プラナリアの咽頭神経 ~食べるか食べないかはのどが決める~ 論文紹介

プラナリアの咽頭神経 ~食べるか食べないかはのどが決める~
論文名 The pharyngeal nervous system orchestrates feeding behavior in planarians
プラナリアでは咽頭神経が摂食行動を調整する
著者名 Mai Miyamoto, Miki Hattori, Kazutaka Hosoda, Mika Sawamoto, Minako Motoishi, Tetsutaro Hayashi, Takeshi Inoue, Yoshihiko Umesono
掲載誌 SCIENCE ADVANCES
掲載年 2020年
リンク https://advances.sciencemag.org/content/6/15/eaaz0882
プラナリアの摂食行動への咽頭神経系の関わりを調べた2020年の論文です。
プラナリアはこれまで主に再生研究のモデル動物でしたが、最近では行動や生殖戦略の研究対象として扱われることが多くなってきたように思います。この論文では、プラナリアの摂食行動に注目しています。
プラナリアはいわゆる脳と呼ばれる神経系があります。この脳は眼の近くにあります。眼は体の移動方向側にあることから、眼のある側が前方と考えられています。つまり、眼と脳が体の前方にあり、いわゆる“頭”があることになります。一般的には頭には口がありますが、プラナリアの口は体の中央部にあり、脳からは離れています。また、エサを食べるときには、口から咽頭(のど)を体外に突出させ、咽頭によってエサを体内に運び込みます。この咽頭は筋肉のかたまりですから、いろいろな方向に自由に動かすことができます。脳と咽頭は神経でつながっていることが分かっています。そこで行われているコミュニケーションについて調べた論文になります。
おもしろいのは、プラナリアのエサの食べ方です。残念ながらプラナリアがエサを食べる様子の動画はこの論文にはありませんが、図1に写真がありますので、そちらを参考にしてください。また、実験では体から切り取った咽頭を使っています。咽頭だけにしてもエサへと移動し、またエサを取り込んでいます。切り取った咽頭はただの管ですから、取り込んだエサは反対側から外に出てしまいます。その様子は論文の動画で見ることができます。ダウンロードしなければなりませんが、ぜひ見てみてください。この動画をみると、咽頭という生物のように見えてきます。咽頭がそれ自体で行動できるということは、咽頭の神経系によってある程度行動の判断を行っていることになります。では、脳にある神経系はどう関わってくるのかという点がこの論文の内容になります。
