鱗食魚の経験と学習 ~若い頃しか学べない~ 論文紹介

鱗食魚の経験と学習 ~若い頃しか学べない~
論文名 Experience‑dependent learning of behavioral laterality in the scale‑eating cichlid Perissodus microlepis occurs during the early developmental stage
鱗食魚ペリソダスの行動左右差の経験依存的学習は初期発育期間に起こる
著者名 Yuichi Takeuchi, Yuna Higuchi, Koki Ikeya, Masataka Tagami & Yoichi Oda
掲載誌 Scientific reports
掲載年 2022年
リンク https://doi.org/10.1038/s41598-021-04588-8
鱗を食べる魚ペリソダスの行動左右差の学習を成長段階毎に調べた2022年の論文です。
ペリソダス(Perissodus microlepis、和名がないため、以下便宜的にペリソダスとします。)はアフリカのタンガニーカ湖に生息する他の魚の鱗を食べる魚です。他の魚の鱗を奪う時には、獲物の右または左側から近づく必要がありますが、その行動が左右のどちらかに偏っていることがこの魚では知られています。また、口の形が左右非対称で左側が大きくなっている場合は「左利き」で左側から攻撃する、右側が大きくなっている場合は「右利き」で右側から攻撃することが分かっています(漫画「ペリソダス」参照)。口の形は遺伝的に決まっているようで、「左利き」と「右利き」は1対1になるようです。この行動の左右差は成長とともにどのように獲得されるのか、について本論文では調べています。
「背景」にあるように、行動の左右差は、色々な動物で知られています(漫画「左右の好み ~利き〇〇~」参照)。特にヒトでは利き手や利き足があることは誰でも知っていると思いますが、それらがどのようにして形成、獲得されるのかと言うのははっきりと分かっていません。ヒトの場合は一定の条件を維持しながら長期間観察することは困難です。そこで他の動物を使った研究が行われています。
本論文では、経験が行動左右差の学習に影響を与えるかどうかを3つの成長段階について調べ、成長のどの段階での経験がその後の行動にどのような影響を与えるかを明らかにしています。本研究で分かったことから、脳内での神経回路形成の仕方や領域が成長段階によって異なることが考えられます。そのあたりについては本論文では触れられていませんが、今後の研究で明らかになる可能性があります。成長段階ごとに適した学習をすることの意味やそのメカニズムなど非常に興味深いです。
行動左右差については以前にスナメリの行動について紹介しています。よろしければそちらも読んでみて下さい。
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
ペリソダスは幼魚から成魚まで、鱗食の経験がなくても鱗食を行う動機を保持しており、幼魚と若魚では、鱗食経験によりその動機は強化される。
幼魚は生来の利き側からの攻撃側選好性を持っており、鱗食経験により強化されるが、経験が無ければ生来の攻撃側選好性は発育とともに徐々に失われ、成魚では消失してしまう。
