ニセクロスジギンポの攻撃擬態 ~エサが無い時の最後の手段~ 論文紹介

ニセクロスジギンポの攻撃擬態 ~エサが無い時の最後の手段~
論文名 The false cleanerfish relies on aggressive mimicry to bite fish fins when benthic foods are scarce in their local habitat
偽清掃魚は海底の食料が少ない時には攻撃擬態に頼る
著者名 Misaki Fujisawa, Yoichi Sakai & Tetsuo Kuwamura
掲載誌 Scientific Reports
掲載年 2020年
リンク https://www.nature.com/articles/s41598-020-65304-6
ニセクロスジギンポの攻撃擬態についての2020年の論文です。
擬態とは、動物が他の動物や周囲のものに似た色形を持つことです。良く知られているのが、毒を持つチョウ(ウォーレスドクチョウ)にそっくりな羽を持つ毒を持たないチョウ(ヘリコリウスタイマイ)です。毒を持たないチョウは、毒チョウに似せることで、捕食者に毒を持っていると思わせ、捕食されないようにしています。この擬態はベイツ型擬態と呼ばれる保護擬態のひとつです。また、カマキリは、周囲の草に似せることで自身を草に紛れ込ませ、油断した相手を捕食しますが、これも擬態のひとつになります。この場合は捕食するための擬態ということで、攻撃擬態と呼ばれています。 このように、擬態には自身を守るための保護擬態と、捕食するための攻撃擬態があります。
この論文では、ニセクロスジギンポを研究対象にしています。漫画で紹介したように、見た目がホンソメワケベラにそっくりです(漫画「ホンソメワケベラとニセクロスジギンポ」参照)。ホンソメワケベラは、清掃魚として知られていますが、他の魚にそのように認識されているようで、他の魚に近寄っても攻撃されません。ニセクロスジギンポはホンソメワケベラに似せることで、他の魚から攻撃されない、つまり、保護擬態を利用していると考えられているようです(この論文によると、この保護擬態について実験的に確かめられてはいないようです)。また、別な報告では、ホンソメワケベラのふりをして魚に近づきそのヒレを食べているとも言われています。この場合は捕食のための擬態となり、攻撃擬態として使用していることになります。ニセクロスジギンポの攻撃擬態については、生息地の違いによって報告がことなることから、この論文では、その違いを生み出している条件について野外観察によって調べています。
動物の生態研究において、飼育している動物を使って研究した場合に、どうしても、野生の動物でもそうなのか?という疑問が残ってしまいます。その疑問を解決するためには自然環境、つまりその動物が生息している環境で観察するしかありません。しかし、その生息地に研究者が入り込むことが難しい場合が多々あります。この論文では、石垣島の海中に生息しているニセクロスジギンポを、そのまま観察しています。観察した個体の数が少ないことや、魚卵を食べる行動を観察できなかったことなどから、この海中観察が簡単なものではなかったことを伺わせます。データ数の少なさから、はっきりとしていない点もありますが、ニセクロスジギンポの生態の一部が明らかにされています。大変な研究だとは思いますが、より多くのデータの蓄積が進み、保護擬態も含めたニセクロスジギンポの生態が明らかになることを期待したいと思います。
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
・ニセクロスジギンポは攻撃擬態を利用し魚のヒレを食べる。
・攻撃擬態を利用するかどうかは、海底の食料の量によって変化する。
[背景]
攻撃擬態は、捕食者や寄生者(擬態者)がより捕食しやすくするために第3者の生物(騙される者)には魅力的もしくは無害にみえる別の生物(モデル)に良く似せる擬態の型です。この擬態は、クダクラゲ、クモ、昆虫、蛇、鳥、魚などの多くの動物種の間で広がっています。捕食者から身を守るための保護擬態(ベイツ型擬態)も、場合によっては攻撃擬態に関与しています。