考察にて議論されていますが、プラナリアが含まれる扁形動物の祖先型では、口の位置が前方の脳に近いところにあったようですが、進化とともに後方へと移動したようです。どうして口が移動したのか、口の移動によって何が起こったのか、その進化上の利点は何か、などボディプランの進化についても興味がそそられます。
*同じ研究グループによるプラナリアの摂食行動についての別の論文はこちらで紹介しています
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
・ナミウズムシの咽頭はそれ自体でエサへ移動し取り込む。
・ウコンが含まれるエサの場合は、咽頭はエサへ移動するが取り込むことはない。
・咽頭のエサへの移動と取り込みはオクトパミン作動性神経とコリン作動性神経により制御されている。
・咽頭がウコンを取り込まない摂食拒否はドーパミン作動性神経とセロトニン作動性神経により制御されている。
・咽頭のオクトパミン作動性神経は個体がエサへ移動するために必要となる
[背景]
左右相称動物は、形態や行動が多様な膨大な数の種からなり、特定の環境下での生存と繁殖を可能にしています。淡水プラナリアのナミウズムシは、体の前端に脳と光感覚のための眼と化学感覚のための外耳を持つ頭化形質を持っています。(補足:頭化とは体の前端部に神経組織や感覚器が集中すること。)これとは対照的に、その摂食器官(口と咽頭)は前端の頭部からかなり離れた体の中央部に位置しています。プラナリアがどのようにしてエサを摂取するかを明らかにすることは、独特な形態学的表現型と行動の間をつなぐ神経機構を同定することに繋がります。
プラナリアの摂食行動は、複雑な一連の運動から構成されています。最初に、プラナリアの個体はエサへ向かって移動しそこに留まります(エサ局在運動)。次に、エサに反応した口から咽頭を突出します(目標指向性突出運動)。最後に、咽頭の先端開口部から腸へ向かってエサを積極的に取り込みます(図1A、B)。これまでの研究から、エサ局在運動を含む多くのプラナリアの行動を実行するための感覚情報の統合と処理において、脳が中心的役割を果たしていることが分かりました。しかし、化学感覚シグナルに応答した咽頭の目標指向性突出運動の調節における脳の役割はまだ分かっていません。特に、プラナリアの咽頭は主に筋細胞から構成されており、摂食時には柔軟な動き(伸縮し、あらゆる方向に曲がる)をすることから、タコやイカといった頭足類の触手やゾウの鼻と同様の骨格のサポートがない筋肉の一例であると考えられます。体の中央部である体幹に位置する咽頭が、エサ局在運動中に適切なエサの検出と取り込みを安定して行うことから、咽頭神経系が以前考えられていたより大きな機能的能力を持っていると考えられます。
