鉄欠乏はバクテロイデーテスの代謝様式を変化させ、炎症時の回復力を促進する

2023年6月14日
鉄欠乏はバクテロイデーテスの代謝様式を変化させ、炎症時の回復力を促進する
https://journals.asm.org/doi/10.1128/spectrum.04733-22
著者紹介 ジャニナ・P.Lewis https://orcid.org/0000-0002-0986-6201 jplewis@vcu.edu, Qin GuiAUTHORS INFO & AFFILIATIONS
DOI: https://doi.org/10.1128/spectrum.04733-22
PDF/EPUB
スペクトル
オンラインファースト
ABSTRACT
はじめに
結果
ディスカション
材料と方法
謝辞
補足資料
参考文献
情報・寄稿者
指標と引用
参考文献
図版・メディア
表
シェア
ABSTRACT
細菌は、宿主の栄養免疫に適応するために、低鉄条件下で持続する必要がある。バクテロイデーテスの鉄刺激に関する知見は乏しいため、我々は口腔内細菌(Porphyromonas gingivalisとPrevotella intermedia)と腸内細菌(Bacteroides thataiotaomicron)について、鉄欠乏および鉄過剰の条件に適応する能力について調べた。トランスクリプトミクスと比較ゲノム解析の結果、多くの鉄制御機構がこの動物門内で保存されていることがわかった。低鉄分で発現が増加する遺伝子としては、fldA(フラボドキシン)、hmu(ヘミン取り込みオペロン)、ABCトランスポーターをコードする遺伝子座などが挙げられる。低下した遺伝子は、frd(フェレドキシン)、rbr(ルブレリトリン)、sdh(コハク酸デヒドロゲナーゼ/フマル酸レダクターゼ)、vor(酸化グルタル酸酸化還元酵素/脱水素酵素)、pfor(ピルビン酸:フェレドキシン/フラボドキシン酸化還元酵素)。また、糖質代謝をコードするB. thetaiotaomicronのsusや、ゼノシデロフォア利用をコードするxusABCなど、属特異的なメカニズムも確認された。また、亜硝酸塩還元をコードするnrfAHオペロンを持つ細菌は、培養液中の亜硝酸塩濃度を下げることができるが、B. thetaiotaomicronのみ鉄依存的にオペロンを発現することがわかった。我々の研究とB. thetaiotaomicronの大腸炎研究(W. Zhu, M. G. Winter, L. Spiga, E. R. Hughes et al., Cell Host Microbe 27:376-388, 2020, https://doi.org/10.1016/j.chom.2020.01.010 )で見つかった調節遺伝子が大きく重なっていたことが注目される。それらの共通に制御される遺伝子の多くは、口腔内細菌属でも鉄が制御されていた。本研究は、鉄が細菌の宿主内での持続性を可能にするマスターレギュレーターであることを指摘し、バクテロイデーテスの鉄ホメオスタシスの分子機構をより一般的に調査する道を開くものである。
重要性 バクテロイデーテスは、口腔内や腸内細菌叢に多く存在する重要な嫌気性細菌群である。鉄はほとんどの生物にとって必要な栄養素であるが、この細菌群では鉄レベルの変化に対する適応の分子機構はよく分かっていない。我々は、口腔内細菌叢に属するPorphyromonas gingivalisとPrevotella intermedia、腸内細菌叢に属するBacteroidetes thetaiotaomicronのトランスクリプトーム応答を調べることにより、Bacteroidetesの鉄刺激機構を定義しました。その結果、鉄を制御するオペロンの多くが3属間で共有されていることが明らかになりました。さらに、バイオインフォマティクス解析により、我々のin vitro研究と大腸炎研究から得られたトランスクリプトームデータとの間に有意な重複があることを確認し、我々の研究の生物学的意義が強調されました。バクテロイデーテスの鉄依存性刺激性を明らかにすることは、鉄依存性制御の分子メカニズムを明らかにし、ヒト宿主における嫌気性菌の存続をより良く理解することにつながるであろう。
はじめに
鉄は、微生物-肝臓脂肪軸の重要な構成要素として浮上している(1, 2)。さらに、鉄は、歯周ポケット内の嫌気性病原性細菌の選択への寄与を通じて、歯周炎の発症に関与している(3、4)。上記のような状態は、バクテロイデーテス門に属する細菌の増加と関連している。バクテロイデーテス門は、口腔内(5-7)と腸内(8-11)の両方で最も多く存在する4つの門の1つである。主な特徴は、それらの細菌が微好気性/嫌気性条件下で増殖できることと、エネルギー生成のために鉄ベースの代謝に依存していることである(12)。実際、ピルビン酸からアセチル・コエンザイムA(CoA)への脱炭酸に必要な中心酵素であるピルビン酸フェレドキシン酸化還元酵素(PFOR)(13)には、大気中の酸素濃度下で容易に酸化され失活する鉄硫黄クラスターが酵素の表面に周辺に4個存在している(13)。その細菌群には、鉄に決定的に依存する代謝機能を媒介する他の多くの酵素も含まれている。したがって、鉄レベルが細菌の代謝調節において中心的な役割を果たすことは直感的である。
これまで私たちは、口腔内細菌であるPorphyromonas gingivalisを対象に、鉄のホメオスタシスを決定することに注力してきました。この細菌は、歯周病の発症と進行に関与することが示されている口腔病原体である。私たちは、ヘミンの主要な取り込み遺伝子座hmuを同定し(14)、また、この細菌の鉄刺激性について報告した(15)。さらに、口腔内細菌であるPrevotella intermedia(16)にも研究を広げ、P. gingivalisと同様の遺伝子やタンパク質が鉄量に応じて制御されることを発見した。しかし、P. intermediaの鉄レベルに応じた遺伝子制御の変化を調べるための包括的なアプローチは行われていない。さらに、腸内バクテロイデーテスの鉄依存性トランスクリプトームもまだ決定されていない。これまで、Bacteroides fragilisとBacteroides vulgatusの外膜タンパク質の発現差が調べられたが、鉄依存性の遺伝子発現差を決定する包括的なアプローチは報告されていない(17)。また、口腔内のバクテロイデス類における鉄依存性制御は、腸内細菌の鉄依存性制御と比較して、どのような分子メカニズムがあるのか、まだ明らかにされていない。
ここでは、トランスクリプトームシーケンス(RNAseq)と比較ゲノム解析により、以下の3種のBacteroidetesの鉄刺激性を明らかにした: Bacteroides thetaiotaomicron、P. gingivalis、P. intermediaである。その結果、鉄が豊富な環境と鉄が不足している環境では、適応に大きな重複があることが明らかになった。このことは、研究対象としたすべての細菌で制御される遺伝子のセットが共通していることからも明らかである。また、ユニークなメカニズムも検出され、それは主にB. thetaiotaomicron菌に限定されていた。
結果
P. gingivalisの鉄依存的な刺激。
鉄欠乏条件下で増殖したP. gingivalis W83において、鉄過剰条件下と比較して最も発現量が増加した遺伝子を表1に示す。また、選択した遺伝子のゲノム遺伝子座を図1に示す。合計96個の遺伝子が1.4倍以上アップレギュレートされていた。これらの遺伝子のうち、11のオペロンが同定された。最も高い発現量を示したのは、MATEファミリーの排出トランスポーターをコードするPG0827であった(920倍の発現量)。この遺伝子はシスエンコードされたアンチセンス転写物を持つことが報告されており、small RNAによって制御されている可能性がある (18) 。T9SS type A sorting domain-containing proteinをコードするPG0495は87倍にアップレギュレートされた。また、いくつかのトランスポーターをコードするオペロンがアップレギュレートされ、以下のように含まれた:Hmuヘミン取り込みシステムをコードするPG1551〜PG15516オペロン(アップレギュレート11.4-30. 1倍)、ABCトランスポーターATP結合タンパク質とTetR/AcrRファミリー転写制御因子が続く2成分リポタンパク質/MMPLトランスポーターをコードするPG1175からPG1181(13.4-136.2倍上昇)、PG1019からPG1022 TonB依存トランスポーターシステム(遺伝子によって20.8-214.5倍上昇)。注目すべきは、代謝過程を媒介するタンパク質をコードする遺伝子も制御されていたことである。このクラスで最も高い発現量を示したのは、機能未知のタンパク質(PG1857、48倍)とフラボドキシン(PG1858、133倍)をコードするPG1857〜PG1858オペロンである。また、PG1461からPG1469を持ち、仮説タンパク質や推定イソプレニルシステイン、カルボキシメチル基転移酵素ファミリータンパク質、クラスI SAM依存性メチル基転移酵素をコードする遺伝子座は、遺伝子によって2-から27倍に発現が上昇した。ヘメリトリンドメイン含有タンパク質(RluAファミリー擬似ウリジン合成酵素)や応答制御因子/転写因子をコードするPG1236〜PG1238オペロンなど、鉄と会合するタンパク質をコードする遺伝子は、遺伝子によって2〜7倍に上昇した。鉄(FeoB2、PG1294)およびマンガン分解酵素(FeoB1、PG1043)の輸送をコードする遺伝子や、ペルオキシレドキシンをコードするahpC遺伝子がわずかに上昇した(2.4倍)。最後に、T9SSタイプAソーティングドメイン含有タンパク質をコードする遺伝子、PG0654とPG1374は、それぞれ1.5-と7.3-倍にアップレギュレートされていた。
表1
表1 P. gingivalis W83の鉄欠乏条件下で1.4倍以上発現が上昇した遺伝子a
Fold changebP valuecGene locus_tagOld_locus_tagProduct4.40.336PG_RS10350 Hypothetical protein2.80.241PG_RS10910 DUF1661 domain-containing protein2.50.174PG_RS11125 DUF1661 domain-containing protein1.90.316PG_RS11315 Hypothetical protein1.80.040PG_RS05700 FeoB associated Cys-rich membrane protein1. 60.038PG_RS00305 ハイポhetical protein1.50.441PG_RS10915 DUF1661 domain-containing protein1.50.100PG_RS00455 Hypothetical protein1.40.538PG_RS09100 IS5 family transposase1.60.030PG_RS00020PG0004NAD-dependent deacylase1.60.035PG_RS01345PG0300 Tetratricopeptide repeat protein1.50.50 026PG_RS01395PG_RS01400PG0311, PG0312Glycosyltransferase family 2 protein, DUF4199 domain-containing protein1.40.062PG_RS01535PG0345Hypothetical protein2.85.41712E-08PG_RS01560PG0350Leucine-rich repeat domain-containing protein1.4-1.50.031-0. 081PG_RS01620-30PG0364-66Class I SAM依存性rRNAメチルトランスフェラーゼ、3′-5′エキソヌクレアーゼドメイン含有タンパク質2、DUF5063ドメイン含有タンパク質1.40.168PG_RS01865PG0419DUF2807ドメイン含有タンパク質1. 70.026PG_RS01945PG0437多糖類生合成/輸出ファミリータンパク質1.50.056PG_RS02085PG0469ハイポheticalタンパク質2.30.007PG_RS02165PG0487IS4様要素IS1598家族トランスポゼ87. 10PG_RS02195PG0495T9SS type A sorting domain-containing protein2.40.0005ahpC PG_RS02725PG0618Peroxiredoxin1.50.014PG_RS02890PG0654T9SS type A sorting domain-containing protein2.31.74918E-05PG_RS03020PG0686PAS domain-containing protein2.00.0002PG_RS03105PG0707TonB-dependent receptor1.50. 099PG_RS03365PG0768DUF2891ドメイン含有タンパク質1.40.07-0.078PG_RS03400-05、tsaBPG0777-78電子伝達フラビンタンパク質サブユニットβ/FixAファミリータンパク質、tR NA(アデノシン(37)-N6)-スレオニルカルバモイル基転移酵素複合体の二量化サブユニット1型 TsaB11.00.083PG_RS03595PG0821仮説状タンパク質920.63. 36987E-06PG_RS03625, PG_RS03630PG0827MATE family efflux transporter, hypothetical protein1.80.090PG_RS04015PG0914Hypothetical protein1.40. 042PG_RS04045PG0920ポリプレノールモノホスホマンノース合成酵素1.70.006PG_RS04080PG0928PglZドメイン含有タンパク質1.90.045PG_RS10280PG0943、PG_0943IS3様要素ISPg5ファミリートランスポゼ1。 4-1.70.005-0.101PG_RS04350-60PG0985-87Sigma-70 family RNA polymerase sigma factor, hypothetical protein, DUF4252 domain-containing protein214.5-20.80PG_RS04495-505PG1019-22DUF4876 domain-containing protein, TonB-dependent receptor, hypothetical proteins3.3-1.69.25226E-09-0. 008PG_RS04595-605, feoB1PG1042-44Glycosyltransferase, ferrous iron transport protein B, DtxR family transcriptional regulator1.50.068PG_RS04645PG1055Thiol protease1. 40.057PG_RS04960PG1119NAD(P)H 依存性酸化還元酵素1.50.044PG_RS05005PG1129Adenosylcobalamin-dependent ribonucleoside-diphosphate reductase13.4-136. 20PG_RS05205-235PG1175-81ABCトランスポーターATP結合タンパク質、IS5ファミリートランスポザーゼ、仮説タンパク質、外膜リポタンパク質選別タンパク質、MMPLファミリートランスポーター、TetR/AcrRファミリー転写制御因子6.9-7.30PG_RS05435-40PG1236-38Hemerythrinドメイン含有タンパク質、応答制御転写因子1. 90.002PG_RS05445PG1238RluA family pseudouridine synthase1.50.025PG_RS05650PG1281DUF2027 domain-containing protein2.21.77345E-05feoB2, PG_RS05695PG1294 鉄輸送タンパク質B1. 80.042PG_RS05705PG1296仮説蛋白質1.40.008PG_RS05870、PG_RS05875PG1333、PG1334脂質A脱アシラーゼ、SPFH/バンド7/PHBドメイン蛋白質7. 30PG_RS06055PG1374T9SSタイプAソーティングドメイン含有タンパク質1.50.077PG_RS06085PG1383LysEファミリートランスポーター1.40.042PG_RS0619、PG_RS06200PG1408、PG1409陽イオン輸送体、仮説的タンパク質2. 0–27.30–0.207PG_RS06415–55PG1461–69Hypothetical protein1.60.275PG_RS06510PG1480DUF4141 domain-containing protein11. 4-30.10-1.82188E-13hmuYRSTUV PG_RS06840-65PG1551-56HmuYファミリータンパク質、TonB依存性受容体、コバルトキラターゼサブユニットCobN、仮説的タンパク質、MotA/TolQ/ExbBプロトンチャネルファミリータンパク質、DUF2149ドメイン含有タンパク質1.40.062PG_RS07200PG1635仮説的タンパク質1.40.132PG_RS08040PG1823PorTファミリータンパク質1。 50.034PG_RS08065PG1829AMP結合タンパク質1.8-133.60-0.005PG_RS08150-70、fldAPG1854-585-ホルミルテトラヒドロ葉酸シクロリガーゼ、S41ファミリーペプチド、dCMPデアミ長ファミリータンパク質、DUF2023ファミリータンパク質、フラボドキシン3.73.00015E-07PG_RS08200PG1868アイソプレニルシステインカルボキシルメチル転移酵素ファミリーのタンパク質15. 80PG_RS08205PG1870クラスI SAM依存性メチル基転移酵素1.5-1.60.033-0.065PG_RS08235-40PG1878-79システインtRNAリガーゼ、パタチン様リン脂質酵素タンパク質1. 50.034-0.073deoC, mnmG, PG_RS08805-790PG1995-92 PG1996Nucleotide pyrophosphohydrolase, deoxyribose-phosphate aldolase, d-tyrosyl-tRNA(Tyr)deacylase, MnmG3. 22.13807E-05PG_RS08850PG2006Hypothetical protein2.09.4623E-05PG_RS08855PG2008TonB-dependent receptor1.40.089PG_RS09320PG2102T9SS type A sorting domain-containing protein1.40.128PG_RS09460PG2133FimB/Mfa2 family fimbrial subunit1.40.032ruvX PG_RS09775PG2202Holliday junction resolvase RuvX
a
記載された遺伝子の順序は、旧ローカスタグによるものです。染色体上にまとまって存在する遺伝子(オペロンの可能性があるもの)はグレーで網掛けしてある。株はNC_002950(CDS [コードDNA配列])の下にある。
b
鉄欠乏条件下で増殖した細菌と鉄分豊富な条件下で増殖した細菌の遺伝子発現の比率を示す。
c
4つの独立した生物学的複製で行われた実験からのP値。
図1
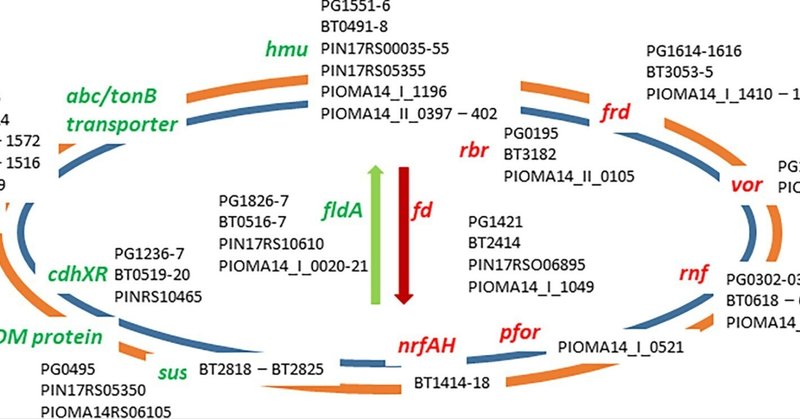
図1 バクテロイデスで発現が増加した遺伝子座の比較。(A) バクテロイデス類におけるヘミン取り込みhmu遺伝子座。P. gingivalis W83(hmuY〜hmuT、PG_RS0)、B. thetaiotaomicron VPI-5482(BT_0491〜PG_RS0598)、P. intermedia 17(PIN17_RS00035〜55)におけるゲノム遺伝子座の比較。(B) バクテロイデーテスのフラボドキシンfld遺伝子座(黄色で指定)。P. gingivalis W83 (PG_RS08170, PG1858), B. thetaiotaomicron VPI-5482 (BT_0517), P. intermedia 17 (fldA) におけるゲノム遺伝子座の比較。(C) バクテロイデーテスの外膜輸送系遺伝子座。ゲノム遺伝子座P. gingivalis W83 (PG_RS04495 to PG_RS04505; PG1019〜21)、B. thetaiotaomicron VPI-5482 (BT_1938 to 9), P. intermedia 17 (PIN17_RS07775 to 90) の比較。(D) バクテロイデーテスのTetR/AcrRレギュレーターと輸送系遺伝子座。ゲノム遺伝子座P. gingivalis W83 (PG_RS05220 to 235; PG1178 to 81), B. thetaiotaomicron VPI-5482 (BT_0507-9), P. intermedia 17 (PIN17_RS07705 to 25, PIN17_RS02240 50, PIN17_RS07725 to 30) を比較した結果.(E) バクテロイデーテスの細胞表面細胞外タンパク質。ゲノム遺伝子座P. gingivalis W83 (PG_RS02195, PG0495)、B. thetaiotaomicron VPI-5482 (オルソログ発見できず)、P. intermedia 17 locus (PIN_RS05350) の比較。
鉄欠乏条件下で生育したP. gingivalis W83で、鉄分充足条件下と比較して最もダウンレギュレーションが多かった遺伝子を表2に示す。また、選択した遺伝子のゲノム上の位置を図2に示す。合計67個の遺伝子が2倍以上ダウンレギュレートされた(Table 2)。最も高度にダウンレギュレートされた遺伝子であるPG0195はルブレリトリンをコードし、46倍ダウンレギュレートされている。その他のダウンレギュレートされた遺伝子は、鉄系代謝タンパク質をコードしていた。まず、電子輸送Rnf複合体をコードするオペロンPG0302〜PG0309である。遺伝子によって異なるが、2-から3.8倍ダウンレギュレートされている。また、フマル酸還元酵素/コハク酸脱水素酵素をコードするオペロンPG1614〜PG1616は、遺伝子によって3.5〜3.7倍にダウンレギュレートされている。また、オキソグルタル酸酸化還元酵素/脱水素酵素系Vorをコードし、4Fe-4S結合タンパク質(フェレドキシン)に挟まれたオペロンPG1809〜PG1813は、遺伝子によって5.2〜7.4倍にダウンレギュレートされました。その他、仮説タンパク質(PG_RS11335、PG_RS11350、PG_RS10005;新しい遺伝子座タグによる命名)をコードする遺伝子が顕著にダウンレギュレートされた(それぞれ30.8-、9.2-、6.3-倍)。これらの遺伝子座には、古い遺伝子座タグはなかった(表2)。
TABLE 2
表2 P. gingivalis W83において、鉄欠乏条件下で2倍以上発現が低下した遺伝子a
Fold changebP valuecGene locus_tagOld_locus_tagProduct-30.80.058PG_RS11335 Hypothetical protein-9.20.226PG_RS11350, PG_RS10690 DUF1661 domain-containing protein-6.30.055PG_RS10005 Hypothetical protein-5.64. 9036E-12PG_RS10650 ハイポhetical protein-3.80.082PG_RS10940, PG_RS11290 DUF1661 domain-containing protein-3.70.491PG_RS11360 ハイポhetical protein-3.10.00203383PG_RS10460, PG_RS06160 DUF1661 domain-containing protein, transposase-3. 00.106PG_RS11285 仮説的タンパク質-2.90.045PG_RS10395 仮説的タンパク質-2.60.227PG_RS11115 DUF1661ドメイン含有タンパク質-2.60.338PG_RS10535 DUF1661ドメイン含有タンパク質-2. 60.0003PG_RS04140 IS5 family transposase-2.40.252PG_RS10550 DUF1661 domain-containing protein-2.30.669PG_RS10870, PG_RS10875 DUF1661 domain-containing protein, hypothetical protein-2. 20.302PG_RS11000 DUF1661ドメイン含有タンパク質-2.10.473PG_RS10580 DUF1661ドメイン含有タンパク質-2.10.005PG_RS02100 DUF4248 domain-containing protein-2.00. 035PG_RS05410 IS5ファミリートランスポザーゼ-2.00.115PG_RS10200 DUF1661ドメイン含有タンパク質-1.90.152PG_RS11050 IS3ファミリートランスポザーゼ-1.90.212PG_RS10250 グリシン切断系Hタンパク質-2.00. 0009PG_RS00510PG0110グリコシルトランスフェラーゼ-2.50.135PG_RS00810PG0177IS4様エレメントIS1598ファミリートランスポザーゼ-46.10PG_RS00900PG0195ルブリトリンファミリープロテイン-2.20. 118PG_RS01030PG0225IS4様エレメントIS1598ファミリートランスポザーゼ-3.8〜-2.38.9495E-13〜0.008PG_RS01350 〜 390PG0302 〜 310RnfABCDGE型電子輸送体-2.00 127PG_RS11275PG0442, PG_0442ハイポテジカル蛋白4. 10.195PG_RS02045PG0460IS5-like element ISPg8 family transposase-3.60.042PG_RS10105PG0524, PG_0524DUF1661 domain-containing protein-2.20. 077PG_RS10115, PG_RS10120PG0545, PG_0546, PG_0546Restriction endonuclease subunit S, helix-turn-helix domain-containing protein-2.10.064PG_RS02500, PG_RS02505PG0563, PG0564Hypothetical protein-4.34.4923E-10PG_RS04540PG1031IS5 family transposase-2. 01.5145E-08~0.0002PG_RS05185~90PG1171~72(Fe-S)-binding protein, lactate utilization protein-2.60.044PG_RS05275PG1197IS5 family transposase-1.90. 0007nrdG PG_RS05545PG1259嫌気性リボヌクレオシド-三リン酸還元酵素活性化タンパク質-2.00.139PG_RS06340PG1445RteCドメイン含有タンパク質-3.14.5733E-06PG_RS06360PG1448IS5ファミリートランスポゼー2.10. 186PG_RS06425PG1463Hypothetical protein−2.00.002PG_RS06610PG1500Helicase−2.10.196PG_RS10525PG1527, PG1527Helix-turn-helix domain-containing protein−2.00.005PG_RS06820PG1545Superoxide dismutase−1.90. 056PG_RS06925PG1573Crp/Fnrファミリー転写制御因子-3.7~-3.55.6732E-14~2.5498E-12sdhB、PG_RS07115~25PG1614~16サクシネート脱水素酵素/フマル酸還元酵素鉄-イオウ、フラビンタンパク質、シトクロムBサブユニット-7. 4~5.10~1.8874E-15PG_RS07975~990PG1809~13Oxidoreductaseファミリータンパク質、FeS結合タンパク質-2.8084PG_RS10710, PG_RS11190PG1980, PG_1980DUF1661 domain-containing protein-3. 00.013PG_RS11390PG2114, PG_2114Hypothetical protein-2.00.184PG_RS09375PG2115DUF1661 domain-containing protein-2.50.008PG_RS09740PG2194IS4-like element IS1598 family transposase
