ゼブラフィッシュの尾ビレ再生 ~切られた長さを知っている~ 論文紹介

ゼブラフィッシュの尾ビレ再生 ~切られた長さを知っている~
論文名 Regrowth of zebrafish caudal fin regeneration is determined by the amputated length
ゼブラフィッシュの尾ビレ再生は切断された長さで決まる
著者名 Toshiaki Uemoto, Gembu Abe and Koji Tamura
掲載誌 Scientific Reports
掲載年 2020年
リンク https://www.nature.com/articles/s41598-020-57533-6
ゼブラフィッシュの尾ビレの再生についての新しい概念を提唱した2020年の論文です。
両生類の四肢再生では、必ず、元の長さの四肢が再生します。これは、切断された場所に最初にできる再生芽と呼ばれる細胞の塊が、切断された位置を認識して、失われた部分だけを再生するためであることが分かっています。ですので、この再生芽を移植すると、移植先では、移植元の位置に応じた部分だけが再生されます(漫画「あしの再生」参照)。この現象を説明する概念として「位置記憶」というものがあります。つまり、再生芽には位置記憶があり、その記憶に従って、失われた部分が再生されます。
この論文ではゼブラフィッシュの再生について研究を行っています。ゼブラフィッシュというと、発生や行動、遺伝子機能などの研究でよく使われています。ゼブラフィッシュでは、尾ビレが再生します。この尾ビレの再生が両生類の「位置記憶」という概念で説明できるのかを、調べた論文になります。論文では、結論として「位置記憶」とは違う記憶と、そのメカニズムの一端を明らかにしていますが、どこがどう違うのかについてはちょっと分かりにくいかもしれません。漫画では、なるべく違いが分かるように書きましたので、漫画を一読してからのほうが理解しやすいと思います。
新しい概念は、それを提唱したら終わりではなく、始まりになります。今後は提唱者だけでなく、他の研究者によって、この概念が検討されて、間違っていなければ徐々に受け入れられ、間違っていれば修正されたり、受け入れられなくなったりします。この先、この概念が生き残り、教科書に載るようになれば面白いと思います。
この論文では、ひれすじの幅が位置記憶に関わっていることを明らかにしています。では、一体どうやってひれすじの幅を位置記憶とつなげているのでしょうか?その解明が待たれます。また、ゼブラフィッシュ以外の魚、それ以外の生物ではどうなのか?気になる点はいくつもありますので、続報を期待しています。
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
・尾ビレ再生の終了点はこれまで考えられていた時期よりも長い。
・切断されたひれすじの長さによって、再生しているひれすじの成長期間と成長率が決まる。
・再生するひれすじの長さは、切断部位に残されたひれすじの幅によって変わる。