本研究では、ナミウズムシを用いて、咽頭神経系が独特な形態学的表現型と摂食行動の間の繋がりを確立する機能的モジュールとして働くことを示しました。これは、左右相称動物が進化の間に形態学的および行動的な多様性をどのように増加させるかについて理解を深めることにつながります。

[結果]
咽頭の摂食プログラムは咽頭自身によって生み出されている
ナミウズムシの体から切り取った咽頭の行動を、目標物となるエサの条件を変えながら45秒間記録しました。エサにすりつぶした肝臓が入っていない場合は、切り取られた咽頭は、先端側を前方にして這って動きましたが、エサに決して触れることなく、摂食することはありませんでした(図1C、F、I、動画1)。(補足:研究室で飼育されているナミウズムシは鶏の肝臓をすりつぶしたものをエサとして与えられている。先端側は切り取られる前は体と反対側にあった側。)エサにすりつぶした肝臓が入っている場合は、切り取られた咽頭はエサに向かって移動し、再現性高く先端側を広げエサを取り込みました(図1D、F、I、動画2)。量的な解析によって、プラナリアの咽頭は、エサがあってもなくても、一定の速度で移動する独自の活性を持つことが分かりました(図1H)。リュウキュウナミウズムシの切り取られた咽頭も、同じ条件でエサを摂食することに成功しましたが、アメリカナミウズムシの咽頭はエサへ向かう移動能力が乏しく、摂食することはできなかったことから、同じナミウズムシ属の間でも行動特性が違うことは注目すべきです。
次に、切り取られた咽頭が適切なエサを選択できるかどうかを調べました。カレーのメインスパイスとして知られるウコンは、日本では着物の防虫剤として使われてきました。プラナリアにすりつぶした肝臓とウコンを混ぜたエサ(ウコン濃度は62.5 μg/μL)を与えたところ、強い摂食拒否を示し、決してそのエサを食べませんでした。ウコンの混ざったエサに対しては、ウコンの量を半分に減らしても、切り取られた咽頭も強い摂食拒否を示しました(図1E、I、動画3)。一方で、このエサへの走化性は常に見られました(図1F、G、動画3)。このことから、プラナリアの咽頭がすりつぶした肝臓を感知していることが分かります。これらの観察は、咽頭それ自身が異なった化学刺激を同時に感知し、エサを食べるかどうかについて決定していることを示しています。
咽頭神経系は咽頭の摂食行動に必要である