a
記載された遺伝子の順序は、旧ローカスタグによるものです。染色体上にまとまって存在する遺伝子(オペロンの可能性があるもの)は灰色で表示されている。株はNC_002950(CDS [coding DNA sequence])以下。
b
鉄欠乏条件下で増殖した細菌と鉄分豊富な条件下で増殖した細菌の遺伝子発現の比率を示す。
c
4つの独立した生物学的複製で行われた実験からのP値。
図2
図2 バクテロイデスでダウンレギュレーションされた遺伝子座の比較。(A) バクテロイデス属のコハク酸デヒドロゲナーゼ/フマル酸還元酵素のsdh遺伝子座。P. gingivalis W83(PG_RS07115→125、PG1614→16)、B. thetaiotaomicron VPI-5482(BT_3053→55)、P. intermedia 17(PIN17_RS04460 →50)のゲノム遺伝子座の比較。(B) バクテロイデーテスの電子輸送rnf遺伝子座。P. gingivalis W83 (PG_RS01350 to 80, PG0302 to 308) と B. thetaiotaomicron VPI-5482 (BT_0617 to 22) のゲノム遺伝子座を比較したもの。P. intermedia 17ではこの遺伝子座のオルソログは見つからなかった。 (C) バクテロイデーテスのFerredoxin- oxidoreductase (vor) 遺伝子座。P. gingivalis W83 (PG_RS07975 to 90, PG1809013), B. thetaiotaomicron VPI-5482 (BT_0329 to BT_0333), P. intermedia 17 (PIN17_RS08380 to 95) のゲノム遺伝子座を比較した結果。(D) バクテロイデーテスのルブレリトリン(rbr)遺伝子座。P. gingivalis W83 (PG_RS00900, PG0195)、B. thetaiotaomicron VPI-5482 (BT_3182), P. intermedia 17 (PIN17_RS01635) のゲノム遺伝子座を比較した結果。(E) バクテロイデーテスの4Fe-4Sジクラスタードメイン含有タンパク質(frd)遺伝子座(紫矢印)。P. gingivalis W83 (PG_RS10480, PG1421)、B. thetaiotaomicron VPI-5482 (BT_2414), P. intermedia 17 (PIN17_RS06895) のゲノム遺伝子座を比較する。
上記の制御された遺伝子からの結論は、鉄/ヘミンの取り込みと鉄に依存しない代謝機構は劇的にアップレギュレートされ、鉄に依存する代謝と酸化ストレス防御機構は鉄キレート条件下でダウンレギュレートされるということである。これらの結果は、P. gingivalis ATCC33277を使用しても確認された(補足資料の表S1およびS2参照)。特筆すべきは、3つのDUF1661ドメイン含有タンパク質コード遺伝子(9倍、3.7倍、3倍、2倍以上のアップレギュレーション)と13のDUF1661ドメイン含有タンパク質コード遺伝子がダウンレギュレーションされたことである。その多くはP. gingivalisに特異的な低分子タンパク質をコードする遺伝子であった(PG0174 [PG_RS00800]がコードする1つを除く)。
P.intermediaの鉄依存的な刺激性。
鉄を添加せずに培養したP. intermedia OMA14において、鉄を添加した条件下で培養した菌と比較して、最も急激に制御された遺伝子を表3および表4に示す。P. intermedia OMA14で2倍以上発現が上昇した遺伝子は101個あった(表3)。中でも代謝タンパク質PIOMA14_I_0020~PIOMA14_I_0021(fldA、フラボドキシン)とDUF2023(ファミリータンパク質)をコードする遺伝子は、それぞれ159倍、164倍と最も高いレギュレーションを示している。また、TetR/AcrRファミリーの転写制御因子をコードする複数のオペロンが、低鉄条件下で劇的に発現を増加させていた。このように、大型染色体では、TonB依存性受容体、仮説タンパク質、TetR/AcrRファミリー転写制御因子をコードするオペロンPIOMA14_I_0603〜PIOMA14_I_0605(それぞれ129-、289-、268-倍)が著しく発現が上昇した。また、TetR/AcrRファミリー転写制御因子、2つのABCトランスポーターATP結合タンパク質、MMPLファミリートランスポーター、外膜リポタンパク質選別タンパク質、仮説タンパク質、仮説タンパク質をコードするオペロンPIOMA14_I_0908からPIOMA14_I_0914、その部位特異的インテグラーゼは、有意にアップレギュレートした(131から137倍、オペロン内の遺伝子によって違う)。さらに、TetR/AcrRファミリー転写制御因子、ABCトランスポーターATP結合タンパク質、ABCトランスポーターATP結合タンパク質、MMPLファミリートランスポーター、外膜リポタンパク質選別タンパク質、仮説的タンパク質、仮説的タンパク質、および部位特異的インテグラーゼをコードするオペロンPIOMA14_I_1565からPIOMA_I_1572はそれぞれ40-、81-、223-、106-、76-、99-、54倍に遺伝子によりアップレギュレートした。最後に、反対側では、PIOMA14_I_1509からPIOMA14_I_1516の遺伝子座の遺伝子がコードするタンパク質と同様の構成を持つオペロンとして、TetR/AcrRファミリー転写制御因子、ABCトランスポーターATP結合タンパク質があった、 ABC transporter ATP-binding protein, MMPL family transporter, outer membrane lipoprotein-sorting protein, hypothetical protein, hypothetical protein (putative site-specific integrase) が含まれ、それぞれ43-, 96-, 136-, 108-, 48-, 99-, 48倍アップレギュレーションした。小染色体には、TetR/AcrRファミリー転写制御因子、ABCトランスポーターATP結合タンパク質、ABCトランスポーターATP結合タンパク質をコードするオペロンPIOMA14_II_0447からPIOMA14_II_0449があり、それぞれ246倍、102倍、851倍にアップレギュレートされている。
表3
表3 鉄欠乏条件下で増殖したPrevotella intermedia OMA14で2倍以上発現が上昇した遺伝子a
Fold changebP valuecGene locus_tag (genome CDS)Old_locus_tag (genome CDS)dProduct (genome CDS)8.52.42E-07PIOMA14_RS09340 Hypothetical protein159.3-163.70fldA PIOMA14_RS00100-05PIOMA14_I_0020-21 Flavodoxin FldA and DUF2023ファミリータンパク質 3.07. 83E-05PIOMA14_RS00245PIOMA14_I_0051Helix-turn-helix transcriptional regulator2.50.008996–0.018805groL PIOMA14_RS01175-80PIOMA14_I_0239–40Cochaperone GroES2. 20.033994PIOMA14_RS01655PIOMA14_I_0334DnaJ domain-containing protein34.50.030959PIOMA14_RS01765PIOMA14_I_0355IS982 family transposase2. 40.02828PIOMA14_RS01930PIOMA14_I_0382Nucleotide exchange factor GrpE3.20.003992PIOMA14_RS02095PIOMA14_I_0415Hypothetical protein2.7914970.041754PIOMA14_RS02745–50PIOMA14_I_0549–50Hypothetical protein, putative porin128.7–288. 60PIOMA14_RS03025-35PIOMA14_I_0603-5TonB依存性輸送タンパク質およびTetR/AcrR調節因子2.20.038591PIOMA14_RS03345PIOMA14_I_0663DUF4359ドメイン含有タンパク質4.60.029493PIOMA14_RS03625PIOMA14_I_0716HlyDファミリー分泌蛋白2.3〜2.40〜0. 003328PIOMA14_RS03855-70PIOMA14_I_0759-62Cof型HAD-IIBファミリーヒドロラーゼ、パタチン様リン脂質酵素、金属輸送系5.2-33.20-5.3E-10PIOMA14_RS04100-120PIOMA14_I_0804-08TonB依存輸送系64. 10PIOMA14_RS04195PIOMA14_I_0824クラスI SAM依存性メチルトランスフェラーゼ2.10.042773PIOMA14_RS04425PIOMA14_I_0867TlpAファミリータンパク質ジスルフィド還元酵素37.2-131.70-6.5E-09PIOMA14_RS04615-650PIOMA14_I_0908-15TetR・AcrRファミリー転写制御機構とABC輸送システム 12.27. 41E-12PIOMA14_RS04770PIOMA14_I_0939イソプレニルシステインカルボキシルメチルトランスフェラーゼファミリー蛋白質3.90. 033802PIOMA14_RS05150, PIOMA14_RS05155PIOMA14_I_1016, PIOMA14_I_1017Hypothetical protein2.0–2.70.000167–0.029173PIOMA14_RS05715–20PIOMA14_I_1122–23Hypothetical protein8. 5-116.10-8.16E-12hmuY PIOMA14_RS06100-110PIOMA14_I_1196-98HmuYファミリータンパク質、T9SSタンパク質4.44.89E-06PIOMA14_RS06320PIOMA14_I_1238C69ファミリージペプチドーゼ 70. 20PIOMA14_RS06705PIOMA14_I_1314TonB-dependent receptor3.19.03E-05PIOMA14_RS06815PIOMA14_I_1336Hypothetical protein2. 10.021645PIOMA14_RS06835PIOMA14_I_1340Hypothetical protein3.10.006622PIOMA14_RS06860PIOMA14_I_1344Hypothetical protein2.20.031637PIOMA14_RS07025PIOMA14_I_1378DUF3781 domain-containing protein2.50.026707PIOMA14_RS07075PIOMA14_I_1387Acyl carrier protein3. 95.15E-05PIOMA14_RS07460PIOMA14_I_1468重金属輸送P型ATPase42.7-136.40-2.26E-08PIOMA14_RS07670-7705PIOMA14_I_1509-1516TonB dep transpot system and TetR/AcrR transcriptional regulator40.46-222.630-6. 99E-08PIOMA14_RS07920-55PIOMA14_I_1565-72TetR/AcrRファミリー転写制御因子およびABC輸送系2.10.035693PIOMA14_RS08015PIOMA14_I_1586仮説タンパク質2.30.041384PIOMA14_RS09030-35PEOMA14_I_1784-85Sigma-70ファミリーRNAポリメラーゼ・シグマ因子は仮説のタンパク7。 8-11.30-1.11E-15PIOMA14_RS09315-335PIOMA14_I_1836-40ハイポhetical protein, PorV/PorQ protein, S8 family serine peptidase, BACON domain protein9.4-72.40-6.09E-07PIOMA14_RS09520-30PIOMA14_I_1877-79 TonB-dep.receipt, peptidase, 50S protein2.20. 030232PIOMA14_RS09745PIOMA14_I_1919Trypsin-like peptidase domain protein2.00.028127trxA PIOMA14_RS09905PIOMA14_I_1948Thioredoxin2.30.000906PIOMA14_RS10235PIOMA14_II_0010Methylaspartate ammonia-lyase2.2–2.50.004026–0. 012522PIOMA14_RS10240-45PIOMA14_II_0011-12非環状テルペン利用AtuAファミリータンパク質3.60.022164PIOMA14_RS14585PIOMA14_II_0078仮説的タンパク質2.4-8.66.99E-15-0.014783PIOMA14_RS10720-745PIOMA14_II_0099-104YeiHファミリーに帰属する硫酸輸出トランスポーターツーピ. 03463PIOMA14_RS11675PIOMA14_II_0286Hypothetical protein21.6 - 238.60-2.01E-05hmuV - Y, PIOMA14_RS12200PIOMA14_II_0397Hmu TonB依存のヘミン取り込み5.7 - 6.71.62E-09-6.44E-09IOMA14_RS12410-20PIOMA14_II_0430-32TonB transport system102.3 - 850.60-3.66D - 3. 66E-15PIOMA14_RS12490–500PIOMA14_II_0447–49TetR/AcrR regulator and ABC transporter2.2–2.40.045456PIOMA14_RS12590–95PIOMA14_II_0466–7Hypothetical proteins2. 80.023653htpG PIOMA14_RS13475PIOMA14_II_0665Molecular chaperone HtpG2.00.010571PIOMA14_RS13585PIOMA14_II_0685Dipeptidase
a
記載されている遺伝子の順番は、旧ローカスタグによるものです。染色体上にまとまって存在する遺伝子(オペロンの可能性があるもの)はグレーで示した。
b
鉄欠乏と鉄分の多い条件下で培養したバクテリアの遺伝子発現の比率の範囲。
c
4つの独立した生物学的複製で行われた実験から得られた0.1のP値範囲。
d
PIOMA14_Iは大型染色体、PIOMA14_IIは小型染色体を意味する。
表4
表4 P. intermedia OMA14の鉄欠乏条件下でダウンレギュレートされた遺伝子群a
Fold changebP valuecGene locus_tag (genome CDS)Old_locus_tag (genome CDS)dProduct (genome CDS)-2.50.038PIOMA14_RS11990 Hypothetical protein-2.40.003PIOMA14_RS00585PIOMA14_I_0117TonB-dependent receptor-2.2 to -3.40.030 to 0. 056PIOMA14_RS00635から45、PIOMA14_RS00640PIOMA14_I_0128から31仮説タンパク質とN-アセチルムラモイル-L-アラニナミダーゼ-3.40.041PIOMA14_RS01695PIOMA14_I_0342仮説タンパク質-2.30.008PIOMA14_RS02580PIOMA14_I_0518Porin4。 35.425E-06nifJ/pfor PIOMA14_RS02595PIOMA14_I_0521Pyruvate:ferredoxin (flavodoxin) oxidoreductase−2.90.017PIOMA14_RS02650PIOMA14_I_0531Hypothetical protein−2.60.002PIOMA14_RS02655PIOMA14_I_0532Choice-of-anchor J domain-containing protein−5. 30.011PIOMA14_RS02795PIOMA14_I_0557IS982 family transposase-2.10.004PIOMA14_RS02865PIOMA14_I_0571Bifunctional dihydroorotate dehydrogenase B NAD binding subunit/NADPH-dependent glutamate synthase-2. 40.035PIOMA14_RS03010PIOMA14_I_0600ハイポhetical protein-3.0 to -2.0040257PIOMA14_RS04855 to 60PIOMA14_I_0955 to 56YitTファミリータンパク質およびポリン8 90.003PIOMA14_RS05315PIOMA14_I_10494Fe-4S binding protein-2に相当。 40.013PIOMA14_RS05915PIOMA14_I_1159Acyltransferase family protein−3.20.095PIOMA14_RS06410PIOMA14_I_1256Site-specific integrase−3.20.027PIOMA14_RS06655PIOMA14_I_1304GntR family transcriptional regulator−9. 3~8.62.933E-12~1.7193E-11fum1 PIOMA14_RS07200~210PIOMA14_I_1410~12Succinate dehydrogenase/fumarate reductase cytochrome b subunit, flavoprotein subunit, Fe-S subunit-2.60. 001PIOMA14_RS08040PIOMA14_I_1591チョイスオブアンカーJドメイン含有タンパク質-2.10.006PIOMA14_RS08120PIOMA14_I_1606Nucleoside recognition domain-containing protein-4.9 to -2.10.00097PIOMA14_RS08795 to 830PIOMA14_I_1739 to 46TonB dep. RagB/SusDEF外膜タンパク質による輸送系-3.40.0001PIOMA14_RS10155PIOMA14_I_1992Fumarate hydratase-22.30PIOMA14_RS10750PIOMA14_II_0105Rubrerythrin-2.50.001PIOMA14_RS10830PIOMA14_II_0117DUF481 domain-containing protein-2.20. 031PIOMA14_RS11290PIOMA14_II_0201Aspartate 1-decarboxylase−12.50.100PIOMA14_RS11515PIOMA14_II_0247Helix-turn-helix domain-containing protein−3.20. 081PIOMA14_RS11580, PIOMA14_RS11585PIOMA14_II_0265, PIOMA14_II_0266Relaxase/mobilization nuclease domain-containing protein, hypothetical protein-2.9 to -2. 30.007〜0.029PIOMA14_RS12025〜35PIOMA14_II_0361〜63Hypothetical and BACON domain-containing proteins-2.70.026PIOMA14_RS12045PIOMA14_II_0365Hypothetical protein-3.50.0002PIOMA14_RS12290PIOMA14_II_0411Choice of anker J domain-containing protein-2.00. 008PIOMA14_RS12435PIOMA14_II_0435制限酵素サブユニットS-2.8〜-2.30.0002〜0.044PIOMA14_RS13230〜35PIOMA14_II_0622〜23Hypothetical proteins-2. 9から-2.30.010から0.085PIOMA14_RS14000から20PIOMA14_II_0765から69ハイポheticalタンパク質とN-アセチルムラモイル-L-アラニンアミドーゼ
a
記載された遺伝子の順序は、旧ローカスタグによる。染色体上にまとまって存在する遺伝子(オペロンの可能性があるもの)はグレーで示した。
b
鉄欠乏条件下と鉄分豊富な条件下で培養したバクテリアの遺伝子発現の比率を示す。
c
独立した4つの生物学的複製で行われた実験からのP値。
d
PIOMA14_Iは大型染色体、PIOMA14_IIは小型染色体を意味する。
図3
図3 バクテロイデーテスの鉄によって制御される他の遺伝子座の比較。(A) B. thetaiotaomicronの遺伝子座(BT_2063〜2065)をコードするゼノシデロフォアを介した鉄獲得。TonB-外膜受容体をコードするBT_2065遺伝子座は、P. intermediaにオルソログが存在する。(B) バクテロイデーテスのahpCF遺伝子座。P. gingivalis W83(新座標タグ:PG_RS02725〜30、旧座標タグ:PG0618〜19)、B. thetaiotaomicron VPI-5482(BT2811〜12)、P. intermedia 17(AHPC-AHPF)のゲノム遺伝子座を比較したところ。(C)バクテロイデーテスのSAM-メチル基転移酵素コード遺伝子座。P. gingivalis W83 (new locus tag, PG_RS06445; old locus tag, PG1467) と B. thetaiotaomicron VPI-5482 (BT_1855) のゲノム遺伝子座を比較したところ。P. intermedia 17では同様の遺伝子座は確認されなかった。 (D) バクテロイデーテスにおけるnrfAH遺伝子座。P. gingivalis W83(新遺伝子タグ:PG_RS0825〜30、旧遺伝子タグ:PG1820〜21)、B. thetaiotaomicron VPI-5482(BT_1417〜18)、P. intermedia 17のゲノム遺伝子座比較。 E) Hemerythrin-domain containing regulator (P. gingivalisではCdhR). (i) P. gingivalis(新遺伝子タグ:PG_RS05435 - cdhR、旧遺伝子タグ:PG1236〜37)およびB. thetaiotaomicron(BT_0519〜520)に、ヘメリトリンドメインとDNA結合制御因子(CdhR)をコードする同一のオペロンが存在する。(ii) P. intermediaでは、オペロンが融合しているため、1つの制御性PIN17_RS10465タンパク質が生成される。(F) B. thetaiotaomicron(BT_2038〜2040)およびP. gingivalis(新しい遺伝子タグ、PG_RS00295、PG_RS00300、PG_RS00290、古い遺伝子タグ、PG0063〜65)において存在するカチオン排出系。(G) BT_2063から2065まで。B. thetaiotaomicronのXenosiderophore-mediated iron acquisition encoding locus。(H) B. thetaiotaomicronのシトクロムc生合成タンパク質(CcsA)コード化遺伝子座. (I)B.thetaiotaomicronのBT_3630〜BT_3633遺伝子座。(J)B.thetaiotaomicronのBT1834から37の遺伝子座がダウンレギュレーションしている。(K) B. thetaiotaomicronにおいてBT0552から53の遺伝子座がダウンレギュレートされている。(M) B. thetaiotaomicronにおいてBT0543の遺伝子座がダウンレギュレートされた。