・両生類の四肢再生とは異なり、切断部位の残された断端の状態が位置記憶として働いている可能性がある。
[背景]
両生類や魚類は哺乳類よりも器官の高い再生能力を持っています。酷い損傷を受けたり、失われたりした臓器を再生することができ、その臓器再生は、しばしば付加的に行われます。(補足:付加とは、増殖などにより、失われた部分が修復されること。真の再生。)付加的器官再生の過程では、最初に、傷ついた表皮が切断部位の表面を急速に覆い、その後、脱分化した間葉細胞からなる再生芽が出現します。再生芽が形成された後、適切な組織が再生芽細胞から新たに形成され、元の形態を再構築します。両生類の四肢再生に関する研究から、再生芽細胞が位置記憶をもつ可能性があることを示されています(図1a)。肢は、柱脚、軛脚、自脚のように近位-遠位軸に沿ったパターンを示します。(補足:肢を体幹から関節ごとに区分けすると柱脚、軛脚、自脚となる。近位-遠位は体幹を基準とした位置関係を示す。)四肢が近位-遠位軸上のある点で切断されると、位置記憶といわれるものに従って、切断部位から遠位の四肢部分の組織と形態的構造を再生します。位置記憶は、各再生芽細胞の肢上の相対的な位置(図1a、切断場所3または5)と再生芽が再構築すべき構造に関する情報を提供します。位置記憶は前後軸と背腹軸に沿っても存在すると考えられています。
魚類は切断された尾ビレを完全に再生することができます。ゼブラフィッシュの尾ビレは双葉型の形態を持ち、長いひれすじを持つ背側と腹側の葉型領域と短いひれすじを持つ中央の裂け目領域の3つに分けることができます。ひれすじは神経組織、血管、間葉細胞を包む表皮で覆われた二つのへこんだ皮骨(鱗状ひれすじ)の近位-遠位方向への連なりによってできています。(補足:ひれすじは細かく分節しており、それが連なっている。)再生ヒレは再生芽を形成し、再生芽の細胞は活発に増殖し、両生類の四肢再生と同様に新しいひれすじを形成するために再分化します。脱分化、再生芽形成、および組織の再分化はゼブラフィッシュで集中的に研究されていますが、再生ヒレが元の形態をどのように再構築するか、つまり、元の長さのひれすじをどのように再生するかは不明です。ゼブラフィッシュの尾ビレ再生に位置記憶が関与し、両生類の四肢再生と同様に元の形態を再構築するかどうかも不明です(図1a、b)。近位(尾ビレの根本)で切断したひれすじは遠位(先端)で切断したひれすじよりも速く成長することから、異なる位置の再生芽は異なる位置記憶(成長率として表される)を持つと考えられています。さらに、移植したひれすじの再生能力に関する研究では、再生芽における位置記憶が尾ビレの再生に関与することを明らかにしました。一方で、他の研究から、切断部位(残された部分の状態)におけるいくつかの条件が、ヒレの長さの回復とヒレの形態の再構築にむしろ重要であることが考えられています。尾ビレの完全な形態を再構築するための再生芽における位置記憶の寄与についてはまだ議論されているところです。