次に、切り取られた咽頭の摂食行動の制御における咽頭神経系の役割を調べました。咽頭神経系は先端基部軸に沿って偏った組織を持っており、主要神経輪はエサの取り込みに使われる先端側に位置しています(図2)。この解剖学的特徴はプラナリアの他種にも保存されています。化学感覚機能は咽頭の先端にあります。切り取られた咽頭を先端基部軸に沿って二つに切断した場合、主要神経輪を含む先端側はすりつぶした肝臓に反応して完全は摂食を行うだけでなく、ウコンによる摂食拒否も示しました(図3)。対照的に、主要神経輪を含まない基部側はエサに向かった移動といったものは見られませんでした(図3)。
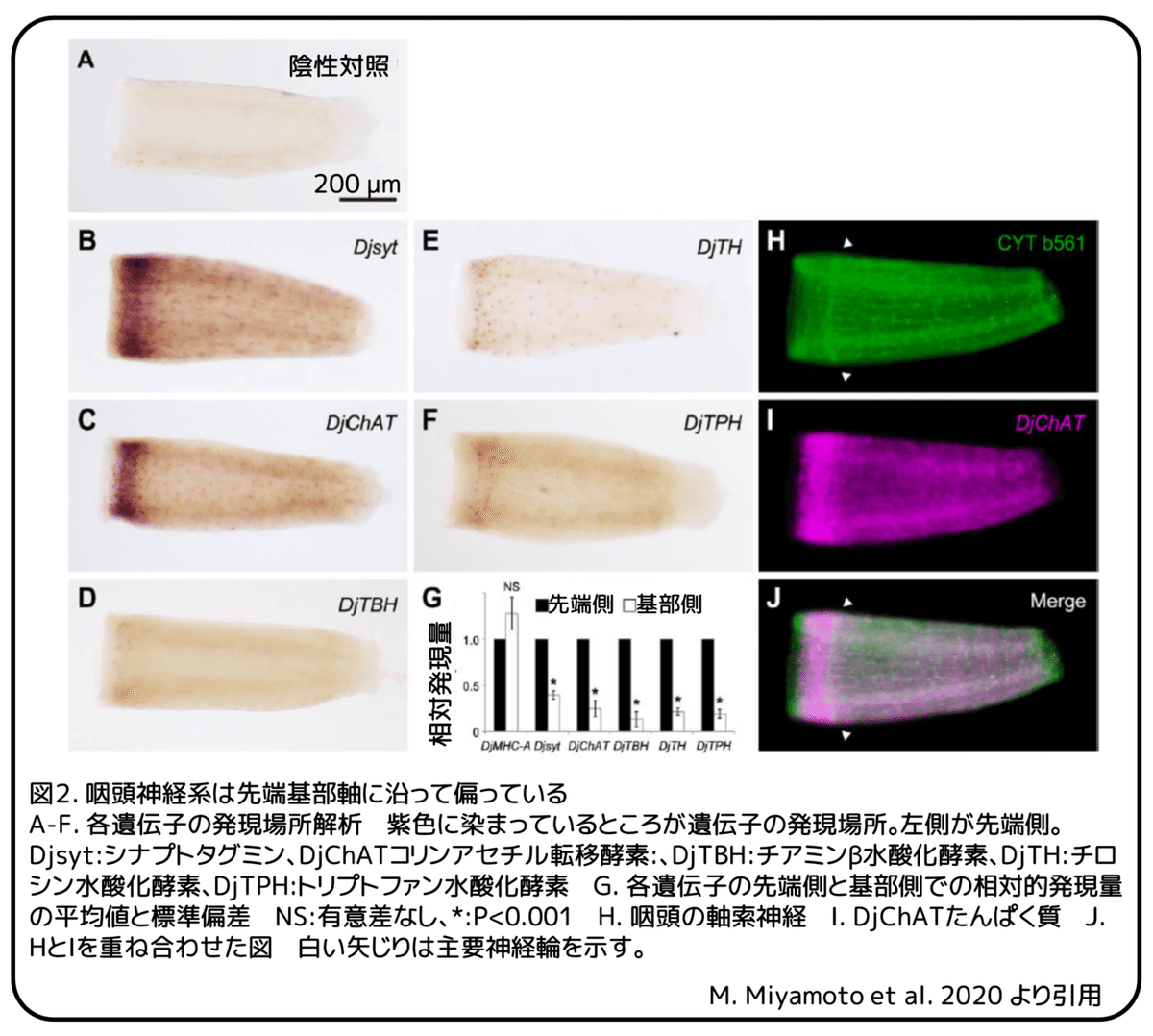

続いて、シナプス小胞に発現し、神経伝達物質の放出を引き起こす膜たんぱく質をコードするシナプトタグミン(Djsyt)遺伝子を咽頭神経系において再生依存的に遺伝子ノックダウンを行いました。シナプトタグミン遺伝子に対するRNA干渉処理をした後に咽頭全体を再生させることによって、咽頭でのシナプトタグミンたんぱく質の発現が大きく減少しました。おそらく咽頭の運動機能の異常のために、切り取られた咽頭はスタート地点から移動できませんでした。(補足:再生によって、咽頭ではシナプトタグミンたんぱく質が作られなくなった。RNA干渉については図4参照。)対照的に、咽頭の先端側半分の再生によって、咽頭先端部ではシナプトタグミンたんぱく質の発現が大きく減少しましたが、再生していない基部側では変化はありませんでした(図5A、C)。この対照的な効果は、未分化な幹細胞からあたらしく再生した神経では、半減期が長いシナプトタグミンたんぱく質を多量に持つ、すでに最終分化している神経よりもRNA干渉がより効果的であるためです。(補足:神経分化によってシナプトタグミンたんぱく質が新しく作られる際にRNA干渉が大きく影響するため。)注目すべきは、これらのシナプトタグミン遺伝子をノックダウンされ、切り出された咽頭が、正常な運動機能を持ちながらもエサを見つけることができなかったことです(図5D、E、G)。これらの発見から、咽頭神経系の先端側半分における特定の神経伝達物質によるシナプス伝達が咽頭の摂食機能の制御において司令的な機能を持っていることが分かりました。


特異的神経伝達物質の生合成に関わる様々な律速酵素遺伝子の発現場所解析と定量的PCR解析によって、少なくとも、コリン、オクトパミン、ドーパミン、セロトニンの4つのモノアミン作動性神経が、咽頭の先端側に集中していることが明らかになりました。咽頭神経系における、コリンアセチル転移酵素(DjChAT)遺伝子に対するRNA干渉によるコリン作動性シグナルの機能不全または、チアミンβ水酸化酵素(DjTBH)遺伝子対するRNA干渉によるオクトパミン作動性シグナルの機能不全によって、切り出された咽頭のエサ方向への移動が見られなくなりました(図5D、G)。