2つのヘミン取り込み遺伝子座が有意にアップレギュレートされた。PIOMA14_I_1196(PIOMA14_RS06100)はヘミン取り込み受容体HmuYをコードし、次いでT9SS type A sorting domain-containing protein、ADP-リボシルグリコハイドロラーゼファミリー蛋白質がそれぞれ116-, 104-, 8.5 fpsの発現量に上昇した。小型染色体PIOMA14_II_0397からPIOMA14_II_0402に位置する第2のhmuヘミン取り込み遺伝子座は、HmuYファミリータンパク質、TonB依存性受容体、コバルトケラトースサブユニットCobN、仮説的タンパク質、MotA/TolQ/ExbBプロトンチャネルファミリータンパク質、DUF2149ドメイン含有タンパク質で、それぞれ239-、140-、 127-、115-、60-および22-倍にアップレギュレートされていました。このP. intermedia hmuオペロンは、Lewisら(14)が同定したP. gingivalis hmuオペロンと大きな類似性を持っている。興味深いことに、アップレギュレートされた遺伝子の中には、硫酸トランスポーター、鉄輸送タンパク質(FeoB)、嫌気性リボヌクレオチド-三リン酸還元酵素(NrdDとNrdG)をコードする遺伝子座(PIOMA14_II_0099〜PIOMA14_II_104)もある。
少なくとも2倍以上ダウンレギュレートされたのは49遺伝子であった(表4)。ルブレトリンをコードするPIOMA14_II_0105は、最も大幅にダウンレギュレートされた遺伝子であった(22.3倍)。また、フマル酸還元酵素/コハク酸脱水素酵素系をコードするオペロンPIOMA14_I_1410からPIOMA14_I_1412も大幅にダウンレギュレートした(それぞれ8.5-, 9.2-, 9.3-fold, )。4Fe-4S結合タンパク質(フェレドキシン、PG1421とBT2414に類似)をコードするPIOMA14_I_1049 (PIOMA14_RS05315) も8.9倍のダウンレギュレーションを示した(図2E;補足資料の図S1参照)。主要な鉄依存性代謝酵素であるピルビン酸:フェレドキシン(フラボドキシン)酸化還元酵素、PIOMA14_I_0521は、4.3倍ダウンレギュレーションされた。最後に、ヘリックスターンヘリックスドメイン含有タンパク質をコードするPIOMA14_II_0247は、12.5倍ダウンレギュレートされた。その他、SusD/Eシステム(PIOMA14_I_1739〜PIOMA14_I_1746)、TonB依存システム(PIOMA14_I_0117)、ポリン(PIOMA14_I_0518とPIOMA14_I_0956)、仮説タンパク質などの輸送系をコードする多くの遺伝子が、ダウンレギュレーションされていた(表4)。
以上のことから、P. intermediaでは、鉄の枯渇により、鉄に依存しない代謝機構と鉄の取り込み機構が過剰発現し、鉄に基づく代謝機構と酸化ストレス応答機構がダウンレギュレーションすることがわかった。
B. thetaiotaomicronの鉄依存的な刺激性。
B. thetaiotaomicron VPI BT5482 Δtdkの鉄レベルによって制御される遺伝子を表5および表6に示す。また、選択した遺伝子のゲノム遺伝子座を図1〜図3に示した。 少なくとも2倍以上発現が上昇した遺伝子を323個確認した(表5)。最も急激に制御されたのは、BT2063からBT2065の3つの遺伝子からなる遺伝子座であった(遺伝子によって、それぞれ489.4倍、676.1倍、663倍にアップレギュレート)。この遺伝子座は、TonB依存性の受容体と、DUF4374ドメインとPepSYドメインを持つ2つのタンパク質をそれぞれコードしている(2)。この遺伝子座は、最近、ゼノシデロフォアの利用に関与し、大腸炎モデルで高度に発現が上昇することが示された(2)。なお、大腸炎モデル研究と今回の解析では、同じ菌株を使用しているため、データを直接比較することが可能です。また、オペロンBT0491からBT0498は、Hmu様ヘミン取り込み系をコードし、非常に高い発現量を示した(発現量は遺伝子によって108.9倍から290.8倍の範囲)。TetR/AcrRファミリー転写制御因子と2つのABCトランスポーターATP結合タンパク質をコードするBT0507からBT0509は、それぞれ51.2-、516.2-、675.5-倍にアップレギュレーションされた。高レギュレーションだったのは、fimbrillin-familyタンパク質、2つのシトクロムc生合成タンパク質(CcsA)、ポリン、チオール酸化還元酵素、ペプチダーゼM75、ヘリックス-ターン-ヘリックス転写制御因子、3つの仮説的タンパク質をコードするBT2473〜BT2482遺伝子群で、遺伝子によって69.4〜323.7倍のアップレギュレーションだった。また、BT2479は大腸炎モデルでもアップレギュレートされていたことが注目される(2)。PepSY関連TMヘリックスドメイン、DUF4857ドメイン、フィンブリリン蛋白、TonB依存性受容体、ABCトランスポーターATP結合蛋白、仮説蛋白をコードする遺伝子座BT3625からBT3633は、その遺伝子座内の遺伝子により36.7-128倍のアップレギュレーションが見られた。
TABLE 5
表5 B. thetaiotaomicron BT5482 Δtdkにおいて、鉄欠乏条件下で少なくとも2倍以上発現が増加した遺伝子a
Fold changebP valuecGene locus tagOld_locus_tagProduct2.1550.003BT_RS06005 Helix-turn-helix domain-containing protein2.2690.147BT_RS09290 Hypothetical protein2.5460.199BT_RS09765 Site-specific integrase3. 2260.097BT_RS12005 仮説的タンパク質3.5510.437BT_RS05690 DUF3408ドメイン含有タンパク質3.5510.437BT_RS05715 仮説的タンパク質3.5510.437BT_RS13325 チロシン型再結合酵素/インテグラーゼ3. 5510.437BT_RS20280 ヘリックス-ターン-ヘリックスドメイン含有タンパク質3.5510.437BT_RS12405 ヘリックス-ターン-ヘリックスドメイン含有タンパク質5.9210.25BT_RS13060 潜伏タンパク質5.9680. 248BT_RS05060 AAAファミリーATPase5.9680.248BT_RS05725 ハイポテティカルプロテイン7.0340.053BT_RS12380 ハイポテティカルプロテインメント 8.2910.161BT_RS05905 HUファミリーDNA結合タンパク質17.770.049BT_RS00115 トランスポーザー42.960. 01BT_RS12570、BT_RS12575 Abiファミリータンパク質54.980.007BT_RS24385仮説タンパク質150.90BT_RS02430コバルトケラトースサブユニットCobN2.2070.272BT_RS00045、BT_RS00050BT0010-11仮説タンパク質2. 6140.232BT_RS00060BT0013Hypothetical protein8.2910.161BT_RS00075BT0016RteC domain-containing protein3.5510.437BT_RS00090BT0019Hypothetical protein15.40.062BT_RS00120BT0028Alkaline phosphatase3.2010. 021BT_RS00360BT0076サイト特異的インテグラーゼ5.9210.25BT_RS00370BT0078JABドメイン含有タンパク質3.5510.437BT_RS00485、BT_RS00490BT0100-101ハイポhetical protein, relaxase/mobilization nuclease domain-containing protein3.5510. 5510.437BT_RS00525、BT_RS00530BT0108-09AAAファミリーATPase、ヘリックス-ターン-ヘリックスドメイン蛋白質2.4850.089BT_RS00550BT0113ハイポhetical protein5.1460.071BT_RS00640BT0130SDR family oxidoreductase2. 5060.28BT_RS00860BT0175TIGR03987ファミリータンパク質5.9680.248BT_RS00965BT0196MFSトランスポーター2.6728E-05BT_RS01060BT0215転写抑制因子8.1-3.61E-14-0.437BT_RS01100-1135BT0224-31ハイポテジカルタンパク質およびヘリックスターンヘリックスタンパク質2. 23E-04BT_RS01175BT0238嫌気性硫酸化酵素・成熟タンパク質2.1-2.72E-05-0.001BT_RS01210-240BT0245-51AAAファミリーATPase、メタロフォスフォエステラーゼ、膜タンパク質、Σ-70RNAポリメラーゼ・シグマ因子、ジヒドラターゼ2. 2–2.96E-05–4E-09BT_RS01275–80BT0258–59Glucosamine-6-phosphate deaminase, aminohydrolyse5.9220.25BT_RS01295BT0263Six-hairpin glycosidase3.0890. 14BT_RS01435BT0292YjbHドメイン含有タンパク質2.290.097BT_RS01480BT0302仮説的タンパク質2.1380.336BT_RS01655BT0339アルファキシロシダーゼ2.0550.009Tyrosine型DNA invertase cluster 3b3.BT_RS01830BT0375 1370.02BT_RS01970BT0404DUF3987ドメイン含有タンパク質11.1-290.80-4E-08BT_RS02415-450BT0491-99Hmuヘミン輸送系2.4-675. 50-2E-04BT_RS02470-2505BT0503-09TetR/AcrR family転写制御因子、TonB-dep受容体、ABCトランスポーター2.3160.068BT_RS02520BT0512Hypothetical protein37.1-598.40BT_RS02535-45 (fldA)BT0515-17Quinol oxidase、DUF2023 protein、FldA16. 3-20.00-7E-13BT_RS02555-60BT0519-20Hemerythrin domain-containing protein, response regulator transcription factor2.0155E-04BT_RS02575BT0523Sugar transferase2. 1439E-06BT_RS02580BT0524Response regulator2.1730.079BT_RS02600BT0528Phosphoribosylanthranilate isomerase2.0–3.02E-06–0.104BT_RS02615–25, trpBT0531–33Aminodeoxychorismate/anthranilate synthase compon. II、アントラニル酸合成酵素コンポーネントIタンパク質、トリプトファン合成酵素βサブユニット2.4560.088BT_RS02685BT0548Diaminopimelate epimerase2.1053E-05BT_RS02830BT0577LysM peptidoglycan-binding domain-containing protein27.230.025BT_RS02870BT0585Flavin reductase2.2-3. 35E-07-5E-04BT_RS03105-115、xthBT0629-31Nrampファミリー二価金属輸送体、エキソデオキシリボヌクレアーゼIII、C-GCAxxG-Cファミリータンパク質3.5510.437BT_RS0325BT0663ハイポーティカルプロテイン4.3540. 113lepBBT0667Signal peptidase I4.3540.119BT_RS03315BT0669Efflux RND transporter periplasmic adaptor subunit2.7-3.68E-05-0.437BT_RS03630-35BT0726-27RtcB family protein, hypothetical protein3. 5670.02BT_RS03955BT0791NUDIX hydrolase3.5510.437BT_RS04255BT0851Hypothetical protein2.6270.007BT_RS04400BT0878ATP-binding cassette domain-containing protein2.0–3. 47E-08htpG、BT_RS04515BT0897-98分子シャペロンHtpG2.0-5.08E-05-0.074BT_RS04640BT0922PepSY様ドメイン含有タンパク質、Pasドメインセンサーkinase2. 4433E-04BT_RS05110-115BT1015-16SDRファミリー酸化還元酵素、パタチン様ホスホリパーゼファミリー蛋白質 3.5620.178BT_RS05170BT1028RagB/SusD family栄養摂取外膜蛋白質 2.5-8.30.002-0.279BT_RS05245-55BT1043-45SusD/RagB family栄養結合性外膜蛋白質。 リポタンパク質、エンドグリコシダーゼ、DUF1735ドメインタンパク質3.5510.437BT_RS05365BT1068Hypothetical protein3.5510.437BT_RS05505BT1095Helix-turn-helix domain-containing protein2. 090.03BT_RS05630BT1116DUF296ドメイン含有タンパク質2.0390.049BT_RS05655BT1121Carboxymuconolactone decarboxylaseファミリータンパク質3. 5510.437BT_RS05705-10BT1131ハイポhetical protein3.1-3.52E-10-4E-07BT_RS05740-50BT1138-39Hypothetical protein, site-specific integrase, TonB-dep protein2.1780.005BT_RS05875BT1163TolC family protein2.5150.214BT_RS05925BT1173DUF4373 domain-containing protein2.1-2.30.032-0. 271BT_RS06120-30BT1212-14TolCファミリータンパク質、排出RNDトランスポーター、ABCトランスポーター2.9330.032BT_RS06235BT1232Hypothetical protein2.0170. 087BT_RS06340-45BT1255-56ハイポheticalタンパク質、DUF2089ファミリータンパク質2.660.219BT_RS06370BT1262ヘリックス-ターン-ヘリックス転写制御因子2.6224E-05BT_RS06380BT1264反応制御因子3.5660. 017BT_RS07020BT1385AraCファミリー転写制御因子3.5650.056BT_RS07100BT1401MFSトランスポーター3.5650.09BT_RS07230BT1423ACHドメイン含有タンパク質3.5510. 437BT_RS07225BT1426DUF1062 domain-containing protein1.9850.003trxABT1456Thioredoxin2.0480.067BT_RS07380-85BT1457-59Response regulator transcription factor, histidine kinase2. 0888E-04BT_RS07480BT1477フコースイソメラーゼ2.2-3.05E-06BT_RS07965BT1575-772ヒドロキシ酸脱水素酵素、ピリンファミリー蛋白、ピリドキサミン5´-リン酸オキシダーゼ3. 2410.002BT_RS08050BT1590仮説タンパク質5.9210.25BT_RS08070、feoBBT1592仮説タンパク質、第一鉄輸送タンパク質B2. 95E-08BT_RS08100、BT_RS08105BT1597-98テトラトリコペプチド反復タンパク質、HAMPドメイン含有ヒスチジンキナーゼ2.30. 007BT_RS08130, ccsABT1604-5Cytochrome c biogenesis protein ResB, cytochrome c biogenesis protein CcsA2.30.05BT_RS08205BT1618FecR family protein15.40.062BT_RS08300BT1637Hypothetical protein15. 4-23.90BT_RS08405 (fsa), BT_RS08410, BT_RS0415 (gpmA)BT1658-60Fructose-6-phosphate aldolase, 2, 3-diphosphoglycerate-depen. phosphoglycerate mutase3.60.179-0.437BT_RS08685-95BT1713-15HAD-IIIA family hydrolase, sialic acid synthase, glycosyltransferase family protein2-3.60.338-0. 437aepX, BT_RS08725BT1720-21Phosphoenolpyruvate mutase, phosphocholine cytidylytransferase family protein3.60.437BT_RS09045-50BT1785-86Hypothetical protein, DUF4270 family protein2.2-3.10.249BT_RS090-95BT1794-95Response regulator transcription factor, histidine kinase2.10. 004BT_RS09105BT1797Helix-turn-helix transcriptional regulator2.1-2.33E-04groL - BT_RS09270BT1829-30 シャペロニン GroEL, コシャペロン GroES2.80. 297BT_RS09460BT1864Class I SAM-dependent methyltransferase27.00.039BT_RS09485BT1869Transposase2.20.125BT_RS09520BT1876DUF4974 domain-containing protein4. 4-6.47E-04-0.011BT_RS09595-605BT1890-92UvrD-ヘリカーゼドメイン含有タンパク質、仮説タンパク質、IS3ファミリートランスポーザー2.1-3.17E-04BT_RS09620BT1895-973仮説タンパク質、ロイシンが多い反復タンパク質、LemAファミリータンパク質3.5-7.40-0. 009BT_RS09805-35BT1933-39IS256ファミリートランスポザーゼ、仮説タンパク質、HEATリピートドメイン含有タンパク質、DUF4876ドメイン含有タンパク質、TonB依存性受容体3.60 437BT_RS09850BT1942Hypothetical protein2.10.342BT_RS09855-60BT1944-45 ヘリックス-ターン-ヘリックスドメイン含有タンパク質、ParAファミリータンパク質 2.7-3.60.288-0. 437BT_RS09875–85BT1948–49Hypothetical protein3.60.437BT_RS09950BT1964TetR/AcrR family transcriptional regulator3.60.178BT_RS10035–4040BT1981–82Hypothetical protein2.00. 007rpiABT1986 リボース5リン酸イソメラーゼ A2.00.083BT_RS10245BT2024 仮説タンパク質2.5-3.31E-08-1E-05BT_IRS10325-35BT2038 40Efflux RNDトランスポーター周辺領域アダプター
サブユニット、CusA/CzcAファミリー重金属排出RNDトランスポーター、TolCファミリータンパク質489.4-676.10BT_RS10455-65BT2063-65PepSYドメイン含有タンパク質、DUF4374ドメイン含有タンパク質、TonB依存受容体2. 20.009BT_RS10795-800BT2132-33ハイポheticalタンパク質、LPS集合タンパク質LptD16.50.077BT_RS10825BT2138Transposase2.00.01BT_RS11045BT2182DUF1349ドメインを含むタンパク質3. 60.437BT_RS11055-60BT2184-85RNAポリメラーゼシグマ因子、仮説タンパク質3.60.437BT_RS11235BT2221DUF3244ドメイン含有タンパク質2.10.209BT_RS11255BT2225DsbAファミリータンパク質3. 60.437BT_RS11545BT2283ハイポーティカルプロテイン6.00.248BT_RS11560BT2286DUF3873ドメイン含有タンパク質2.80.297BT_RS11570BT2288コンジュガール転送タンパク質 TraO2. 00.129ltrABT2297グループIIイントロン逆転写酵素/マチュラーゼ3.60.437traG、BT_RS11625BT2298-99TraGファミリー結合性トランスポゾンATPase、DUF4133ドメイン含有タンパク質8.30.161BT_RS11645BT2303ParAファミリータンパク質3.60.437BT_RS11695、BT_RS11700T2312-13仮説タンパク質、RNA指向DNAポリメラーゼ3.5510。 437BT_RS11695、BT_RS11700BT2312、BT_2312 BT2313、BT_2313Hypothetical protein, RNA-directed DNA polymerase6.00.25BT_RS11750BT2323DUF3945 domain-containing protein5.60. 006BT_RS11770-75BT2327-28仮説蛋白質、DUF3800ドメイン含有蛋白質 2.50.323BT_RS11850BT2345CPBP family膜内メタロ蛋白質 5.90.25BT_RS11880, tnpBBT2350-51IS66ファミリー挿入配列仮説蛋白質