本研究では、位置記憶がその形態を再構築するために使用されていると考えられる根拠となる背腹軸に沿ったひれすじの位置的違い、および近位‐遠位軸に沿った切断レベルに依存して、尾ビレ再生中の各ひれすじにおける特徴的な成長あるかどうかを調べました。最初に、ヒレ形態の再構築の終了を定義し、各ひれすじの成長期間と成長速度を測定しました。ひれすじの成長期間と速度を比較することにより、同じ近位‐遠位レベルで切断されたひれすじの成長期間と速度は同じではないことが分かりました。また、近位で切断したひれすじはより長い成長期間とより高い成長速度を示しました。統計解析によって、2つのパラメータの値がひれすじの位置的違いに関係なくひれすじの切断長さと相関することを示したことから、同じ近位‐遠位レベル(図1b)ではなく、背腹軸に沿った波状線(図1c、遠位端から同じ長さで示される)で切断されたひれすじは同じ成長期間と成長率をもつと考えられます。これは両生類の四肢再生のための再生芽における位置記憶の概念とは一致しません。また、切断部位(ひれすじの幅など)の構造的/物理的条件(生体組織の状態)が切断されたひれすじが再生する長さを決定するかもしれないという、いくつかの証拠も示しました。
[結果]
ひれすじ毎の成長期間と成長率の違い
一般的に、器官のような成長している動物の構造の最終的な大きさは、成長期間と成長率の2つのパラメータによって表すことができます。これらのパラメータを定義するために、再生の成長の終了点を明確にしなければなりませんが、魚のヒレの再生では元の形態を再構築するためにどれほど時間がかかるかはあまりわかっていません。これまでの研究は、主にヒレ再生の比較的早い時期に注目していたため、ヒレの形態再構築の終了点について明確に定義されていませんでした。そのため、双葉型の形態を再構築するために必要な成長期間と成長率はわかっていません。急激なヒレの成長を追跡し、その成長の終了点(日数)を決定するために、ヒレの長さと体幹の長さの比(F-B比)を使用しました。ゼブラフィッシュの成体は、その一生を通して一定の長さの割合を保ったまま成長するため、F-B比は一定に保たれます。一方で、ヒレを再構築している過程では、それぞれのひれすじのF-B比は大きくなっていき、最終的には、切断前の値に達したときに一定となります。そのため、急激なヒレの成長の終了点をひれすじの成長が一定になったときと定義しました。
尾ビレ再生中の各ひれすじの成長期間と成長率を計測して比較するために、背腹軸に平行に尾神経骨とつながった前起鰭条の先端部で尾ビレを切断しました(直線切断、図2、図3a)。切断した尾ビレで、全てのひれすじ(図2、背側8本と腹側8本)のF-B比を3日ごとに33日間追跡し、成長率を計算し、ひれすじ長の修復の終了点を評価しました。例えば、図3bでは、終了点がそれぞれ切断後24.2日と17.5日である背側3と背側8のデータを示しています。これは、成長期間がそれぞれ24.2日と17.5日であることを意味しています。