性を阻害することにより、免疫細胞の活性を変化させ、それにより
炎症性サイトカインの放出を調節し、粘膜バリアにおける免疫学的寛容の維持に役立つ。
粘膜バリアーでの免疫寛容の維持に寄与している (24-26)。免疫調節活性が知られている他のGM由来代謝物
NFκB 活性化物質であるトリメチレンN-オキシドなど、コリン分解物も免疫調節活性を持つことが知られている(27)。
(27). デオキシコール酸などの二次胆汁酸は、一次胆汁酸の微生物による脱共役化によって生成される。
デオキシコール酸などの二次胆汁酸は、Bifidobacterium属などの微生物が一次胆汁酸を脱共役することによって生成され、また、ビフィズス菌などの微生物群によって
は、樹状細胞やマクロファージによる炎症性メディエーターの産生を制御し、T-レギュラトリー(Treg)細胞を誘導します(28)。
制御性(Treg)細胞を誘導することができる(28, 29)。免疫細胞活性の調節に加えて、発酵由来の代謝物自体が
発酵由来の代謝産物は、クロスフィーディングにより腸内細菌叢の組成を変化させ、酢酸などのSCFAを産生する。
酢酸などのSCFAは、乳酸菌やフェカリバクテリウム属などの他の細菌によって利用される。
乳酸菌(LAB)は免疫細胞と直接相互作用することができ、宿主の免疫機能や耐性に有益な影響を与えることがよく知られている。
乳酸菌は、宿主の免疫機能や寛容性反応に有益な影響を与えることがよく知られています。樹状細胞のモジュレーション。
を介した樹状細胞のモジュレーションは、常在菌のLABがin vitroで免疫細胞の機能を変化させるメカニズムの一つであることが知られている(30)。
の機能を変化させるメカニズムの一つであることが知られている(30)。
食事成分や栄養素の中には、それ自体が宿主の免疫細胞を直接修飾するものもある。
は、宿主の免疫細胞を直接調節することができる。
マクロファージや樹状細胞が上皮細胞の間に細胞突起を形成する結果、GMとは無関係に宿主の免疫細胞を直接変調させることができる。
が上皮細胞間の細胞突起を突き出し、管腔内の内容物を採取することにより、GMとは無関係に宿主の免疫細胞を直接修飾するものもある(31, 32)。食事成分と腸管上皮の直接接触
食事成分と腸管上皮細胞との直接接触もまた、腸の形態に大きな影響を与える可能性がある。
機能および病原体の侵入を防御する能力に大きな影響を与える可能性がある。例えば、食事で摂取したn-3系多価不飽和脂肪酸は、腸の形態や機能に大きな影響を与える。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文は、該当するクリエイティブ・コモンズ・ライセンスが適用されます。脂肪酸(PUFAs)は、絨毛の長さと粘液の厚さを増加させる一方で、分泌性上皮細胞の数を減少させます。
の数を減らす一方で、絨毛やパネス細胞などの分泌性上皮細胞の数を増やした(33)。
食餌の構成が免疫機能に及ぼす影響は深刻であろう。高脂肪、低ショ糖の食事は
脂肪とショ糖が多く、食物繊維やその他の植物性多糖類が少ない食事(すなわち「西洋」食)は、GMを変化させる。
は、ファーミキューテス/バクテロイデーテス比を大幅に増加させ、食事からエネルギーを獲得し、植物多糖類を蓄積する代謝経路を促進することによって
という代謝経路を促進することによってGMを変化させる(34-36)。食事誘発性肥満のマウスモデルにおいて
肥満モデルマウスでは、欧米食の摂取により炎症性マクロファージの分化が誘導され、その結果、慢性的な炎症(特に脂肪細胞)が引き起こされる。
慢性炎症(特に脂肪組織)、T2糖尿病の発症、インスリン抵抗性を引き起こす(34)。
抵抗性をもたらす(34)。興味深いことに、肥満の結果生じる低悪性度の慢性炎症は
の結果、好酸球や交互に活性化されたマクロファージなどの2型免疫細胞が著しく減少している(37)。
マクロファージが減少していることが知られています(37)。さらに、ILC2は、脂肪細胞の代謝を直接制御するペプチドを分泌している。
また、ILC2は、脂肪細胞の代謝を直接制御するペプチドを分泌し、脂肪組織におけるビーギング(エネルギー消費)を促進し、脂質の蓄積を抑制する(38)。
2型応答の障害が肥満のカジュアルファクターであるという考えと一致するように、食事により
肥満(および耐糖能などの関連する免疫パラメータ)は、蠕虫であるリトアニア菌の感染によって改善することができる。
Litomosoides sigmodontisという蠕虫の感染により、好酸球依存的に肥満が改善されることがわかった(37)。このように
食事とGMの相互作用は、異なる粘膜免疫細胞サブセットのバランスを著しく変化させる可能性がある。
(例えば、1型と2型のように)、それによって異なる腸管病原体に対する抵抗性や感受性を決定しているのかもしれない。
蠕虫感染症における食事-遺伝子-免疫のクロストークの例
4.1 栄養、食事と蠕虫族
蠕虫感染の特徴的な症状は栄養失調であるが、感染がどの程度の栄養ストレスを引き起こすかは、その程度に依存する。
栄養ストレスの程度は、感染の強さや期間、寄生虫の種類だけでなく、年齢などの宿主要因にも依存する。
感染による栄養ストレスの程度は、感染の強さや期間、寄生虫の種類だけでなく、年齢などの宿主要因にも依存する。例えば、ヒトの場合、T. trichiuraは小児期の栄養不良により顕著な影響を与える可能性がある。
lumbricoidesよりも顕著に小児期の栄養不良に影響を与える可能性があります(8)。そのため、栄養学的な観点からの研究が必要である。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. Wiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照してください。OA論文は、該当するCreative Commons Licenseパラサイトの相互作用が長年にわたって実施されています。豚のアスカリス・スム(Ascaris suum)感染を利用した初期の実験では
豚のアスカリス感染を利用した初期の実験では、蠕虫の感染が実際に顕著な栄養代謝の変化と関連することが証明された(39)。
栄養代謝が著しく変化することが示された(39)。より最近の研究では、野生のキジネズミの集団に標準的なペレット飼料を与えたところ
標準的なペレット状飼料(自然食に加えて)を与えることで、自然食に対する抵抗力が向上した。
Heligmosomoides polygyrus 感染に対する抵抗性を向上させた。さらに、駆虫剤で処理されたマウスに補充した飼料を与えたところ、免疫応答が強くなり
さらに、駆虫剤を投与したマウスは、免疫反応と再感染に対する抵抗力が強くなり、その結果、H. polygyrusの感染も抑制された。
H. polygyrus 感染の可能性を減少させた(40)。この研究により
この研究は、感染に対する宿主の効果的な反応には、バランスの取れた食事が有効であることを示している。宿主の免疫力
は、大栄養素と微量栄養素の状態に依存していることをさらに「ピンポイント」に指摘できる (41-43)。
マウスと子羊を使った研究では、タンパク質が乏しい餌を与えられた動物は、感染に対する感受性が高まり
マウスと子羊の研究により、タンパク質が乏しい餌を与えられた動物は感染に対する感受性が高まり、それに伴って寄生虫特異的な抗体や免疫細胞の産生が低下することが明らかにされている
また、タンパク質を補給することで、これらのパラメータと感染抵抗性が速やかに回復することも明らかにされている(44, 45)。
これらのパラメータと感染に対する抵抗力を回復させることがわかった(46)。同様に、亜鉛などの微量栄養素の欠乏も、感染症への抵抗力を低下させる。
亜鉛などの微量栄養素の欠乏も蠕虫に対する免疫力の低下と関連している(47)。
動物用蠕虫の駆虫薬耐性が急速に高まっていることから、感染負荷を下げる手段として食餌組成を操作すること
動物用蠕虫の駆虫抵抗性が急速に高まる中、感染負荷を低減する手段として、食餌組成の操作が1990年代に盛んに研究されるようになった。
1990s. それ以前の研究では、マウスにおける H. polygyrus 感染の経過は、食餌組成そのものに大きく影響されることが示されていた。
その結果、精製された半合成飼料(粗繊維の少ない飼料)を与えると
(粗繊維が少ない) 精製半合成飼料は市販のペレット状飼料に比べて感染率が低かった (48) 。
野外条件下での研究(商業農場から自然感染した豚)で、以下のことが判明した。
食物繊維(非デンプン性多糖類)の含有量とAscaris suumおよびAscaris suumの有病率の間に正の相関があることが判明した。
Ascaris suum およびTrichuris suis 感染の有病率との間に正の相関関係があることがわかった (49). その後、豚の感染を制御して観察したところ
その後、豚のコントロールされた感染実験では、飼料成分によってA. suumとOesophagostomum dentatumの感染強度が異なることが示された(50)。
の感染強度が異なることが示され (50)、「栄養学的駆除」を達成するための飼料成分の操作の可能性が開かれた。
を操作して「栄養学的駆除」を達成する可能性が出てきた。これは O. dentatum の感染で劇的な形で達成された。
不溶性食物繊維を発酵性の高い炭水化物に置き換えることで、ほぼ完全な脱虫が達成された。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. このような場合、「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」「痒いところに手が届く」。さらに、ある種の牧草を食べた反芻動物が、虫の発生を著しく減少させるという観察結果も得られました。