タンパク質、IS66ファミリー挿入配列要素付属タンパク質TnpB2.80.003BT_RS11910BT2356ヘリックス・ターンヘリックス転写制御因子3.30.008BT_RS11975BT2370DinBファミリータンパク質2. 00.09BT_RS12065BT2387O-アセチルホモセリンアミノカルボキシプロピルトランスフェラーゼ/システインシンターゼ4.00.097BT_RS12120-25BT2398-99ハイポheticalタンパク質、IS3ファミリートランスポーザーズ3.60. 437BT_RS12255BT2423K(+)-輸送型ATPaseサブユニットC3.00.147kdpABT2425カリウム輸送型ATPaseサブユニットKdpA20.10.041BT_RS12300-05BT2427-28ISS3ファミリートランスポゼ、仮説上のタンパク質3. 55E-04BT_RS12410BT2450Hypothetical protein3.10.159BT_RS12425BT2453Hypothetical protein5.90.25BT_RS12430BT2455Reverse transcriptase3.60.437BT_RS12500BT2468Helix-turn-helix domain-containing protein62.7–323. 70BT_RS12520-65 (ccsA)BT2473-82Fimbrillin family protein, cytochrome c biogenesis protein CcsA, hypothetical proteins, porin, thiol oxidoreductase, peptidase M75, helix-turn-helix transcriptional regulator2.0-2. 83E-05-0.006BT_RS12620-35BT2494-97ABC トランスポーター ATP 結合タンパク質、ABC トランスポーター パーミアーゼ、排出型 RND トランスポーター ペリプラズム アダプター サブユニット 3.22E-04msrBBT2499 ペプチド-メチオニン (R)-S-oxide 還元酵素 MsrB2. 2-9.62E-16-0.03BT_RS12705-10 (cadA)BT2511-12Transcriptional regulator, cadmium-translocating P-type ATPase3.30.024BT_RS12820BT2535Hypothetical protein2.24E-04BT_RS12845BT25402 component sensor histidine kinase29.BT2540T2.C.C.C. 70.034BT_RS12925BT2556Transposase4.40.119BT_RS13010BT2572Two pore domain potassium channel family protein3.20.069BT_RS13030-35BT2575-76Hypothetical protein, site-specific integrase8.30. 161traM, BT_RS13140, traKBT2596, BT_2596 BT2597, BT_2597-98共役トランスポゾンタンパク質TraM、DUF3989ドメインタンパク質、共役トランスポゾンタンパク質 TraK5.90.25BT_RS13320BT2634Tyrosine-type recombinase/integrase3. 60.437BT_RS13360BT2645PRTRC system protein E2.10.342BT_RS13370BT2647Hypothetical protein11.50.007BT_RS13510-15BT2673-74Hypothetical protein, IS3 family transposase2.02E-04BT_RS13630BT2697Hypothetical protein 2.30. 001BT_RS14075BT2778Sigma-70ファミリーRNAポリメラーゼシグマ因子2.20.249BT_RS14160BT2794HlyDファミリー分泌タンパク質2.4-8.06E-05-0.005AHPF-AHPFBT2811、BT_2811アルキルヒドロパーオキサイド還元酵素サブユニットF、パーオキサイドオキシンの18.1-4. 54E-10-0.049BT_RS14290-225BT2818-25TonB 依存型受容体、RagB/SusDファミリー栄養取り込み外膜タンパク質、SusC/RagAファミリーTonB連動外膜タンパク質、BACONドメイン含有タンパク質、キチナーゼ3. 60.437BT_RS14520BT2863Polysaccharide biosynthesis/export family protein3.60. 437BT_RS14585-910BT2876-81グリコシルトランスフェラーゼ、アシルトランスフェラーゼ、リポ多糖生合成タンパク質、CDP-グリセロールグリセロフォストランスフェラーゼファミリーのタンパク質、NAD依存エピメラーゼ/デヒドラターゼファミリーのタンパク質、2-C-メチル-d-エリスリトール4リン酸シチルリルトランスフェラーゼ、3. 60.437BT_RS14645BT2888ウンデカプレニル/デカプレニルリン酸α-N-アセチルグルコサミニル1-リン酸トランスフェラーゼ3. 60.437BT_RS14980BT2954Hypothetical protein2.60.232BT_RS15070–75BT2974–75Hypothetical protein, NAD-dependent deacylase2.60.036BT_RS15160BT2990Tyrosine-type recombinase/integrase3. 10.153BT_RS15535BT3064DUF4248ドメイン含有タンパク質5.90.25BT_RS16085BT3177Sulfatase4.60.021BT_RS16185-90BT3197-98仮説的タンパク質、DUF4373ドメイン含有タンパク質2.90. 027BT_RS17260-65BT3410重金属結合ドメイン含有タンパク質、仮説タンパク質4.20.021BT_RS17295BT3416ISS1様要素ISBthe4ファミリートランスポザーゼ2.30. 103BT_RS17400BT3438Hypothetical protein16.50.076BT_RS17600BT3480Transposase2.50.041BT_RS18045BT3575Hypothetical protein2.60.35BT_RS18235BT3614Oxidoreductase38.1–128. 10BT_RS18290-325BT3625-33DUF4857ドメイン含有タンパク質、仮説タンパク質、ABCトランスポーターATP結合タンパク質、PepSY関連TMヘリックスドメイン含有タンパク質、仮説タンパク質、DUF4876ドメイン含有タンパク質、TonB依存受容体5. 20.084BT_RS18570BT3682DUF4361 domain-containing protein16.50.076BT_RS18690BT3707Transposase3.60.437BT_RS19185–90BT3805Response regulator, hypothetical protein16.50.076BT_RS19290BT3826Transposase2.3–5.04E-07–0. 006BT_RS20020-40BT3968-72エフラックスRNDトランスポーター ペリプラズムアダプターサブユニット、CusA/CzcAファミリー重金属排出RNDトランスポーター、グルタチオンパーオキシダーゼ、NUDIXドメイン含有タンパク質3.60. 437BT_RS20295BT4021部位特異的インテグラーゼ5.90.25BT_RS20335BT4029仮説タンパク質3.60.437BT_RS20550BT4071仮説タンパク質2.10.02BT_RS20730BT4107PTS糖輸送体サブユニットIIC7.50.031BT_RS20790BT4119ペクチン酸リアセ 3.60. 437BT_RS20855BT4132ディスコイジンドメイン含有タンパク質2.00.004BT_RS21325BT4227Mfa1フィンブリリンC末端ドメインタンパク質2.2-3.37E-04BT_RS21360BT4233-36DUF3868ドメイン含有タンパク質、DUF3575ドメイン含有タンパク質、反応制御因子6. 00.248BT_RS21575-80BT4274DUF5030 ドメイン含有タンパク質、仮説タンパク質2.80.297BT_RS21590、tnpBBT4276-77IS66ファミリー挿入配列仮説タンパク質、アクセサリータンパク質TnpB2.70. 034BT_RS21980BT4355RNAポリメラーゼシグマ-70因子2.46E-05BT_RS22275BT4416銅ホメオスタシス蛋白CutC3.60.437BT_RS22400BT4441ロイシンに富む繰り返し蛋白2.40.015BT_RS22590BT4479サイトスペシフィックインテグレ-ゼ2.17E-04BT_RS22655BT4492ハイポシェティック蛋白2. 3-3.60.437BT_RS22685-30BT4498-49ハイポheticalタンパク質、部位特異的インテグラーゼ2.20.008BT_RS22945BT4552応答制御因子2.07E-04BT_RS22965BT4556NfeDファミリータンパク質3.40.002BT_RS23025BT4567ヒスチジンキナーゼ2. 0-7.70.144BT_RS23040-45、enoBT4570-72仮説タンパク質、RNAポリメラーゼシグマ因子、ホスホピルビン酸ヒドラターゼ3.20.097BT_RS23075-80BT4577-78仮説タンパク質、IS3ファミリートランスポーザー2.00. 011BT_RS23250BT4613 ハイポhetical protein2.0-6.04E-04dnaK, BT_RS23265-85BT4615 19分子シャペロンDnaK、ヘリックス-ターン-ヘリックスドメイン含有タンパク質、部位特異的インテグラーゼ、VirEタンパク質、DUF3987ドメイン含有タンパク質2. 1-3.60.099-0.359BT_RS23290-300BT4621-23MobC familyプラスミド動員リラクソーム蛋白質、リラクサース/動員ヌクレアーゼドメイン含有、蛋白質、仮説蛋白質2.70.012BT_RS23395-400BT4643-44RNAポリメラーゼシグマ-70因子、FecR ドメイン含有蛋白質 3.40.152BT_RS23610BT4685RagB/SusD family栄養摂取外側膜蛋白質5. 84E-05BT_RS23755BT4715DNA starvation/stationary phase protection protein2.6-8.30-0.043BT_RS23855-860, pflA, pflBBT4735-38DsbA family protein, hypothetical proteins, N-acetylmuramoyl-L-alanine amidase, Pyruvate Formate lyase-activating protein, formate C-acetyltransferase2. 20.259BT_RS23885BT4740IS110ファミリートランスポザーゼ2.10.343BT_RS23930BT4749N-6DNAメチラーゼ6.00.248BT_RS24005BT4763DUF3408ドメイン含有たんぱく質
a
記載された遺伝子の順序は、旧遺伝子座タグによるものです。染色体上にまとまって存在する遺伝子(オペロンの可能性があるもの)は、グレーで示した。株はNC_004663(CDS)以下。
b
鉄分が豊富な条件下と比較して鉄分が不足した条件下で培養したバクテリアにおける遺伝子発現の比率。
c
4つの独立した生物学的複製で行われた実験からのP値。
TABLE 6
表6 B. thetaiotaomicron BT5482 Δtdkにおいて、鉄欠乏条件下で少なくとも2.5倍ダウンレギュレートされた遺伝子群a
Fold changebP valuecLocus_tagOld_locus_tagProduct-9.30.132BT_RS16240 Hypothetical protein-4.51E-04 -4.20.353BT_RS21305 Hypothetical protein-4.20.353BT_RS24330 Hypothetical protein-4.20.117BT_RS00315 Hypothetical protein-3.20. 14BT_RS22770 Helix-turn-helix転写制御因子-3.10.235BT_RS11535 仮説的タンパク質-3.10.235BT_RS17910 PLDc N末端ドメイン含有タンパク質-2.50.57BT_RS24325 仮説的タンパク質-2.50.57 -2.50.57BT_RS05605 仮説的タンパク質-2. 50.57BT_RS10270 DUF4373ドメイン含有タンパク質-2.50.57BT_RS12485 仮説タンパク質-2.50.57BT_RS05530 トランスポーザー2.50.57BT_RS09845 仮説タンパク質-2.50.57BT_RS13180 DUF4134ドメイン含有タンパク質-2.50.57 -2.50. 57BT_RS12025 アシルトランスフェラーゼ-2.50.57BT_RS04725 仮説蛋白質-2.50.57BT_RS08645 Smalltalk protein-2.50.57BT_RS15170 DUF3853 family protein-2.50.57BT_RS24395 Hypothetical protein-2.50.57BT_RS21300 Hypothetical protein-2.50. 326BT_RS05520 ヘリックス-ターン-ヘリックスドメイン含有タンパク質-2.80.109BT_RS03120 翼状ヘリックス-ターン-ヘリックスドメイン含有タンパク質-8.10.169BT_RS00010BT0002ハイポhetical protein-4.20.353BT_RS00020BT0004Hypothetical protein-5.90. 241BT_RS00105BT0022ヘリックス-ターン-ヘリックスドメイン含有タンパク質-2.50.042BT_RS00190、BT_RS00195BT0044グリコシルトランスフェラーゼファミリー2タンパク質、アシルトランスフェラーゼ-3.10.003BT_RS00270BT0060多糖類生合成/輸出ファミリータンパク質-4.4〜-2.92E-06〜0. 005BT_RS00275〜80BT0061〜62多糖類生合成チロシンオートキナーゼ、仮説タンパク質-3.490.044BT_RS00305BT0067仮説タンパク質-4.2〜-2.50.353〜0.57traNMK、BT_RS00410〜425BT0086〜89共役トランスポゾンプロテイン tranmk-4.20. 352BT_RS00435BT0091、BT_0091DUF4141ドメイン含有タンパク質-2.50.57traG、BT_RS00445~50BT0093~94TraG共役トランスポゾンATPase、DUF4133ドメイン-タンパク質-2. 50.57BT_RS00460BT0096仮説的タンパク質-4.20.353BT_RS00470、BT_RS00475BT0098AAAファミリーATPアーゼ、仮説的タンパク質-4.20. 352BT_RS00500〜05BT0103〜04DUF3945ドメイン含有タンパク質、仮説的タンパク質-4.2〜-2.50.353〜0.57BT_RS00535〜40BT0110〜11RteCドメイン含有タンパク質、仮説的タンパク質-4.10.177BT_RS00560BT0115アーセネート還元酵素 ArsC-5. 45E-09BT_RS00610BT01242Fe-2S鉄硫黄クラスター結合タンパク質-4.20.353BT_RS00645, BT_RS00650BT0131IS4 family transposase-12.8 to -110BT_RS00850 to 55BT0173から74GGtGRTタンパク質、窒素固定タンパク質 NifU-2.70.038BT_RS00935TB0190TonB dependent receptor-12.7 to -2. 50.084~0.57BT_RS01020~25BT0207~08RagB/SusDファミリー栄養取り込み外膜タンパク質・DUF4984ドメインタンパク質-5.9~-2.60.152~0.241BT_RS01040~50BT0211~13BACONドメイン含有タンパク質、S8ファミリーセリンペプチダーゼ、DUF1573ドメインタンパク質-29. 5→-2.60.007→0.071BT_RS01325→355BT0269→75RagB/SusDファミリー栄養取り込み外膜タンパク質、仮説タンパク質、DUF5007ドメイン含有タンパク質、RagB/SusDファミリー栄養取り込み外膜タンパク質、DUF5124ドメイン含有タンパク質-7. 6~-2.60.033~0.18BT_RS01365~75BT0277~79Carbohydrate-binding protein, alkaline phosphatase family protein, hypothetical protein-3.10.227BT_RS01440BT0293YjbH domain-containing protein-2.90.003BT_RS01470BT0299efflux RND transporter permease subunit-3.BG_RT_T0299 5~-2.63E-10~0.002BT_RS01555~60BT0317~18TonB依存性受容体、SusD/RagBファミリー栄養結合外膜リポ蛋白質-3.5~-2.76E-05~0.004BT_RS01700~05BT0348~49α・N・アラビノフラノシダーゼ、グリコシドヒドロラーゼファミリー 127タンパク質-8. 4~-4.13E-09~2E-05BT_RS01755~80BT0360~65ファミリー43グリコシルヒドラーゼ、ragb/susdファミリー栄養取り込み外膜タンパク質、tonb依存性受容体、仮説的タンパク質-4.740. 007BT_RS01790BT0367アラビナンエンド-1,5-α-L-アラビノシダーゼ-2.8~-2.66E-04~0.021BT_RS02145~65BT0438~40α-N-アセチルグルコサミ二ゼ、トンドベクトル、ragb/susd family nutrient uptake outer membr. タンパク質-2.60.021BT_RS02165BT0442DUF4855ドメイン含有タンパク質-6.1~-2.69E-04~ 0.025BT_RS02190~ 205BT0446~ 49DUF5009 domain-containing protein, family 10 glycosylhydrolase, calcineurin-like phosphoesterase C-terminal domain-containing protein, family 10 glycosylhydrolase-7. 1~-2.58E-04~0 0.154BT_RS02215~30BT0451~54RagB/SusD family 栄養取り込み外膜タンパク質、TonB依存性受容体、シュガーポーターファミリーMFSトランスポーター-4.220. 353BT_RS02670BT0545P-IIファミリー窒素調節因子-3.5~-2.50.09~0.326asnB、BT_RS02700~05BT0551~52Asparagine synthase B, glutamate synthase subunit beta-2.960.014BT_RS02775T0566,BT_0566Hypothetical protein-3.4~-2.61E-05 ~ 0. 007rsxC、BT_RS03050BT0618~22電子輸送複合体サブユニットrsxc、rnfabcdge型電子輸送複合体サブD、rnfabcdge型電子輸送複合体サブG、電子輸送複合体サブユニットE、電子輸送複合体サブユニットrsxa~9. 30.132BT_RS03280BT0662O-抗原リガーゼファミリー蛋白質-4.20.353BT_RS03290BT06646-ブレードβプロペラ-6.8~-3.10.002~0. 015BT_RS03390〜400BT0678〜80TolCファミリータンパク質、排出RNDトランスポーターペリプラズムアダプターサブユニット、CusA/CzcAファミリー重金属排出RNDトランスポーター-3.6〜-2.56E-07〜2E-04BT_RS03430、35(hcp)BT0686〜87Uracilキサンティンペルメアーゼ、ヒドロキシルアミン還元酵素-4. 20.353BT_RS03535BT0706N-アセチルムラモイル-L-アラニンアミダーゼ-5.9~-2.96E-04~0.241BT_RS03550~55BT0709~10DIF3987ドメイン含有タンパク質、仮説的タンパク質-3.60. 02BT_RS03775BT0755RagB/SusDファミリー栄養取り込み外膜タンパク質-4.20.117arsAB、BT_RS04010~15BT0802~03ヒ素ポンプ駆動ATPase、ACR3ファミリー亜ヒ酸排出トランスポーター3.00. 08BT_RS04210〜15BT0841〜42Hypothetical protein-5.90.241BT_RS04295BT0857TolC family protein-2.50.135BT_RS04315BT0861ABC transporter permease-3.05E-06BT_RS04345BT0867TonB-dependent receptor-2.83E-05BT_RS04355BT0869Lipocalin family protein-2. 90.05BT_RS04620~25BT0918~19IS3ファミリートランスポザーゼ、仮説的タンパク質-16.1~-2.50.065~0.57BT_RS04690~700BT0933~35仮説的タンパク質-2.50.57BT_RS04705BT0937仮説的タンパク質-3. 10.248BT_RS04730BT0940仮説蛋白質-2.60.018BT_RS04760BT0945仮説蛋白質-4.2~-2.50.353~0.57BT_RS0470~75BT0947~48チロシン型組み換え/インテグラーゼ、DUF3871ファミリー蛋白質-2.50. 57BT_RS04870BT0967DUF3098 domain-containing protein−2.50.57BT_RS04920BT0977Hypothetical protein−7.60.175BT_RS04960BT09859-O-acetylesterase−4.50.02mgtA, BT_RS04975BT0988Magnesium-translocating P-type ATPase−4.20. 057BT_RS05040BT1001ファミリー78グリコシドヒドロラーゼ触媒ドメイン-2.8~-2.50.113~0.57BT_RS05160~65BT1025~26TonB依存性受容体、DUF4957ドメイン含有タンパク質-2. 61E-04~4E-04BT_RS05260~65BT1046~47SusC/RagA family TonB結合外膜タンパク質、SusD/RagB family 栄養結合外膜リポタンパク質-5.40BT_RS05280BT1050Discoidin domain-containing protein-3.10BT_RS05360BT1067Hypothetical protein-2. 70.008BT_RS05390BT1072DUF4251ドメイン含有タンパク質-7.60.175BT_RS05720BT1133Sigma-70ファミリーRNAポリメラーゼシグマ因子-4.003BT_RS05775BT1143YecHファミリータンパク質-4.20.353BT_RS05900BT1168N-アセチルムラモイル-アラニンアミドーゼ-2. 50.26BT_RS06405BT1269EffluxRNDトランスポーターperiplasmic adaptor subunit-5.0~-3.14E-05~0.001BT_RS06435~45BT1275~77Rhamnulokinase, L-rhamnose mutarotase, L-fucose:H+ symporter permease-7.8~-3.14BT0629 021BT_RS06460~85BT1280~85TonB 依存型受容体、SusD/RagBファミリー栄養結合性外膜リポ蛋白、仮説蛋白、DUF1735ドメイン含有蛋白、エンド-β-N-アセチルグルコサミニダーゼ-4. 00.005BT_RS07015BT1384Pyridoxamine 5′-phosphate oxidase family protein-2.50.57BT_RS07035BT1388Multidrug efflux SMR transporter, cytochrome c biogenesis protein ccsa, cytochrome c biogenesis protein resb-36.4 to -30. 40BT_RS07165〜85BT1414〜18アルギン酸輸出ファミリータンパク質、シトクロムc生合成タンパク質ccsa、シトクロムc生合成タンパク質resb、アンモニア形成シトクロムc亜硝酸還元酵素、シトクロムc亜硝酸還元酵素小型サブユニット6. 3〜-5.22E-13〜7E-06BT_RS07335〜340BT1448〜49ビオチン/リポイル結合タンパク質、アセチル-コアカルボキシラーゼビオチンカルボキシラーゼサブユニット5.12E-12BT_RS07345BT1450アセチル-コアカルボキシラーゼサブユニットβ4.20. 353BT_RS07405BT1462仮説タンパク質-2.50.57BT_RS07435BT1468TolCファミリータンパク質-3.10.222~0BT_RS08345~50BT1646~47糖転移酵素ファミリー2タンパク質、仮説タンパク質-2.50~-4.2. 57BT_RS08660→70BT1708→10糖転移酵素、グリコシルトランスフェラーゼ、EpsGファミリータンパク質-2.50.57BT_RS08700→05BT1716→17CDP-グリセロールグリセロフォストランスフェラーゼファミリータンパク質、リポポリサッカライド生合成タンパク質-3. 05E-06BT_RS09005BT1777Hlycoside hydrolase family 95 protein-4.80.095BT_RS09070BT1790Hypothetical protein-3.6 to -3.02E-09 to 3E-09BT_RS09295 to 300 (hydEFG)BT1834 to 374Fe-4S dicluster domain protein, [FeFe] hydrogenase H-クラスタ radical SAM maturase HydE, {FeFe] hydrogenase H-クラスタの maturation GTPase HydF-7. 60.175BT_RS09480BT1868Tetratricopeptide repeat protein−3.02E-07BT_RS09725BT1918Arylsulfatase−4.20.353BT_RS09870BT1947DUF3408 domain-containing protein−4. 20.353BT_RS10275BT2029Hypothetical protein-2.60.002BT_RS10620BT2097Glycoside hydrolase family 127 protein-2.80.013BT_RS10660BT2105GH92 family glycosyl hydrolase-9.30.138BT_RS10670BT2107TonB-dependent receptor-2.308BCT_RESERVICE-2. 9〜-2.62E-06〜1E-04BT_RS10915〜25BT2156〜58糖リン酸イソメラーゼ/エピメラーゼ、DUF1080ドメイン含有タンパク質、Gfo/Idh/MocAファミリー酸化還元酵素5.20.1BT_RS11095BT2193仮説的タンパク質3. 73E-04BT_RS11125BT2199GH92ファミリーグリコシルヒドロラーゼ-3.00.075BT_RS11135BT2201RagB/SusDファミリー栄養摂取外膜タンパク質-3.10.1BT_RS11145BT2203LamGドメイン含有タンパク質-2.60.155BT_RS11365BT2247仮説タンパク質-7. 3〜-3.02E-09〜2E-06BT_RS11405〜10BT2255〜26アシルトランスフェラーゼ、フマル酸ヒドラターゼ-2.50.57BT_RS11555BT2285Hypothetical protein-7.60.175traN, BT_RS11575BT2289Conjugative transposon protein tran-2.50. 57traJ、BT_RS11595〜600BT2293〜94Conjugative transposon protein traj, DUF4141 domain-containing protein-2.50.327traG, BT_RS11625BT2298 〜99TraG family conjugative transposon ATPase, DUF4133 domain-containing protein-2. 50.57BT_RS11670BT2307RteCドメイン含有タンパク質-4.30.343BT_RS11805BT2335サイト特異的インテグラーゼ-2.70.23BT_RS11825BT2339ハイポーティカルプロテイン-13.7~-7. 84E-14〜3E-06BT_RS12090〜105BT2392〜95IPT/TIGドメイン含有タンパク質、TonB依存性受容体、RagB/SusDファミリー栄養摂取外膜タンパク質、DUF4361ドメイン含有タンパク質-2.80.046BT_RS12130BT2400DNA-3-methyladenine glycosylase I-4. 84E-05BT_RS12205BT24144Fe-4S結合タンパク質(FdxA)-7.60.175BT_RS12375BT2442OmpAファミリータンパク質-2.50.57BT_RS12505BT2469チロシン型組み換え酵素/インテグラーゼ-2.50.056BT_RS12805BT2532RagB/SusDファミリーの栄養摂取外膜.
