プラナリアの有性化物質5 ~いろいろな扁形動物~ 論文紹介

プラナリアの有性化物質5 ~いろいろな扁形動物~
論文名 Sex-inducing effects toward planarians widely present among parasitic flatworms
プラナリアに対する有性化効果は寄生性扁形動物の間に広く存在する
著者名 Kiyono Sekii, Soichiro Miyashita, Kentaro Yamaguchi, Ikuma Saito, Yuria Saito, Sayaka Manta, Masaki Ishikawa, Miyu Narita, Taro Watanabe, Riku Ito,1 Mizuki Taguchi, Ryohei Furukawa, Aoi Ikeuchi, Kayoko Matsuo, Goro Kurita, Takashi Kumagai, Sho Shirakashi, Kazuo Ogawa, Kimitoshi Sakamoto, Ryo Koyanagi, Noriyuki Satoh, Mizuki Sasaki, Takanobu Maezawa, Madoka Ichikawa-Seki, and Kazuya Kobayashi
掲載誌 iScience
掲載年 2023年
リンク doi.org/10.1016/j.isci.2022.105776
プラナリアの有性化物質が寄生性の扁形動物にもあるのかを調べた2023年の論文です。
プラナリアの有性化物質については当サイトで既に4報(「プラナリアの有性化物質 ~アミノ酸の不思議~」、「プラナリアの有性化物質その2 ~いろいろなプラナリア~」、「プラナリアの有性化物質その3 ~卵巣誘導物質セロトニン~」、「プラナリアの有性化物質その4 ~手がかりはアミノ酸輸送体?~」)を紹介しています。ほぼ1年に1報ペースで同じ研究室から継続的に論文が発表されており、この研究に対する熱量が伝わってきます。
プラナリアのリュウキュウナミウズムシは有性生殖と無性生殖の2つの生殖様式によって増殖することができます。生殖器官を持たない無性リュウキュウナミウズムシは分裂と再生によって増殖し、生殖器官を持つ有性リュウキュウナミウズムシは他の個体と交接して卵を産むことで増殖します。同じ個体が環境の変化などで、無性から有性へ、有性から無性へと変化します。最近では、有性化する時期が一年ごとに周期的に現れるという報告が出ています[1]。リュウキュウナミウズムシは、特定のエサを与えることで、無性から有性への変化を人工的に起こすことができます。また、有性化の過程も調べられており、卵巣が作られた後に交接器官ができると、その後は特定のエサがなくても完全に有性化することも分かっています(漫画「リュウキュウナミウズムシの有性化」参照)。そのため、無性から有性への変化、有性化を研究するために適した実験動物になっています。
特定のエサを与えることで有性化するため、そのエサには有性化を引き起こす化合物、有性化物質が含まれていると考えられます。それを発見しようというのが、本論文も含めてこれまで当サイトで紹介してきた研究となります。これまでに、リュウキュウナミウズムシと同じ三岐腸目のプラナリアであるイズミオオウズムシ、オオミスジコウガイビルをすりつぶした溶液からトリプトファンと(漫画「トリプトファンと有性化物質」参照)、有性化すると発現が変化する遺伝子を手がかりに、トリプトファンの代謝物であるセロトニンを、卵巣の形成を誘導する物質、卵巣誘導物質として発見していますが、完全に有性化させる有性化物質はまだ見つかっていません。