さらに、特定の牧草を食べた反芻動物が寄生虫を著しく減少させるという観察から、食餌に含まれる植物代謝物を特定する生産的な研究分野が開かれた。
さらに、特定の牧草を食べた反芻動物が寄生虫の数を著しく減らすという観察から、食餌に含まれる植物代謝物が抗寄生虫作用を持つことを特定する生産的な研究分野が始まった。その結果、凝縮タンニン(例:セインフォイン)、セサミオイル(例:セイヨウノコギリソウ)が同定されました。
セインフォイン)、セスキテルペンラクトン(チコリなど)を含む植物が発見され、現在では、寄生虫の多い地域における防除の補完的な選択肢になっている。
現在では、駆虫剤耐性の強い地域や薬剤治療が困難な地域における虫害対策の補完的な選択肢となっている。
を含む植物が、駆虫薬耐性の高い地域や薬物治療が問題となる地域(例:有機農業システム)において、虫の駆除のための補完的な選択肢となっている(52、53)。このように、食事要因が蠕虫の治療結果の形成に大きな役割を果たすことは明らかである。
このように、食事が蠕虫感染の結果を形成する上で主要な役割を果たすことは明らかである。
4.2 プレバイオティクス食品成分と蠕虫に対する免疫力
食餌性化合物のプレバイオティクス効果およびGMに依存した粘膜免疫機能の変化についての理解が進んだことにより、蠕虫に対する免疫力の向上が疑問視されている。
粘膜免疫機能における遺伝子組み換え依存性の変化に関する理解が進んだことで、食物繊維などのプレバイオティクス食品成分が、遺伝子組み換え食品と相互作用して抗蠕虫機能を形成できるかどうかという疑問が生じました。
と相互作用し、抗蠕虫免疫を形成することができるのかという疑問が生じました。その結果
宿主のGM組成は蠕虫の感染時に劇的に変化するが、蠕虫の感染に反応して常に変化する核となる分類群は
しかし、異なる蠕虫に反応して一貫して変化する核となる分類群はまだ定義されていない(54-57)。このような
虫の定着にGMが重要であることは、マウスや子豚を用いた研究で証明されている。
マウスや子豚を用いた研究で、鞭毛虫(大腸に生息するTrichuris属)の定着が
の定着が著しく低下するか、あるいは完全に消失することが示された。
GMの存在が必要であることが示唆された(7, 58)。さらに、蠕虫によって
さらに、蠕虫によって誘発されたGMの変化が、感染による免疫学的作用のいくつかに関与しているという概念
さらに、蠕虫に感染したマウスから無菌マウスにGMを移植すると、感染による免疫学的影響の一部が再現されることが示された。
を生殖細胞非接種マウスに移植したところ、蠕虫に起因する免疫機能のいくつかの特徴が再現された。
特にTreg応答が顕著であった(59, 60)。注目すべきは、無菌マウスでは一般的に
型免疫応答を示すが、これは腸内細菌のコロニー形成により徐々に減少する。
これは、3型RORγt+ TregやTh17細胞の誘導が一因である(61)。これに対して
抗生物質処理によるGMの擾乱の後、GMの再コロニー化(co-colonization)により誘導される。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文には、該当するクリエイティブ・コモンズ・ライセンスが適用されます)。
murisのクリアランスを低下させる(62)。小腸回虫H. polygyrusは、微生物相の存在によって恩恵を受けるようである。
GMによって誘導されたTregがTh2細胞を抑制するため、小腸回虫H. polygyrusは微生物叢の存在に恩恵を受けるようである。
を抑制し、その結果、無菌マウスに比べ、虫の繁殖力が向上した(63)。しかし、最近の報告では
しかし、最近の報告では、無菌マウスや抗生物質処理マウスにH. polygyrusを感染させると、虫体量が増加し、腸内環境の悪化により免疫力が低下することが示唆されている。
しかし、最近の報告では、無菌マウスや抗生物質投与マウスにH. polygyrusを感染させると、腸の蠕動運動が低下するため、虫体量が増加し、免疫力が低下することが示唆されている(64)。このように、蠕虫に対する宿主の反応に及ぼすGM組成の影響は
蠕虫に対する宿主の反応に及ぼすGM組成の影響は、実験デザイン、感染量および/または期間、および
また、大腸菌(T. muris)と小腸菌(T. muris)では、状況が大きく異なる可能性がある。
(muris)と小腸に棲息する蠕虫の間で状況が大きく異なる可能性がある。まとめて
これらの研究
寄生虫と宿主GMのダイナミックな相互作用が免疫機能に影響を及ぼしていることが明らかになった。
このクロストークをプレバイオティクスによって操作することで、免疫防御反応を促進できる可能性があります。
保護的な免疫反応を促進する可能性を示しています。
プレバイオティクスがどのように抗蠕虫免疫に影響を与えるかは、現在解明されつつあるところです。
遺伝子組み換えと免疫のクロストークは複雑であるため、まだまだ解明されることはないだろう。直接
GM由来の代謝産物が蠕虫に対する防御を媒介するという直接的な実験的証拠は、Petkeviciusらによって提供された。
Petkeviciusらは、水溶性食物繊維がO. dentatumに対して前述の抗寄生虫効果を発揮することを示した。
に対する水溶性食物繊維の前述の抗寄生虫効果が、SCFA をブタの結腸に直接注入することで再現されることを示した。
豚の大腸に直接注入することで再現できることを示した (65). SCFAやその他多くのGM由来の分子が特異的な免疫調節特性を持つことを考えると
このことは、多糖類を分解して生物活性代謝物を生成させることが、免疫反応を引き起こす戦略であることを示唆している
は、望ましい抗蠕虫免疫反応を引き起こすための戦略である可能性がある。このように
このように、微生物が利用しやすい糖質を豊富に含む食餌によってGMを改変することは、免疫機能を高めることで感染症から身を守ることができるかもしれない。
免疫機能を高めることによって、感染症から身を守ることができるかもしれない。これと一致して、Schneiderらは、発酵性繊維を豊富に含む飼料を与えることで、感染症から身を守ることができることを示した。
Schneiderらは、発酵性繊維を豊富に含む飼料を与えることで、腸内環境の感知と免疫機能の強化に関与する重要な細胞であるタフト細胞が増殖することを明らかにした。
タフト細胞は、腸内環境を感知し、IL-25とロイコトリエンを放出することで2型免疫を活性化する重要な細胞である。
ロイコトリエンを放出し、ILC2を活性化する。これらの著者らは、高繊維食によって
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規約はWiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は、該当するクリエイティブ・コモンズ・ライセンスに準拠する。腸内常在菌Tritrichomonas murisの拡大により、酢酸とコハク酸が生成され、GPR91を介してタフト細胞が活性化された。
GPR91受容体を介してタフト細胞を活性化する。しかし、発酵性食物繊維を含まない飼料を与えたマウスでは、小腸の房状細胞が著しく減少していた。
しかし、発酵性食物繊維を含まない飼料を与えたマウスでは、小腸の房状細胞が著しく減少していた。したがって、発酵性炭水化物の摂取により、房細胞-ILC2軸が活性化され
は、2型免疫を誘発する可能性がある。しかし、Myhillら
らは、発酵性食物繊維(イヌリン)を多く含む飼料を与えたマウスは、T. murisの感染が
murisへの感染が、不溶性食物繊維(セルロース)を含む飼料を与えたマウスと比較して、大幅に増加したことを明らかにした(68)。GMは
は、Actinobacteria(放線菌)門とAkkermansia(粘菌)門に富んでいた。
が濃縮され、糞便中のSCFA濃度が上昇した。
プレバイオティック多糖類のポジティブな効果に特徴的である。しかし、免疫学的解析の結果、T. muris感染時にイヌリン
T. muris感染時のイヌリン補給は2型応答を著しく低下させることが示された。
このことは、プレバイオティクス繊維が防御免疫力を高めるのではなく、むしろ抑制していることを示している。これらの観察結果の背景にあるメカニズムは
これらの観察結果の背景にあるメカニズムはまだ明らかではないが、プレバイオティック食物繊維と感染に対する免疫反応の相互作用が
食物繊維と感染に対する免疫反応との相互作用は、複雑で、おそらく文脈依存的であることが予想される。
依存する可能性があります。このことと矛盾しないように、T. murisを用いた結果とは対照的に、発酵性食物繊維は他のマウス病原体の感染除去を助ける。
のような大腸の他のマウス病原体の感染クリアランスを助ける。
Citrobacter rodentium(シトロバクター・ローデンティウム)など、大腸の他の病原体の感染除去を助ける(69、70)。このモデルでは、抗細菌性免疫応答は食物繊維の摂取中も
このことから、イヌリンを摂取したマウスにおけるT. murisに対する免疫の阻害は、蠕虫によって誘導されるタイプ2の反応に特異的であることが示唆される
に対する免疫の抑制は、蠕虫によって誘導されるタイプ 2 の反応に特有であることが示唆された。興味深いことに、実験用マウスを再野生化すると、タイプ2に偏った免疫反応から
マウスを再び野生化させると、2型に偏った免疫反応から1型に偏った免疫反応へと変化し、T. murisへの感受性も同様に変化する(71)。
murisに感染しやすくなる(71)。再野生化の過程で、より多様な「自然」食を摂取できるようになったと考えられる。
再野生化の過程で、より多様な「自然」食を摂取できるようになることを考えると、複雑な植物繊維や変化したGMの摂取へと徐々にシフトしていくことが、T. muris感染の抑制と関連しているのかもしれない。
