
レプリコンワクチン (コスタイベ筋注用 開発コードARCT-154) の遺伝子構造解析

ベネズエラウマ脳炎ウイルス
Meiji Seikaファルマのレプリコンワクチンは自己増殖型mRNAワクチンであり、販売名は「コスタイベ筋注用」です。このレプリコンワクチンの設計に使用されたベネズエラウマ脳炎ウイルス (VEEV) はトガウイルス科のアルファウイルスです。ベネズエラウマ脳炎ウイルスは主に蚊に媒介され、もともと小型齧歯類と蚊の間での感染のサイクルが存在しています。そして蚊を媒介して馬に感染すると、馬に高い死亡率での高力価ウイルス血症を引き起こします。さらには馬と密接に接触する人間にも蚊を介して感染し得ます。
タイトルの画像はWikipediaからベネズエラウマ脳炎ウイルスと近縁の東部ウマ脳炎ウイルスです (ウイルスが赤く色分けされています)。
ベネズエラウマ脳炎ウイルスについては以下のSharmaとKnollmann-Ritschelによる総説論文を参考にしました。
Current Understanding of the Molecular Basis of Venezuelan Equine Encephalitis Virus Pathogenesis and Vaccine Development
Sharma and Knollmann-Ritschel (2019) viruses
https://www.mdpi.com/1999-4915/11/2/164
ベネズエラウマ脳炎ウイルスは主に嗅覚神経を通って脳に侵入しますが、効率は低いものの血液脳関門を通過する経路がある事も知られています。このウイルスはまず脳の嗅球で複製されます。一般的に脳内に侵入したウイルスは免疫系のグリア細胞によって検出され、放出されるケモカインやサイトカインが炎症反応を起こします。また、感染初期の炎症反応が血液脳関門を損傷すると、血液中を循環するウイルス粒子や白血球が脳内に侵入できるようになり炎症を憎悪させます。最終的にニューロンはウイルスによる直接感染によって誘発されるアポトーシスに加え、炎症によっても傷害されます。このようにしてベネズエラウマ脳炎ウイルスはヒトに重篤な脳炎を引き起こします。
また、高い感染力と脳を障害するという軍事的な「魅力」から、ベネズエラウマ脳炎ウイルスは生物兵器の材料としても知られています。実際、冷戦中にはアメリカとソビエト連邦の両国がベネズエラウマ脳炎ウイルスの兵器使用を目的とした研究を行っていました。
ベネズエラウマ脳炎ウイルスのゲノム構成
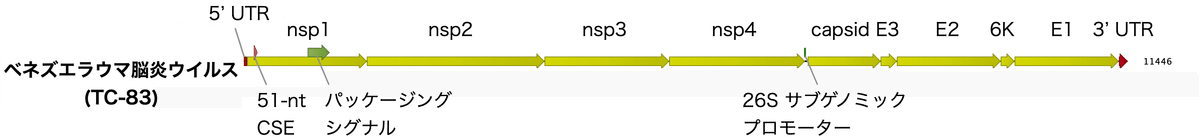
図1は私自身で描いたもので、ベネズエラウマ脳炎ウイルスのゲノム構成です。遺伝子配列は弱毒性ワクチン株 (TC-83株) のものです。例えば研究材料として、ベネズエラウマ脳炎ウイルスの弱毒化生株であるTC-83はバイオセーフティレベル2で対処できるのに対し、病原性ベネズエラウマ脳炎ウイルス株はバイオセーフティレベル3での取り扱いが求められます。
ベネズエラウマ脳炎ウイルスのゲノムは11.4 kbの一本鎖のプラス鎖RNAです。このゲノムは4つの非構造タンパク質 (nsp1、nsp2、nsp3、nsp4) と5つの構造タンパク質 (キャプシド (C)、エンベロープ (E) 3、E2、6K、E1) をコードしています。それらの遺伝子以外には5’非翻訳領域 (UTR)、26Sサブゲノムプロモーター、3’非翻訳領域 (UTR) があり、ゲノムの3’末端にはポリ(A) 尾部が存在します。
nsp1、nsp2、nsp3、nsp4はまず1つのタンパクとして翻訳され、nsp2のプロテアーゼ活性によって4つのタンパク質に分解されます。nsp1-3はウイルスRNAをウイルス粒子にパッケージングするために重要な酵素ですが、その中でもnsp2はウイルスゲノムRNAのウイルス粒子へのパッケージングを制御し、ウイルス粒子の成熟にも関与します。また、nsp2は宿主細胞の翻訳を阻害し、感染細胞においてウイルスを効率的に産生します。こうした酵素の働きのため、レプリコンワクチンを受け取った細胞そのものが損傷を受ける可能性があります。
nsp4はRNA依存性RNAレプリカーゼであり、ウイルスRNAのプラス鎖およびマイナス鎖の合成を担います。また、nsp4は末端アデニルトランスフェラーゼとしても働き、ウイルスRNAゲノムにポリAテールを付加します。