本論文では、三岐腸目以外の扁形動物として、進化的に三岐腸目と共通の祖先を持つと考えられる寄生性扁形動物を対象として、リュウキュウナミウズムシを有性化させる有性化物質を探索しています(漫画「寄生性扁形動物」参照)。この論文を読むまではいわゆるサナダムシといった寄生虫がプラナリアと進化的に近い扁形動物であるとは知りませんでした。寄生虫は、その生涯を通して宿主を乗り換えたり、いくつかの宿主を乗り換えないと成虫にならなかったりと、複雑な生活環を持つことが多く、培養して増やすことは非常に難しいと思います。また、すりつぶして溶液にする必要が有ることから、ある程度の量が必要となります(漫画「扁形動物の抽出物」参照)。そのため、材料を確保することは非常に大変だったと思います。それでも、本論文では9種もの寄生虫を使用していることから、著者たちのなみなみならぬ熱意や探究心が伺えます。
本論文は二部構成になっています。前半は上記の寄生性扁形動物を材料にした有性化物質の探索です。後半はリュウキュウナミウズムシが産む複合卵に注目し、複合卵に含まれるたんぱく質の直接的な設計図であるmRNAを調べ、有性化に関わる遺伝子を見つけることで、有性化メカニズムを解明するとともに有性化物質の手がかりを探索しています。また、後半ではメタボローム解析を行っています。これは、生体内に含まれる、核酸、たんぱく質以外の糖、有機化合物、アミノ酸などを全て調べる解析です。無性リュウキュウナミウズムシと有性リュウキュウナミウズムシまたは複合卵の解析結果を比較することで、有性化に関わる有機化合物を探索しています。
本論文では、残念ながら有性化物質を突き止めることができていません。しかしながら、トリプトファンとは別の有性化物質の存在が明確となり、メタボローム解析により有性化に関わりそうな有機化合物のデータもそろってきました。一歩ずつ着実に有性化物質の同定に近づいていると思います。その日を期待して続報を待ちたいと思います。
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
寄生性扁形動物である単生類、吸虫類、条虫類の抽出物のうち、単生類と吸虫類の抽出物は有性化効果を示した。
吸虫類の双口吸虫にはトリプトファンとは異なる有性化効果を示す化合物があり、プラナリアであるオオミスジコウガイビルの有性化物質と同一物質であると考えられる。
リュウキュウナミウズムシの複合卵では、有性化によって発現が変化するプリン体代謝物経路に関わる遺伝子が発現していた。
無性リュウキュウナミウズムシと比べて、有性リュウキュウナミウズムシか複合卵で量が多い代謝物として6種のプリン体代謝物を含む計18種の卵巣誘導物質を同定した。
[背景]
寄生性扁形動物は、有性生殖サイクルを完成させために多くの脊椎動物に感染します。それらは、宿主から得るエネルギーの大半を卵の生産に費やし、しばしば宿主に貧血、炎症、その他の問題を引き起こします。このような宿主の病気は、人の健康問題や畜産・水産業の生産性低下に起因する大きな社会経済的損失をもたらす可能性があります。そのため、寄生性扁形動物の有性生殖のメカニズム、特にその基盤となる生理活性物質の解明は、寄生性扁形動物の急速な無性生殖や有害な有性生殖を阻害する抗寄生虫薬の設計を容易にすることから、経済・生物医学的に特に重要であると考えられています。しかし、複雑な扁形動物の生活環を実験室で維持することは、宿主内での性成熟を観察することと同様に困難であるため、これらのメカニズムについてはほとんど分かっていません。