GMを摂取するようになったことが、T. muris感染時の2型応答の減衰に関係しているのかもしれない。しかし、他の感染モデルにおいて、相反する
しかし、他の感染モデル、例えば、Trichinella spiralisに感染させたマウスに
β-グルカン(これも食餌性オリゴ糖)を与えたマウスは、β-グルカンが介在することによって感染から保護されることがわかった。
グルカンを介したA. muciniphilaのGM内での拡大が、TLR2依存性の免疫機構を引き起こし、虫の排出を促進することによって、感染から守られることがわかった(72)。
を誘発し、虫の排出を促進することがわかった(72)。これらの研究結果を総合すると、A. muciniphilaの感染経路はβ-グルカンであることが示唆される。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文については、該当するクリエイティブ・コモンズ・ライセンスが適用される。
この関係を理解するためには、いくつかの課題が残されています。この関係を理解するためには、いくつかの課題が残されている。
寄生虫の種類や宿主の種類によって、食事介入に対する反応に違いがある可能性がある。
また、腸内寄生虫の位置(小腸と大腸)も重要であろう。これらの相互作用を解明するためのさらなる
これらの相互作用を解明するためのさらなる研究の優先順位は高い。
4.4 特定の代謝産物-受容体相互作用
食事摂取による粘膜免疫機能の変化は、特定の代謝物の産生によって媒介される可能性がある。
特定の受容体に対するリガンドとして作用する特異的な代謝物の産生が介在している可能性がある。
免疫調節を行う食事成分や代謝物が免疫に及ぼす影響を模倣した薬物治療の可能性がある(図2)。
図2)。GMが食餌成分の発酵中に産生する膨大な数の腸内代謝産物は
粘膜免疫細胞は、免疫反応を感知して反応するように高度に発達しているため、蠕虫の感染に重要な影響を与える可能性が高い。
細胞は、代謝や栄養の擾乱を感知し、それに反応するように高度に発達している。例えば
例えば、ILCは高度に制御された代謝ネットワークを持ち、細胞内のアミノ酸レベルの変化を素早く感知し、それに反応する。
また、特定のアミノ酸受容体を欠損したマウスでは、その機能が著しく変化している。
また、特定のアミノ酸受容体を欠損させたマウスでは、機能が著しく変化する(73-75)。興味深いことに、免疫細胞の活動もまた、概日リズムによって調整されている
興味深いことに、免疫細胞の活性は、概日リズムと食物摂取量によって調整される。ILC3活性は食物摂取に応答して抑制される。
腸管ニューロン依存的に、食物摂取に反応してILC3の活性が抑制される。これはおそらく、栄養調節に役立つ分節化された糸状菌の活動を可能にするためであろう。
糸状菌の活動を可能にし、栄養吸収や脂質代謝を調節していると考えられる(76)。ILC3の欠損
日内リズムのマスターレギュレーターであるBMAL1が欠損したILC3は、過活動でアポトーシスを起こしやすいことから
このことは、栄養素と環境の両方を感知することで、腸内の免疫細胞(ILC3など)の活性が調整されることを示している。
ILC3などの)腸内免疫細胞の活性を調整することを示唆している(77)。概日時計もまた、寄生虫に対する免疫反応と密接に関連しているようであることを考えると
を考えると、食物摂取量や日内リズムに応じた特定の腸内代謝産物の産生は、免疫細胞が寄生虫に対して効果的に反応するようプログラムされているのかもしれない。
免疫細胞が病原体に効果的に反応するようにプログラムされているのかもしれない。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規約はWiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用される。蠕虫感染の文脈では、自然免疫細胞の異なるクラスが、変化する病原体に異なる反応をすることが特に重要である。
このような場合、自然免疫細胞は、ある種の栄養素や代謝物のレベルの変化に対して異なる反応を示すように見えることが、蠕虫感染との関連で特に重要です。一例として
異なる病原体に対する免疫反応を制御する役割を果たすと思われる特異的な代謝産物感知受容体の一例として
アリール炭化水素受容体(AhR)は、腸管バリアー表面に高発現する転写因子であり、さまざまな病原体に対する免疫反応を制御する役割を担っている。
これは、腸管バリアー表面に高レベルで発現している転写因子で、異種物質や多くの異なる食餌由来の代謝物を感知する(79)。
を感知する転写因子である(79)。AhRシグナルは、細菌または蠕虫の感染に対する適切なILC応答を制御する上で重要な役割を果たす可能性がある。
シグナルは、細菌または蠕虫の感染に対する適切なILC反応を制御する上で重要な役割を果たすと考えられる。AhR欠損マウスは、C. rodentium感染に非常に感受性が高い。
しかし、H. polygyrus感染にはより抵抗性である。さらに、ILC2特異的なAhRの切除は、抗蠕虫免疫を促進する。
さらに、ILC2特異的なAhRの欠損は蠕虫に対する免疫を促進するが、ILC3のAhR欠損はC. rodentiumに対する抵抗性を消失させる(80)。
このように、自然免疫細胞の相反するクラスの活性が、食事感受性受容体によって制御される経路が明らかにされた
の活性を調節する経路を示した。このように、ある特定の受容体を腸内代謝産物でライゲーションすることで
これは、腸内代謝物によって特定の受容体がライゲーションされると、2型免疫が犠牲になって3型反応が選択的に促進されるという具体的な例を示している。
食事や遺伝子組み換え食品に由来する手がかりが、自然免疫細胞のタイプ1/3とタイプ2の間の「スイッチ」を作動させるというメカニズムが示唆された。
の機能を切り替えるというメカニズムを示唆している。一般的なAhRリガンドには、以下のような植物栄養素が含まれる。
グルコシノレート由来化合物などの植物栄養素や、遺伝子組換え食品によるトリプトファンの分解で生じる代謝物など
トリプトファンの分解によって産生される代謝物がある。特に、発酵性食物繊維(レジスタントスターチなど)の摂取は、GMタキソプラズマの増殖を促進することが知られています。
トリプトファンを積極的に分解する乳酸菌のようなGM分類群の増殖を促進することが知られている。
のようなAhRリガンドを産生する乳酸菌のようなGM分類群の増殖を促進することが知られている(81)。このことは、場合によっては、なぜ
このことは、発酵性食物繊維を大量に含む食事を摂取した場合、抗蠕虫性免疫の働きが弱まることがあることを示唆しているのかもしれない。
このことは、発酵性食物繊維を大量に含む食餌を摂取した場合に、抗蠕虫免疫の働きが弱まる場合があることを示唆していると考えられる。もう一つの例はビタミンAで、食事中の濃度が異なると、腸内のILCの反応も多様になる。
の反応も異なる。食事中のビタミンA濃度が高いと、3型ILC(ILC3)が優位になり、C. rodentium感染に対する抵抗性が高まる。
一方、ビタミンAが不足すると、2型ILC(ILC2)が優位になり、虫のコロニー形成に抵抗性を示すようになる(図1)。
が優勢となり、虫のコロニー形成に抵抗性を示すようになる(82)。このような状況は、腸内環境を強化するための進化的な適応から生まれたのかもしれない。
この状況は、栄養不足の時代に、どこにでもいる寄生虫に対する腸管バリア防御を強化するために、進化的に適応して生まれたのかもしれない。
このような状況は、栄養不足の時期に寄生虫に対する腸管バリア防御を強化するための進化的適応から生じたと考えられる。このように、食物成分の生理活性代謝物への分解は複雑である。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文は、該当するクリエイティブ・コモンズ・ライセンスが適用され、腸管免疫細胞に機能的に多様な影響を与え、異なる病原体クラスに対する免疫の発現に重要な影響を与える。
に対する免疫の発現に重要な影響を及ぼします(図2)。
このような食事-代謝物-免疫経路を実証的に明らかにするためには、まだ多くの課題が残されています。
腸管感染症の免疫系を制御する代謝産物を同定することは
腸管感染症において免疫機能を制御する代謝物を同定することができれば、薬理学的介入によって食事の効果を模倣し、効果的な免疫反応を促進する前例のない機会を提供できるかもしれません。
免疫反応を効果的に促進するための薬理学的介入によって、食事の影響を模倣する前例のない機会を提供することができるかもしれない。前述のように、コハク酸は微生物の代謝物の 1 つであり
型免疫反応を促進する上で重要な役割を果たすことが既に示されている。
ハイスループットなメタボロームプロファイリングの急速な進歩により、さらなる候補物質の検出と検証が可能になるはずである。
検出され、検証されるはずである。例えば、胆汁酸と結合して胆汁酸塩を形成するアミノ酸の一種であるタウリンは、最近、胆汁酸と結合して胆汁酸塩を形成することが明らかになりました。
胆汁酸と結合して胆汁酸塩を形成するアミノ酸であるタウリンは、最近メタボロームプロファイリングにより、病原性細菌のコロニー形成抵抗性を損なうGM由来の代謝物であることが明らかになりました。
が病原性細菌に対するコロニー形成抵抗性を低下させることが、メタボロームプロファイリングにより明らかになりました(83)。これらの回路を、2型免疫との関連で利用することで
2型免疫の文脈でこれらの回路を利用して望ましい免疫反応を引き起こすことは、栄養免疫学の新たなパラダイムを示す可能性がある。
栄養免疫学の新たなパラダイムとなり、さまざまなタイプの病原体に対する粘膜免疫反応を微調整する精密栄養学的アプローチが可能になる。