対照的に、咽頭神経系における、チロシン水酸化酵素(DjTH)遺伝子に対するRNA干渉によるドーパミン作動性シグナルの機能不全または、トリプトファン水酸化酵素(DjTPH)遺伝子対するRNA干渉によるセロトニン作動性シグナルの機能不全では、切り出された咽頭はgfp遺伝子に対するRNA干渉を行った対照群と同程度の完全な摂食機能を持ち、エサの取り込みを行いました。摂食拒否実験では、咽頭神経系においてドーパミン作動性シグナルまたは、セロトニン作動性シグナルが機能不全である切り出された咽頭は、予想通りウコンとすりつぶした肝臓を混ぜ合わせたエサへの走化性を見せましたが、摂食拒否については明らかな異常を見せました(図6A-E)。これらの咽頭は、対照群の咽頭よりも積極的にウコンを含むエサを取り込みました(図6B、C、F)。

咽頭のオクトパミン作動性シグナルは個体の摂食行動に必要である
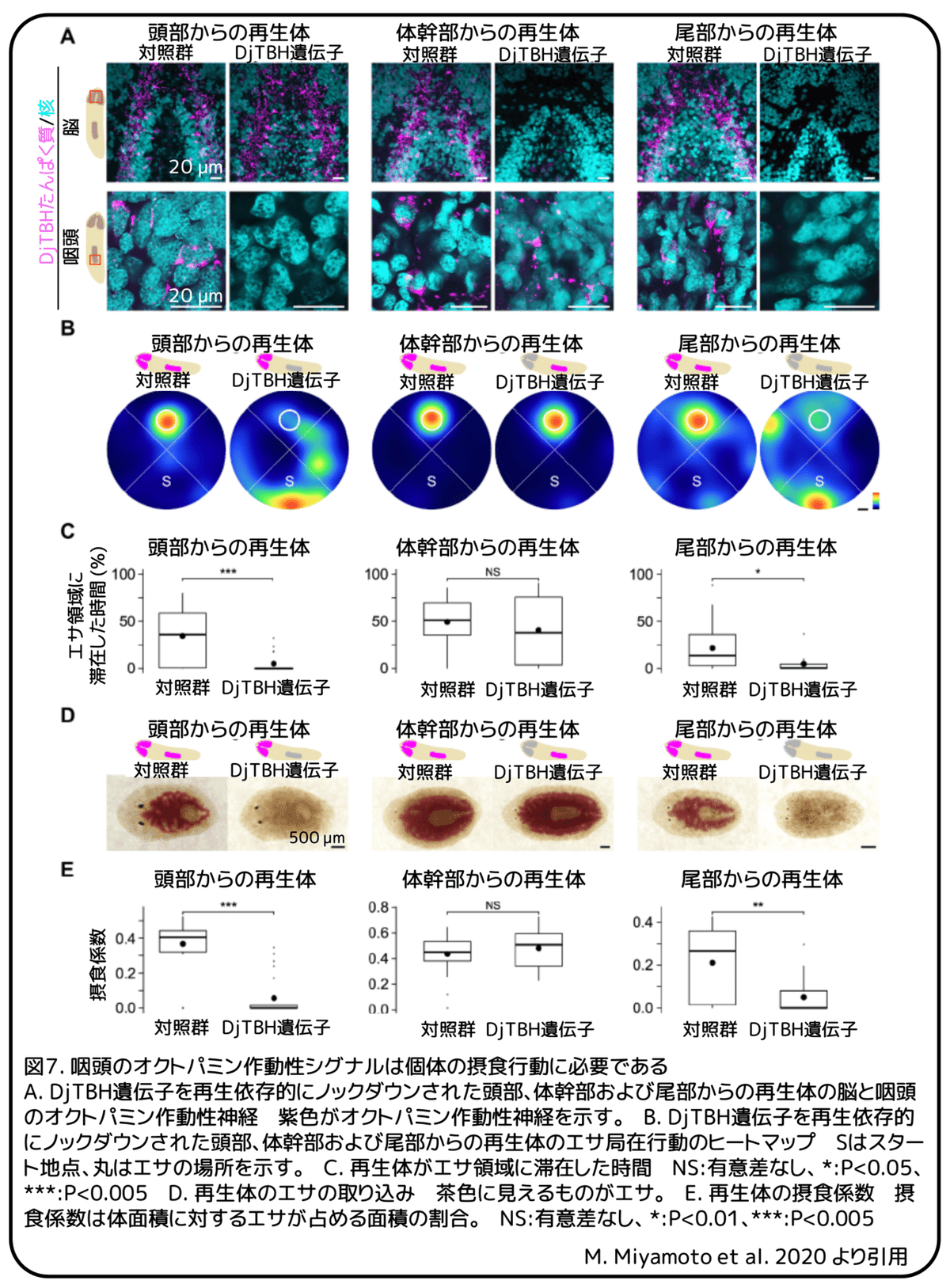
最後に、個体の摂食行動における咽頭のオクトパミン作動性シグナルの役割についてDjTBH遺伝子に対する再生依存的遺伝子ノックダウン実験によって調べました。DjTBH遺伝子または対照群としてgfp遺伝子に対するRNA干渉処理をされたプラナリアを、頭部、咽頭を含む体幹部、尾部の3つに切断し、通常の条件で摂食機能が回復する十分な期間である10日間をかけて再生させました。抗DjTBHたんぱく質抗体による染色によると、対照群においてDjTBHたんぱく質は主に、脳と咽頭の2つの器官に発現していました(図7A)。DjTBH遺伝子に対するRNA干渉処理をした場合、頭部から再生したプラナリアでは、脳に変化は見られませんでしたが、咽頭のDjTBHたんぱく質が選択的に消失しました(図7A)。一方で、体幹部から再生したプラナリアでは、咽頭に変化は見られませんでしたが、脳のDjTBHたんぱく質が選択的に消失しました(図7A)。そして、尾部から再生したプラナリアでは、咽頭と脳のDjTBHたんぱく質が消失しました(図7A)。対照群では、3つの断片からの再生体全てで、エサ局在運動、咽頭の突出、エサの取り込みという全ての摂食運動が回復していることを確かめました(図7B-E)。DjTBH遺伝子に対するRNA干渉処理をし、体幹部から再生した個体は、対照群の体幹部からの再生体と同程度の摂食行動を示したことから、脳のオクトパミン作動性シグナルは摂食行動に重要ではないと考えられます(図7B、C)。一方で、DjTBH遺伝子に対するRNA干渉処理をし、頭部から再生した個体は、エサ局在運動に大きな異常がみられ、その結果エサを取り込むことができませんでした(図7D、E)。DjTBH遺伝子に対するRNA干渉処理をし、尾部から再生した個体でも、頭部から再生した個体と同様の異常がみられました(図7B-E)。