寄生性扁形動物の有性生殖の基盤となる共通原理を明らかにすることは、これらの幅広い種に対する理解と防除能力を大きく向上させるでしょう。本研究では、非寄生性扁形動物であるプラナリアで報告されている特異的な生理活性物質である有性化物質に着目しました。これらの物質が寄生性扁形動物にも存在する場合、共通の有性生殖メカニズムの鍵となる要因になると仮定しました。三岐腸目のプラナリアは寄生性扁形動物の進化的祖先であり、寄生性扁形動物群(新皮亜綱)および別の扁形動物群(ヒメヒラウズムシ目)と共に単系統群を構成しています(図1A)。プラナリアは、比較的取り扱いが容易で安全なため、寄生性近縁種を研究するための優れたモデル生物として台頭してきています。寄生性扁形動物がその複雑な生活環の中で両方の生殖様式を兼ね備えているように、一部のプラナリアは環境条件によって無性生殖と有性生殖を使い分けることができます。プラナリアには、無性生殖時には新しい個体に必要なあらゆる種類の組織に、有性生殖時には生殖細胞や他の生殖器官に分化できるネオブラストと呼ばれる多能性幹細胞が存在し、両方の生殖様式を利用することできます。これまでに、無性のヨーロッパナミウズムシに別種の有性プラナリアを与えると有性化することが発見され、有性プラナリアには種特異的ではないホルモン様物質(以下、有性化物質)があることが明らかになっています。給餌による実験的な有性化は ヨーロッパナミウズムシに限った現象ではなく、いくつかのプラナリア種で観察されています。有性化物質は、幹細胞の機能や挙動を有性生殖用に変化させることで、無性生殖のプラナリアを有性生殖に誘導することができます。生殖様式を切り替えることができるプラナリアは、環境要因などに応じてそのような物質の産生を制御することでこれを実現しています。しかし、寄生性扁形動物における有性化物質の存在とその正体については、いまだ分かっていません。寄生性扁形動物に有性化物質が存在すれば、その物質が寄生性扁形動物の性成熟の引き金になっている可能性があり、今後の寄生性扁形動物の研究において重要な意味を持つ可能性があります。

プラナリアのリュウキュウナミウズムシ(扁形動物門 渦虫綱 三岐腸目)は、特に有性化物質の研究に適したモデル生物です。リュウキュウナミウズムシのOH株は、主に無性生殖である横裂とその後の再生によって繁殖します。しかし、他のプラナリア種とは異なり、リュウキュウナミウズムシに有性化物質を含む有性の扁形動物を与えることで、実験的かつ安定的に有性化させることができます。この給餌実験系により、対象種が有性化物質を持っているかどうか、そして特定の化学物質の有性化作用を検討することができます。
寄生性扁形動物に有性化物質が存在すると予想される理由が2つあります。1つ目は、有性化物質の正体はまだ明らかにされていませんが、少なくとも三岐腸目の非寄生性プラナリアでは種を超えて保存されていると考えられ、多数の卵黄細胞と数個の受精卵(すなわち外黄卵)を含む複合卵に有性化物質が存在することが報告されています。また、外黄卵を持たない海産扁形動物であるミノヒラムシと非扁形動物のチャコウラナメクジを リュウキュウナミウズムシに与えたところ、いずれも完全に有性化するほどの有性化作用はありませんでした。これらの結果から、卵黄腺が有性化物質の産生と貯蔵の両方もしくは一方に重要であり、有性化物質が外黄卵を持つ種内で保存されていると考えられます。そのため、外黄卵を持つ寄生性扁形動物には、有性化物質が存在すると予想されます(図1A)。2つ目として、近年の研究から、吸虫と非寄生性プラナリアの幹細胞の不均質性や遺伝子発現特性の類似性が明らかになり、寄生性扁形生物の複雑な生活環は、非寄生性である祖先に既に存在する発生プログラムを適応させて進化したと考えられています。そのため、有性化物質を介してネオブラストを制御するシステムも寄生性扁形動物間で共有されていると予想されます。