鱗食経験によって幼魚は攻撃時の体の屈曲動作を、若魚は接近速度を改善し、捕食成功率を上げる。
[背景]
ヒトの身体はほぼ左右対称ですが、手仕事のように高度で力強い作業を行う場合には、身体の片側を使うことを好みます。行動する際に体の片側を他方よりも好む"行動左右差 "は、チンパンジーやカラスの道具使用時の好み、マウスやヒキガエルに見られる利き足、魚類や頭足類の利き目など、動物界を通して観察されています。行動左右差はどのようにして生じるのでしょうか?ヒトの利き手は、遺伝的要因と環境要因の複雑な相互作用によるものであると考えられています。利き手は胎内から観察されており、小児期に発達すると考えられています。しかし、ヒトの利き手の発達過程を研究することは、長期間の観察が困難であること、実験の目的のために身体に処理を施すことができないこと、あるいは社会的制約(例えば、左利きの子供が右手で文字を書くことを強いられるケースなど)により、これまで困難でした。動物を用いた研究では、出生前後の影響が脳や視覚の左右差に見られる家畜のニワトリを除いては、成体になってからの行動の記述に限定されています。現在までに、動物の利き手を引き起こす要因や、生涯を通じてどのように行動左右差が生じるのかについては、ほとんど分かっていません。
魚類では、捕食行動や逃避行動、同種間の反応に行動左右差があることが報告されています。行動左右差の顕著な例として、タンガニーカ湖の鱗食性シクリッド魚ペリソダス(Perissodus microlepis)の「左利き」「右利き」が挙げられます。(補足:左利きは利き側が左になる。)本種はタンガニーカ湖に豊富に広く分布し、斜めにねじれた形態を持つ口で他の魚の鱗を主に食べることに特化した魚です。成魚は、獲物である魚の横腹の鱗を食べるために、利き側から攻撃します。左利き(個体数の約50%)は左側の下顎骨が大きく、口をやや右側に開き、獲物の左側の横腹を攻撃しますが、右利きの魚は逆の形態と行動を示します。この口の形態の左右差は、遺伝的に決定されていると考えられています。この左右差のある捕食行動の発達を探るこれまでの研究では、以下の知見が報告されています。 まず、タンガニーカ湖でのペリソダスの胃内容物と口の形態の大規模解析から、幼魚は35-45mmの標準体長で鱗食を始め、成長とともに徐々に攻撃の利き側が発達することが明らかとなりました。また、標準体長が 80 mmより大きい大型成魚は鱗のみを食べ、利き側から捕食者を攻撃する行動が定着していました。次に、標準体長が約38-48mmに達する生後4ヶ月まで顆粒餌で人工飼育され、鱗食経験のない未経験幼魚は、最初の捕食攻撃で、利き側から獲物を攻撃する弱い選好性を示しましたが、鱗食経験の増加とともに、利き側から攻撃して効率的に鱗を剥がす能力を向上させました。
本研究では、タンガニーカ湖のペリソダスの行動左右差は、ある特定の発育段階でのみ確立するのか、それとも学習する機会があれば成体を含むどの発育段階でも確立するのかを明らかにしました。鳴禽類であるキンカチョウやミヤマシトドは発育初期の短い特定の時期にしか歌を覚えられませんが、ホシムクドリやサヨナキドリは発育後期まで歌を覚えることができることが報告されています。このように、自然環境における発育過程での動物の行動学習は、生活史における行動経験と密接に関連している可能性があります。