ホンソメワケベラ(ベラ科)によく似た偽清掃魚であるニセクロスジギンポ(イソギンポ科)の場合では、擬態の2つの機能、つまり攻撃と保護が考えられています。これは清掃共生がよくみられるサンゴ礁に生息する魚の中でも最もよく知られている擬態の例です。主に水槽内の魚の観察によって、この魚の攻撃擬態の機能に注目されました。つまり、ニセクロスジギンポは騙されて清掃をお願いする魚や、近づくことを許した魚のヒレの一部に咬み付くことが出来ます。しかし、沖縄のサンゴ礁での定量的野外観察と胃の内容物解析から、ニセクロスジギンポは主にチューブワームであるイバラカンザシとケヤリムシの触手を食べ、時にはスズメダイの巣に入り込み、その卵を食べていましたが、魚のヒレに咬み付くことはほとんどありませんでした。ヒレに咬み付く以外に、攻撃擬態はニセクロスジギンポの摂食行動に関与していないように見え、このような食料の多様性については、他の動物でみられる攻撃擬態の例では報告されていません。他の擬態動物は食べるために攻撃擬態を常に利用しています。さらに、擬態魚ではなく主に糸状藻類や時にイバラカンザシを食べ、魚のヒレは決して食べない近縁のクロスジギンポと食料や摂食行動を比較すると、ニセクロスジギンポがホンソメワケベラに擬態する主な機能は保護擬態であると結論できます。
近年、ニセクロスジギンポの摂食行動の地域による変化が報告されました。グレートバリアリーフ、インドネシア、紅海では、ニセクロスジギンポが魚のヒレに咬み付くことはほとんどありませんでした。ニセクロスジギンポは沖縄で観察されたようにスズメダイの卵やチューブワームを食べていました。しかし、フランス領ポリネシアでは、ニセクロスジギンポはたびたびヒレに咬み付いていました。このことから、地域による変化をもたらす因子は不明で、今後の研究が必要ですが、攻撃擬態の相対的重要性は場所によって変化すると結論されました。また、インドネシアではヒレ咬みは幼若体でたびたび観察されることから、擬態タイプの相対的重要性は、生活史においても変化すると考えられます。
これまでに、年齢による擬態の変化は沖縄の瀬底島での野外観察で報告されています。ニセクロスジギンポは攻撃擬態を全長が小さい時のみ利用します。ヒレ咬みの頻度は成長とともに低下し、卵を食べる頻度が増加しました。対照的に、イバラカンザシの触手やヒメシャコガイのヒダを体の大きさに関わらず食べていました。イバラカンザシやヒメシャコガイといった海底資源の量は場所によって変化し、ニセクロスジギンポによるヒレ咬みの頻度に影響を与える可能性が考えられます。
本研究では、生活史に加えて、生息地の限られた海底食料がニセクロスジギンポの摂食行動において攻撃擬態への依存度に影響すると仮説を立てました。この仮説を検証するために、瀬底島の南西約400 Kmに位置する石垣島の裾礁に生息するニセクロスジギンポの摂食行動を野外観察しました。石垣島の裾礁ではニセクロスジギンポが好むイバラカンザシやヒメシャコガイといった海底食料はほとんどみあたりませんが、瀬底島のサンゴ礁では非常に豊富です。石垣島で得られたデータを以前報告された瀬底島のデータと比較することで、ニセクロスジギンポの摂食行動におけるヒレ咬みの依存度の違いを調べ、条件付き摂食戦略について地域による変化を促す因子を検討しました。
[結果]
本研究では、石垣島で8個体のニセクロスジギンポを野外観察し、以前に報告された瀬底島の40個体のデータと比較しました。
食料の量
瀬底島と同様に、石垣島では、ニセクロスジギンポはイバラカンザシの触手、ヒメシャコガイのヒダ、他の魚のヒレ、スズメダイの卵の4つを食料として利用しました。
ヒレ咬みの対象となるルリスズメダイやネッタイスズメダイといった小さなスズメダイの密度に、石垣島と瀬底島で有意な差はありませんでした(図1)。イバラカンザシとヒメシャコガイは石垣島では非常にまれで、5平方メートルあたりの数は瀬底島よりも有意に少なくなりました(図1、P=0.002)。魚卵についての量的なデータは得られませんでしたが、瀬底島では多いロクセンスズメダイのような大きなスズメダイの成魚は石垣島ではほとんどいませんでした。