この結果は、これらのひれすじでの間では成長期間が1週間違うことを示しています。さらに、切断前のひれすじの長さによって、ひれすじごとに成長期間が違うことが分かりました(図3c)。葉型領域の長いひれすじは、裂け目領域の短いひれすじよりも長い成長期間を示しました。背側と腹側の葉型領域に対称的に位置する同じ長さのひれすじは、同じような成長期間を示しました。さらに、大きな魚と小さな魚の成長期間を比べてみました。実際のヒレの大きさは魚の大きさによって変わりますが、どのような大きさの魚でも、同じような成長期間を示したことから、ヒレ再生の成長期間は体やヒレの大きさとは無関係であると考えられます。

ひれすじ再生中の成長率の変化に関しては、全てのひれすじで切断後3-6日に成長率がもっとも大きくなり、その後は徐々に低下しました(図3d)。成長期間を通して、成長率は常に短いひれすじの背側8より長いひれすじの背側3が大きくなったことから、長いひれすじは短いひれすじよりも常に速く成長すると考えられます。さらに、背側3と腹側3のように葉型領域に対称的に位置するひれすじは、成長期間中に同様の成長率を示しました。

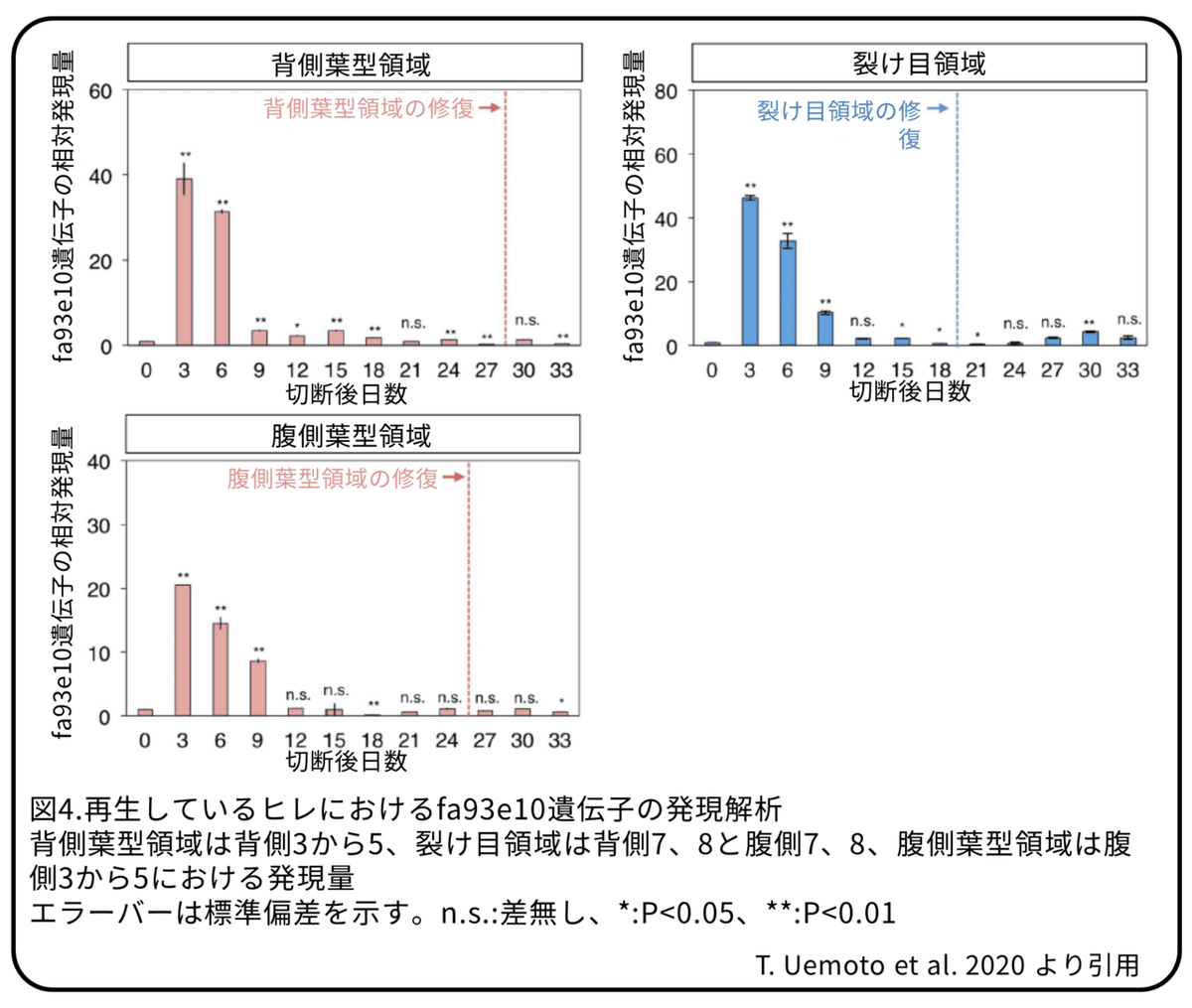
尾ビレでは、増殖性の間葉細胞を覆っている表皮細胞が、ヒレ成長マーカーとして知られているfa93e10遺伝子を発現しています。fa93e10遺伝子に対するqRT-PCRによって、葉型と裂け目の両領域でfa93e10遺伝子の発現は切断後3日目にピークに達することが示されました(図4)。(補足:qRT-PCRは特定の遺伝子の量をPCRによって相対的に測定する方法。この場合はfa93e10遺伝子の量を測定している。)これは、ヒレ再生中の成長率と一致することから、細胞増殖が成長率に寄与していると考えられます。一方で、fa93e10遺伝子の発現レベルは切断後9日目に劇的に減少し、背側の葉型領域では切断後21日目に、切れ目領域と腹側の葉型領域では切断後12日目に切断前と同じレベルになりました。fa93e10遺伝子の発現の低下は、終了点よりもかなり早くなったことから、fa93e10遺伝子の発現が低下する時期は、ひれすじの修復の終了点とは一致しないことが分かりました。細胞増殖と同様に、細胞内外の基質の集積や細胞の形の変化などの他の要因が、形態再構築のための急激な成長の後期には寄与している可能性があります。
切断程度による成長期間と成長率の違い
ゼブラフィッシュの尾ビレ再生に関するこれまでの研究は、ヒレの根本で切断したひれすじは先端で切断したひれすじよりも速く成長するため、切断の程度に関わらず切断されたひれすじの修復には同じ時間がかかることを前提としていました。そして、本研究で、成長期間はひれすじの長い葉型領域とひれすじの短い裂け目領域で違うことが明らかになりました。そのため、それぞれのひれすじは背腹軸にそった位置的違いによって、決まった成長期間を持っていると考えました。
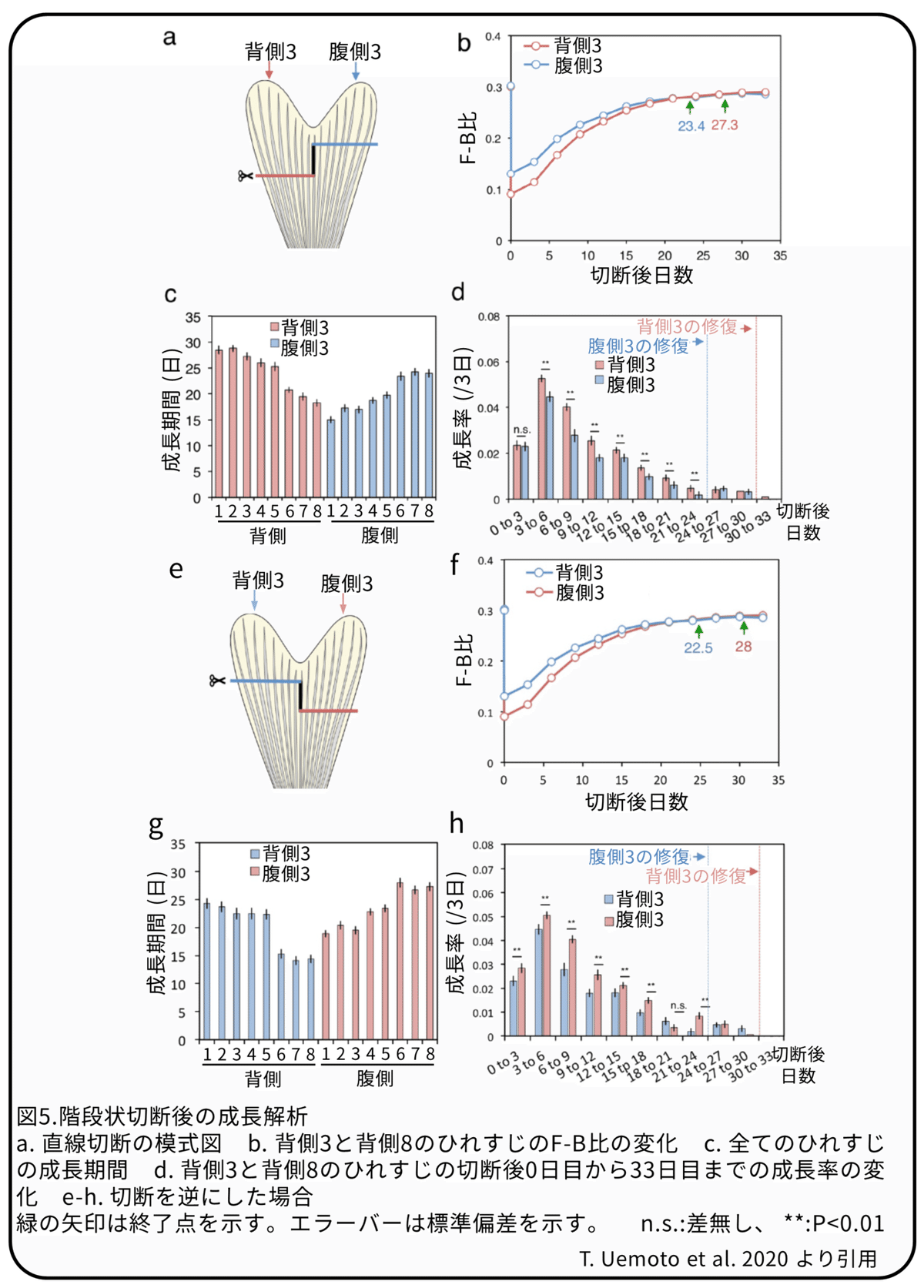