行動左右差の確立過程を明らかにするために、ペリソダスの個体が鱗食を開始する「幼魚期」(発育初期、標準体長が 50 mm以下)、プランクトンと鱗食の両方を食べる混合食の「若魚期」(標準体長が51 mm以上 64 mm以下)、鱗食が確立した「成魚期」(標準体長65 mm以上)の3つの発育段階(図1a)で鱗食体験が行動左右差の確立に与える影響を比較検討しました。3つの発達段階は、る野生魚の3つの食餌段階におけ標準体長に対応して決定されました。成魚期の定義は、シクリッドの体長と性腺の重さの相関関係に基づいて行いました。生後4ヶ月(幼魚)、8ヶ月(若魚)、12ヶ月(成魚)の3つの発育段階まで顆粒餌のみを与えた鱗食未経験ペリソダスを使用しました。 (1)3つの発育段階の未経験ペリソダスは、捕食の経験がなくても鱗食の動機を保持しているか、(2)経験とともに鱗食行動が発達するか、(3)口の形態に応じた利き側からの攻撃を学習するか、(4)捕食経験によって捕食運動は動力学的に発達するか(体の屈伸角とその速度、餌への接近速度)、について明らかにするために捕食実験を行いました。
[結果]
研究室でふ化したペリソダスは、攻撃実験を行うまで個別の水槽で顆粒餌を与えて飼育しました(図1a)。3つの発育段階の未経験ペリソダスは、実験水槽に獲物である金魚を一緒に入れられると、金魚を自分から攻撃しその鱗を捕食しました(図1b)。未経験ペリソダスはセッション1、つまり生涯で初めて遭遇した時から、獲物である金魚に攻撃を行いました。(補足:1セッションは1時間の攻撃実験で、本研究では2、3日おきにセッションを行い合計5回のセッションを行った。)このことから、ペリソダスは獲物である魚に攻撃を行う生来の選好性があることが分かりました。しかし、最初のセッションでの攻撃頻度は発育段階によって異なりました。統計解析によって、3つの発育段階の間に有意な差が検出されました(P=0.013)。1時間のセッション中の攻撃回数は若魚と成魚よりも幼魚で優位に多くなりました(図1c)。セッションを繰り返した後は、幼魚と若魚は獲物に気がつくとすぐに攻撃を始めました。ペリソダスのいる水槽に獲物の金魚を入れた後に、ペリソダスが最初の攻撃をするまでの時間は、幼魚と若魚ではセッション1よりセッション5で優位に短くなりましたが、成魚ではそうではありませんでした(図1d)。このことから、成魚ではなく幼魚と若魚は経験によって攻撃のための動機が高くなること分かりました。

攻撃行動の左右差を調べるために、攻撃を仕掛ける方向が口の形態の利き側と一致しているかどうかを確かめました(図2a)。セッション1では、未経験の幼魚は偶然から予想される確率よりも頻度高く利き側から攻撃を行いました(図2b)。(補足:利き側とその反対側の2通りと考えると、利き側からの攻撃確率は50%と予想される。)対照的に、未経験の若魚と成魚はセッション1でそのような選好性を示しませんでした(図2b)。この結果から、利き側から攻撃を行う生来の選好性を幼魚は持っており、それは成長とともに徐々に失われ鱗食の経験のない成魚では最終的に消失してしまうと考えられます。
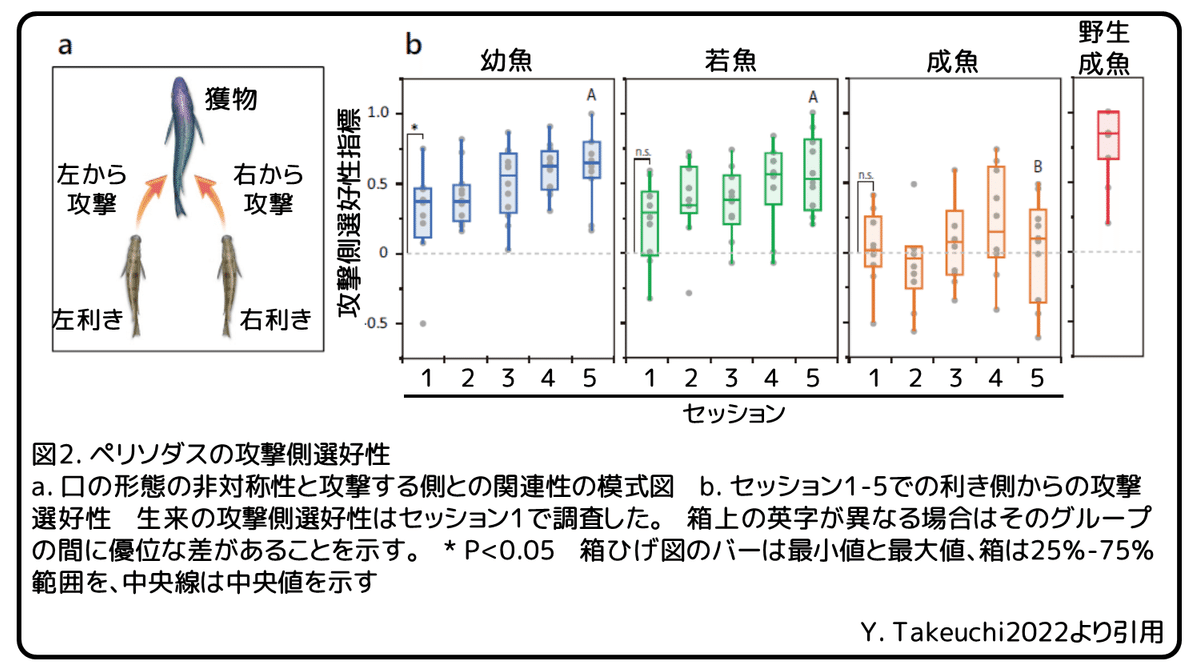