体の大きさと摂食頻度との関係性
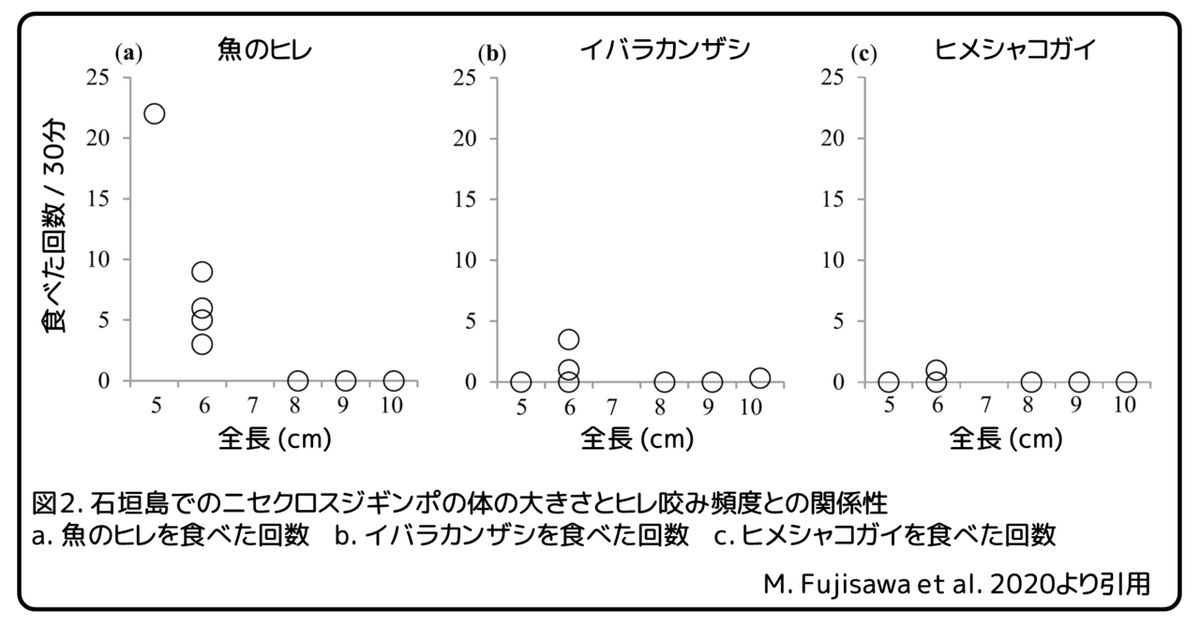
瀬底島と同様に、石垣島では、ヒレ咬みの頻度とニセクロスジギンポの体の大きさとの間には有意な負の相関性がありました(図2a、P=0.016)。卵を食べることは石垣島ではたった1回しか観察されませんでしたので、体の大きさと卵を食べる頻度との間の相関性を解析することは出来ませんでした。瀬底島では、大きなニセクロスジギンポはたびたび集団をつくり、大きなスズメダイの巣を襲いました。石垣島では、5個体の集団を観察しましたが、スズメダイの巣を実際に襲うところは観察されませんでした。石垣島での、イバラカンザシとヒメシャコガイを摂食する頻度は、瀬底島と同様に、ニセクロスジギンポの体の大きさとの間で有意な相関性はみられませんでした(図2b、c)。
摂食頻度の地域による変化
石垣島での、30分あたりの卵を食べる頻度は瀬底島での頻度と有意な差はありませんでした(図3a)。石垣島での、イバラカンザシとヒメシャコガイを食べる頻度は瀬底島での頻度に比べて有意に低くなりました(図3b、c、イバラカンザシ:P=0.0005、ヒメシャコガイ:P=0.003)。