タンパク質-5.43E-04BT_RS12945BT2560TonB依存性受容体-2.50.57BT_RS13100BT2589SAM依存性DNAメチルトランスフェラーゼ-2.50.57BT_RS13105BT2590仮説タンパク質-7.60.175BT_IRS13120ないし25BT2593ないし94DUF3872ドメイン含有蛋白、結合体転送蛋白質 TraO-5. 9から-2.60.346BT_RS13120から25、traNBT2593から95DUF3872ドメイン含有タンパク質、共役転送タンパク質TraO、共役トランスポゾンタンパク質TraN-2.5 BT_RS13155BT2600 -7.6 から -4.20.178 から 0. 353BT_RS13165→75、traGBT2602→03マチュラーゼ、TraGファミリー共役トランスポゾンATPase、DUF4133ドメイン含有タンパク質-31.2~-3.13E-13~0. 18BT_RS13225〜3310、ltrABT2614〜32YWFCYドメイン含有タンパク質、グループIIイントロン逆転写酵素/マチュラーゼ、シグマ54依存Fisファミリー転写制御因子、応答制御因子、グリコシド水解酵素ファミリー97タンパク質、仮説タンパク質、α-グルクロニダーゼ、グリコシド水解酵素ファミリー88タンパク質、DUF4361ドメイン含有タンパク質、RagB/SusDファミリー栄養摂取外膜. タンパク質、SusC/RagAファミリーTonBリンク外膜タンパク質、IPT/TIGドメイン含有タンパク質、応答制御因子、GH92ファミリーグリコシルヒドロラーゼ、エンドヌクレアーゼ/エキソヌクレアーゼ/ホスファターゼファミリータンパク質、グリコシドヒドロラーゼファミリー125タンパク質ー2.50BT_RS13340BT2643DUF4099ドメイン含有タンパク質ー4.20. 352BT_RS13375〜80BT2648〜49Prokaryotic E2 ligase family D protein, PRTRC system ThiF family protein-4.20.353BT_RS13395BT2652Helix-turn-helix domain-containing protein-3. 1~-2.73E-04~0.003BT_RS13455~75BT2664~68基質結合ドメイン含有タンパク質、生体高分子輸送体ExbD、MotA/TolQ/ExbBプロトンチャネル・ファミリータンパク質-6. 4~4.50~4E-08rbsK、BT_RS14220BT2804、BT_2804リボキナーゼ、TonB依存性受容体、RagB/SusDファミリー栄養摂取外膜タンパク質、DUF4969ドメイン含有タンパク質、ヌクレオシドヒドロラーゼ、多剤DMTトランスポーターパーミーズ4. 20.352BT_RS14270BT2814Dihydrodipicolinate合成酵素ファミリータンパク質-110.107BT_RS14485BT2856DUF5013ドメイン含有タンパク質-4 20.353BT_RS14490BT2857DUF4959ドメイン含有タンパク質-3. 50.065BT_RS14495BT2858RagB/SusDファミリー栄養摂取外膜タンパク質-3.10.232BT_RS14600〜05BT2879〜80CDP-グリセロールグリセロフォストランスフェラーゼファミリータンパク質、NAD依存エピメラーゼ/デヒドラーゼファミリープロテイン-3. 10.222BT_RS14615BT2882グリコシルトランスフェラーゼ-4.3〜-2.50.343〜0.347BT_RS14625〜30BT2884〜85WbqCファミリータンパク質、DegT/DnrJ/EryC1/StrSファミリー アミノトランスフェラーゼ-2. 50.57BT_RS14650BT2889ヘリックス-ターン-ヘリックス転写調節因子-4.20.353BT_RS14675BT2895ファミリー43グリコシルヒドロラーゼ-4.20.352BT_RS14700BT2900ファミリー43グリコシルヒドロラーゼ-14.2~-3.10. 002〜0.241BT_RS14710〜30BT2902〜06仮説タンパク質、DUF4959ドメイン含有タンパク質、TonB依存性受容体、グリコシド水解酵素ファミリー99様ドメインタンパク質-4.2〜-350.073〜0. 154BT_RS14800〜810BT2920〜22TonB依存性受容体、グリコシド水分解酵素ファミリー88タンパク質、β-ガラクトシダーゼ-14.3〜-9.80.01〜0.07BT_RS15040〜45BT2968〜69TonB依存性受容体、αガラクトシダーゼ2. 80.043BT_RS15115BT2982仮説蛋白質-2.50.57BT_RS15130~140BT2985~86JABドメイン含有蛋白質、チロシン型リコンビナーゼ/インテグレーゼ、仮説蛋白質-2.50. 326BT_RS15165BT2991仮説蛋白質-4.20.353BT_RS15195BT2995リラクサーゼ/動員ヌクレアーゼドメイン含有蛋白質-8.3~-2.61E-04~0. 179BT_RS15285〜305BT3012〜16TonB依存性受容体、RagB/SusDファミリー栄養摂取外膜タンパク質、DUF4959ドメイン含有タンパク質、ディスコイジンドメイン含有タンパク質、TonB依存性受容体-9.5〜 -3.05E-09 to 0. 011BT_RS15340〜60BT3024〜28TonB依存性受容体、RagB/SusDファミリー栄養摂取外膜タンパク質、グリコシルヒドロラーゼ、仮説的タンパク質、グリコシドヒドロラーゼファミリー130タンパク質-4.9〜-4.09E〜080. 015BT_RS15440〜50BT3045〜47RagB/SusDファミリー栄養取り込み外膜タンパク質、TonB依存性受容体IPT/TIGドメイン含有タンパク質-2.53E-04BT_RS15495BT3055Succinate dehydrogenase/fumarate reductase iron-sulfur subunit-4. 80.087BT_RS15525BT3061DUF3575ドメイン含有タンパク質-7.9~-3.10,001~0.024BT_RS15645~60BT3087~90グリコシド水解酵素ファミリー66タンパク質、SusF/SusEファミリー外膜タンパク質、RagB/SusDファミリー栄養摂取外膜. タンパク質、TonB依存性受容体-3.80.009BT_RS15675~80BT3093~94アリルスルファターゼ、ファミリー43グリコシルヒドロラーゼ-5.9~-4.89E-07~0. 241BT_RS15725→35BT3103→05TonB依存性受容体、仮説タンパク質、BACONドメイン含有タンパク質-2.6→-2.50372→057BT_RS15880→90BT3135→37部位特異的インテグラーゼ、部位特異的インテグラーゼ、仮説タンパク質-7. 10.035BT_RS15985BT3157RagB/SusDファミリー栄養摂取外膜タンパク質-3.70.024BT_RS16035BT3167Helix-hairpin-helix domain-containing protein-2.50.24BT_RS16070BT3174TonB-dependent receptor-3. 03E-06BT_RS16110BT3182Rubrerythrinファミリータンパク質-2.90.244BT_RS16215〜20BT3202〜03RHSリピート関連コアドメイン含有タンパク質、仮説的タンパク質-14.6〜-10.002〜0. 084BT_RS16305~15BT3221~23DUF3244 ドメイン含有タンパク質、DUF4848 ドメイン含有タンパク質、仮説タンパク質-3.70.02BT_RS16670BT3291Hypothetical protein-3.20.028BT_RS16695BT3296RagB/SusD family nutrient uptake outer memb. タンパク質-2.60.001BT_RS16820BT3322レドキシンファミリータンパク質-2.90.005BT_RS16855BT3329DUF4988ドメイン含有タンパク質-11.1〜-5.30〜4E-07BT_RS16930〜45BT3344〜47DUF4973ドメイン含有タンパク質・RagB/SusDファミリー栄養素摂取外膜(Nature Upter Outumn Member)? タンパク質、TonB依存性受容体、IPT/TIGドメイン含有タンパク質-19.40.044BT_RS17260~265BT3410重金属結合ドメイン含有タンパク質、仮説的タンパク質-3. 13E-06BT_RS17300BT3418チオール:ジスルフィド交換タンパク質-2.70.139BT_RS17405BT3439仮説タンパク質-3.0〜-2.70.004〜0.065BT_RS17570〜80BT3474〜76RagB/SusDファミリー栄養摂取外膜(NTM).タンパク質、TonB依存性受容体、IPT/TIGドメイン含有タンパク質-2.530.57BT_RS17605BT3481DUF1793ドメイン含有タンパク質-19.40.045BT_RS17625BT3485DUF4973ドメイン含有タンパク質-7.60. 175BT_RS17685〜90BT3498〜99DUF4956ドメイン含有タンパク質、DUF2490ドメイン含有タンパク質-5.30.002BT_RS17715BT3505TonB依存受容体-4.20.353BT_RS17740BT3510ハイポhetical protein-3.20. 001BT_RS18035BT3572ハイポhetical protein-2.70.086BT_RS24405BT3582Hypothetical protein-11.00.104BT_RS18130BT3593SGNH/GDSL hydrolase family protein-3.70.01BT_RS18170BT3601SIS domain-containing protein-4.20. 353BT_RS18420BT3651DUF3137ドメイン含有タンパク質-3.25E-04BT_RS18450BT3657糖質結合ドメイン含有タンパク質-3.10.222BT_RS18475、 BT_RS18480BT3663Hypothetical protein, glycoside hydrolase 43 family protein-2.970.019BT_RS18560BT3680, BT_3680TonB-dependent receptor-4.2 to -2.1. 91E-09〜5E-05BT_RS18645〜70、susDEFBT3698〜703α-アミラーゼSusG、DUF5115ドメイン含有タンパク質、SusF/SusEファミリー外膜タンパク質、デンプン結合性外膜リポタンパク質SusD、TonB依存性受容体、グリコシド ヒドロラーゼ ファミリー 97タンパク質-2. 530.57BT_RS18900BT3747、BT_3747Hypothetical protein-3.146E-05BT_RS18915BT3750, BT_3750SusC/RagA family TonB-linked outer memb. protein-6.4 to -3.49E-06 to 4E-05BT_RS18920 to 30BT3752から54SusD/RagB family nutrient-binding outer memb. リポタンパク質、エンド-β-N-アセチルグルコサミニダーゼ、DUF1735ドメイン含有タンパク質-4.6~-2.86E-08~0.007BT_RS19040~55BT3775~78仮説タンパク質-12.6~-3.42E-08~9E-08BT_RS19065~75BT3780~82仮説タンパク質、グリコシド ヒドロラーゼ ファミリー 125タンパク質、グリコシド ヒドロラーゼ ファミリー 88タンパク質-22.3~-2. 90〜2E-08BT_RS19085〜120BT3784〜92GH92ファミリーグリコシルヒドロラーゼ、ハイブリッドセンサー ヒスチジンキナーゼ/レスポンスレギュレーター転写因子、IPT/TIGドメイン含有タンパク質、TonB依存性受容体、RagB/SusDファミリー栄養摂取外膜タンパク質、DUF4361ドメイン含有タンパク質、ラムGドメイン含有プロテアー、グリコシルヒドロラーゼファミリー76タンパク質、-5. 3~-3.52E-06~0.013BT_RS19140~55BT3796~99Sulfatases、α-galactosidase、α-L-fucosidase-2.70.007BT_RS19275BT3823Ferritin-6.4~ -2. 84E-11〜2E-05BT_RS19460〜80BT3854〜58SusC/RagAファミリーTonB結合外膜タンパク質、RagB/SusDファミリー栄養摂取外膜タンパク質、DUF3823ドメイン含有タンパク質、仮説的タンパク質、GH92ファミリーグリコシルヒドロラーゼ-2. 75E-04BT_RS19495BT3861DUF4972ドメイン含有タンパク質-3.1~-2.77E-06~2E-04BT_RS19695~705BT3902~04ABCトランスポーター透過酵素、HlyDファミリー分泌タンパク質-2.7~-2.60.038~0. 106BT_RS19750→55BT3913→14DUF5034ドメイン含有タンパク質、仮説タンパク質-3.0→-2.85E-06→0.099BT_RS19935→55BT3951→53ヘリックス-ターンヘリックスドメイン含有タンパク質、TonB依存受容体、RagB/SusDファミリー栄養摂取外膜タンパク質-4. 80.037BT_RS19955BT3955仮説蛋白質-3.6~-2.64E-10~7E-05BT_RS19970~75BT3958~59TonB依存性受容体、RagB/SusDファミリーの栄養取り込み外膜蛋白質-3. 22E-07BT_RS19985BT3961DUF4302 domain-containing protein−2.70.003BT_RS20110BT3985Hypothetical protein−2.60.043BT_RS20125BT3988DUF4989 domain-containing protein−2.50. 57BT_RS20270BT4016仮説蛋白質-4.20.353BT_RS20305〜310BT4023〜24チロシン型リコンビナーゼ/インテグラーゼ、仮説蛋白質-2.50.57BT_RS20325〜330BT4027〜28仮説蛋白質-5.3〜 -3. 38E-12~3E-04BT_RS20380~90BT4038~40RagB/SusD family栄養取り込み外膜タンパク質、TonB依存性受容体、仮説タンパク質-6.4~-3.49E-06 ~0. 011T_RS20585~610BT4078~83仮説タンパク質、IPT/TIGドメイン含有タンパク質、RagB/SusDファミリー栄養取り込み外膜タンパク質、DUF4361ドメイン含有タンパク質-2.50.57BT_RS20625BT4086DUF5004ドメイン含有タンパク質-14. 3~-3.41E-04~0.353BT_RS20740~65BT4109~14Pectinesterase, pectin esterase, DUF5123 domain-containing protein, TonB-dependent receptor-7.5~-3.50.007~0.105BT_RS20950~55BT4152~53Beta-galactosidase, exopolygalacturonase-6. 3→-3.34E-06→0.005BT_RS21010→35BT4164→69SusC/RagAファミリー TonBリンク型外膜タンパク質、DUF5108ドメイン含有タンパク質、仮説タンパク質、TonB依存性受容体-3. 1〜-2.50.074〜0.23BT_RS21070〜75、rhaMBT4176〜77グリコシドヒドロラーゼファミリー88タンパク質、L-ラムノースミュータロターゼ-3.20.047BT_RS21090BT4180シアル酸O-アセチルエステラーゼ-3.20. 018BT_RS21195〜200、miaABT4202〜03仮説タンパク質、tRNA(アデノシン(37)-N6)-ジメチルアリルトランスフェラーゼMiaA-2.50.221BT_RS21545BT4268RagB/SusDファミリー栄養摂取外膜タンパク質-17. 0〜8.21E-14〜-0.103BT_RS21680〜705BT4294〜99仮説タンパク質、ディスコイジンドメイン含有タンパク質、DUF4361ドメイン含有タンパク質、RagB/SusDファミリー栄養取り込み外膜タンパク質、TonB依存受容体、仮説タンパク質-4. 20.015BT_RS21995BT4358RagB/SusDファミリー栄養取り込み外膜タンパク質-2.50.57BT_RS22120BT4384翼状ヘリックス-ターンヘリックスドメイン含有タンパク質-2. 90.071BT_RS22160BT4392IS4様エレメントISBthe3ファミリートランスポザーゼ-2.70.013BT_RS22225~30BT4406~07仮説タンパク質、DUF1735ドメインを含むタンパク質-2.50. 57BT_RS22300→05BT4421→22仮説タンパク質-2.50.57BT_RS22385BT4438仮説タンパク質-3.2→-2.50.011→0.147BT_RS22435→40BT4448→49Gfo/Idh/MocAファミリー酸化還元酵素-3.1→-2.80.001→0. 007BT_RS22495〜505BT4460〜62カルボキシペプチダーゼ様調節ドメイン含有タンパク質、シグマ-70ファミリーRNAポリメラーゼシグマ因子、1-アシル-sn-グリセロール-3-リン酸アシルトランスフェラーゼ-2・50. 57BT_RS22810〜15BT4523〜24制限酵素サブユニットS、ヘリックス-ターン-ヘリックスドメイン含有タンパク質-3.10.017BT_RS22820BT4526メタロフォスポエステラーゼ-3.00.028BT_RS23090BT4581、BT4581グリコシド ヒドロラーゼ ファミリー97タンパク質-4.20. 352BT_RS23275〜80BT4618〜19Helix-turn-helix domain-containing protein、VirE protein-4.20.353BT_RS23290 〜95BT4621〜22MobC family plasmid mobilization relaxosome protein、laxase/mobilization nuclease domain-containing protein-3. 8~-2.97E-06~0.004BT_RS23480~90BT4660~62TonB依存性受容体、DUF4958ファミリータンパク質、ヘパリン硫酸リアーゼ-6.60.005BT_RS23640BT4693Efflux RNDトランスポーターペリプラズム・アダプター・サブユニット8. 4〜-4.00.001〜0.002BT_RS23645〜50BT4694〜95CusA/CzcAファミリー重金属排出RNDトランスポーター、TolCファミリータンパク質-4.80.098BT_RS23720BT4708SusD/RagBファミリー栄養結合外膜? リポタンパク質-2.50.349BT_RS23725BT4709グリコシルヒドロラーゼ-6.9~-2.92E-05~0.152BT_RS23800~805BT4724~25TonB依存性受容体とRagB/SusD外膜タンパク質-2.60. 053BT_RS23815BT4727Glycerophosphodiester phosphodiesterase family protein−2.50.57BT_RS23915BT4746DNA-binding protein−4.80.097BT_RS23945BT4752DUF3945 domain-containing protein−7.60.175BT_RS24010BT4764Hypothetical protein−4. 2から-2.50.326から0.353BT_RS24065から75、traKBT4776から78共役トランスポゾンタンパク質TraK、仮説的タンパク質、共役トランスファータンパク質-2.50.57traN、 BT_RS24085BT4780共役トランスポゾンタンパク質 TraN
a
記載された遺伝子の順序は、旧遺伝子座タグによるものです。染色体上にまとまって存在する遺伝子(オペロンの可能性があるもの)は灰色で表示されている。株はNC_004663(CDS)以下。
b
鉄分が豊富な条件下と比較して鉄分が不足した条件下で培養したバクテリアにおける遺伝子発現の比率。
c
独立した4つの生物学的複製で行われた実験からのP値。
ヘメリトリンドメイン含有タンパク質と応答制御因子/転写因子をコードする2つの遺伝子、すなわちBT0519とBT0520は、それぞれ16.3倍と20倍に発現が増加した。BT1658〜BT1660からなる糖質代謝をコードするオペロンは、それぞれ15.4倍、16.8倍、20倍に発現が上昇した。また、SusD/SusCシステムを含む遺伝子座をコードするBT2818~BT2825は、遺伝子によって4.5~22.5倍アップレギュレートされた。
アルカリホスファターゼをコードする遺伝子BT0028などの酸化ストレス保護機構は、15.4倍アップレギュレートされた。アルキルヒドロペルオキシドレダクターゼサブユニットFをコードするBT2811も8倍アップレギュレートされました。最後に、DNA飢餓/定常期保護タンパク質(Dps)をコードする遺伝子BT4815は、5.8倍にアップレギュレートされた。
最後に、フラボドキシン(FldA; BT0517)とDUF2023タンパク質(BT0516)をコードする鉄非依存性代謝オペロンBT0516〜BT0517は、それぞれ326倍と598倍にアップレギュレートされた。このオペロンは、大腸炎モデル(2)でも高度にアップレギュレートされていることが報告されている((表7)。
TABLE 7
表7 鉄依存性調節因子の臨床的相関性
-Fe/+Feb遺伝子名colitis studydProtein annotation遺伝子の存在ine:1.898914BT_01592.111094VIT1/CCC1トランスポーターファミリー蛋白Bt8.06201BT_02249. 704741Hypothetical proteinBt4.40866BT_022511.11973Hypothetical proteinBt, Pi4.883878BT_022610.46389Hypothetical proteinBt3.419034BT_02277.563568Porin family proteinBt1. 633861BT_0239-1.05735DUF4369ドメイン含有タンパク質Bt, Pg, Pi108.9247BT_049118.86303DUF2149 ドメイン含有タンパク質, MotA/TolQ/ExbB プロトンチャネル・ファミリータンパク質 Bt, Pg, Pi186.9863BT_049228.50811DUF2149 ドメイン含有タンパク質, MotA/TolQ/ExbB プロトンチャネル・ファミリータンパク質 Ba, Pi136. 0809BT_049316.425ハイポhetical proteinBt, Pg, Pi282.4144BT_049624.69941TonB-dependent receptorBt, Pg, Pi290.8443BT_049729. 40818HmuY family proteinBt263.7668BT_049831.73038Hypothetical proteinBt, Pg,2.392774BT_05038.93965Hypothetical proteinBt, Pg2.430991BT_05044. 850745TonB-dependent receptorBt, Pg, Pi51.24636BT_05075.508571TetR/AcrR family transcriptional regulatorBt, Pi, Pi675.5204BT_050815.40959TetR/AcrR family transcriptional regulatorBt, Pi516.2514BT_050911.91826ABC transportter ATP-binding proteinBt,37.04651BT_05153. 708254Quinol oxidaseBt, Pg, Pi598.3778BT_051627.22851DUF2023 family proteinBt, Pg, Pi325.7BT_051729.3FlavodoxinBt, Pg, Pi19.95236BT_05199. 597707Hemerythrinドメイン含有タンパク質、応答制御因子Bt、Pg、Pi16.32074BT_05209.573912Hemerythrinドメイン含有タンパク質、応答制御因子転写因子Bt2. 014698BT_05234.325417Sugar transferaseBt2.142719BT_05241.586668Response regulatorBt,2.007205BT_05327.851734Anthranilate synthase component I family proteinBt, Pg-4.22BT_0545-174P-II family nitrogen regulatorBt-2.5BT0551-167.1Asparagine synthaseBt-3.Bt_Pt_T_Tokyo.BT-0553.1 5BT0552-1,267グルタミン酸合成酵素βサブユニットBt-1.99BT0553-290グルタミン酸合成酵素大サブユニットBt3.711937BT_09224.113622PepSY様ドメイン含有タンパク質Bt, Pg5.042992BT_09233.241869PepSY様ドメイン含有タンパク質Bt2.027212BT_09951. 799071PASドメイン含有センサーヒスチジンキナーゼ、ヘリックスターンヘリックスドメイン含有タンパク質Bt3.760234BT_104310.62892SusD/RagBファミリー栄養結合性外膜タンパク質Bt8. 318422BT_104510.67078DUF1735ドメイン含有タンパク質Bt, Pg, Pi3.099403BT_1138-1.56722hypothetical protein, site-specific integraseBt2.279712BT_15973. 318077Tetratricopeptide repeat protein, HAMP domain-containing histidine kinaseBt3.294873BT_189535.68394Hypothetical proteinBt3.128725BT_189640.56019Leucine-rich repeat proteinBt, Pg3.320197BT_20385.169461Efflux RND transformter periplasmic adaptor subunitBt, Pg2.477183BT_20396.Bt2.29385BT_20485.Hypothetical protein 533055CusA/CzcAファミリー重金属排出RNDトランスポーターBt, Pg2.549452BT_20404.430047TolC family proteinBt489.3635BT_206334.05744PepSY domain-containing proteinBt676. 0915BT_206434.84073DUF4374ドメイン含有タンパク質Bt, Pi662.945BT_206516.46505TonB-dependent receptorBt, Pi3.463556BT_245026. 23424Hypothetical proteinBt62.68564BT_247314.21087Fimbrillin family proteinBt103.9787BT_247529.12251Cytochrome c biogenesis protein ccsa, hypothetical proteinBt225. 3287BT_247628.66125Hypothetical proteinBt146.1537BT_247725.62196PorinBt133.2681BT_247830.08073Thiol oxidoreductaseBt230.1729BT_247932. 79333Peptidase M75Bt283.9951BT_248026.77281Hypothetical proteinBt323.7068BT_248220.39152Helix-turn-helix transcriptional regulatorBt2.264999BT_27781. 409648シグマ70ファミリーRNAポリメラーゼシグマ因子Bt-2.26543BT_3053-3.48253コハク酸脱水素酵素/フマル酸還元酵素シトクロムbサブユニットBt、Pg、Pi-2. 29584BT_3054-2.86087フマル酸還元酵素/コハク酸デヒドロゲナーゼフラボプロテインサブユニットBt, Pg, Pi(p)65.02367BT_362515.21476DUF4857 domain-containing proteinBt36. 69843BT_362614.07917hypothetical proteinBt39.85516BT_362718.25618ABC transporter ATP-binding proteinBt,38.27088BT_362817. 59845PepSY-associated TM helix domain-containing proteinBt43.33606BT_362917.27476PepSY-associated TM helix domain-containing proteinBt97.71608BT_363019.14698Fimbrillin family proteinBt128.0625BT_363114.74463Hypothetical protein, DUF4876 domain-containing proteinBt125.3197BT_363216. 11913仮説タンパク質、DUF4876ドメイン含有タンパク質Bt112.1206BT_363318.93561TonB依存性受容体Bt5.035874BT_3963.185176CusA/CzcAファミリー重金属排出RNDトランスポーターBt2.017194BT_422713.80192Mfa1フィンブリンC末ドメイン含有タンパク質Bt、2.305598BT_42339. 275554DUF3845ドメイン含有タンパク質Bt3.306148BT_42349.429591DUF3575ドメイン含有タンパク質Bt, Pi2.182647BT_4236-1.25097 応答制御因子Bt4.047878BT_45713.342238ハイポhetical protein, RNA polymerase sigma factorBt5.844437BT_471518.7437DNA starvation/stary phase protection proteinBt,Pg,Pi
a
B. thetaiotaomicron VPI BT5482のゲノム上のクラスタリングを示す遺伝子の濃淡ブロック。
b
本研究でのフォールドチェンジ(-Fe/+Fe)。
c
B. thetaiotaomicron VPI BT5482の遺伝子の名称。
d
大腸炎試験(2)におけるFold change。
e
Bt, B. thetaiotaomicron, Pg, P. gingivalis, Pi, P. intermedia.