本研究では、リュウキュウナミウズムシの給餌実験系を用い、寄生種を中心に、異なる扁形動物に含まれる物質がリュウキュウナミウズムシの有性化を誘導するかどうかを明らかにするとともに、扁形動物の有性化物質に何が含まれるのかを明らかにしました。
[結果]
寄生性扁形動物における有性化効果の存在
扁形動物門は1つの非寄生性群(渦虫綱)と、単生類(単生下綱)、吸虫類(吸虫下綱)、条虫類(条虫下綱)からなる進化的に近い寄生性群から構成されています(図1A)。単生類は水生脊椎動物のエラや皮膚の上で生活する外寄生性生物ですが、吸虫類と条虫類は水生または陸生脊椎動物の消化管、血球、または内部臓器の内部で内寄生性的に生活し、しばしば宿主を変えるような複雑な生活環を持っています。扁形動物に有性化効果が存在しているかどうかを調べるために、非寄生性群と3つの寄生性群からの9種を含む幅広く多様な種を対象にしました(図1A)。リュウキュウナミウズムシとは異なる科である非寄生性プラナリアのイズミオオウズムシは陽性対照群として使用しました。イズミオオウズムシは既に給餌実験でリュウキュウナミウズムシに対して有性化効果を持つことが分かっています。その他の対象種は、単生類のネオヘテロボツリウム(和名なし、Neoheterobothrium hirame)とハダムシ、吸虫類の肝蛭と巨大肝蛭の雑種(以下、雑種肝蛭)と双口吸虫とマンソン住血吸虫、条虫類のヒメノレピス(和名なし、Hymenolepis microstoma)とベネデン条虫とジフィロボツリウム(和名なし、Diphyllobothrium hottai)とマンソン裂頭条虫です。
それぞれの扁形動物に含まれる化学物質はその疎水性に従ってM0、M10、そしてM100の3つの画分に分けられました。(補足:M0が最も疎水性が低く、M100が最も疎水性が高い。)これらの画分の無性リュウキュウナミウズムシに対する有性化効果は4週間の給餌実験を通して調べられました。無性リュウキュウナミウズムシの有性化過程は大雑把に6つの形態的に区別できるステージに分けられます(図1B)。ステージ1と2では、卵巣の発生が起こりますが、他の生殖器官はステージ3-6の間に形成されます。ステージ2とステージ3の間には、さらなる有性化物質の投与がなくても有性化が自律的に完遂する能力をプラナリアが示す、引返し限界点があります。本研究では、有性化物質を、無性リュウキュウナミウズムシが引返し限界点を超えるまで有性化させる能力があるものと定義し、有性化効果はリュウキュウナミウズムシに交接器官が形成されるかどうかで評価しました(図1B)。完全な有性化効果は、プラナリアが引返し限界点に到達し、交接器官が観察された場合と定義されました。卵巣形成だけが誘導され、引返し限界点に到達しない場合は、卵巣誘導効果としました。
この定義に従って、イズミオオウズムシと同様に、単生類から吸虫類までの全ての実験種で完全な有性化効果が観察されました(図2)。これまでに報告されたイズミオオウズムシの結果と同様に、3つの画分の中で、一貫してM10画分は完全な有性化効果を示しました(図2、3)。吸虫類のマンソン住血吸虫 に関しては、オスとメスを分けて実験しました。ほとんどの扁形動物種が雌雄同体ですが、この吸虫やその属する住血吸虫科の扁形動物は雌雄異体で、それぞれの個体はオスまたはメスです。注目すべきは、メスは小さく採集することが難しいために、画分に分けることなく直接食べさせられましたが、完全な有性化効果がオスでもメスでも観察されたことです(図2)。一方で、条虫ではどの画分でも完全な有性化効果は観察されませんでしたが、卵巣を形成させる部分的な有性化効果が一部の種で観察されました(図2、3)。