次に、セッション中に鱗食を経験するに伴い行動左右差が発達するかどうか、またどのように発達するのかを調べるために、攻撃側選好性指標(Index of Attack side Preference, IAP)を測定しました。(補足:IAPは攻撃側が利き側と反対側が同数であれば0、利き側が多い場合は正、反対側が多くなれば負の値に、全ての攻撃側が利き側であった場合は1になる。)約2週間の間に行われた計5回のセッションを通して、幼魚と若魚は攻撃側選好性を発達させました(図2b)。もし、もう少しセッションを続けていたならば、これらの攻撃側選好性は野生成魚と同程度になったと推測されます(図2b、野生成魚)。対照的に、未経験の成魚は実験期間を通して攻撃側選好性の発達は見られず、セッション5での成魚の攻撃側選好性指標は、幼魚と若魚よりも有意に小さくなりました。さらに、各発育段階での個体レベルの解析を行いました。幼魚では、12匹中の2匹がセッション1で利き側からの有意な攻撃側選好性を示しました(図3a、J1とJ3)。セッション1での集団レベルでの解析と併せると、ほとんどの個体は有意な左右差を示しませんでしたが、集団としての攻撃方向は主に利き側である可能性が考えられます。5回のセッションを終えて、有意な行動左右差を示す個体の数は増加し、セッション5では、ほとんどの個体が行動左右差を獲得しました(図3a、b)。全ての場合で、攻撃側選好性は非対称な口の形態よって決定される利き側と一致していました。若魚では、10匹中の1匹だけがセッション1で利き側からの有意な攻撃側選好性を示しました(図3a、Y10)。5回のセッションを終えて、有意な行動左右差を示す個体の数は増加しましたが、セッション5では、有意な行動左右差を示したのは半数だけでした(図3a、b)。つまり、有意な攻撃側の偏りの発達が幼魚の80%以上で見られたのに対し、若魚では半数にとどまったことから、これら2つの発育段階で学習効率が異なる可能性が示されました。幼魚や若魚とは対照的に、セッション中に攻撃側選好性を見せた未経験の成魚はほとんどいませんでした(図3a)。たった1匹の成魚だけがセッション4で有意な攻撃側選好性を示しましたが、この選好性はセッション5では見られませんでした。これらの結果から、経験を通して行動左右差を獲得する能力は一般的に鱗食行動を開始すると考えられているふ化後4ヶ月(幼魚)で高いが、ふ化後8ヶ月(若魚)までに鱗食の経験をすることがなければ、徐々に失われ、ふ化後12ヶ月(成魚)までに完全に消失してしまうと考えられます。

続けて、攻撃の成功に関わる左右差のある行動をどのように学習するのかを調べました。予想外に、幼魚と同様に若魚でも経験とともに攻撃成功率が上昇することが分かりました(図4a)。一方で、成魚では、セッション2で一過的な成功率の上昇が見られましたが、セッション1とセッション5で成功率に有意な変化は見られませんでした(図4a)。セッション5では、成魚の攻撃成功率は幼魚や若魚よりも有意に低くなりました。そこで、行動左右差を学習することにより攻撃が成功するのかどうかを、利き側からの攻撃と反利き側からの攻撃の成功率を比較することで調べました。統計解析によって、全てのセッションを通して、幼魚では利き側からの攻撃成功率が反利き側からのものよりも有意に高いことが分かりました(図4b)。しかし、若魚と成魚では成功率に有意な差はありませんでした。これらの結果から、幼魚はわずかにねじれた口で獲物の鱗を効率的の捕食するために利き側からの攻撃を学習しますが、若魚は攻撃を改善するために異なる戦術を学習する可能性が明らかになりました。