合計444の遺伝子が2.5倍ダウンレギュレーションされた。制御された遺伝子は51のオペロンに分類された。最も大幅にダウンレギュレートされたオペロンの中には、アンモニアへの異化性亜硝酸還元をコードするオペロンBT1414〜BT1418があり、nrfAH(BT1417〜BT1418)およびシトクロムc生合成コード遺伝子ccsA(BT1415)およびccsB(BT1416)が先行するシトコロームレダクターゼを含む(表6、図3I)。そのオペロンには、アルギン酸輸出ファミリータンパク質をコードする遺伝子も含まれていた(BT1414)。これは、BT1413がコードするCrp様制御因子によって制御されていると予測される。
窒素調節サイクルを拡張するために、窒素固定タンパク質NifUをコードする遺伝子(BT0174)および推定ヒドロキシルアミン還元酵素(Hcp)をコードするhcp(BT0687)(ダウンレギュレート[-3.6倍])もダウンレギュレートした。また、hcpに隣接するウラシル-キサンチンペルミアーゼをコードする遺伝子(BT0686)も2.5倍減少しており、この2つの遺伝子はオペロンを形成していると考えられる。hcpはBT0688がコードするHcpRによって制御されていると予測される。
コハク酸デヒドロゲナーゼ/フマル酸レダクターゼ(BT3055)、RnfABCDGE電子輸送複合体(BT0618〜BT0622)など鉄依存性の代謝機構をコードする遺伝子は、遺伝子によって2.5〜3.4倍のダウンレギュレーションだった。アスパラギン生合成をコードする遺伝子(BT0551)は2.5倍、グルタミン酸合成酵素をコードする遺伝子座(BT0552〜BT0553)は3.5倍ダウンレギュレートされた(図3L)。最後に、4Fe-4Sクラスターを含むヒドロゲナーゼをコードする遺伝子座hydGEF(BT1834〜BT1837)は、遺伝子によって2.9〜3.6倍ダウンレギュレートされた。
フェリチンコード遺伝子(ftn;BT3823)やルブレトリンスリンファミリータンパク質コード遺伝子(BT3182)を含む鉄ホメオスタシスおよび酸化ストレスコード遺伝子は、遺伝子によって2.7倍から3倍にダウンレギュレートされていた(表6)。また、スルファターゼをコードする遺伝子座(BT3796〜BT3799)は、遺伝子によって3.5〜5.3倍減少している(表6)。
TonB依存性受容体およびRag/Susファミリー栄養取り込みタンパク質をコードする遺伝子を含む2成分Sus様トランスポーター系をコードする多数のオペロン(BT4724〜BT4725、BT4294〜BT4299、BT4038〜BT4040、BT3958〜BT3959、BT3951〜BT3953、BT3854〜BT3858.BT3786〜BT3792、BT3344〜BT3347、BT3087〜BT3090、BT3045〜BT3047、BT3024〜BT3028、BT3012〜BT3016、BT2804〜BT2809、BT2614〜BT2632、BT2392〜BT2395、BT1280〜BT1285、BT1046〜BT1047、BT0438〜BT0440、BT0360〜BT0365及びBT0317〜BT0318)はダウンレギュレートを示した(表6)。最後に、共役型トランスポゾンをコードする遺伝子traN、traM、traK(BT0086~BT0089)も、遺伝子によって2.5~7.6倍のダウンレギュレーションが見られた。
全体として、鉄/ヘミンの取り込み、糖質代謝、酸化ストレス、鉄非依存性代謝がアップレギュレートされた。低下したのは、代謝酵素(nrfAH、ccsA、hcp、rnf、hyd)や酸化ストレス(rbr、ftn)、TonBやRagA/Sus輸送系など、鉄補因子を含むタンパク質をコードする遺伝子だった。また、鉄の量が少ないと、DNAの動員に関与する遺伝子の発現も低下した。
Bacteroidetesの鉄依存性刺激。
本研究で解析した3つの細菌種で制御された遺伝子を比較したところ、多くの共通遺伝子が確認された(図4)(遺伝子配列と予測される機能の類似性)(図1〜3)。さらに、多くの遺伝子は、研究対象とした3つの細菌種で同様のゲノム構成をとっていた。共通して制御されているオペロンの中には、ヘミンの更新機構をコードするhmu遺伝子があった(表1、表3、表5、図1A)。また、フラボドキシンをコードする遺伝子は、試験した3つの細菌種すべてで高度にアップレギュレートされていた(図1B)。この遺伝子座は、以下のように、fldAと仮説タンパク質をコードする下流遺伝子の2つで構成されている。B. thataiotaomicronでは、fldA遺伝子と下流遺伝子(BT0517とBT0516)はオペロンを形成していると予測される。このような予測は、両遺伝子の著しいアップレギュレーションによってさらに立証される(表5)。両遺伝子は口腔内細菌にも存在することから、これらも共転写されている可能性が高い。TonB依存性受容体を含む2つのオペロンは、それぞれ3菌(図1C)および2菌(図1D)でアップレギュレートされていた。口腔内細菌種(P. gingivalisとP. intermedia)で劇的にアップレギュレートされたのはPG0495(87倍アップ)とPIOMA14_RS06105(ChI;104倍アップ)(P. intermedia 17ではPIN17_RS05350)(図1E、図S1)。この遺伝子はT9SSのC末端ターゲットドメインタンパク質をコードしている。注目すべきは、P. intermediaでは、この遺伝子座はhmuYの隣に位置していることである(Fig. 1E)。
図4
図4 バクテロイデーテスにおける鉄依存的な遺伝子制御の模式図。鉄制限条件下で増殖した細菌において、アップレギュレートされた遺伝子は緑色で、ダウンレギュレートされた遺伝子は赤色で示されている。
また、共通にダウンレギュレートされる遺伝子もいくつかあった。コハク酸デヒドロゲナーゼ/フマル酸レダクターゼをコードするオペロンsdhABC(図2A)、電子伝達鎖をコードするrnf遺伝子(図2B)、2-オキソグルタル酸フェリドキシン酸化還元酵素をコードするvor遺伝子(図2C)などの鉄系代謝コード遺伝子が、試験した3種のバクテリアすべてでダウンレギュレーションされていました。ルブレイトリンコード遺伝子rbrは、B. thetaiotaomicron(BT3812が3倍ダウンレギュレート)よりもP. gingivalisとP. intermedia(PG0195とPIOMA14_II_0105がそれぞれ46倍と22.3倍の変化)で、より劇的にダウンレギュレートした(図2D、表2、4および6)。なお、B. thetaiotaomicronは、推定鉄取り込み制御タンパク質FurをコードするBT0215に続く別のルブレリトリン(BT0216)をコードしているが、この遺伝子座は鉄による有意な影響を受けなかった。また、アルキルヒドロペルオキシド還元酵素をコードするahp遺伝子座(図3B)は、P. gingivalisとB. thetaiotaomicronで発現が上昇することが判明した。
その他の有意に制御された遺伝子の中には、ゼノシデロフォア獲得機構をコードするBT2063〜BT2065遺伝子座があった(2)。その遺伝子座はB. thataiotaomicronにのみ存在し、P. gingivalisとP. intermedia 17には存在しない(図3A)。また、B. thetaiotaomicronにのみ存在するのは、グルタミン酸合成酵素をコードする遺伝子座BT0552〜BT0553と、ヒドロゲナーゼ系をコードする遺伝子座BT1834〜BT1837だった。もう一つの注目すべき遺伝子座であるメチルトランスフェラーゼをコードする遺伝子座は、アップレギュレートされたが(図3C)、P. gingivalis W83においてのみであった。最後に、亜硝酸還元酵素をコードするnrfAH遺伝子座は、試験した3つの細菌すべてで見つかった(図3D)。しかし、B. thetaiotaomicronのみ、鉄欠乏によって劇的な影響を受けることが判明した(表6)。興味深かったのは、P. gingivalisにのみ存在したDUF1661ドメインタンパク質コード遺伝子群である(補足資料の図S4参照)。このグループは、鉄欠乏に対するP. gingivalisの適応において独自の機能を持つ可能性のある低分子タンパク質をコードする遺伝子ファミリーである。
全体として、フラボドキシンをコードする遺伝子が大幅に増加し、フェレドキシンをコードする遺伝子が同時に減少していることが非常に印象的であった。一貫して、多くの鉄ベースの代謝タンパク質がダウンレギュレートされた。鉄の取り込み機構は、おそらく細胞内の鉄不足を緩和するために発現量が増加した(図4)。
B. thetaiotaomicronの亜硝酸塩還元は、鉄制限条件下で減少する。
調査したすべての細菌種は、亜硝酸還元をコードする遺伝子、アンモニア形成性チトクロームc亜硝酸還元酵素系NrfAH(図3D)を保有しているが、細菌の亜硝酸利用能は実証されていない。ここでは、3つの細菌種すべてが、培養混合物中に存在する亜硝酸レベルを還元することを示す(図5AおよびB)。この能力は菌種によって異なり、B. thetaiotaomicronは亜硝酸塩を最も効率よく還元することができる。B.taiotaomicronは、48時間以内に培養液中に存在する2mMの亜硝酸塩を検出できないレベルまで還元することができる(Fig. 5B)。P. intermedia OMA14は亜硝酸塩を還元する能力で2位、P. gingivalis W83は0.3mMの亜硝酸塩を72時間以内に検出できないレベルまで還元することが確認されました(図5A)。鉄の枯渇はB. thetaiotaomicronの亜硝酸還元能力を低下させたが(48時間の時点でP = 0.065)(図5B)、P. gingivalisとP. intermediaによる亜硝酸還元には影響を与えなかった(結果は示さず)。これらの知見は、鉄制限条件下でB. thetaiotaomicronの転写レベルの有意な減少が観察されるという、トランスクリプトームシーケンス解析によって得られたデータをさらに機能的に検証するものである。
図5
図5 バクテロイデスには亜硝酸塩を還元する能力がある。(A) B. thetaiotaomicron VPI VPI BT5482 Δtdkを、TSB培地中、150μM DPの有無にかかわらず、2mM亜硝酸ナトリウムの存在下および非存在下で培養した。(B) P. gingivalis W83およびP. intermedia OMA14を、BHI培地中、0.3mM亜硝酸ナトリウムの存在下および非存在下で増殖させた。OD660を測定することで成長をモニターし、亜硝酸塩のレベルはGriess試薬で測定した。
バクテロイデーテスにおける鉄依存性制御の臨床的相関性
大腸炎時にB. thetaiotaomicronで発現が上昇する遺伝子(2)と、本研究で鉄欠乏条件下で発現が上昇する遺伝子には、大きな重複があった(表7)。また、多くの遺伝子がオペロン構造に編成されている68の遺伝子(15のオペロン様組織)が、2つの研究間で重複していることが確認された(Table 7)。いくつかの遺伝子座は、試験した3つの細菌全てに存在した。
まず、フラボドキシン(FldA;BT0517)とDUF2023タンパク質(BT0516)をコードする遺伝子座は、我々のモデルと大腸炎モデルの両方で劇的に発現が上昇した(表5、7)(2)。P. gingivalis のフラボドキシンをコードする遺伝子 fldA は、ヒトの歯周炎患者由来のマイクロバイオームにおいて最も高発現する遺伝子の一つであることが判明しており、鉄欠乏によってアップレギュレートされる細胞外炎症環境における遺伝子も高発現することが示されている (19).
さらに、本研究と大腸炎時の両方で有意に発現が上昇した遺伝子座はhmu遺伝子座である(表7)。さらに、TetR/AcrRファミリー転写制御因子と2つのABCトランスポーターATP結合タンパク質をコードするBT0507〜BT0509遺伝子座も、本研究と大腸炎中に発現が上昇した(表7、図1D)。注目すべきは、P. intermediaのゲノム上に同様のオペロンのコピーが2つ存在し(一方、小型染色体上のコピーは調査した他の細菌と同一)、大型染色体上のコピーには別の遺伝子(ABCトランスポーターコード遺伝子)が含まれていたことだ(図1D, ii)。同様の遺伝子座はP. gingivalisにも存在し、ABCトランスポーターをコードする遺伝子に続いて、外膜リポタンパク質選別タンパク質(PG1179)と仮説タンパク質(PG1178)をコードする遺伝子が存在している。興味深いのは、トランスポザーゼをコードするPG1177に続いて、BT0508からBT0509のオルソログをコードするPG1176からPG1175が存在することである(図1D、ii)。また、試験した他の2つの細菌に類似の遺伝子を持つモバイルエレメントをコードする遺伝子BT1138も、両方の試験で制御されていた。さらに、コハク酸デヒドロゲナーゼ/フマル酸還元酵素SdhABCをコードする遺伝子座BT3053〜BT3055は、両方の研究でダウンレギュレーションされた(表7、図2A)。最後に、DNAスターベーション/定常期保護タンパク質をコードするBT4715 dpsは、本研究で調査した3つの細菌全てに存在する。興味深いことに、B. thetaiotaomicronではdpsの上流に逆向きのoxyR(BT4716)が存在し(表7)、dpsの発現に関与している可能性を示している。
鉄量や大腸炎によって制御されるB. thetaiotaomicronの遺伝子座で、本研究で調査した他の細菌のうち1つだけに対応する遺伝子座が存在した。BT0503〜BT0504遺伝子座は、B. thetaiotaomicronとP. gingivalisで、仮説タンパク質をコードする最初の遺伝子(BT0503)が両菌で同じで、その後に、配列は異なるが機能的に同様にTonB依存性のタンパク質をコードする遺伝子(BT0504とPG2008)が続いている構成である。同様に、BT0519からBT0520は、2成分のヘメリトリンドメイン制御タンパク質をコードしている(表7、図3F)。このオペロンはP. gingivalis W83にも存在し、CdhR制御因子をコードしている(20)。さらに、PepSY様ドメイン含有タンパク質をコードするBT0922〜BT0923遺伝子座、および重金属排出トランスポーター系をコードするBT2038〜BT2040遺伝子座がB. thataiotaomicronとP. gingivalis両方に存在した(表7)。
最後に、B. thetaiotaomicronにのみ存在する、鉄と大腸炎の条件で制御される遺伝子座があった(表7)。これらの遺伝子座には、糖転移酵素をコードするBT0523〜BT0524とその同族応答制御因子BT0532をコードするアントラニル酸合成酵素コンポーネントIファミリータンパク質、PASドメイン含有センサーヒスチジンキナーゼをコードするBT0995、SusD/RagBトランスポーターシステムをコードするBT1042〜BT1045の遺伝子座がある、 HAMPドメイン含有ヒスチジンキナーゼをコードするBT1597(次いで、システムの第2の構成要素と陽イオン排出ポンプBEXをコードする遺伝子)、および大腸炎で非常にアップレギュレーションされる外膜ロイシンリッチ受容体システムをコードするBT1895〜BT1896(表7)。
シグマ70ファミリーRNAポリメラーゼシグマファクターであるBT2778、推定フィンブリリン合成/輸送遺伝子座をコードする多遺伝子座BT3625〜BT3633、重金属トランスポーターをコードするBT3969、MfaIフィンブリリン蛋白質をコードするBT4227、2成分応答レギュレーターをコードするBT4236、RNAポリメラーゼECFタイプシグマファクターをコードするBT4771はB. thetaiotaomicronのみに存在していました (Table 7).