魚のヒレに咬み付く頻度はニセクロスジギンポの体の大きさと負の相関性がありましたので、全長7 cm未満と7 cm以上に分けて比較しました。小さなニセクロスジギンポでは、石垣島の30分あたりのヒレ咬み頻度は瀬底島の頻度よりも有意に高くなりました(図4a、P=0.004)。大きなニセクロスジギンポでは、ヒレ咬み頻度は2つの観察地点で有意な差はありませんでした(図4b)。観察数(3個体)が少ないこともあり、石垣島では、大きなニセクロスジギンポのヒレ咬みは観察されませんでした。

小さいニセクロスジギンポでは、石垣島での総摂食に対するヒレ咬みの割合は瀬底島の頻度よりも有意に高くなりました(図5a、P=0.005)。対照的に、大きなニセクロスジギンポでは、ヒレ咬みの割合は2つの観察地点で有意な差はありませんでした(図5b)。

小さいニセクロスジギンポでは、石垣島での30分あたりのヒレ咬み試行回数も瀬底島での回数よりも有意に多くなりましたが(図6a、P=0.002)、大きなニセクロスジギンポでは有意な差はありませんでした(図6b)。

小さいニセクロスジギンポでは、ヒレ咬み成功率は2つの観察地点で有意な差はありませんでした(図7)。石垣島の大きなニセクロスジギンポのヒレ咬みは観察されませんでしたので、大きなニセクロスジギンポのヒレ咬み成功率は比較できませんでした。

[考察]
本研究から、ニセクロスジギンポの攻撃擬態行動における地域による変化の根底には、他の食料の量があることが明らかになりました。イバラカンザシやヒメシャコガイが少ない石垣島の小さいニセクロスジギンポのヒレ咬み頻度は、これらが豊富な瀬底島のものよりも有意に高くなりました。そのため、フランス領ポリネシアで他の場所よりもヒレ咬み頻度が高いのは、その量について調査はされていませんが、海底食料が不足していたためである可能性も考えられます。
小さいニセクロスジギンポと比べて、大きなニセクロスジギンポでは石垣島と瀬底島でヒレ咬み頻度に有意な差はありませんでした。瀬底島では、小さい時にニセクロスジギンポがたびたびヒレに咬み付くことが報告されています。本研究では、石垣島において同様の傾向がみられました。インドネシアでもニセクロスジギンポの幼若体のみに高いヒレ咬み頻度がみられることが報告されています。そのため、どの生息地でも、ヒレ咬み頻度は成長とともに減少すると考えられます。しかし、瀬底島では、卵を食べる頻度は成長とともに増加しました。石垣島では、おそらく大きなニセクロスジギンポの観察数が少なく、卵を食べる様子は1回だけしか観察されなかったため、そのような傾向をとらえることは出来ませんでした。
石垣島でのイバラカンザシとヒメシャコガイを食べる頻度は瀬底島の頻度よりも非常に低くなりました。これは単純に、石垣島ではこれらの海底食料がほとんどないためです。一方で、ヒレ咬みの対象となる魚の数に違いはありませんでしたが、石垣島の小さいニセクロスジギンポは瀬底島のものよりも頻繁にヒレに咬み付いていました。石垣島の小さいニセクロスジギンポは、イバラカンザシやヒメシャコガイを食べることが出来ないために、ヒレ咬みに依存していると考えられます。以前に、ヒレ咬み頻度は他の食料の量に関連している可能性が示唆されていましたが、本研究によって、このことが初めて確かめられました。
小さいニセクロスジギンポでは、石垣島において、ヒレ咬み頻度だけでなく、ヒレ咬み割合も瀬底島よりも高くなりました。ヒレ咬みの成功率に差は見られませんでしたが、ヒレ咬み試行回数は石垣島で有意に高くなりました。これらの結果は、石垣島の小さいニセクロスジギンポは、その成功率は高くないものの、ヒレ咬みに依存しなければならないという結論を裏付けています。つまり、小さいニセクロスジギンポは、他の食料が少ない時に、攻撃擬態に強く依存しています。以前に、この擬態の主な機能は攻撃擬態ではなく、保護擬態であることが示唆されていましたが、本研究では、特に海底食料が少ない生息地において、攻撃擬態は小さいニセクロスジギンポが生き残るために重要であると結論します。
大きなニセクロスジギンポは、魚卵、イバラカンザシ、ヒメシャコガイを利用することができるのであれば、攻撃擬態に依存する必要はありません。石垣島の大きなニセクロスジギンポの観察数が本研究では少ないため、同じような条件下で攻撃擬態に依存するかどうかを調べるためには、魚のヒレ以外の食料が少ない時の大きなニセクロスジギンポの捕食戦略に注目したさらなる観察が必要です。さらに、この擬態は保護機能を持つことが考えられていますが、それを調査した実験データは報告されていないことから、さらなる研究が必要です。
よろしくお願いします。