幼魚と若魚の攻撃の違いをさらに調べるために、ハイスピードビデオカメラを使って、攻撃行動の運動を詳細に解析しました。最初に、攻撃中の体の屈曲角に注目しました。これは野生成魚の利き側と反利き側からの攻撃で異なることが分かっています(図5a)。幼魚では、セッション1とセッション5の両方で、利き側からの攻撃時の屈曲角が反利き側よりも有意に大きくなりました(図5b)。(補足:屈曲角は口吻端・重心・尾ヒレの3点をつないだ線によってなす角度を180度から引いた値であると思われる。)その差はセッション1からセッション5までに大きくなりました。同様に、幼魚の最大角速度もセッション1とセッション5の両方で、利き側で大きくなりました(図5c)。しかし、若魚と成魚では、セッション1でもセッション5でも、利き側と反利き側との間で屈曲角と角速度に差は見られませんでした。これらの結果から、鱗食行動中の効率的な攻撃のための屈曲の左右差は、幼魚期の経験を通して確立されることが分かりました。
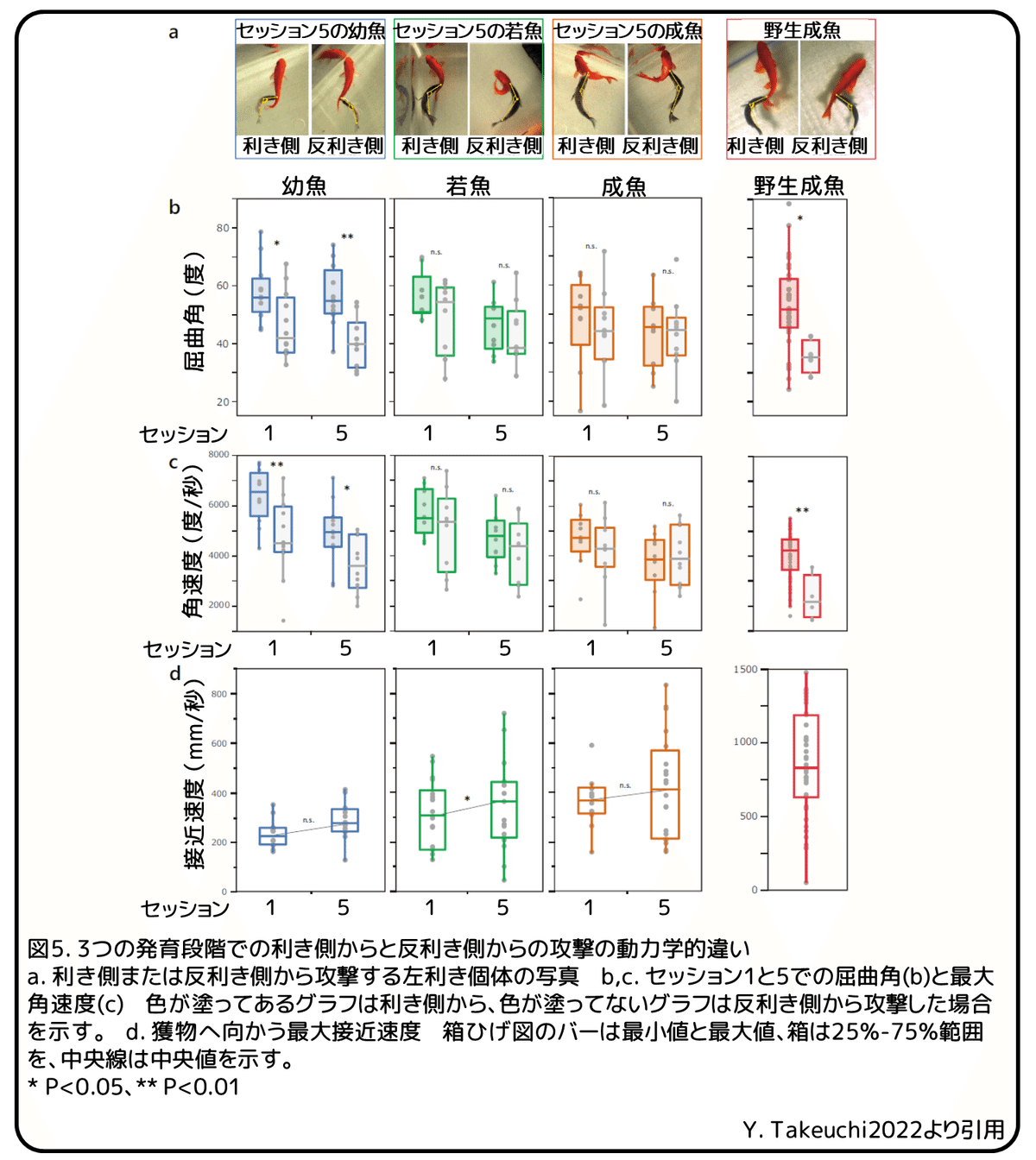
さらに、追跡はじめから鱗の捕食終了までの一連の攻撃行動を観察することによって、セッション中に獲物に近づくときの速いスピードを若魚は発達させていることが分かりました。セッション5では、若魚の最大接近速度はセッション1よりも有意に速くなりましたが、左右差はありませんでした(図5d)。しかし、幼魚と成魚は接近速度のそのような変化は示しませんでした。そのため、若魚はその左右差に関係なく接近速度を増加させることを学習し、鱗食効率を改善しましたが、幼魚も成魚もそのような経験に依存した接近時の速度上昇を獲得しませんでした。これらの結果から、攻撃中の屈曲角の左右差と攻撃側選好性は、初期発育段階での鱗食経験を通して最初に獲得、発達すると考えられます。さらに、ペリソダスは若魚期に接近速度を上昇させることを学びました。野生成魚は、それぞれの発育段階を通して、利き側からの接近速度と屈曲角の両方を恐らく改善します。これらの技術は獲物の鱗捕食の成功に貢献しているように見えます。しかし、運動能に関する生来の左右差と学習能力は、成長期に鱗食経験を持たないと失われてしまう可能性があります。
[考察]
本研究は、左右差をもった攻撃行動を示す鱗食魚ペリソダスの行動左右差の獲得に対する鱗食経験の役割と発育段階依存的学習の重要性を明らかにすることを目的としました。行動実験では最初に、発育段階に関係なく、ペリソダスはこれまでに一度も遭遇したことのない金魚をすぐに攻撃することが分かりました(図1b)。鱗食をはじめる時期である未経験の幼魚(ふ化後4ヶ月)は鱗を捕食するための生来の強い動機を持ち、短い期間のセッション中の経験を通して、その動機を強くしました。未経験の若魚(ふ化後8ヶ月)は、当初は攻撃意欲が低下していましたが、経験によって、攻撃的になることを学習しました。未経験の成魚(ふ化後12ヶ月)は獲物を攻撃しようとしましたが、経験によってその行動は発達しませんでした(図1c、d)。2番目に、未経験の幼魚は攻撃方向の生来の選好性と、利き側でのより強力な行動を示しました(図2b、5b、c)。