栄養学的アプローチにより、様々なタイプの病原体に対する粘膜免疫応答を微調整したり、制御不能な免疫極性を修正したりすることが可能になる(例えば、炎症性腸疾患やアレルギーなど)。
炎症性腸疾患やアレルギーなど)。
4.5 プロバイオティクス細菌と蠕虫(ぜんちゅう)感染症
過去20年間、マイクロバイオーム分野の最大の成果の1つは、「有益な」細菌、すなわちプロバイオティクスの利用である。
細菌、すなわちプロバイオティクスを使用して、ディスバイオーシスを修正し、バランスのとれた免疫機能を回復させ、炎症性疾患を緩和することである。
炎症性疾患を緩和することです。一般的なプロバイオティクスの候補は、乳酸菌や腸内細菌などの乳酸菌です。
Enterococcus faeciumなどの乳酸菌、および放線菌門のビフィズス菌である(84, 85)。これらの
これらの細菌は、健康を増進する食事成分に反応して増殖することが知られている。例えば
イヌリンおよび関連する食物繊維は、ビフィズス菌の増殖を強く刺激する(86)。その結果
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. OA論文は、該当するクリエイティブ・コモンズ・ライセンスが適用されます。プロバイオティクス細菌の供給は、多くの前臨床感染モデルにおいて免疫力を高めることが示されています。
を提供することで、細菌およびウイルス感染症の多くの前臨床感染モデルにおいて免疫力を高めることが示されている(87-89)。
プロバイオティクス細菌が蠕虫に対する免疫を調節する役割を持つかどうかはまだ明らかではないが、様々な食事成分の影響が大きいことを考えると
異なる食事成分が蠕虫感染に対する免疫反応に大きな影響を与えることを考えると、プロバイオティクス治療が蠕虫に対する免疫反応を調節することが合理的に期待される。
プロバイオティクスによる治療が、蠕虫感染に対する免疫反応に大きな影響を与えることを考えると、その効果の一部を再現することが期待できるかもしれない(有益なものも有害なものもある)。
を再現することが期待される。) 現在までのところ、この結果はやや矛盾している。
を投与したマウスは、T. muris 感染に対してより感受性が高いことが示された(90)。
murisに感染しやすいことが示された(90)。一方、Lacticaseibacillus rhamnosus(JB-1)を投与したマウスは、同じ寄生虫に対する免疫が強化された。
一方、Lacticaseibacillus rhamnosus(JB-1)を投与したマウスは、IL-10依存的に同じ寄生虫に対する免疫力を向上させた(91)。T. spiralis感染症は
A. muciniphilaの熱処理による免疫強化により、T. spiralis感染を抑制することができる(72)。
(一方、Lactobacillus lactisは、マウスにおけるA. suumの肺病期に対する免疫を、sIgA産生を促進することにより、向上させることができる(92)。
sIgA産生を促進することにより、マウスの A. suum の肺病期に対する免疫を促進する可能性がある (92) 。豚の蠕虫感染モデルにおいて、いくつかの研究が以下のことを示した。
プロバイオティクスは、蠕虫に特異的な免疫を促進するのではなく、むしろ減少させる傾向があることをいくつかの研究が示している。ブタ
suum に感染し、Lacticaseibacillus rhamnosus(LGG)を投与されたブタでは、type-2 関連遺伝子(例えば、type-1)の発現が減少した。
型関連遺伝子(IL13など)の発現が低下していること(93)。
Bifidobacterium animalis subsp. lactis Bb-12 を与えた豚は、A. suum 誘発性好酸球増多と肥満細胞関連遺伝子の発現が減少した。
の発現を低下させた(94)。これと一致するのが
O. dentatumに感染したブタの結腸における2型免疫関連遺伝子の発現上昇も、混合菌処理により抑制される
E. faeciumと2種類のバチルス菌の混合菌で処理すると抑制された(95)。さらに
また、Lactobacillus taiwanensisをマウスに投与すると、H. faluciumに対する免疫の発現が抑制される。
H. polygyrusに対する免疫の発現を抑制した。
polygyrusに対する免疫の発現を抑制し、強いTreg応答を促進することがわかった(96)。このように、プロバイオティクス細菌と免疫系の間の相互作用
と免疫系の相互作用は、多くのモデルで抗蠕虫免疫を損なうようであり、これはおそらく、いくつかのプロバイオティクス細菌について報告されている効果と一致する。
これは、いくつかのプロバイオティクスがアレルギーや喘息の症状を抑制する効果があると報告されていることと、おそらく一致する(97-99)。
喘息の症状を抑制する効果があるとされていることと、おそらく一致している (97-99) 。さらに、これらの結果は、乳酸菌の産生を刺激するプレバイオティクス食品成分が
また、乳酸菌やその関連菌の産生を刺激するプレバイオティクス食品成分も、アレルギーや喘息に対して同様の減衰効果を持つ可能性がある。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規約はWiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は、該当するCreative Commons Licensetype-2の対応に従う。蠕虫の流行地域におけるプロバイオティクス補給に明確な意味を持つ。
る。
4.6 食事性化合物の微生物に依存しない効果
免疫調節における多くの食事成分の作用機序は、しばしばプレバイオティクス効果(すなわち、微生物に依存しない効果)に起因する。
プレバイオティクス効果(すなわち、遺伝子組換え組成および関連代謝産物の産生を変化させる)とされることが多いが、一部の食事性化合物の遺伝子組換え非依存性効果もまた、免疫系に影響を及ぼす。
に依存しない効果も観察されており、このことは、それらが宿主細胞と直接相互作用して
このことは、それらが宿主細胞と直接相互作用して免疫反応に影響を与えることができることを示唆している。例えば、いくつかの複合
例えば、さまざまな食品に含まれる複合糖質ポリマー(チコリやエルサレム・アーティチョークなど)は、TLRまたはアーティチョークと直接相互作用することが示されている。
免疫細胞上のTLRや他の受容体と直接相互作用し、その活性を変化させ、ワクチンや感染症に対する反応を調節することが示されている。
ワクチンやB型肝炎ウイルスなどの病原体の感染に対する反応を調節することが示されている(100-102)。
同様の調節作用が蠕虫感染に対するタイプ2免疫の文脈でも見られるかどうかは今のところ不明である
現在のところ不明である。揮発性の低分子化合物、例えば
シナモンの精油の主成分であるシンナムアルデヒドのような小さな揮発性化合物が、蠕虫の感染に対する粘膜反応に影響を与えることが示されている。
マウスではH. polygyrus、ブタではA. suum)が、蠕虫感染に対する粘膜反応に影響を与えることが示されている。
にもかかわらず、シンナムアルデヒド投与後のGM組成には変化が見られなかった(103, 104)。おそらく
シンナムアルデヒドまたは関連化合物が腸内で急速に吸収されると、Nrf2-を介した抗酸化反応が誘発されるようである。
を介した抗酸化反応が起こり、局所免疫細胞の活性に影響を与える可能性がある。さらに、これらの食餌性化合物の
これらの食物化合物の無毒化生成物(カルボン酸など)は、それ自体、局所免疫細胞の受容体のリガンドとして作用することができる。
免疫細胞上の受容体に対するリガンドとして作用することができる(105)。このように、食物性化合物の宿主代謝は、遺伝子組み換えとは無関係に
は、GMとは無関係に、粘膜免疫応答を制御することができるかもしれない。
免疫反応を制御することができるかもしれない。
宿主の代謝と免疫の間の関係を理解することは、免疫系の栄養学的操作におけるもう一つの道となることが期待される。
栄養学的な免疫操作のもう一つの方法として、特定の免疫反応を制御するための標的介入を可能にすることが期待される。
免疫系を操作するための新たな手段となることが期待される。
13653024, ja, https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022] からダウンロードしたものです。利用規則については、Wiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照してください。OA論文は、該当するクリエイティブ・コモンズ・ライセンスが適用されます5。未解決の質問と今後の展望
食餌の構成が、宿主の蠕虫感染に対する反応を著しく変化させる可能性があることは明らかである。
を大きく変える可能性があることは明らかである。現在の課題は、食餌成分の代謝を、蠕虫の感染を阻止する免疫回路の発達と関連付けることである。
免疫回路の発達と関連付けることである。食餌の相互関係を明らかにするための詳細な実験
食餌成分、GM、そして抗蠕虫免疫に関与する様々な免疫細胞の間の相互関係を明らかにするための詳細な実験が重要である。
の相互関係を明らかにすることが重要である。最近の合成微生物群集の開発手法の進歩により
を開発するための方法論は、近年進歩している。
が、その一助となるであろう(2)。
ILCのような自然免疫細胞は、腸内の栄養状態の変化に非常に敏感であることが発見された。
ILCのような自然免疫細胞が腸内の栄養量の変化に強く反応するという発見は、食事中の炭水化物やタンパク質がどのように小さな代謝産物に分解されるのかを理解する機会を与えてくれる。
2型免疫の発達を制御していることを理解する機会を与えてくれる。このような回路を理解することで