B. thetaiotaomicronの遺伝子座の中には、調査した他の細菌に存在する選択した遺伝子のオルソログを持つものもあった。したがって、BT2063からBT2065遺伝子座は、P. intermediaにも存在するTonB依存性受容体をコードするBT2065のオルソログを除いて、B. thetaiotaomicronに主に存在した。BT2450は、仮説タンパク質であるパイロジェニックエキソトキシンB(BT2451)と2つの仮説タンパク質(BT2452〜BT2453)をコードする遺伝子座の最初の遺伝子であり、BT2451に類似する遺伝子であるpdnAは、P. gingivalis(PG1427)に存在し、ペリオドンテイン軽鎖をコードするほか、P. intermedia(PIOMA14_I_1549)に存在し、C10ファミリーペプチダーゼをコードする(補足資料の図 S3参照)。また、BT4233はB. thetaiotaomicronにのみ存在したが、BT4234はP. intermediaにオルソログが存在した。
最後に、両方の研究(2)(鉄レベルの高低は本研究と参考文献2に記載の大腸炎研究を指す)で高度にアップレギュレートされたのは、BT2473〜BT2482 fimbrillin-family protein、2つのシトクロムc biogenesis proteins (CcsA)、ポリン、thiol oxidoreductase、ペプチド酵素M75 (helix-turn-helix transcriptional regulator, and three hypothetical proteins with 69. 4-から323.7倍のアップレギュレーション)であった(表5、7、図3G)。
我々の研究でも大腸炎の研究でも、ダウンレギュレートされたパスウェイがあった。主なダウンレギュレーション遺伝子座は、グルタミン酸合成酵素(glt)をコードするBT0552からBT0553であった。また、アスパラギン合成酵素をコードする遺伝子も両試験でダウンレギュレートされた。さらに、BT0545がコードする窒素制御因子が有意にダウンレギュレートされていた。上記の遺伝子はB. thetaiotaomicronでのみ見つかったが、コハク酸デヒドロゲナーゼ/フマル酸レダクターゼをコードするダウンレギュレーションオペロンは、研究した3菌全てに存在する(表7)。
考察
バクテロイデーテス門に属する3つの細菌種を並行して研究した結果、これらの生物における鉄のホメオスタシスについて洞察することができた。感染症やそれに伴う炎症時には鉄が不足するため、宿主の状態は我々の研究で報告されたような鉄制限状態に類似している(21-24)。ほとんどのin vitro研究は、鉄が十分に存在する条件下で行われているため、本研究では、宿主環境への適応をより正確に描写することができます。さらに、後者については、大腸炎モデルを用いたin vivoでの研究で得られた結果と比較することで検証した(2)。我々は、P. gingivalisの鉄依存性トランスクリプトームについて報告したが、本研究では、この研究をさらに発展させ、バクテロイデーテスに属する他の細菌と比較し、この細菌群における鉄依存性制御に関するコンセンサスを得ることを最終目標としている。我々は、鉄によって発現が変化するP. intermediaのタンパク質を報告したことがあるが、P. intermediaの鉄依存性制御に関する最初の包括的な報告であり、このデータは、以前に発表した我々の発見(16)および他の研究室からのデータ(25)を強化するものである。しかし、B. thetaiotaomicronの鉄依存性制御については、これまで比較的知られていませんでした。本研究では、鉄分充足および鉄分キレート条件下で増殖させた細菌由来のRNAを用いて、スチムロンの包括的な解析を行ったことを示す。これは、B. thetaiotaomicronのゲノムサイズが非常に大きいこと(26)、およびB. thetaiotaomicronのライブラリーはHiSeq Illuminaシーケンサーで、他の2つのライブラリーセットはMiSeq Illuminaシーケンサーでシーケンスしたため、HiSeq Illuminaシーケンサーでのリード数が非常に多くなっているためと考えられる。
鉄/ヘムの獲得、代謝、酸化ストレスなど、いくつかの経路を仲介するタンパク質をコードする遺伝子発現に変化が見られた。これらの経路の多くは、3つの細菌種の間で共有されています。最も劇的に変化した経路のひとつは代謝で、最も発現量が増加したのはフラボドキシンをコードする遺伝子(FldA)であった。FldAの構造は、最初に大腸菌のFldAを用いて決定された(27)が、P. gingivalisのFldAはDesulfovibrioのFldA構造とより似ていることに注目した(28)。Flavodoxinは電子伝達タンパク質で、様々な細菌、藻類、下等植物に存在する(29-31)。低分子のフラビンモノヌクレオチド(FMN)含有タンパク質で、微生物の様々な代謝過程で電子伝達物質として機能する。電子移動は、結合した補酵素FMNの露出したジメチルベンゼン環を介して行われ、その後FMNH2に還元されます。FldAは、半世紀以上前に、最初はシアノバクテリアから、後にクロストリジウムから単離された (32, 33) 。FldAの発現は、鉄制限条件下で誘導されることが示されたが、他のストレスによっても発現が制御されることがある。FldAの生物学的役割はフェレドキシンと同様で、FldAがドナーから細菌細胞膜にあるRnf複合体をはじめとする電子伝達系構成要素に電子を伝達し、ナトリウムプロトン勾配を形成してATP生成を促進する(34)。フェレドキシン(Frds)は鉄-硫黄クラスターを用いて電子を移動させるが、FldAは補酵素であるフラビンモノヌクレオチド(FMN)と結合するため、フェレドキシンよりも鉄を節約でき、鉄に依存する他の必須酵素の活性や酸化的/硝酸的ストレスによる損傷に強いという利点がある。FldAの発現が劇的に上昇する一方で、鉄制限条件下ではFrdsの発現が減少することも確認された。このような鉄に依存した発現は、他の生物でも指摘されている(29, 35)。炎症が起きると鉄分が減少し、炎症細胞から放出される酸化ストレスメディエーターが増加するため、FldAを上昇させFrdを置換することで適応し、このような状況下で細菌が持続するための直感的な戦略のように思える。また、FldAは大腸炎モデルのB. thetaiotaomicronで非常に発現量が多く、その転写物は歯周病患者のP. gingivalisで非常に豊富であることから(19)、我々のデータは翻訳上の意義もあると思われる。このようなデータは、P. gingivalisが病気の活動状態において、より鉄に依存しない酵素に依存する新しい代謝機構を採用していることを示しており、したがって、実験室で鉄分の多い条件下で増殖した場合に細菌が採用するこれまでの好ましい代謝モデルとは対照的である (36). FldAは、高等真核生物には存在しない細菌特有の特徴を持つ酵素であり、将来的に治療戦略を開発するためのターゲットとなる可能性があることは注目に値する(31, 37)。
もう1つは、鉄・ヘム獲得機構である。ここでは、よく定義されたシステムであるヘミン取り込みオペロンhmuが、試験した3つの細菌種すべてでアップレギュレートされていることが観察された。P. gingivalisとB. thetaiotaomicronでは1つのオペロン/ゲノムが同定されたが、P. intermedia OMA14では、大型染色体(Ch I)にhmuYのみを持つ遺伝子座、小型染色体(Ch II)にhmuオペロンが認められた(表3)。興味深いことに、P. intermedia 17ではhmu遺伝子座も重複して別々の染色体上に存在するが、オーファンhmuYは小型染色体(ChII)に、完全なオペロンはChrIに存在する(図S1)。後者の観察は、Olczak研究室からの最近の報告(25, 38)と一致する。我々は、3つの細菌すべてにおいて、完全なhmuオペロンと、孤児であるP. intermedia 17 hmuY遺伝子の劇的なアップレギュレーションを観察した。このような結果は、P. intermediaの鉄依存性プロテオームに関する我々の過去の報告(16, 17)と一致するものである。また、B. thetaiotaomicronでは、大腸炎条件下でこのオペロンが健康な動物に比べて発現量が増加することも注目される(2)。歯周炎では、全身の鉄を減少させるヘプシジンレベルが上昇する(21, 39)。したがって、歯周炎では、微生物による鉄の取り込み機構が健常部位と比較して著しく上昇する(40, 41)。細胞外鉄が少ない条件下では、hmuシステムを用いたヘミンの輸送による鉄の獲得が直感的な戦略であると思われるが、TonB依存性タンパク質やABC型トランスポーターを含む2つの推定トランスポーターシステムのアップレギュレーションも観察された。これらの系は、試験したほとんどの細菌種において、鉄欠乏条件下で高い発現量を示したことから、低鉄条件下での細菌の増殖と生存を媒介する役割について、さらなる調査が必要であると考えられた。そのようなシステムの一つ、すなわちBT2063〜BT2065は、最近B. thetaiotaomicronで特徴づけられ、Salmonella enterica serovar Typhimuriumが生産するxenosiderophoresの輸送を通じて鉄獲得に重要な役割を果たすことが示された(2)。この遺伝子座はB. thetaiotaomicronで最も劇的に発現が上昇し、調査した他の2つの細菌では類似性(オルソログ)がない。また、炎症状態(大腸炎モデル)に由来するB. thetaiotaomicronでは、非炎症の宿主のものと比べて最も発現量が増加しており(2)、今回の発見の生物学的意義がさらに証明された。
また、ABC型トランスポーターやTonB依存性タンパク質を含む遺伝子座の発現量が高いことから、口腔内または腸内細菌叢に豊富に存在するシデロフォアからの鉄獲得に関与している可能性が指摘されています。P. gingivalisやP. intermediaのような口腔内細菌では、口腔環境に豊富に存在するラクトフェリンから鉄を獲得することが可能な戦略であると思われる(3, 42)。また、精製された多くの細菌性シデロフォアは、in vitroの条件下でラクトフェリンよりも鉄Fe(III)に対する親和性が高い(43-46)という知見も、この戦略を立証するものであろう。したがって、P. gingivalisとP. intermediaは、シデロフォア供給者が豊富に存在する複雑な口腔内微生物群に存在する細菌であるため、他の細菌が生産するシデロフォアを「盗む」能力が存在する可能性が高い(43)。
我々は、アミノ酸レベルでの同一性が低いにもかかわらず、調査した3つの細菌間で類似した機能を持つタンパク質を多遺伝子座の中に見出した。この発見の例として、CdhRを持つ遺伝子座が考えられる。さらに、遺伝子座の中の遺伝子の順序が異なっている可能性もある。例えば、トランスポーターと関連するレギュレーターをコードする遺伝子座PG1180からPG1181は、試験した細菌間で異なっている。遺伝子座内の遺伝子の分布も異なるため、個々の遺伝子を探すよりも、ゲノムの文脈で制御される経路を見た方が、低鉄環境適応時に制御されなくなるメカニズムを予測することができた。
また、生物間の差異も検出され、主にB. thetaiotaomicronの糖質利用を媒介する遺伝子の制御が含まれていた。この結果は、B. thetaiotaomicronが炭水化物の利用に依存していることから、細菌の生息するニッチが異なることに起因すると考えられる(47)。バクテロイデーテスは、糖鎖の分解に使用する精巧な酵素の組み合わせを持っている(48)。P. gingivalisとP. intermediaのゲノムはそれぞれ2.3Mbと2.2〜2.8Mb(さらにミニゲノムが0.6〜0.9Mb)と小さいが、Bacteroides thetaiotaomicronのゲノムサイズは6.26Mb (VPI-4582 strain) で、倍以上大きい (26, 49-51). P. gingivalisは糖鎖分解酵素をコードする遺伝子を持たないため、この細菌はエネルギー生成をペプチドに依存している(27, 49)。鉄制限条件下で、P. intermediaとB. thetaiotaomicronでTonB依存性メカニズムのダウンレギュレーションが観察されたことは注目すべきことである。このような観察は、鉄欠乏条件下でTonB依存性機構を過剰発現させるという予測される結果と相反する。しかし、それらの機構が他の栄養素を輸送する可能性があり、ここでは糖鎖輸送に関与すると予測されることは一致している(47, 52)。
調べたすべての細菌が亜硝酸塩を還元することができたが、B. thetaiotaomicronではその能力は鉄に依存していた。このような知見は、Goffらによって報告された最近の知見とも一致する(53)。さらに、試験したすべての細菌は亜硝酸還元酵素系NrfAHを有しており、B. thetaiotaomicronオペロンだけが鉄制限条件下で劇的にダウンレギュレートされている。従って、このオペロンがその細菌の生理に大きな役割を果たしていると考えられる。鉄キレート剤存在下で培養した菌では、発現量と亜硝酸塩消費量の減少が見られたが、B. thetaiotaomicronは鉄制限条件下でも亜硝酸塩を大量に消費することができることが確認された。このような能力は、B. thetaiotaomicronだけでなく、P. gingivalisやP. intermediaも、硝酸塩還元による有害な副産物を無毒化して宿主内で生き残るのに役立つと考えられる。特に硝酸塩を多く含む食事の後では、口腔内の亜硝酸塩濃度が1mMを超えることが示されている(54-56)。さらに、免疫細胞によって展開される宿主の防御機構の1つは一酸化窒素の生成であり、その一酸化窒素は亜硝酸塩に酸化されるため、それらの細菌には有効なニトロソストレス解毒機構が期待されます。
また、鉄によって制御される小さな遺伝子も多数検出された。DUF1661ドメイン含有タンパク質をコードするこれらの遺伝子の役割は、まだ確立されていない。これらの遺伝子はP. gingivalisに特異的であり、他の細菌におけるこれらの遺伝子との類似性は検出されなかったことは注目に値する。
本研究で鉄分制限条件下で発現が増加した遺伝子と、健常人とは対照的に大腸炎で発現が増加した遺伝子との間の驚くべき類似性は、バクテロイデスが宿主の炎症状態に適応するためのマスターレギュレーターが鉄であることを指摘している。これまでのところ、バクテロイデス属細菌が異なる鉄レベルに適応するために必要なエフェクタータンパク質の発現を制御する分子メカニズムに関する知見は乏しいままである。今回、3つの細菌種で共通に制御されている上記の遺伝子が同定されたことで、これらの細菌における鉄依存的な制御システムの特定に役立つことが予想されます。CdhRはhmuYRの発現を制御することが示されている(20, 57)が、CdhRが鉄と結合することがまだ示されていないため、その分子機構は不明である。さらに、鉄制限条件下で発現する遺伝子と炎症性疾患の活動時に高発現する遺伝子との臨床的相関を考えると、本研究で同定された発現が急激に上昇する機構は生物学的意義を持ち、その機能や分子機構をさらに詳細に検討する必要があると思われる。
以上、バクテロイデーテス門に属する3種の細菌の鉄依存性刺激物について比較検討した。鉄の取り込み機構に加え、代謝機構や酸化ストレス応答機構にも大きな適応が見られた。鉄が制限された状態が炎症を起こした宿主に似ているという観察は、細菌感染に対する自然免疫反応として鉄の隔離を示す数十年にわたるデータ(21-24)と一致する。この結果は、全体として、鉄が細菌の病的状態への適応を制御する主要なプレーヤーであることを示しています。最終的には、鉄ベースのエフェクタータンパク質の分子機構と、バクテロイデーテス門で今のところ解明されていない制御機構について、より詳細な調査が必要である。
材料と方法
細菌株と増殖条件
本研究では、以下の3つの細菌種を使用した: Porphyromonas gingivalis (Strain W83) (58), Prevotella intermedia (strain OMA14), Bacteroides thetaiotaomicron (strain BT5482 Δtdk). このうち2種は口腔内細菌であり、B. thetaiotaomicronは腸内細菌である。すべての細菌は臨床的に重要であり、P. intermedia OMA14株は最近配列が決定された臨床分離株である(49)。また、より毒性の強いW83株との比較のために、P. gingivalisの実験室株、すなわちATCC 33277株も含めている。細菌は、37℃の嫌気性チャンバー(10%H2、10%CO2、80%N2)内の血液寒天プレート(TSA II with 5% sheep blood; BBL, Cockeysville, MD)上で維持されました。ブロス培養は、濃縮脳心筋梗塞(BHI)培地(37 g/L BHI、2.5 g/L 酵母エキス、5 μg/mL ヘミン、および 0.5 μg/mL メナジオン)を用いて調製した。培養は嫌気的条件下で維持し、37℃で攪拌せずに培養した(静置培養)。鉄欠乏条件は、BHIに150μMのジピリジル(DP)を補充することで生成した。細胞はミッドログフェーズで収穫した(600nmの光学密度[OD660]が約0.5)。
RNA単離およびライブラリー調製。
RNA単離およびライブラリー調製は、以前に記載したように行った(59、60)。簡単に言うと、RNeasy minikit(Qiagen)を用いて、製造者のプロトコルに従って細菌ペレットからRNAを単離した。RNAは、製造者のプロトコールに従って、DNAフリーDNaseキット(Ambion、Thermo Fisher)で処理し、残存するDNAを除去した。アガロースゲル電気泳動でRNAの品質を確認し、Ovation complete prokaryotic RNASeq DR multiplex kit (Universal Prokaryotic RNAse multiplex kit [Tecan]) を用いて、メーカーの指示に従いRNAseqライブラリーを作成しました。ライブラリーはバージニア・コモンウェルス大学(VCU)の核酸配列決定コア施設に提出し、バイオアナライザー(Agilent Technologies)を使用してライブラリーの品質を検証した。
ハイスループット配列決定とデータ解析。
検証され、許容できる品質を有するライブラリーは、(VCU)Nucleic Acid Sequencingコアにより、MiSeq Illuminaゲノムアナライザーを用いて(P. gingivalisおよびP. intermediaライブラリー)、およびIllumina HiSeq 4000プラットフォームを用いて(B. thetaiotaomicron)シーケンスされました。その後、CLC Genomic Workbench V12(CLC Bio、Qiagen)を用いてシーケンスデータを解析した。こうして得られたトランスクリプトーム由来の配列を、P. gingivalis W83の参照ゲノム(P. gingivalis W83トランスクリプトーム用)、P. gingivalis ATCC 33277(P.の参照ゲノム)にアラインした。gingivalis ATCC 33277 transcriptome)、P. intermedia OMA14(P. intermedia OMA14 transcriptome)、B. thetaiotaomicron VPI BT5482(B. thetaiotaomicron VPI BT5482 Δtdk transcriptome)。各注釈付き遺伝子のリード数を抽出し、後述の統計解析に使用した。
バイオインフォマティクス解析。
BioCyc genome viewerを使用して、選択した遺伝子のゲノム遺伝子座を整列させた(61)。類似の遺伝子座のアライメントは、本研究で使用した3つの異なる細菌種にわたって行われた。使用した参照ゲノムは、P. gingivalis W83、P. intermedia 17(P.インターメディアOMA14ゲノムはBioCycで入手不可)、およびB. thetaiotaomicron BT5482である。
亜硝酸塩還元アッセイ。
細菌株は血液寒天培地プレートで維持した。アッセイは、0.5 mg/mL システイン、5 μg/mL ヘミン、および 0.5 μg/mL メナジオンを補充したBHIブロス中で行った。鉄欠乏条件は、150μMのジピリジル(DP)を培地に補充することで生成した。亜硝酸塩は、P. gingivalis/P. intermediaおよびB. thetaiotaomicronにそれぞれ0.3および2mMで添加した。BHIブロスで調製した活発に増殖する一夜培養液を1:30(P. gingivalis W83およびP. intermedia OMA14用)または1:50(B. thetaiotaomicron用)に希釈して37℃で72時間嫌気的に培養した。次に、異なる時点で1mLのアリコートを取り出し、細菌の増殖と亜硝酸のレベルを測定するために使用した。コントロールとして、亜硝酸塩を含む鉄強化培地(DPなし)および鉄欠乏培地(DPを補充)を、細菌培養と並行して維持した。
細菌増殖は、660nmでの光学密度の測定によってモニターし、亜硝酸塩レベルは、製造者の説明書(Abcam)に従ってGriess試薬を使用して決定した。
統計解析。
すべての実験は、少なくとも4つの別々の日に繰り返された(生物学的複製)。鉄分補給条件下および鉄分欠乏条件下でのmRNAのレベルの変化の分析については、4つの生物学的複製物から倍率変化およびP値を導出した。解析には、細菌特異的リードと遺伝子特異的リードを使用した。鉄依存性制御の濃縮度は、正規化後にゲノムの各位置について、鉄欠乏条件で培養した細菌由来のリード数を、鉄完全条件で培養した細菌由来のリード数で割ることにより求めた(59、60、62)。RNAseqデータの統計解析は、CLC Workbench 12のToolbox|Workflows|RNA統計ワークフロー(Qiagen)で利用できるプログラムとデフォルト設定を使用して実施した。簡単に説明すると、Robinsonら(63)によって概説されたDGE(EdgeR)の経験的分析が分析に使用された。このテストは、限られた生物学的複製で多くの特徴が同時に調べられるサンプル用に設計されている。解析には、順に正規化および変換された生のカウントが使用された。2群の不対比較は、RPKM(reads per kilobase per million)(RPKMは、転写物に割り当てられたリードの数を転写物長で割り、マップされたリードで正規化したものとして定義される)値(64)を用いて実施された。RNAseqデータの統計解析の実施には、t検定を使用した。また、鉄欠乏条件下および鉄完全条件下で調製した培養物の少なくとも3つの生物学的複製に由来するデータを用いて行った亜硝酸アッセイの統計的有意性を決定するために、t検定を使用した。
データの利用可能性
RNAseqデータは、NCBI Gene Expression Omnibus (GEO)に寄託されている。データは、アクセッション番号GSE169260(B. thetaiotaomicron BT5482 Δtdkについて)、GSE168982(P. intermedia OMA14について)、GSE168570(P. gingivalis W83について)およびGSE174493(P. gingivalis ATCC 33277について)で入手できます。
謝辞
本研究は、NIHグラント1R01DE023304(J.P.L.)、NIHR21DE02555(J.P.L.)、K18 DE029865(J.P.L.)により資金提供されました。図は、BioCyc Pathways Tools version 26.0 (software by SRI International)を用いて作成した。
J.P.L.は実験を行い、すべてのデータを分析した。J.P.L.は原稿を執筆した。Q.G.は実験を行った。
申告すべき利益相反はない。
補足資料
File (spectrum.04733-22-s0001.pdf)
ダウンロード
602.94 KB
ASMは、論文にリンクされている、または論文を通じてアクセスできる補足資料の著作権を保有していません。著者はASMに対して、Supplemental Materialファイルを公開するための非独占的かつ世界的なライセンスを付与しています。再利用する場合は、対応する著者に直接連絡してください。
参考文献
1.
Mayneris-Perxachs J, Cardellini M, Hoyles L, Latorre J, Davato F, Moreno-Navarrete JM, Arnoriaga-Rodríguez M, Serino M, Abbott J, Barton RH, Puig J, Fernández-Real X、 Ricart W, Tomlinson C, Woodbridge M, Gentileschi P, Butcher SA, Holmes E, Nicholson JK, Pérez-Brocal V, Moya A, Clain DM, Burcelin R, Dumas M-E, Federici M, Fernández-Real J-M. 2021. 鉄の状態は、腸内細菌叢を通じて肥満における非アルコール性脂肪性肝疾患に影響を与える。Microbiome 9:104.