この仮説を確かめるために、双葉型の対象に位置するひれすじが同じ成長期間と成長率を持つことを利用して、同じ個体で背側と腹側の葉型領域を異なるレベルで切断しました(図5a、階段状切断)。背側のひれすじを根元側で、腹側のひれすじを鱗状鰭条の1または3分節分先端側で切断し、成長期間を決定するためにF-B比を追跡しました(図5b,c)。例えば、根元側で切断した背側3はひれすじ長を修復するために27.3日かかりましたが、先端側で切断した腹側3は23.4日かかりました(図5b)。全てのひれすじは根本よりも先端で切断した場合に成長期間が短くなりました(図5c)。このことは、切断程度の違いによって成長期間が異なることを示しています。さらに、逆の階段状切断でも、同様の結果が示されました(図5d-f)。そのため、当初の予想に反して、成長期間はそれぞれのひれすじの位置的違いによって必ずしも固定されていないように見えました。成長率では、根本で切断されたひれすじは先端で切断されたひれすじよりも高い成長率を示しました(図5d)。例えばこの実験で、ともに切断後3日目にピークがありながら、根本で切断された背側3は、先端で切断された腹側3よりも成長期間を通して高い成長率を示しました(図5d)。これらの結果から、これまでの研究と同様に、より根元側で切断されるほど成長率は高くなりますが、これまでの研究とは異なり、違った位置で切断されたひれすじの修復にかかる時間は同じではないと考えられます。
上記の結果から、背腹軸に沿ったひれすじの位置的違いに関わらず、成長期間と成長率は切断の長さ(程度)によって決定されると仮説を立てました。この仮説を確かめるために、ひれすじの切断された長さと成長期間および成長率との間の相関解析を行いました。この解析によって、成長期間はひれすじの切断された長さと明らかな相関性があることが分かったことから、切断の程度の違う場合に成長期間は同じにはならないことが明らかになりました。(図6a)。ひれすじの切断された長さと切断後3日目から15日目までの成長率の間の相関性についても解析し、成長率についてもひれすじの切断された長さと明らかな相関性があることが分かりました(図6b)。これらの結果は、ひれすじの成長期間と成長率はともに切断されたひれすじの長さによって決定されていることを示していることから、背腹軸に沿ったひれすじの位置的違いに関わらず、ひれすじの先端から同じ距離で切断されたひれすじは同じ再成長をしめすと考えられます。

断端の状態は再生しているひれすじの再成長に寄与する
これまでの結果から、尾ビレの再生において同じ長さの切断は同じ成長期間と成長率をもたらすと考えられます。もしゼブラフィッシュの尾ビレの再生間葉細胞がある位置記憶を持っているとしたら、これはその記憶が同じ遠位-近位軸上で同じ位置にあるひれすじの間で同じではないけれども、波状の曲線に沿っては同じであることを意味しています(図1b,c)。