これらの回路を理解することで、機能性食品成分や新薬の開発が促進されるかもしれない。
を標的とした新薬の開発を促進することが期待される。しかし、基礎的な知見を新しい治療法に応用するためには、多くの障害が残されている。
しかし、基礎的な知見を新しい治療薬に応用するためには、多くの障害が残っている。未解決の重要な問題のいくつかは、対照的なタイプの発酵性糖質がどのように
発酵性糖質がどのように異なる腸管区画で代謝されるのか、そしてこのことが蠕虫の発生時に粘膜免疫の表現型にどのように影響するのか。
蠕虫の感染時に粘膜免疫の表現型にどのような影響を与えるか。さらに、アミノ酸や胆汁酸などの代謝物のセンシングも重要である。
胆汁酸やトリプトファン代謝物など、蠕虫感染時のILC2およびILC3の活性に影響を与える可能性のあるアミノ酸や代謝物の感知もまた、蠕虫感染時の
の活性に影響を与える可能性のある胆汁酸やトリプトファン代謝産物などのアミノ酸や代謝産物を感知することも、さらなる研究が必要な分野である。どの腸内細菌が
どの腸内細菌がAhRリガンドなどの2型調節代謝産物の産生に関与しているのか、また、これらの腸内細菌を治療することができるのか、という疑問がある。
これらの腸内細菌を治療的に利用し(例えばプロバイオティクス)、望ましい免疫反応を引き起こすことができるのかどうかについては、依然として不明である。
は不明なままである。GMの組成がどのように免疫反応を制御しているのか、その基本的な理解には大きな進展が見られますが、その一方で
GMの組成がどのように免疫機能を制御しているかについての基本的な理解は進んでいますが、より実用的な動物モデルがあれば、基本的な知識と免疫機能のギャップを埋めることができます。
基本的な知識と疾病予防のための新しい手段との間のギャップを埋めるために、より実用的な動物モデルが必要とされています。ブタの使用。
ブタは、ヒトと非常に類似したGM組成を持ち、類似した食餌を摂取し、また、ヒトと同じような免疫系に感染している。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するCreative Commons Licenseが適用される。ヒトとほぼ同一の蠕虫種(例:Ascaris suum)が、重要な役割を果たす可能性がある。
が重要な役割を担っている可能性があります。
さらに、実験的な蠕虫感染症の利用も、ヒトと蠕虫の交わりに関する基礎的な調査にとって、ますます有用なモデルとなることが証明されるかもしれない。
さらに、実験的蠕虫感染の利用は、食事、GM、2型免疫の交差点に関する基礎研究のモデルとして、ますます有用であることを証明している。
アレルギーや潰瘍性大腸炎のような疾患との明確な関連性がある。食品成分や遺伝子組み換え由来の代謝物の同定
型免疫細胞の制御に関与する食品成分や遺伝子組み換え由来の代謝産物を同定することは
粘膜バリアにおける2型免疫細胞の制御に関与する食品成分や遺伝子組み換え由来の代謝物を同定することは、非常に価値のあることであろう。
結論として、食事が免疫応答をどのように形成するかをより明確に理解することは、難治性の感染症に対するカスタマイズされた解決策につながる可能性がある。
また、食事によって免疫機能のバランスが保たれるような治療法の発見にもつながるだろう。
また、免疫系のさまざまな働きを広く同時に制御することで、バランスのとれた免疫機能を促進するような治療薬を同定する道も開けるかもしれない。
また、免疫系のさまざまな機能を広く同時に制御することで、バランスのとれた免疫機能を促進する治療薬を特定する道も開けます。これはまた、以下のような新しい治療法の開発を加速させるでしょう。
また、アレルギーや喘息、IBD、その他の免疫機能の異常と関連する病態に対する新しい治療法の展開が著しく加速されるでしょう。
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規約はWiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用される。謝辞
財政的支援に感謝します。
ノボ ノルディスク財団 (助成金 0052422) および
独立研究
Fund Denmark (助成金7026-0094B)からの財政的支援に感謝する。図版は
BioRender.comで作成しました。
利益相反に関する声明
著者らは、申告すべき利益相反はない。
データの利用可能性
この原稿にはオリジナルデータは含まれていません。
使用
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用されますFigure Legends
図1. 図1.自然免疫と適応免疫の対照的な細胞プロファイルの概要。
タイプ1応答は、通常、細胞内の細菌やウイルスによって誘導され、タイプ1
自然リンパ球(ILC1)とTヘルパー細胞(Th1)からなり、IFNγなどのシグネチャーサイトカインを産生する。
タイプ3(またはタイプ17)反応は、通常、細胞外の細菌や真菌によって誘導され、以下の細胞で構成される。
一方、タイプ2反応は蠕虫によって誘導され、ILC2細胞とTh2細胞から構成される。
細胞からなる。これらの細胞サブセットは互いに制御し合っている。したがって、タイプ2応答を引き起こす刺激は、通常
2型反応を引き起こす刺激は、通常1型および3型反応を抑制し、その逆もまた同様である。タイプ-1とタイプ-3応答はまた
タイプ-1およびタイプ-3応答もまた、逆調節されることがあり、場合によっては協調して活性化されることがある(破線の棒グラフで示す)。
図2. 食事由来の代謝産物による免疫調節の可能な結果。
さまざまな食事成分(発酵性食物繊維など)は、腸内細菌叢によって分解され、短鎖脂肪酸などの代謝物を産生する。
短鎖脂肪酸(SCFA)やコハク酸などの代謝産物が得られる。腸内細菌叢の組成もまた、食事摂取によって変化する。
胆汁酸やトリプトファンを代謝する細菌の数が変化し、食事摂取によって腸内細菌叢の組成が変化する。
トリプトファンを代謝する細菌の数を変化させます。これらの代謝物は、いずれも腸管上皮細胞上の特定の受容体と結合し、かつ/または、上皮バリアを通過することができる。
上皮関門を通過して免疫細胞と直接相互作用する。これらの代謝物の受容体として、いくつかの例
これらの代謝物の受容体の例を示している。自然免疫系細胞(ILC)の相対的な割合と活性の変化に関連する
自然免疫系細胞(ILC)やTヘルパー細胞(Th細胞)の相対的な割合や活性の変化と、そのサブセット(タイプ-1、タイプ-2、タイプ-3)が相互に関連しあって、免疫系を制御していると考えられる。
2, type-3)の相対的な割合と活性の変化が,腸管蠕虫に対する免疫の発現を制御している可能性がある.
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online Libraryの利用規約(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用される。References
Blander JM, Longman RS, Iliev ID, Sonnenberg GF and Artis D. Regulation of inflammation by microbiota interactions with host.
微生物叢と宿主との相互作用。Nat Immunol 2017; 18: 851-860.
アレクサンダー・M、ターンボーPJ. 食事-微生物-免疫のメカニズムの解体
相互作用。Immunity 2020; 53: 264-276。
Collins N and Belkaid Y. Control of immunity via nutritional interventions. 免疫2022; 55。
210-223.
Zmora N、スエズJとElinav E.あなたが食べるものです:ダイエット、健康、腸内細菌叢。ネイチャー
レビュー消化器病学&肝臓学 2019; 16: 35-56.
ブニャヴァニッチSとベリンMC。食物アレルギーとマイクロバイオーム。現在の理解と
future directions. Journal of Allergy and Clinical Immunology 2019; 144: 1468-1477.
Cortés A, Peachey L, Scotti R, Jenkins TP and Cantacessi C. Helminth-microbiota cross-talk(蠕虫とマイクロバイオータのクロストーク)-A
脊椎動物の消化器官を介した旅。モレキュラー・アンド・バイオケミカル・パラジトロジー
2019; 233: 111222.
7. White EC, Houlden A, Bancroft AJ et al. Manipulation of host and parasite microbiotas(宿主と寄生虫の微生物相の操作)。サバイバル
線虫の慢性感染時の戦略。サイエンス・アドバンス 2018; 4: eaap7399.
8.
9.
10.
11.
12.
13.
Else KJ, Keiser J, Holland CVら、WhipwormとRoundwormの感染症。ネイチャーレビュー
疾患プライマー2020; 6:44。
Charlier J, Thamsborg SM, Bartley DJ 他、「Mind the gaps in research on the control of the gastrointestinal nematodes
畜産反芻動物および豚の消化管内線虫の制御に関する研究におけるギャップに留意すること。トランスバウンダリー・アンド・エマージング
Diseases 2018; 65: 217-234.