一般的なウイルスと同様に、ベネズエラウマ脳炎ウイルスでもゲノムのほとんどはタンパクをコードする領域と制御領域で構成されており、余分な配列はほとんど存在しません。これは限られた大きさのウイルスゲノムの中に効率的に遺伝子を配置するためです。そして、1つの酵素が複数の機能を持つ事により酵素の数を最小限に抑えています。また、このウイルスのゲノムの特徴としては、転写やパッケージングの制御領域が非翻訳領域だけではなくタンパクのコード領域内にも含まれている事があげられます。
レプリコンワクチンARCT-154の遺伝子構成
レプリコンワクチン「コスタイベ筋注用」の有効成分名が「ザポメラン」、開発コード名は「ARCT-154」です。開発はアメリカのArcturus Therapeutics社によるものです。ARCT-154は起源株に近いB.1 変異株 (D614G変異スパイクタンパク) を用いており、ベトナムなどで治験が行われました。その後、開発者側によりレプリコンワクチンのスパイクタンパクはJN1株に変更され、8匹のマウスでの実験を経て、そのまま日本人への接種が開始されました。事実上、日本は開発国ですら承認されていないレプリコンワクチンを異例のはやさで世界で唯一承認したばかりか、一般への接種を開始したのです。
コスタイベ筋注用 に関する資料
https://www.pmda.go.jp/drugs/2023/P20231122002/780009000_30500AMX00282_B100_2.pdf
公開されている「コスタイベ筋注用に関する資料 」には黒塗りの部分が多いのですが、ARCT-154の遺伝子配列はこの資料の中に記載されていますので、そのデータを用いてARCT-154の遺伝子構成について私自身で解析してみました。今回使用したアプリはCLC Genomics Workbench (Qiagen) です。
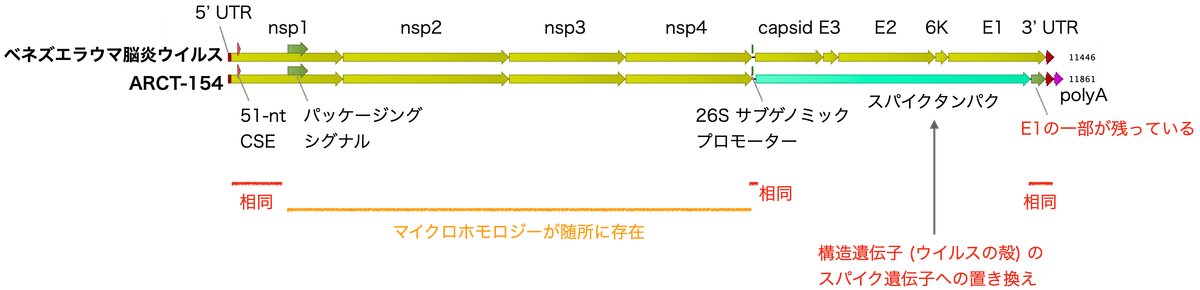
図2はベネズエラウマ脳炎ウイルスゲノムとARCT-154の比較です。図で示されるように、ARCT-154は構造遺伝子 (ウイルスの殻の遺伝子など) がスパイク遺伝子に置き換えられたものです。
ARCT-154のRNAレプリカーゼ複合体やスパイクでは、アミノ酸配列を保ちながらコドン最適化の過程で遺伝子配列が変更されています。配列を詳しく見ると、ARCT-154内にはウイルス由来のE1遺伝子の一部が残っているのが確認できます。相同な配列はウイルスと組換えを起こす原因となるのですが、実際ARCT-154とベネズエラウマ脳炎ウイルスには配列がほぼ同一の箇所 (5' UTR、nsp1のN末端側、サブゲノムプロモーター、E1遺伝子のC末端側、3' UTR側) と微小な相同性 (マイクロホモロジー) を持つ箇所 (nsp1のC末端側とnsp2から4) が存在します。

図3は図2の左端からnsp1までの領域について、ベネズエラウマ脳炎ウイルスとARCT-154の配列を比較したものです。それぞれの列で上がベネズエラウマ脳炎ウイルス (VEEV TC-83株)、下がARCT-154です。両者ともゲノムはRNAですが、ここではデータベースに登録されているTC-83の情報に合わせ、Uの代わりにTで表記しました。
転写制御配列がnsp1遺伝子内に存在するためか (51塩基の保存された配列 (51-nt CSE))、nsp1遺伝子のN末端側では互いの配列は同一です。これは、この領域の機能を失わずに遺伝子配列を変更する事が難しいからでしょう。一方、nsp1遺伝子のC末端側ではコドンを保ちながら配列が置き換えられていますが、随所に微小な相同性 (マイクロホモロジー) が残されています。
また、パッケージングシグナルもnsp1のコード領域内に存在しています。nsp1はレプリカーゼ複合体を構成する要素であり、レプリコンワクチンに必須な遺伝子です。そのため、レプリコンワクチンのデザインからパッケージングシグナル領域を除去する事は原理的に不可能なのです。
そもそもARCT-154はウイルスの構造遺伝子 (ウイルスの殻) をスパイク遺伝子に置き換えた構造をしています。では、「ウイルスの殻」が供給されると、レプリコンワクチンがウイルス粒子に取り込まれる可能性はあるのでしょうか?