これは肢ではある遠位-近位軸上の位置にある再生芽の細胞が位置的価値を持つことを前提とした再生肢芽による位置記憶の概念とは一致しません(図1a)。成長期間と成長率は切断位置近くのひれすじの残された部位(ひれすじの断端)の状態によると予想しました。再生しているひれすじの再成長を制御することに寄与している断端の物理的および生理的状態や遺伝子制御のエピジェネティックな状況といった生体機能の状態を調べるために、物理的状態のひとつであるひれすじの幅(厚さ)に注目しました。それは、最近の研究によって、背腹軸に沿ったひれすじの位置的違いにかかわらず、ひれすじの厚さがひれすじの先端からの距離と相関性があることが示され、損傷したひれすじの周囲長が再生するヒレの大きさと形を決定していると考えられたためです。面白いことに、その研究では切断していない尾ビレでひれすじの周囲長が同じになる部分をつなげると波状になることが示されました。
選択的アドレナリン受容体α1の作動薬であるフェニレフリンはマウスの骨芽細胞様細胞の増殖を制御することが知られています。再生しているヒレをフェニレフリンで処理することで、時々幅が広くなったひれすじができることが分かりました(図7b-h)。さらに、ヒレ再生中にフェニレフリン処理をしても、元と同じ長さのヒレができたことから、フェニレフリン自体は位置記憶に影響を与えないと考えられます。断端のひれすじの幅が再生したひれすじの長さに影響を与えるかどうかを調べるために、切断後7日目に再生領域の真ん中で広がったひれすじを再度切断する連続切断を行いました(図7a)。2度目の再生中には、切断されたヒレはフェニレフリンによる処理を行いませんでした。幅広くなった断端では、1回目の切断前よりも明らかに長いひれすじが再生されることが分かりました(図7i)。これらの結果は、切断位置のひれすじの幅が再生されたひれすじの長さに寄与する可能性を示しています。

[考察]
ゼブラフィッシュの尾ビレ再構築における双葉型ヒレ形態の再構築
再生中に双葉型の尾ビレの形態を再構築するためには、それぞれのひれすじは元の長さを修復するために、背腹軸に沿ったひれすじの位置的違いに依存した適切な長さまで再成長しなければなりません。直線切断実験では、葉型領域の長いひれすじは裂け目領域の短いひれすじよりも長い成長期間と成長率を持つことが分かりました。さらに、背側と腹側に対称的に位置している同じ長さのひれすじは同じ成長期間と成長率を持っていました。階段状切断実験では、根本で切断されたひれすじは先端側で切断されたひれすじよりも長い成長期間と高い成長率も持っており、これらの切断位置特異的な成長期間と成長率は全てのひれすじで見られました。統計解析によって、成長期間と成長率は切断されたひれすじの長さと明らかな相関性を示したことから、背腹軸に沿ったひれすじの位置的違いにかかわらず、ひれすじの成長期間と成長率は切断された長さによって決定されると考えられます(図6c)。言い換えれば、切断されたひれすじは失われた長さを認識し、適切な長さのひれすじを再生させるための成長期間と成長率を制御している可能性があります。その結果、双葉型のヒレ形態は再構築されます。