Loukas A、Maizels RM、Hotez PJ. ヒトの土壌伝染性蠕虫の陰と陽
の感染症。Int J Parasitol 2021; 51: 1243-1253.
ホテツPJ. トキソカリア症。人新世エポックの無視された感染症。アドバンス・イン
寄生虫学 2020; 109: 879-883.
Flay KJ, Hill FI and Muguiro DH. A Review: 牧草地におけるHaemonchus contortus感染症。
を、貧血の病態と血液学的パラメータの変化を中心に解説した。
血球学的パラメータの変化に着目して。アニマルズ 2022; 12.
Fox NJ、Smith LA、Houdijk JGM、Athanasiadou S、Hutchings MR. ユビキタス寄生虫による
a 33% increase in methane yield from livestock. Int J Parasitol 2018; 48: 1017-1021.
14. Morgan ER, Aziz N-AA, Blanchard A et al. 100 Questions in Livestock Helminthology Research(家畜蠕虫学研究における100の質問).
Trends in Parasitology 2019; 35: 52-71.
15.
Kotze AC、Hunt PW、Skuce Pら、Recent advances in candidate-gene and whole-genome
Anthelmintic resistance markers の発見と薬剤/受容体相互作用の記述のための候補遺伝子および全ゲノムアプローチにおける最近の進歩。
薬剤/受容体相互作用の解明に対する候補遺伝子および全ゲノムアプローチの最近の進歩。寄生虫学のための国際ジャーナル 薬剤と薬剤耐性
2014; 4: 164-184.
16.
17.
18.
19.
20.
21.
Krücken J, Fraundorfer K, Mugisha JC et al.ルワンダの学童における Ascaris lumbricoides に対するアルベンダゾールの効力の低下。
ルワンダの学童におけるアスカリス・ルンバリコイデスに対するアルベンダゾールの効力の低下。インターナショナル・ジャーナル・フォー・パラシトロジー・ドラッグス・アンド
drug resistance 2017; 7: 262-271.
Ploeger HWとEverts RR。消化管に対する驚くべきレベルの駆除薬耐性
オランダの羊の線虫。Vet Parasitol 2018; 262: 11-15.
Herath M、Hosie S、Bornstein JC、Franks AE、Hill-Yardin EL. 消化管の役割
Mucus System in Intestinal Homeostasis(腸のホメオスタシスにおける粘液システム)。Implications for Neurological Disorders(神経疾患への影響). 2020; 10.
Gause WC, Wynn TA and Allen JE. 2型免疫と創傷治癒:進化的
ヘルミンスによる適応免疫の進化的洗練。Nature reviews Immunology 2013; 13: 607-.
614.
コロンボSAPとGrencis RK。土壌伝染性蠕虫に対する免疫:フィールドと実験室モデルからの証拠
と実験室モデルからの証拠。Frontiers in immunology 2020; 11: 1286-1286.
アンソニーRM、ルティツキーLI、アーバンJF、Jr.、スタデッカーMJ、ゴースWC. 蠕虫感染における防御的免疫機構
蠕虫感染における保護的な免疫機構。Nature reviews Immunology 2007; 7: 975-987.
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンス22が適用される。
23.
24.
25.
26.
27.
28.
29.
30.
31.
Sorobetea D、Svensson-Frej M、Grencis R.
の感染症。Mucosal Immunol 2018; 11: 304-315.
消化管内線虫に対する免疫性
池田崇、西田亜希子、山野真、木村郁夫 短鎖脂肪酸受容体と腸内環境
としての腸内細菌叢
治療標的
代謝疾患、免疫疾患、神経疾患における治療標的としての短鎖脂肪酸受容体と腸内細菌叢。
薬理学・治療学 2022; 239: 108273.
Hosseinkhani F, Heinken A, Thiele I et al. The contribution of gut bacterial metabolites in the human immune signal pathway of non-communicable diseases.
非伝染性疾患のヒト免疫シグナル伝達経路。腸内細菌 2021; 13: 1-.
22.
Koh A, De Vadder F, Kovatcheva-Datchary P and Bäckhed F. From dietary fiber to host
生理学。主要な細菌代謝産物としての短鎖脂肪酸。セル 2016; 165: 1332-1345.
Nastasi C, Candela M, Bonefeld CM et al. 短鎖脂肪酸のヒトへの影響
monocyte-derived dendritic cells. サイエンティフィック・リポーツ 2015; 5: 16148.
Seldin MM, Meng Y, Qi H et al. Trimethylamine N-Oxide promotes vascular inflammation(トリメチルアミンN-オキシドは血管の炎症を促進する。
マイトジェン活性化プロテインキナーゼと核因子-κBのシグナル伝達を介する。ジャーナル・オブ・ザ
アメリカ心臓協会 2016; 5.
Hu J, Wang C, Huang X et al. Gut microbiota-mediated secondary bile acids regulate dendritic
TGR5シグナルを介して自己免疫性ぶどう膜炎を減衰させる細胞。セル・レポート 2021; 36: 109726.
Campbell C, McKenney PT, Konstantinovsky D et al. 胆汁酸のバクテリアによる代謝は
末梢性制御性T細胞の生成を促進する。ネイチャー2020; 581: 475-479.
フォリニュB、ズンポポウルーG、デウルフ
Jら、プロバイオティクスにおける樹状細胞の重要な役割。
機能性における樹状細胞の重要な役割。PLoS One 2007; 2: e313.
Niess JH, Brand S, Gu X et al. CX3CR1を介した樹状細胞の腸管内腔へのアクセスおよび細菌クリアランス。
と細菌クリアランス。Science (New York, NY) 2005; 307: 254-258.
32. Mazzini E, Massimiliano L, Penna G and Rescigno M. Oral tolerance can be established via gap junction transfer of fed antigens.
ング抗原のCX3CR1⁺マクロファージからCD103⁺樹状細胞へのギャップジャンクショントランスファーによって確立することができる。
Immunity 2014; 40: 248-261.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
Todorov H, Kollar B, Bayer F et al. α-リノレン酸リッチダイエットはマウス小腸の微生物相に影響を与える。
マウス小腸の微生物相組成と絨毛の形態に影響を与える。Nutrients 2020; 12.
Zou J, Chassaing B, Singh V et al. 食物繊維を介した腸内細菌叢の栄養補給は、腸内細菌叢を回復させることにより、ダイエット誘発性肥満から保護する。
IL-22を介した大腸の健康状態を回復させることにより、食事誘発性肥満から保護する。セルホスト&マイクロビー
2018; 23: 41-53.e44.
Ridaura VK, Faith JJ, Rey FE et al. Gut microbiota from twins discordant for obesity modulate(肥満について不一致の双子の腸内細菌叢が調節する
metabolism in mice. Science (New York, NY) 2013; 341: 1241214.
Turnbaugh PJ, Ley RE, Mahowald MA et al.肥満に関連する腸内細菌叢は、エネルギー収穫の能力が向上している。
エネルギー収穫のための増加した能力。Nature 2006; 444: 1027-1031.
Berbudi A, Surendar J, Ajendra J 他 フィラリア感染または抗原投与は、食餌誘導における耐糖能を改善する。
食事誘発性肥満マウスの耐糖能の改善。ジャーナル・オブ・自然免疫 2016; 8: 601-616.
Brestoff JR, Kim BS, Saenz SA et al. グループ2自然リンパ系細胞は白血球のベイジングを促進し
adipose tissue and limit obesity. ネイチャー2015; 519: 242-246。
Stephenson LS、Pond WG、Nesheim MC、Krook LP、Crompton DW. アスカリス・スウム(Ascaris suum):栄養
15-で実験的に感染させた若いブタの栄養吸収、成長、および腸の病理学。
日齢幼虫に実験的に感染させた若ブタにおける栄養吸収および腸管病理。Exp Parasitol 1980; 49: 15-25.
Sweeny AR, Clerc M, Pontifes PA 他:栄養補給は蠕虫の負担を減らし、自然宿主における薬剤の効果を高める。
Sweeny AR, Clerc M, Pontifes PA, et al.栄養補給は蠕虫の負担を減らし、自然宿主-蠕虫系における薬効を高める。プロシーディングス バイオロジカルサイエンス
2021; 288: 20202722.
スティーブンソンLS. 小児の栄養失調に対するアスカリス・ルンブリコイデスの貢献度。
寄生虫学 1980; 81: 221-233。
Ing R、Su Z、Scott MEおよびKoski KG。T ヘルパー 2 免疫の抑制と線虫の生存期間の延長。
タンパク質栄養失調マウスにおける線虫寄生虫のTヘルパー2免疫の抑制と生存期間の延長。Proc Natl Acad Sci U S A 2000; 97: 7078- 7083.
7083.
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンス43が適用される。
44.
45.
46.
47.
48.
49.
50.
51.
52.