パッケージングシグナル
アルファウイルスの中でもベネズエラウマ脳炎ウイルスは特に効率的にウイルス粒子を形成します。このウイルス粒子はエアロゾルとしても感染しますが、その感染力が非常に強いのです。「パッケージングシグナル」とはウイルスゲノムをウイルス粒子に「配達するための荷札 (タグ)」の様なものであり、ウイルス粒子に取り込む目印として使われます。
パッケージングシグナルの配列自体はアルファウイルス間ではさほど保存されていません。パッケージングシグナルの機能は配列そのものよりも、その領域の複雑な折れ曲がり構造によるもので、とりわけステム (幹) とループ (輪) による折れ曲がり構造とループ内の「GGG」配列がパッケージングシグナルに重要です。
実際、mRNAは一本鎖であるため折れ曲がりやすく、分子内でA-T、C-Gの対により複雑な構造を取ります。ヒトゲノムなどのDNAは二重螺旋構造を取りますが、RNAとは対照的に二本鎖DNAは「曲がりにくい」という物理化学的性質をもっています。
図4はCLC Genomics Workbenchによってパッケージングシグナル領域のRNA高次構造を私自身で解析したものです。図4 Aはベネズエラウマ脳炎ウイルスのパッケージングシグナルの高次構造です。RNAが折れ曲がりステムとループを形成しており、ループ内にいくつかの「GGG」配列が見つかります。
折れ曲がりのパターンは異なりますが、ARCT-154のパッケージングシグナル領域も折れ曲がります (図4 B)。そして数は減少しているものの、ループ内には「GGG」や「GG」配列も見られます。しかしながら、実際の実験データが開示されない限りは、パッケージングシグナルの改変配列のみによってレプリコンワクチンのウイルス様粒子への取り込みのレベルを判断する事は難しいでしょう。
ベネズエラウマ脳炎ウイルスにおいては、パッケージングシグナルの配列を大きく変化させたり「GGG」を取り除いたりするとウイルス粒子産生は減少するものの、残存活性が見られます。また、ゲノムの他の領域にも隠れたパッケージングシグナルが存在する可能性があります。
レプリコンワクチンはウイルス様粒子に取り込まれるか
結論として、ARCT-154の遺伝子構造から考察できる事は以下になります。
レプリコンワクチンのウイルス粒子への取り込みについて
懸念1) パッケージングシグナルは取り除けていない。
懸念2) パッケージングシグナルがnsp1遺伝子上にあるためにパッケージングシグナルを完全に取り除く事は不可能である。
懸念3) ウイルス粒子へのパッケージングに必要な酵素はレプリコンワクチンの中に含まれている。
レプリコンワクチンと野生ウイルスの組換えについて
懸念4) 部分的な相同性のためにアルファウイルスとの組換えは起こり得る。
懸念5) 組換えによって完全なパッケージングシグナルを再取得する事もあり得る。
懸念6) 組換えによって新規キメラウイルスを誕生させる可能性がある。
レプリコンワクチン接種者がアルファウイルスに感染すると、接種者の体内でワクチンがウイルス様粒子に取り込まれる可能性があります。また、そうしたウイルス様粒子が他者に感染するとレプリコンワクチンの個体間伝播が起こり得ます。ベネズエラウマ脳炎ウイルスが蚊によって媒介されるようにレプリコンワクチン接種者からの蚊による個体間伝播も現実的な懸念事項です。
もともとベネズエラウマ脳炎ウイルスは脳炎を引き起こすウイルスであり、その性質を利用した兵器としても研究されてきた経緯のあるウイルスです。レプリコンワクチンによる脳への影響についての懸念も払拭できないと私は考えます。
#コロナワクチン
#ワクチン
#コロナ
*記事は個人の見解であり、所属組織を代表するものではありません。