引用文献に移動する
クロスフィルム
PubMed
Google Scholar
2.
Zhu W, Winter MG, Spiga L, Hughes ER, Chanin R, Mulgaonkar A, Pennington J, Maas M, Behrendt CL, Kim J, Sun X, Beiting DP, Hooper LV, Winter SE. 2020. ゼノシデロフォア利用は大腸炎時のBacteroides thetaiotaomicronの回復を促進する。Cell Host Microbe 27:376-388.e8.
Crossref
PubMed
アイエスアイ
Google Scholar
3.
Chathoth K, Martin B, Cornelis P, Yvenou S, Bonnaure-Mallet M, Baysse C. 2020年。歯肉縁下ディスバイオシスの原因となりうる事象:鉄、硫化物、酸素の相互作用に着目して。FEMS Microbiol Lett 367:fnaa100.
クロスレフ
PubMed
アイエスアイ
Google Scholar
4.
Boyer E, Le Gall-David S, Martin B, Fong SB, Loréal O, Deugnier Y, Bonnaure-Mallet M, Meuric V. 2018年. トランスフェリン飽和度の上昇は、遺伝性ヘモクロマトーシスにおける歯肉縁下微生物叢異種および重度歯周炎と関連している。Sci Rep 8:15532.
引用文献へ移動する
クロスレフ
PubMed
Google Scholar
5.
Kirst ME, Li EC, Alfant B, Chi Y-Y, Walker C, Magnusson I, Wang GP. 2015. 慢性歯周炎における歯肉縁下マイクロバイオームの予測される機能におけるディスバイオシスと変化。Appl Environ Microbiol 81:783-793.
引用文献へ移動する
クロスレフ
パブコメ
ISI
Google Scholar
6.
Jiang Y, Song B, Brandt BW, Cheng L, Zhou X, Exterkate RAM, Crielaard W, Deng DM. 2021. 歯周炎患者の唾液と歯肉縁下プラーク間の赤色複合細菌の比較:システマティックレビューとメタアナリシス。Front Cell Infect Microbiol 11:727732.
引用文献に移動する
クロスフィルム
PubMed
ISI
Google Scholar
7.
マーク・ウェルチJL、デューハーストFE、ボリジGG. 2019. 口腔内マイクロバイオームの生物地理学:サイトスペシャリスト仮説。Annu Rev Microbiol 73:335-358.
引用文献へ移動する
クロスレフ
パブコメ
国際標準化機構
Google Scholar
8.
Ghosh S, Pramanik S. 2021年。ヒト腸内細菌叢の構造的多様性、機能的側面、および将来の治療的応用。Arch Microbiol 203:5281-5308.
引用文献に移動する
クロスフィルム
パブコメ
アイエスアイ
Google Scholar
9.
ドナルドソンGP、リーSM、マズマニアンSK. 2016. 細菌性微生物叢の腸内生物地理学。Nat Rev Microbiol 14:20-32.
引用文献に移動する
クロスレフ
パブコメ
アイエスアイ
Google Scholar
10.
ウェクスラーHM. 2007. Bacteroides: the good, the bad, and the nitty-gritty. Clin Microbiol Rev 20:593-621.
引用文献に移動する
クロスレフ
パブコメ
国際標準化機構
Google Scholar
11.
Hayase E, Hayase T, Jamal MA, Miyama T, Chang C-C, Ortega MR, Ahmed SS, Karmouch JL, Sanchez CA, Brown AN, El-Himri RK, Flores II, McDaniel LK, Pham D, Halsey T, Frenk AC, Chapa VA, Heckel BE, Jin Y, Tsai W-B, Prasad R、 Tan L, Veillon L, Ajami NJ, Wargo JA, Galloway-Peña J, Shelburne S, Chemaly RF, Davey L, Glowacki RWP, Liu C, Rondon G, Alousi AM, Molldrem JJ, Champlin RE, Shpall EJ, Valdivia RH, Martens EC, Lorenzi PL, Jenq RR. 2022. 粘液分解性バクテロイデスがカルバペネムを移植片対宿主病の悪化に繋げる。Cell 185:3705-3719.e14.
引用文献に移動する
クロスフィルム
PubMed
国際標準化機構
Google Scholar
12.
Cavazza C, Contreras-Martel C, Pieulle L, Chabrière E, Hatchikian EC, Fontecilla-Camps JC. 2006. ピルビン酸-フェレドキシン酸化還元酵素とピルビン酸との反応の動力学的結晶学的研究により明らかになったチアミン二リン酸の柔軟性。Structure 14:217-224.
引用文献に移動する
クロスフィルム
PubMed
アイエスアイ
Google Scholar
13.
Chabrière E, Vernède X, Guigliarelli B, Charon MH, Hatchikian EC, Fontecilla-Camps JC. 2001. ピルビン酸:フェレドキシン酸化還元酵素のフリーラジカル中間体の結晶構造。Science 294:2559-2563.
クロスフィルム
PubMed
アイエスアイ
Google Scholar
14.
Lewis JP, Plata K, Yu F, Rosato A, Anaya C. 2006. Porphyromonas gingivalis W83 hmu haemin-uptake遺伝子座の転写組織、制御および役割。微生物学(読書)152:3367-3382.
Crossref
PubMed
ISI
Google Scholar
15.
Anaya-Bergman C, Rosato A, Lewis JP. 2015年. Porphyromonas gingivalisの鉄およびヘミン依存的な遺伝子発現. Mol Oral Microbiol 30:39-61.
引用文献に移動する
クロスレフ
PubMed
Google Scholar
16.
Yu F, Anaya C, Lewis JP. 2007. Prevotella intermedia 17の外膜プロテオーム:チオレドキシンと鉄抑制性ヘミン取り込み遺伝子座の同定。Proteomics 7:403-412.
クロスフィルム
PubMed
ISI
Google Scholar
17.
Otto BR, Verweij-van Vught AM, van Doorn J, Maclaren DM. 1988. Bacteroides fragilisおよびBacteroides vulgatusの外膜タンパク質と鉄の取り込みおよび病原性との関係。Microb Pathog 4:279-287.
クロスレフ
PubMed
アイエスアイ
Google Scholar
18.
Phillips P, Progulske-Fox A, Grieshaber S, Grieshaber N. 2014. マイクロアレイおよびRNA-seq解析を用いて特定されたヘミン利用可能性に応じたPorphyromonas gingivalis small RNAの発現. FEMS Microbiol Lett 351:202-208.
引用文献に移動する
クロスフィルム
PubMed
国際標準化機構
Google Scholar
19.
Lewin GR, Stocke KS, Lamont RJ, Whiteley M. 2022年。定量的な枠組みにより、従来の実験室での増殖がヒト口腔内感染の高精度モデルであることが明らかになった。Proc Natl Acad Sci USA 119:e2116637119.
Crossref
PubMed
国際標準化機構
Google Scholar
20.
Chawla A, Hirano T, Bainbridge BW, Demuth DR, Xie H, Lamont RJ. 2010. Streptococcus gordoniiとPorphyromonas gingivalis間のコミュニティシグナルは、転写制御因子CdhRによって制御されている。Mol Microbiol 78:1510-1522.
クロスレフ
PubMed
ISI
Google Scholar
21.
Han Y, Huang W, Meng H, Zhan Y, Hou J. 2021年。炎症性サイトカインであるインターロイキン6が誘発するヘプシジン、歯周炎関連炎症性貧血のキーメディエーター。J Periodontal Res 56:690-701.
Crossref
PubMed
アイエスアイ
Google Scholar
22.
カルヴァーリョRCC、レイテSAM、ロドリゲスVP、ペレイラAFV、フェレイラTCA、ナシメントFRF、ナシメントJR、ゴメス-フィリョIS、バストスMG、ペレイラALA.2016. 慢性歯周炎とヘプシジンおよびヘモグロビンの血清レベル。オーラル・ディス22:75-76.
クロスレフ
PubMed
アイエスアイ
Google Scholar
23.
Nemeth E, Ganz T. 2021年。ヘプシジン-フェロポーティン相互作用は全身の鉄ホメオスタシスを制御する。Int J Mol Sci 22:6493.
クロスレフ
PubMed
アイエスアイ
Google Scholar
24.
Nairz M, Weiss G. 2020. 感染と免疫における鉄。Mol Aspects Med 75:100864.
Crossref
PubMed
国際標準化機構
Google Scholar
25.
Bielecki M, Antonyuk S, Strange RW, Siemińska K, Smalley JW, Mackiewicz P, Śmiga M, Cowan M, Capper MJ, Ślęzak P, Olczak M, Olczak T. 2020年. Prevotella intermediaは、Porphyromonas gingivalis HmuYに相同だがヘム配位様式が異なる2つのタンパク質を産生する。Biochem J 477:381-405.
クロスレフ
PubMed
アイエスアイ
Google Scholar
26.
Xu J, Bjursell MK, Himrod J, Deng S, Carmichael LK, Chiang HC, Hooper LV, Gordon JI. 2003. ヒト-Bacteroides thetaiotaomicron共生のゲノム的見解。Science 299:2074-2076.
クロスレフ
PubMed
国際標準化機構
Google Scholar
27.
Hoover DM, Ludwig ML. 1997. 酵素活性化に必要なフラボドキシン:Escherichia coli由来酸化フラボドキシンの1.8A分解能での構造。Protein Sci 6:2525-2537.
クロスフィルム
PubMed
ISI
Google Scholar
28.
O'Farrell PA, Walsh MA, McCarthy AA, Higgins TM, Voordouw G, Mayhew SG. 1998. Desulfovibrio vulgaris flavodoxinにおけるFMNの酸化還元電位の調節:グリシン61変異体の熱力学的性質と結晶構造。Biochemistry 37:8405-8416.
引用文献に移動する
クロスフィルム
PubMed
アイエスアイ
Google Scholar
29.
Pierella Karlusich JJ, Lodeyro AF, Carrillo N. 2014. 長いお別れ:植物進化におけるフラボドキシンの盛衰。J Exp Bot 65:5161-5178.
クロスフィルム
PubMed
国際標準化機構
Google Scholar
30.
Simondsen RP, Tollin G. 1980. フラボドキシンにおける構造-機能関係。Mol Cell Biochem 33:13-24.
引用文献へ
クロスレフ
PubMed
アイエスアイ
Google Scholar
31.
Cremades N, Velázquez-Campoy A, Martínez-Júlvez M, Neira JL, Pérez-Dorado I, Hermoso J, Jiménez P, Lanas A, Hoffman PS, Sancho J. 2009年。ヘリコバクター・ピロリ感染に対する治療薬となりうる特異的なフラボドキシン阻害剤を発見した。ACS Chem Biol 4:928-938.
クロスフィルム
PubMed
国際標準化機構
Google Scholar
32.
Smillie RM. 1965. アオミドロから葉緑体フェレドキシン活性を持つ2つのタンパク質を単離した。Biochem Biophys Res Commun 20:621-629.
引用文献へ
クロスフィルム
PubMed
アイエスアイ
Google Scholar
33.
Knight E, D'Eustachio AJ, Hardy RW. 1966. Flavodoxin: a flavoprotein with ferredoxin activity from Clostrium pasteurianum. Biochim Biophys Acta 113:626-628.
引用文献へ
クロスフィルム
PubMed
Google Scholar
34.
バッケルW、タウアーRK. 2018. フラビンベースの電子分岐、フェレドキシン、フラボドキシン、およびプロトン(Ech)またはNAD+(Rnf)を電子受容体とする嫌気性呼吸:歴史的レビュー。Front Microbiol 9:401.
引用元へ
クロスフィルム
PubMed
国際標準化機構
Google Scholar
35.
ホイットニーLP、リンスJJ、ヒューズMP、ウェルズML、チャペルPD、ジェンキンスBD. 2011. 鉄ストレスの種特異的な指標となる鉄応答性遺伝子の特性評価(Thalassiosiroid Diatoms). Front Microbiol 2:234.
引用文献へ
クロスフィルム
パブコメ
アイエスアイ
Google Scholar
36.
Meuric V, Rouillon A, Chandad F, Bonnaure-Mallet M. 2010. Porphyromonas gingivalis の推定呼吸鎖。Future Microbiol 5:717-734.
引用文献に移動する
クロスフィルム
PubMed
国際標準化機構
Google Scholar
37.
Salillas S, Sancho J. 2020. ヘリコバクター・ピロリおよび他の胃病原体に対する新規治療標的としてのフラボドキシン。Int J Mol Sci 21:1881.
引用文献に移動する
クロスフィルム
PubMed
アイエスアイ
Google Scholar
38.
Siemińska K, Cierpisz P, Śmiga M, Olczak T. 2021年。Porphyromonas gingivalis HmuYとBacteroides vulgatus Bvu-新規競合ヘム獲得戦略。Int J Mol Sci 22:2237.
引用文献に移動する
クロスレフ
PubMed
国際標準化機構
Google Scholar
39.
Wu D, Lin Z, Zhang S, Cao F, Liang D, Zhou X. 2019年. 歯周炎におけるヘモグロビン濃度の低下と鉄代謝障害:システマティックレビューとメタアナリシス。Front Physiol 10:1620.
引用文献へ移動する
クロスレフ
パブコメ
アイエスアイ
Google Scholar
40.
Solbiati J, Frias-Lopez J. 2018. 健康と病気における口腔内マイクロバイオームのメタトランスクリプトーム(Metatranscriptome of the oral microbiome in health and disease. J Dent Res 97:492-500.
引用文献に移動する
クロスレフ
PubMed
国際情報通信学会
Google Scholar
41.
Yost S, Duran-Pinedo AE, Teles R, Krishnan K, Frias-Lopez J. 2015. 微生物メタトランスクリプトーム解析により明らかになった歯周炎進行中の口腔内ディスバイオシスの機能的シグネチャー。Genome Med 7:27.
引用文献に移動する
クロスレフ
PubMed
アイエスアイ
Google Scholar
42.
Wang R, Kaplan A, Guo L, Shi W, Zhou X, Lux R, He X. 2012. 鉄の利用可能性がヒトの唾液微生物群集組成に与える影響。Microb Ecol 64:152-161.
引用文献に移動する
クロスフィルム
PubMed
アイエスアイ
Google Scholar
43.
Butaitė E, Baumgartner M, Wyder S, Kümmerli R. 2017. Siderophore cheating and cheating resistance shape competition for iron in soil and freshwater Pseudomonas communities. Nat Commun 8:414.
クロスレフ
PubMed
グーグル・スカラー
44.
Schryvers AB, Stojiljkovic I. 1999. 病原性ナイセリアにおける鉄獲得システム。Mol Microbiol 32:1117-1123.
引用文献に移動する
クロスレフ
PubMed
アイエスアイ
Google Scholar
45.
Wilson BR, Bogdan AR, Miyazawa M, Hashimoto K, Tsuji Y. 2016. 鉄代謝におけるシデロフォア:メカニズムから治療の可能性まで。Trends Mol Med 22:1077-1090.
引用文献へ移動
クロスレフ
PubMed
アイエスアイ
Google Scholar
46.
ホールデンVI、バックマンMA. 2015. 感染時の細菌性シデロフォアの乖離した役割。メタロミクス7:986-995.
引用文献に移動する
クロスレフ
PubMed
グーグル奨学生
47.
Lapébie P, Lombard V, Drula E, Terrapon N, Henrissat B. 2019. Bacteroidetesは何千もの酵素の組み合わせを使って糖鎖を分解している。Nat Commun 10:2043.
クロスレフ
PubMed
グーグル・スカラー
48.
McKee LS, La Rosa SL, Westereng B, Eijsink VG, Pope PB, Larsbrink J. 2021. バクテロイデーテス類による多糖類分解:機構と命名法。Environ Microbiol Rep 13:559-581.
引用文献に移動する
クロスフィルム
PubMed
国際標準化機構
Google Scholar
49.
内藤真也、小倉由美子、伊藤貴志、庄司正人、岡本真也、林哲也、中山恭子...2016. Prevotella intermedia OMA14株の全ゲノム配列決定とその後の微細な種内ゲノム比較により、共役トランスポゾンと移動性トランスポゾンの異常な増幅が明らかになり、新規Prevotella系統特異的反復が特定された。DNA Res 23:11-19.
クロスレフ
PubMed
国際標準化機構
Google Scholar
50.
Haigh RD, Crawford LA, Ralph JD, Wanford JJ, Vartoukian SR, Hijazi K, Wade W, Oggioni MR. 2017年. 歯周病菌Porphyromonas gingivalis、Prevotella intermedia、Tannerella forsythiaのドラフト全ゲノム配列は、相変化型制限修飾系を含む。Genome Announc 5:e01229-17.
引用元へ
クロスフィルム
PubMed
Google Scholar
51.
コムストック・LE、コイン・MJ. 2003. Bacteroides thetaiotaomicron: a dynamic, niche-adapted human symbiont. Bioessays 25:926-929.
引用文献に移動する
クロスレフ
パブコメ
国際標準化機構
Google Scholar
52.
仁平亨、鈴木絵理子、北岡正樹、西本雅彦、大坪和彦、中井浩之、2013年。N-糖鎖の代謝に関与するβ-1,4-D-マンノシル-N-アセチル-D-グルコサミンホスホリラーゼを発見した。J Biol Chem 288:27366-27374.
引用文献に移動する
クロスフィルム
PubMed
アイエスアイ
Google Scholar
53.
Goff JL, Chen Y, Thorgersen MP, Hoang LT, Poole FL, Szink EG, Siuzdak G, Petzold CJ, Adams MWW. 2023. 混合重金属ストレスは、グローバルな鉄飢餓応答を誘導する。ISME J 17:382-392.
引用文献に移動する
クロスフェード
パブコメ
国際標準化機構
Google Scholar
54.
Lundberg JO, Weitzberg E, Cole JA, Benjamin N. 2004. 硝酸塩、細菌、人間の健康。Nat Rev Microbiol 2:593-602.
引用文献に移動する
クロスフィルム
パブコメ
国際標準化機構
Google Scholar
55.
Velmurugan S, Gan JM, Rathod KS, Khambata RS, Ghosh SM, Hartley A, Van Eijl S, Sagi-Kiss V, Chowdhury TA, Curtis M, Kuhnle GGC, Wade WG, Ahluwalia A. 2016年. 食餌性硝酸塩は高コレステロール血症患者の血管機能を改善する:無作為化二重盲検プラセボ対照試験。Am J Clin Nutr 103:25-38.
引用文献へ移動する
クロスレフ
PubMed
アイエスアイ
Google Scholar
56.
Hezel MP, Weitzberg E. 2015. 口腔内マイクロバイオームと一酸化窒素のホメオスタシス。オーラル・ディス21:7-16.
引用文献に移動する
クロスレフ
PubMed
情報通信研究機構
Google Scholar
57.
太田紘一、菊池洋一、今村和彦、喜多大輔、吉川和彦、斉藤晃、石原和彦...2017. Porphyromonas gingivalisの細胞質外機能シグマ因子SigCHはcdhRとhmuYRの発現を制御している。Anaerobe 43:82-90.
引用元へ
クロスフィルム
PubMed
ISI
Google Scholar
58.
Belvin BR, Gui Q, Hutcherson JA, Lewis JP. 2019年. Porphyromonas gingivalis hybrid cluster protein Hcp is required for growth with nitrite and survival with host cells. Infect Immun 87:e00572-18.
引用文献に移動する
クロスレフ
PubMed
国際標準化機構
Google Scholar
59.
Karp PD, Billington R, Caspi R, Fulcher CA, Latendresse M, Kothari A, Keseler IM, Krummenacker M, Midford PE, Ong Q, Ong WK, Paley SM, Subhraveti P. 2019年. 微生物ゲノムと代謝パスウェイのBioCycコレクション。Brief Bioinform 20:1085-1093.
クロスレフ
PubMed
アイエスアイ
Google Scholar
60.
Yanamandra SS, Sarrafee SS, Anaya-Bergman C, Jones K, Lewis JP. 2012. Porphyromonas gingivalis の細胞質外機能シグマ因子、SigH の役割。Mol Oral Microbiol 27:202-219.
クロスレフ
PubMed
Google Scholar
61.
Robinson MD, McCarthy DJ, Smyth GK. 2010. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. バイオインフォマティクス26:139-140.
引用文献に移動する
クロスフィルム
パブコメ
国際標準化機構
Google Scholar
62.
Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B. 2008. RNA-Seqによる哺乳類トランスクリプトームのマッピングと定量化。Nat Methods 5:621-628.
引用文献に移動する
クロスフィルム
PubMed
国際標準化機構
Google Scholar
63.
Nelson KE, Fleischmann RD, DeBoy RT, Paulsen IT, Fouts DE, Eisen JA, Daugherty SC, Dodson RJ, Durkin AS, Gwinn M, Haft DH, Kolonay JF, Nelson WC, Mason T, Tallon L, Gray J, Granger D, Tettelin H, Dong H, Galvin JL, Duncan MJ, Dewhirst FE, Fraser cm. 2003. 口腔病原性細菌porphyromonas gingivalis strain W83の全ゲノム配列。J Bacteriol 185:5591-5601.
引用文献に移動する
クロスフィルム
PubMed
ISI
Google Scholar
64.
内藤真一、Belvin BR、庄司真一、Gui Q、Lewis JP. 2021年。Prevotella intermedia OxyRの挿入型不活性化により、酸化ストレスおよび宿主細胞存在下での生存率が低下する。Microorganisms 9:551.
引用文献に移動する
クロスフィルム
PubMed
Google Scholar
全文を見るPDFをダウンロードする
この記事が気に入ったらサポートをしてみませんか?