幼魚は利き側からの攻撃による鱗食の経験によって、攻撃側選好性を強化し、屈曲動作を発達させ、攻撃成功率を徐々に上昇させました(図4a、5b)。3番目に、未経験の若魚は生来の左右差を失いましたが、いくつかの個体では鱗食経験を通してそれを徐々に発達させました(図2b、3b、4a)。さらに、若魚のほとんどは攻撃を成功させるために獲物に素早く近づくための別の技術を発達させました(図5d)。幼魚や若魚と対照的に、未経験の成魚は左右差のある動作を発達させることはなく、利き側でより高い成功率を示すこともありませんでした。そのため、鱗食行動は幼魚から成魚までの全ての発育段階で備わっているようですが、鱗食に対する動機と攻撃側選好性の発達は発育段階に依存していました。初期発育段階での経験はその後の発育段階の攻撃行動に影響する可能性があります。特に、幼魚と若魚は短い期間の鱗食経験によって自身の攻撃技術を改善したことから、各発育段階のわずかな経験によって、ペリソダスは攻撃行動の重要な動作を学習し獲得しました。攻撃中の左右差のある屈曲動作が鱗食を始めた直後である幼魚期で学習されることは、強調すべきことです。
タンガニーカ湖のペリソダスの生活史についてこれまでの研究では、稚魚は群れをなして動物プランクトンを食べ、標準体長が約45 mmの幼魚は群から離れ、徐々にその食性を鱗へと移行していくことが分かっています。本研究で使用した幼魚は、最初のセッションで偶然に予想される確率よりも頻度高く利き側から攻撃を行いました(図2b)。わずかにねじれた口で鱗を捕食するときに、予め左右差のある攻撃はある程度有利であり、鱗への食性移行を加速させる可能性があります。付け加えて、口の形態的非対称性も、鱗食経験の結果として発達する可能性があります。特に、野生の個体は人工的に飼育した個体に比べ、より非対称な口の形態を見せます。これは、野生個体が研究室の水槽で育った個体よりも獲物を攻撃する機会が多いことが理由であるように見えます。口形態の必然的な変化は、利き側からの攻撃を増やすことに貢献している可能性があります。
幼魚と若魚は鱗食経験によって行動の異なった側面を学習します。より早い発育段階は左右差のある攻撃行動を学習することに適しており、中期発育段階は追跡のための速度を向上させることに適しています。注目すべきことは、鱗食経験によって、幼魚は攻撃側選好性を強化するだけでなく、利き側からの攻撃中の屈曲動作も発達させました。特に、幼魚は鱗食の開始時期に生来の左右差のある捕食行動を発達させます。しかし、その期間に鱗食の機会がなければ、生来の左右差は徐々に失われます。自然環境では、捕食成功率を最大化させ、その行動によるコストとリスクを最小化するために、ペリソダスは異なる発育段階で捕食行動の異なる面を学習している可能性があります(図5b、c、d)。本研究では、幼魚と若魚の攻撃方向の学習能力の差について慎重な結論を与えました。二項検定の結果から、幼魚は若魚よりも攻撃方向の学習に適していると考えられます(図3b)。しかし、二項検定の結果は集団の数が多くなるほど有意となる傾向があるため、これらの結果は注意深く解釈する必要があります。(補足:図3aでは二項検定によって有意であるかを調べているが、攻撃回数が多い個体で有意差が出やすくなるため、有意差のある個体数を示した図3bの解釈には注意が必要となる。)そのため、幼魚と若魚の間の左右差をもった攻撃のための学習能力の差は今後さらに研究される必要があります。