M10画分に加えて、雑種肝蛭と双口吸虫のM100画分も完全な有性化効果を示しました(図3)。これは以前の研究で実験されたイズミオオウズムシやその他のプラナリアでは観察されませんでした。これは、M10画分に溶出されなかった過剰量の有性化物質が、M100画分に持ち込まれてしまったためである可能性があります。もしくは、より疎水性の高い別の化合物が吸虫類では有性化物質として機能しているかもしれません。これら2つの可能性を切り分けるために、双口吸虫を使い、M0、M10、M30、M50、M100の5つの画分に分け、追加実験を行いました。最も高い有性化効果はM10画分で観察され、全てのプラナリアで引返し限界点を超える交接器官の形成が確認されました。さらに、M100画分で観察された有性化効果はM30画分に移動しました。そのため、完全な有性化効果がM100画分で示された理由は、M10画分に含まれるものと同様の化学物質が持ち込まれたためと思われます。双口吸虫がM30画分に溶出されるわずかに疎水性が異なる有性化物質を持っているという可能性を完全に否定することはできませんが、全ての結果から、調べた全ての扁形動物でM10画分に含まれる同じ化学物質が完全な有性化効果を担っていると考えられるため、有性化物質はこれらの扁形動物で共有されている可能性があります。
精巣と卵黄腺は顕微鏡で観察できないために、生殖器官のマーカー遺伝子の発現を定量的RT-PCRによって調べました。この解析では、精巣と卵黄腺のマーカー遺伝子として、それぞれDrY1遺伝子とDryg遺伝子が使用されました。リュウキュウナミウズムシでは卵巣のマーカー遺伝子が同定されていませんが、地中海プラナリアでは報告されています。本研究では、卵巣マーカー遺伝子としてTR34905|c0_g1_i1遺伝子を単離することに成功しました。これら3つのマーカー遺伝子を使用して、それぞれの給餌実験で性的に発達した10または11匹のプラナリアを定量的RT-PCRによって調べました。顕微鏡観察の結果と同様に、イズミオオウズムシや単生類、吸虫類といった完全な有性化効果を持つ種のM10画分を給餌されたプラナリアでは、卵巣、精巣、卵黄腺のマーカー遺伝子の発現が対照群のプラナリアと比較して有意に上昇しました(図4)。これらの結果から、寄生性扁形動物とイズミオオウズムシでは、M10画分に含まれるわずかに疎水性の高い化学物質が完全な有性化効果を担うことが支持されました。
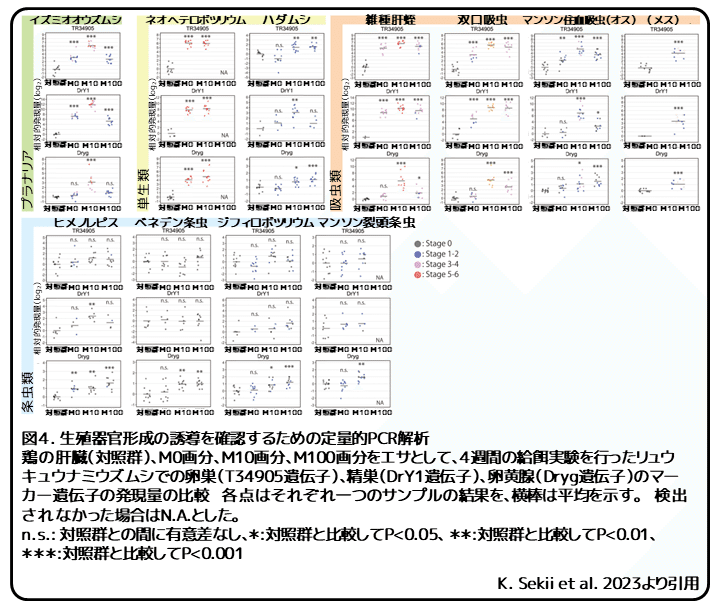
興味深いことに、いくつかの条虫を給餌されたプラナリアでは、精巣と卵黄腺のマーカー遺伝子の発現が、時折上昇しました(図4)。しかし、切片による解剖学的解析では、これらの器官の組織化された構造はほとんどの場合に観察されませんでした。わずかな例外として、ヒメノレピスのM10画分を給餌されたプラナリアで小さな精巣が観察されました。この場合では、精巣のマーカー遺伝子の発現が有意に上昇し、観察結果と一致しています。しかし、観察された精巣は体全体でわずかに2つしかなく、その大きさはイズミオオウズムシのM10画分を給餌されたプラナリアと比べて非常に小さく、精子や精細胞は観察されませんでした。