量的な解析を行いましたが、頭部と尾部からの再生体の間に明らかな違いないことから、脳でのオクトパミン作動性シグナルの消失は行動異常に対して何も影響していない可能性が示されました(図7C、E)。まとめると、咽頭のオクトパミン作動性シグナルは個体のエサ局在運動に必要です。この意外な発見から、プラナリアの咽頭が個体としての摂食行動を統合するために脳とコミュニケーションしている可能性が考えられます。(補足:咽頭からの情報がオクトパミン作動性シグナルによって脳に伝わり、脳からはオクトパミン作動性シグナル以外のシグナルによって個体としての行動につなげている。)
[考察]
本研究では、プラナリアの切り出された咽頭の行動評価システムを構築し、中枢神経系から独立した咽頭の感覚および行動を明らかにしました。この発見は、コリン、オクトパミン、ドーパミン、セロトニンの4つの異なった作動性神経が、摂食誘引と摂食拒否という2つの行動反応に必要であること示しました(図8A)。また、これらの結果から、プラナリアの咽頭が、個体がエサを取り込むために適切な位置取りを行うエサ局在運動のために脳へ司令を送っている可能性が考えられます(図8B)。これまでの標識追跡実験から、特定の咽頭神経の長く伸びた軸索が直接的に脳と接続していることが分かっています。咽頭のオクトパミン作動性神経はそのような神経の有力な候補の一つです。

脳がエサに向けた咽頭突出運動の制御に関連している可能性を排除することはできません。この可能性は、頭を切り落とされたナミウズムシをエサのそばにおいても咽頭を突出させないことから考えられています。しかし、この現象を説明する別の考えがあります。それは、脳はエサに反応して咽頭を突出するために口をあけるシグナルを送っている可能性です。この可能性について、咽頭がエサを感知したときに、脳に対して口を開けるように司令を送っているかもしれません。これらの仮説については今後研究で検証していきます。
本研究結果は、前後軸に沿って様々な位置に摂食器官をもつ扁形動物門内の形態的および行動的多様性の進化に関して重要な意味も持っています。進化的に最も早く枝分かれした小鎖状目や多食目は摂食器官である口と突出しない咽頭を前端近くに持っています。プラナリアにつながる扁形動物系統における摂食器官の先祖型の前方位置から後方への移動は、個体をエサへと向かわせる運動を指揮する感覚―中枢神経の回路から、咽頭の感覚と行動を独立させる圧力となった可能性が推測されます。摂食行動を制御するための咽頭神経系と中枢神経系の分業の確立は、扁形動物のボディプランの大きな変化に関連して進化的適応性を高めた可能性があります。
よろしくお願いします。