発育期間の中で、学習のための特異的な期間は「感受性期」と呼びます。例えばアヒルのようにいくつかの鳥では、適切な期間に刷り込みによって求愛の歌を学習する機会を逃してしまうと、求愛の歌を歌うことができなくなることがよく知られています。さらに、感受性期は脳の成熟と関係していると信じられています。そのため、異なる機能は異なる発育段階で学習されます。子猫の視覚野神経の視覚反応性は異なる感受性期に発達します。両目立体視は生後3-5週間の間に発達し、方位選択性は生後6-12週間の間に発達します。鳴禽類は発育段階のいくつかの習熟期に歌の歌い方を学習します。例えば、キンカチョウは生後65日までにモデルとなる歌の音要素を発することができ、90日までに正確な音要素をもった歌を歌うことができます。同様に、ヒトの言語習得にも感受性期があります。母国語として苦もなく話すことができるためには、子供は3-5歳までにその言語が話されている環境で生活し、その言語で使われている音の特徴を記憶しなければなりません。第二言語は、幼児期から思春期までの感受性期にのみに本当に流暢に話すことができるレベルで習得することができることが経験則から分かっています。本研究結果と併せると、これらのパターンから、多くの脊椎動物は異なる行動の側面を異なる発育期間に学習することが分かります。機能ごとの複数の感受性期は動物の成長環境に適応した知覚・認知能力の確立と関係している可能性があります。もし、本研究で観察されたペリソダスの注目すべき左右差をもった捕食行動の発育期間依存的な学習がタンガニーカ湖のような自然環境でも起こるのであれば、ペリソダスは本来の生息地での発育的学習を研究する新しいモデル動物になる可能性があります。
硬骨魚の学習能力は、場合によっては哺乳類や鳥類のものと同程度と言え、様々な分類学上の分類で相同の神経ネットワーク構造を共有しています。学習と記憶は、魚が効率よく獲物を認識し、行動し、捕食するために決定的な役割を担っています。しかし、発育段階特異的な学習とその機能はほとんど研究されていません。本研究は、近年注目されていなかったテーマである行動左右差に対する個体発生の影響を明らかにするものです。ペリソダスでは、行動左右差は初期発育段階特異的に獲得され、鱗食経験によって誘発される可能性があり、脊椎動物で見られる普遍的な行動左右差の確立を理解するための重要な知見をもたらします。ペリソダスの寿命は短いことから、約1年で性的に成熟します。これまでの研究では、捕食行動にかかわる神経回路が提案されており、左右差に関連した脳機能についての遺伝子解析が行われています。ゲノム配列と遺伝子発現データはタンガニーカ湖に生息するペリソダスの近縁種を含むアフリカのシクリッドの代表的な種から得られています。このように、ペリソダスは、捕食行動の発達のダイナミクスとその根底にある神経および遺伝的基盤を研究する上で明らかな長所を持っています。魚の神経学的、遺伝学的変化を調べる方法がたくさんあることから、ペリソダスは捕食者と獲物の相互作用での期間特異的学習の基礎となる脳システムの理解を深める道を開くことができるでしょう。
よろしくお願いします。