尾ビレ再生の終了
F-B比を追跡することによって、元のひれすじ長の修復の終了点を正確に評価し、ヒレの形態再構築の過程を記述することができました。この測定によって、直線切断時にひれすじ長の修復にかかる最大期間は最も長いひれすじである背側1で25.1日であることが分かりました。この成長期間はこれまで考えられていたものより約1週間長いものでした。さらに、ひれすじはその修復を全て同じ時点で終了するわけではなく、最小成長期間は最も短いひれすじである腹側8で15.6日でした。最も短いひれすじは最も長いひれすじよりもだいたい10日ほど早く再生しました。本研究では、尾ビレ再生中におけるヒレ長の修復の全容を明らかにすることに初めて成功しました。
修復中のひれすじが急激な成長を止める時期は適切なヒレ形態を再構築するために重要です。急激な成長に外部操作を加えると、ひれすじの過剰成長や切断前の形態とは違った伸長した尾の構築が引き起こされます。本研究では、ヒレ成長のマーカー遺伝子であるfa93e10遺伝子の発現はヒレの急成長と相関性があり、発現解析によって、成長率のピークとfa93e10遺伝子の発現量に相関性があることが分かりました。しかし、fa93e10遺伝子の発現は予想されたひれすじ長の修復の終了点よりも約1週間前に切断前のレベルまで減少しました。そのため、fa93e10遺伝子の発現は再生中の成長率のピークを反映していますが、形態再構築の終了の基準としては適切では無いように見えます。同様に、本研究のF-B比に基づいた基準はヒレ形態の再構築の終了を定義していますが、必ずしも解剖学的な修復や色素パターンの再生といった完全な再生過程の完了を意味するものではありません。終了の示す別の指標がある可能性があり、器官再生の全ての過程を理解するために、様々な生物学的視点から終了点を注意深く決めることが重要になります。
ひれすじ長の制御における位置記憶
適切な長さでひれすじを再生するためには、ひれすじは元の長さについて何らかの認識を持っている必要があります。この現象は両生類の四肢再生で詳しく研究されている「位置記憶」の概念で説明することができます。四肢再生では、肢再生芽細胞が近位-遠位軸に沿った位置情報を提供する位置記憶を持っていると考えられています。その情報は、細胞が相対的に肢のどこに位置しているのか(図1a、切断位置)、修復のために必要な細胞は何か(図1a、切断された部分)といったものになります。多くの研究によって、位置記憶の証拠が示されています。例えば、先端側で切断された肢に作られた再生芽がより根本側で切断された肢に移植されると、移植された再生芽は元の切断位置から先端側の構造の形成のみに寄与します。(補足:結果として、再生された肢は短くなる。)このことから、再生芽で再活性された位置記憶は、周囲の状態によって影響を受けることは決してないと考えられます。重要なのは、近位-遠位軸に沿った切断位置にある細胞は、近位-遠位についての再生芽内の位置記憶は同じものであるということです。