Shi HN、Scott ME、Stevenson MMおよびKoski KG。エネルギー制限と亜鉛の欠乏は
消化管線虫感染時のマウスT細胞および抗原提示細胞の機能を損なう。
エネルギー制限と亜鉛欠乏は、消化管内線虫感染時のマウスT細胞と抗原提示細胞の機能を損なう。J Nutr 1998; 128: 20-27.
Boulay M、Scott ME、Conly SL、Stevenson MMおよびKoski KG。食事性タンパク質および亜鉛の制限
ヘリグモソモイデス・ポリギルス(Nematoda)感染症を独立して修正する。
マウスにおけるHeligmosomoides polygyrus (Nematoda) 感染を独立して修正した。Parasitology 1998; 116 ( Pt 5): 449-462.
Kambara T, McFarlane RG, Abell TJ, McAnulty RW and Sykes AR.(神原利一、McFarlane RG、Abell TJ、McAnulty RW、Sykes AR)。子羊の免疫と抵抗性に及ぼす年齢と食餌タンパク質の影響
を接種した子羊の免疫性と抵抗性に及ぼす年齢および食餌タンパク質の影響。
Int J Parasitol 1993; 23: 471-476.
van Houtert MF, Barger IA, Steel JW, Windon RG and Emery DL. 食事性タンパク質摂取の影響
への反応に対する食餌性タンパク質摂取の影響。獣医学
Parasitol 1995; 56: 163-180.
Shi HN, Scott ME, Stevenson MM and Koski KG. 亜鉛の欠乏は、Heligmosylusの一次感染マウスにおけるT細胞機能を低下させる。
亜鉛の欠乏は、Heligmosomoides polygyrus (Nematoda) の一次感染によるマウスのT細胞機能を低下させる。寄生虫免疫学
1994; 16: 339-350.
Dewitt WBおよびWeinstein PP.精製飼料給餌によるマウスの腸内寄生虫の駆除。
精製飼料を与えることによるマウスの腸内寄生虫の駆除。J Parasitol 1964; 50: 429-434.
Pearce GP. 育成期-仕上期の豚における食物繊維、内部寄生虫および Lawsonia intracellularis バクテリアの相互作用。
Interactions between dietary fib, endo-parasites and Lawsonia intracellularis bacteria in grower-finisher pigs. Veterinary Parasitology 1999; 87: 51-61.
Petkevicius S, Bjørn H, Roepstorff A et al.
実験的に感染させたブタにおける Ascaris suum および Oesophagostomum dentatum の個体数に及ぼす 2 種類の餌の影響。Parasitology
1995; 111: 395-401.
Petkevicius S, Bach Knudsen KE, Murrell KD and Wachmann H. The effect of inulin and sugar beet fib on Oesophagostomum dentatum.
イヌリンとテンサイ繊維が豚のOesophagostomum dentatum感染に及ぼす影響。Parasitology 2003; 127: 61-68.
Valente AH, de Roode M, Ernst Mら: Identification of compounds responsible for the anthelmintic effects of chicles.
チコリ(Cichorium intybus)の駆虫効果に関わる化合物の分子ネットワークとバイオガイデッド分画による同定。
分画。寄生虫学のための国際ジャーナル 薬剤と薬剤耐性 2021; 15: 105-.
114.
53.
54.
Charlier J, Bartley DJ, Sotiraki S et al. Anthelmintic resistance in ruminants: challenges and solutions.
解決策。寄生虫学の進歩 2022; 115: 171-227。
Cortés A, Peachey LE, Jenkins TP, Scotti R and Cantacessi C. Helminths and microbes within vertebrate gut(脊椎動物の腸内の蠕虫と微生物
the vertebrate gut - not all studies are created equal. 寄生虫学 2019; 146: 1371-1378.
55. ウォークST、ブラムAM、ユーイングSA-S、ワインストックJV、ヤングVB. マウスの腸内環境の変化
寄生蠕虫Heligmosomoides polygyrusの感染時のマウス腸内細菌叢の変化(Alteration of mouse gut microbiota during infection with the parasitic helminth Heligmosomoides polygyrus.
Inflammatory Bowel Diseases 2010; 16: 1841-1849.
56.
57.
Li RW, Wu S, Li W ら: Alterations in the porcine colon microbiota induced by the gastrointestinal nematode (ブタの大腸内細菌叢の変化)
により誘発されるブタ大腸微生物叢の変化。Infection and immunity 2012; 80: 2150-2157.
リーSC、タンMS、リムYALら、蠕虫のコロニー形成は、腸内細菌叢の多様性の増加と関連している。
腸内細菌叢の多様性の増加と関連している。PLOS Neglected Tropical Diseases 2014; 8: e2880.
58. Mansfield LS, Gauthier DT, Abner SR et al. Enhancement of disease and pathology by synergy
of Trichuris suis and Campylobacter jejuni in the colon of immunologically naive swine. その
American journal of tropical medicine and hygiene 2003; 68: 70-80.
59.
60.
61.
62.
Zaiss Mario M, Rapin A, Lebon L et al. The intestinal microbiota contributes to the ability of helminths to modulation of allergic inflammity(腸内細菌叢は蠕虫のアレルギー性炎症の調節能力に寄与する)。
helminths to modulate allergic inflammation. Immunity 2015; 43: 998-1010.
Su C, Su L, Li Y et al. Helminth-induced alteration of the gut microbiota exacerbate bacterial
大腸炎を悪化させる。Mucosal Immunology 2018; 11: 144-157.
Ohnmacht C、Park JH、Cording Sら、MUCOSAL IMMUNOLOGY.MUCOSAL IMMUNOGY. 微生物叢が制御するタイプ
2免疫のRORγt⁺ T細胞による制御。Science (New York, NY) 2015; 349: 989-993.
Scott NA, Andrusaite A, Andersen P et al. Antibiotics induce sustained dysregulation of
マクロファージのホメオスタシスを擾乱することによる腸管T細胞免疫。サイエンス トランスレーショナル
medicine 2018; 10.
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用される63。
寄生性線虫は抗菌活性を発揮し、その恩恵を受ける。
benefit From microbiota-driven support for host immune regulation. Front Immunol 2018; 9:
2282.
64. Moyat M, Lebon L, Perdijk O et al. Microbial regulation of intestinal motility provides.
蠕虫感染に対する抵抗性。Mucosal Immunol 2022; 15: 1283-1295.
65.
66.
67.
Petkevičius S, Murrell KD, Bach Knudsen KE et al.短鎖脂肪酸と乳酸の豚のOesophagostomum dentatumの生存に対する影響
を用いた、豚における Oesophagostomum dentatum の生存率に対する短鎖脂肪酸および乳酸の影響。Veterinary Parasitology 2004; 122:
293-301.
Schneider C, O'Leary CE, von Moltke J 他:代謝物トリガーによる房細胞-ILC2回路が小腸のリモデリングを駆動する。
small intestinal remodeling(小腸リモデリング)。セル 2018; 174: 271-284.e214.
Gerbe F, Sidot E, Smyth DJ et al. Intestinal epithelial tuft cells initiate type 2 mucosal immunity(腸管上皮タフト細胞は2型粘膜免疫を開始する。
to helminth parasites. ネイチャー 2016; 529: 226-230.
68. Myhill LJ, Stolzenbach S, Mejer H et al. 発酵性食物繊維は蠕虫の感染を促進し
し、宿主の炎症反応を増悪させる。ジャーナル オブ イムノロジー 2020; 204: 3042-.
3055.
69.
70.
71.
72.
73.
74.
An J, Zhao X, Wang Y et al. Western-style diet impedes colonization and clearance of Citrobacter rodentium.
Citrobacter rodentium. PLoS pathogens 2021; 17: e1009497.
Neumann M, Steimle A, Grant ET et al.特異的病原体除去マウスにおける食物繊維の剥奪は、腸内病原体への感受性を促進する。
腸管粘膜病原体Citrobacter rodentiumに対する感受性を促進する特定病原体フリーマウスにおける食物繊維の欠乏。腸内
Microbes 2021; 13: 1966263.
Leung JM, Budischak SA, Chung The H et al. Rapid environmental effects on gut nematode
rewildedマウスにおける感受性。PLoS biology 2018; 16: e2004108.
Jin X, Liu Y, Wang J et al. β-グルカンをトリガーとするAkkermansia muciniphilaの膨張は、以下のことを促進する。
マウスにおけるTLR2を介した腸内寄生虫の駆除。Carbohydrate polymers 2022; 275: 118719.
Hou Q, Huang J, Xiong X, Guo Y and Zhang B. Role of nutrient-sensing receptor GPRC6A in regulating colonic group 3 innate
大腸グループ3自然リンパ系細胞の制御と炎症性粘膜治癒における栄養感知受容体GPRC6Aの役割。ジャーナル・オブ
Crohn's & colitis 2022; 16: 1293-1305.
Surace L, Doisne JM, Croft CA et al. Dichotomous metabolic networks govern human ILC2
の増殖と機能を支配している。Nat Immunol 2021; 22: 1367-1374.
75. Wang G, Huang S, Wang Y et al. Bridging intestinal immunity and gut microbiota by metabolites(腸管免疫と腸内細菌叢を代謝産物でつなぐ。
代謝産物による腸管免疫と腸内細菌叢の橋渡し。細胞・分子生命科学: cmls 2019; 76: 3917-3937.