そのため、組織化された精巣の形成を誘導するヒメノレピスのM10画分の効果は非常に弱いと思われます。交接器官は切片からは確認されませんでした。条虫を給餌されたプラナリアでの卵巣のマーカー遺伝子の発現では、対照群との有意差がありませんでした(図4)。これらの結果から、組織切片で観察された小さな卵巣からも裏付けられるように、最初の重要なステップでの失敗、つまり卵巣が十分に発達しないことによって有性化が上手く進まなかったと考えられます。
プラナリアと寄生性扁形動物の間で保存されている可能性が高い有性化物質
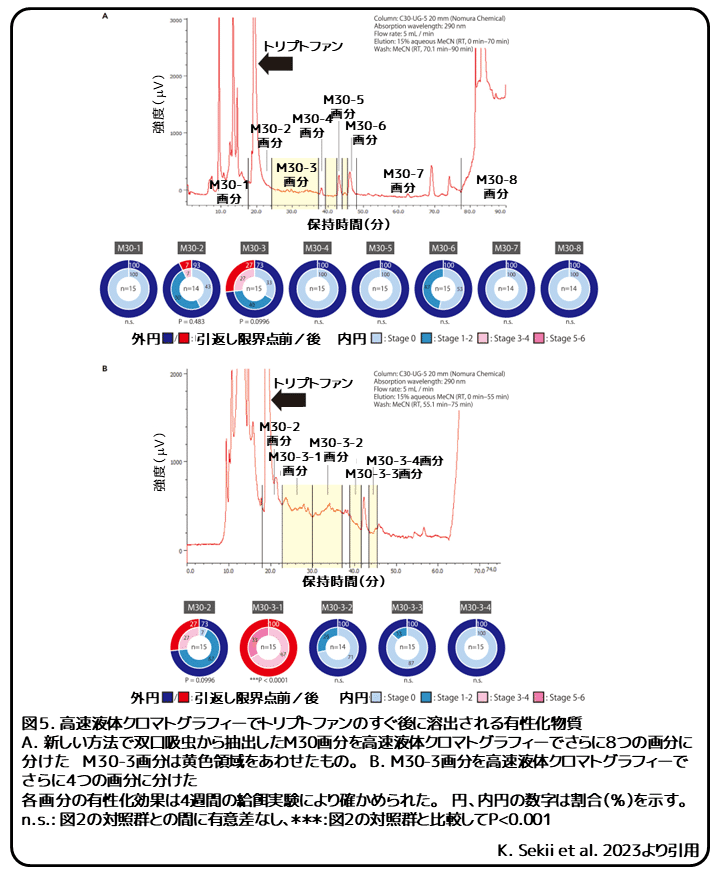
有性化物質はプラナリア、単生類、吸虫類に存在しましたが、NMRなどによる構造予測は行われなかったため、それらが全て同じ化学物質であるかどうかはわかりません。(補足:NMR(Nuclear Magnetic Resonance、核磁気共鳴装置)は強い磁場に置いた試料にラジオ波をあてることで核磁気共鳴を起こし、その際に発生する信号から分子構造を解析する装置。複雑な有機化合物の化学構造の決定のために使用される。) そのため、有性化活性がある画分に大量のトリプトファンが含まれてしまうという従来法の問題点を克服する、有性化物質を精製する新しい方法を確立しました。最初に、体のサイズが大きいために材料を多く用意できるオオミスジコウガイビルを使い、オープンカラムを用いた大量精製法を確立しました。この方法では、従来法のM10画分で観察された完全な有性化効果は、M30画分へと移動しました。双口吸虫を材料に、この方法で精製すると、完全な有性化効果は同様にM30画分へと移動しました。この方法の利点は、有性扁形動物に大量に含まれるトリプトファンは主にM10画分にとどまり、M30画分にはほとんど含まれなかったことです。
双口吸虫のM30画分は高速液体クロマトグラフィーで分画され、主なピークとその他の画分について有性化効果を調べました。(補足:高速液体クロマトグラフィーは溶液中の成分を分離する方法で、分離された成分は検出器の信号強度によってピークとして確認することができる。)M30-2画分にはトリプトファンが、主だったピークのない混合画分であるM30-3画分には完全な有性化効果があることが分かりましたが、その効果は対照群と比較して有意差がみられませんでした(図5A)。M30-3画分をさらに分画して、有性化効果を調べると、トリプトファンのピークのすぐ後に溶出されるM30-3-1画分で有意に強い効果が示されました(図5B)。繁殖期に野外で採集する必要があるオオミスジコウガイビルのM30画分では、画分の量が限られており、有性化活性が不安定であったために、同様の解析を行うことができませんでした。しかし、双口吸虫の結果は、オオミスジコウガイビルでトリプトファンの近くに溶出されるがトリプトファンとは異なる化学物質に有性化効果があることを示したこれまでの報告と一致しました。M30-3-1画分には明確なピークはなく、有性化効果を担う化学物質の量はこの時点で構造予測を行うには十分ではありませんでした(図5A、B)。しかし、本研究で報告された、新しい方法による完全な有性化効果のM30画分への移動、および本研究とこれまでの研究での高速液体クロマトグラフィーの結果から、オオミスジコウガイビルと双口吸虫にある完全な有性化効果を担う化学物質はトリプトファンのすぐ近くに溶出される同じ物質であると考えられます。
有性化効果の鍵となる要因としてのプリン体代謝