両生類の四肢再生に見られる位置記憶の概念をゼブラフィッシュの尾ビレ再生に当てはめると、根本での切断はより長い再生を(図1b、3の切断位置)、先端側で切断した場合は短い再生を引き起こします(図1b、5の切断位置)。ゼブラフィッシュのこれまでの研究から、ヒレの再生にも位置記憶が関係していると考えられます。ひれすじまたはヒレ片を別の領域に移植し、続けて移植されたひれすじを切断すると、移植された領域の長さではなく、提供したヒレ領域の長さに近い長さに修復されます。(補足:移植されたヒレ片がもともと持っていた位置についての情報に依存して再生される長さが決まる。)この結果から、修復されたひれすじの長さは近位-遠位軸に沿ったある種の位置記憶によって決定されると考えられます。これまでの研究で、先端側で切断されたひれすじの先端に作られ、根本で切断されたひれすじの先端に移植された再生芽細胞は、再生芽細胞の近位-遠位位置に関係なく、完全なひれすじを再生することに寄与することが分かりました。これは、上述した両生類の再生芽の位置記憶にみられる特徴とは一致しません。これらの結果は、再生芽細胞ではなく、むしろ切断部位の断端組織が位置記憶を持っていることを示唆しています。
本研究では、ヒレ再生の成長期間と成長率はひれすじの切断された長さとの明らかな相関性がみられたことから、ひれすじは背腹軸に沿ったひれすじの位置的違いについてではなく、切断された長さを元の長さに修復するための情報として使用していると考えられます。この考えは、尾ビレの近位-遠位軸に沿った位置記憶の概念と似ています。しかし、本研究結果は、ひれすじの位置的違いに関わらず、同じ長さで切断したひれすじは同じ成長期間と成長率を持つことも明らかにしました。もし、それぞれのひれすじが切断された長さを認識し位置記憶として使用するならば、その位置記憶は、背腹軸に沿って直線ではなく、波状に切断したときに同じものになると思われます。これは、両生類の四肢再生にみられるように、ヒレの近位-遠位位置で同じであると考えられている再生芽の位置記憶と一致しません。
さらに、最近の研究で示唆されたように、切断位置の断端組織の構造的で物理的な状態がひれすじの切断された長さを認識し記憶として使われている可能性があります。面白いことに、最近の研究で、ひれすじの長さと厚さ(周囲長)には強い相関性があることが明らかになりました。これは、本研究の成長期間と成長率の結果と一致します。ひれすじの厚さは断端の状態の記憶として再生されたひれすじの長さの決定に寄与する可能性があります。本研究では、フェニレフリン処理によって幅が広くなったひれすじの切断はより長いひれすじの再生を引き起こしました。この結果は、再生芽ではなく、ひれすじの幅のような断端の状態にある位置記憶が再生されたひれすじの長さを決定する可能性を支持しています。切断されたひれすじの再生する長さを決定するメカニズムを明らかにするためにはさらなる研究が必要です。
よろしくお願いします。