76.
77.
78.
79.
80.
81.
82.
83.
84.
Talbot J, Hahn P, Kroehling L et al. 摂食に依存したVIPニューロン-ILC3回路は腸管バリアを制御する。
腸管バリアー。Nature 2020; 579: 575-580.
Teng F, Goc J, Zhou Lら:概日時計は、腸のグループ3自然リンパ球のホメオスタシスに不可欠である。
腸のリンパ系細胞。サイエンス免疫学 2019; 4.
Carvalho Cabral P, Tekade K, Stegeman SK, Olivier M and Cermakian N. The involvement of the cradian clock in regulation.
宿主の概日時計の寄生虫感染に対する免疫応答の調節への関与
哺乳類における寄生虫感染に対する免疫反応の制御における宿主の概日時計の関与。寄生虫免疫学 2022; 44: e12903.
腸内微小環境におけるAHR:バリア機能を守る。
バリア機能を守る。Nature reviews Gastroenterology & hepatology 2021; 18: 559-570.
Li S, Bostick JW, Ye J et al. アリール炭化水素受容体シグナル伝達細胞は、本質的に以下のことを抑制する。
intestinal group 2 innate lymphoid cell function. Immunity 2018; 49: 915-928.e915.
ローガーHMおよびリヒトTR. 健康と病気における微生物トリプトファンカタボライト。ネイチャー
communications 2018; 9: 3294.
Spencer SP, Wilhelm C, Yang Q et al. Adaptation of innate lymphoid cells to a micronutrient(自然免疫系リンパ球の微量栄養素への適応
欠乏は2型バリア免疫を促進する。Science (New York, NY) 2014; 343: 432-437.
Stacy A, Andrade-Oliveira V, McCulloch JA et al. Infection trains the host for microbiota-.
は、病原体に対する抵抗力を強化する。セル2021; 184: 615-627.e617.
Bron PA, Kleerebezem M, Brummer RJ et al.プロバイオティクスは腸管バリア機能に影響を与えることでヒトの病気を調節することができるのか?
腸管バリア機能に影響を与えるか?The British journal of nutrition 2017; 117: 93-107.
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は、該当するクリエイティブ・コモンズ・ライセンス85に準拠する。
86.
87.
Franz CM, Huch M, Abriouel H, Holzapfel W and Gálvez A. Enterococci as probiotics and their implications in food safety(プロバイオティクスとしての腸球菌と食品安全におけるその重要性)。
食品安全における意味合い。International journal of food microbiology 2011; 151: 125-140.
Tawfick MM、Xie H、Zhao C、Shao PおよびFarag MA。
食事中のイヌリン・フルクタン。腸内環境における役割
イヌリン・フルクタン:腸内恒常性、免疫、健康アウトカム、潜在的治療薬における役割。国際ジャーナル・オブ
生物学的高分子 2022; 208: 948-961.
Johnson-Henry KC, Nadjafi M, Avitzur Y et al. Citrobacter rodentium 感染の影響を改善するために、マウスに前処理を施す。
プロバイオティクスの前処理によるマウスにおけるCitrobacter rodentium 感染の影響の改善。The Journal of Infectious
Diseases 2005; 191: 2106-2117.
88. Wang W, Zijlstra RT and Gänzle MG. リモシラクトバチルス・ファーメンタムK9-2およびラクチカゼイバシラス・カゼイバを与えることで
Lacticaseibacillus casei K9-1, or Limosilactobacillus reuteri TMW1.656 Feeding Reduces Pathogen Load in Weanling Pigs.
離乳豚における病原体負荷の低減 Frontiers in Microbiology 2020; 11: 608293.
89.
90.
Kandasamy S, Vlasova AN, Fischer D et al. Escherichia coli Nissle and
ラクトバチルス・ラムノサス株GGのヒトロタウイルスへの結合、感染、B細胞免疫に及ぼす差異効果。
Immunity. ジャーナル・オブ・イミュノロジー 2016; 196: 1780-1789.
Dea-Ayuela MA, Rama-Iñiguez S and Bolás-Fernandez F. Enhanced susceptibility to Trichuris
プロバイオティクスLactobacillus caseiで処理したB10Brマウスのmuris感染に対する感受性の向上。インターナショナル
Immunopharmacology 2008; 8: 28-35.
91. Lactobacillus rhamnosus Ingestion Promotes Innate Host
腸管寄生虫感染症における自然免疫系の防御を促進する。Clinical and Vaccine Immunology 2013; 20: 818.
92.
93.
94.
Nogueira DS, de Oliveira LM, Amorim CCO et al. Eosinophils mediate SIgA production triggered by TLR2 and TLR4 to control Ascaris suum infection in the TLR2, TLR4.
TLR2 と TLR4 によって、マウスのアスカリス・スウム感染を制御する。PLoS pathogens 2021; 17:
e1010067.
Jang S, Lakshman S, Beshah E et al. Flavanol-Rich Cocoa Powder Interacts with Lactobacillus
rhamnosus LGGと相互作用して、寄生性線虫の感染に対する抗体応答を変化させる。
Ascaris suum. ニュートリエンツ 2017; 9: 1113.
Solano-Aguilar G, Shea-Donohue T, Madden KB et al. Bifidobacterium animalis subspecies
lactisは、小腸の局所免疫反応とグルコース取り込みを調節する。
寄生性線虫Ascaris suumに感染した幼齢ブタ。ガット・マイクロベス 2018; 9: 422-.
436.
95. Myhill LJ, Stolzenbach S, Mejer H et al. Parasite-Probiotic Interactions in the Gut(腸内寄生虫とプロバイオティクスの相互作用)。バチルス属菌(Bacillus sp.
とEnterococcus faeciumは、Type-2炎症反応を制御し、腸内細菌叢を修正する。
豚の蠕虫感染時の腸内細菌叢を修正する。フロンティアーズ・イン・イミュノロジー 2022; 12.
96.
97.
98.
Reynolds LA, Smith KA, Filbey KJ et al.腸管におけるコンセンサルと病原体の相互作用。
tract. Gut Microbes 2014; 5: 522-532.
Ciprandi GおよびTosca MA. 喘息の子どもにおけるプロバイオティクス。チルドレン(スイス・バーゼル)
2022; 9.
Drago L, Cioffi L, Giuliano Mら. プライマリーケアにおけるプロバイオティクスによる小児喘息管理(PROPAM)研究
プライマリーケアにおけるプロバイオティクス研究。を用いた無作為化対照二重盲検試験。
リギラクトバチルス・サリバリウスLS01(DSM 22775)とビフィドバクテリウム・ブレーベB632(DSM 24706)の無作為化対照二重盲検試験。
ジャーナル・オブ・イミュノロジー・リサーチ 2022; 2022: 3837418.
99.
Fu W, Chen C, Xie Q et al. Pediococcus acidilactici strain alleviates gluten-induced food allergy and regulating gut microbiot
と腸内細菌叢を制御する。Frontiers in Cellular and Infection Microbiology 2022;
12: 845142.
100. Vogt LM, Elderman ME, Borghuis T et al.イヌリン型フルクタン食物繊維の鎖長依存的な効果
B 型肝炎に対するヒト全身性免疫反応に対するイヌリン型フルクタン食物繊維の鎖長依存性効果。分子栄養学&
food research 2017; 61.
101. Vogt L, Ramasamy U, Meyer D et al. 異なるタイプのβ2→1-フルクタンによる免疫モジュレーション
はtoll-like receptor依存性である。PLoS One 2013; 8: e68367.
102.
Fernández-Lainez C, Akkerman R, Oerlemans MMPら、β(2→6)型フルクタンが炎症反応を抑制する。
Toll様受容体を介して構造依存的に炎症性反応を抑制する。
Carbohydrate polymers 2022; 277: 118893.
13653024, ja, Downloaded from https://onlinelibrary.wiley.com/doi/10.1111/pim.12965 by Cochrane Japan, Wiley Online Library on [29/12/2022]. 利用規定はWiley Online LibraryのTerms and Conditions(https://onlinelibrary.wiley.com/terms-and-conditions)を参照。OA論文は該当するクリエイティブ・コモンズ・ライセンスが適用される103。
Zhu L, Andersen-Civil AIS, Myhill LJ et al. The phytonutrient cinnamaldehyde limits intestinal inflammation and enteric parasite infection.植物栄養素シンナムアルデヒドは、腸の炎症と腸内寄生虫の感染を制限する。The Journal of nutritional biochemistry 2022; 100: 108887.
104. Williams AR、Hansen TVA、Krych Lら、Dietary cinnamaldehyde enhances acquisition of specific antibodies following helminth infection in pigs(ブタの蠕虫感染後の特異的抗体の獲得を強化する)。Vet Immunol Immunopathol 2017; 189: 43-52.
105. Kim CH, Park J and Kim M. Gut microbiota-derived short-chain fatty acids, T Cells, and inflammation. Immune Netw 2014; 14: 277-288.


この記事が気に入ったらサポートをしてみませんか?