有性化物質の分離と同定は達成されませんでしたが、その物質が既知の化学物質である可能性を考えました。有性化物質の分子実体に関する手がかりを得るために、別の方法を試しました。本研究の給餌実験結果から、条虫類を除いて、内黄卵を産む寄生性扁形動物は完全な有性化効果を示すことが分かりました(図2)。これは、有性化物質の生産と貯蔵の両方またはどちらか一方に卵黄細胞が重要であり、そのような物質が内黄卵を産む種間に保存されていることを示唆した以前の研究と一致します。そのため、卵黄細胞に存在する有性化物質を同定することを目的として、卵黄細胞を豊富に含むリュウキュウナミウズムシの複合卵と有性扁形動物のトランスクリプトーム解析とメタボローム解析を行いました。(補足:トランスクリプトーム解析は材料中に含まれるmRNAを、メタボローム解析は代謝物を網羅的に解析すること。)
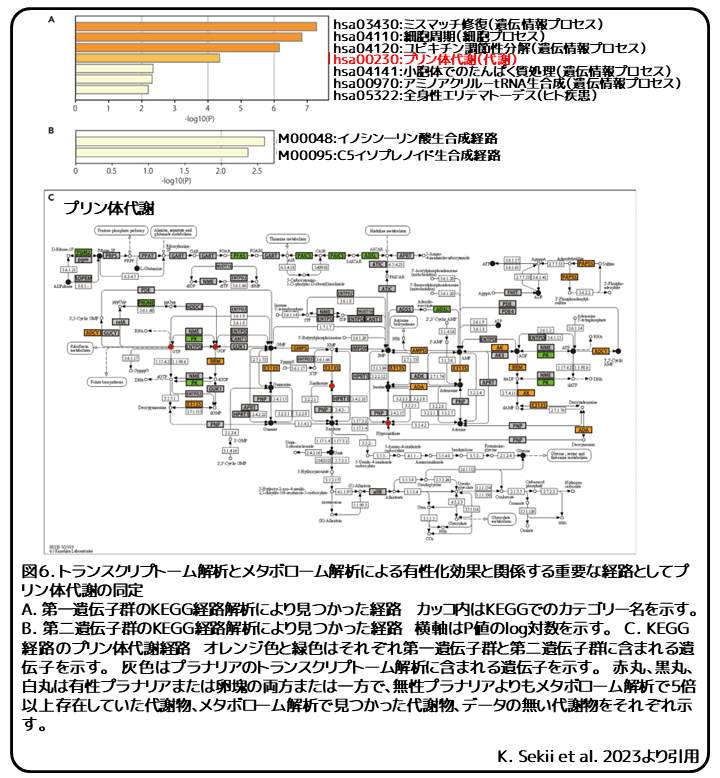
最初に、リュウキュウナミウズムシの複合卵のトランスクリプトーム解析を行い、転写物モデルの新規データセットを作成しました。この複合卵のデータセットと有性扁形動物のデータセットを融合し、複合卵で発現する有性化によって発現が変化した遺伝子群(DEG; differentially expressed gene)を第一遺伝子群としました。これらは卵黄細胞での有性化物質の生産に関わる可能性のある候補遺伝子群です。第一遺伝子群の京都遺伝子ゲノム百科事典(KEGG; Kyoto Encyclopedia of Genes and Genomes)経路解析から、プリン体代謝が代謝カテゴリの中で唯一の発見された経路でしたが、非代謝カテゴリでは遺伝情報プロセス、細胞プロセス、ヒト疾患などが見つかりました(図6A)。(補足: KEGGは京都大学の金久研究所によって作られた遺伝子、ゲノム、化合物などの生命システムを理解するためのデータベースのこと。KEGG経路解析は、そのデータベースにある分子間の相互作用やネットワークなどの経路情報をもとに解析を行うこと。)さらに、有性化物質の生産に関わる酵素が比較的保存されていることが予想されたため、イズミオオウズムシとリュウキュウナミウズムシで共通する酵素を探索するために、イズミオオウズムシの複合卵のトランスクリプトーム解析を行いました。イズミオオウズムシの複合卵の新規データセットを使用して、第一遺伝子群から第二遺伝子群を抽出しました。(補足:方法によると、第一遺伝子群のうちイズミオオウズムシの複合卵で発現する遺伝子と共通する遺伝子群が第二遺伝子群にあたる。)この第二遺伝子群はイズミオオウズムシの複合卵で発現する遺伝子群と非常に似ていました。第二遺伝子群のKEGG経路解析から、プリン体代謝に関わるイノシン一リン酸生合成経路とC5イソプレノイド生合成経路が見つかりました(図6B)。これらの発見に基づいて、第一遺伝子群と第二遺伝子群の遺伝子をKEGG経路のプリン体代謝経路に当てはめることで、第一遺伝子群と第二遺伝子群の遺伝子はGTPやATPといった主なプリン体代謝物の生産に関わることが分かりました(図6C)。プリンヌクレオチドは新規合成経路を介して生合成されるか、サルベージ経路を介して余剰塩基からリサイクルされます。プリン体代謝経路に当てはめられた第二遺伝子群は、多くはプリンヌクレオチド新規合成経路でみられました(図6C、緑)。さらに、GTPもしくはATPとGDPもしくはADPの相互変換を触媒するピルビン酸キナーゼも第二遺伝子群に含まれていました。これらの結果から、プリン体関連代謝物はリュウキュウナミウズムシの複合卵で活発に合成され、この特徴はイズミオオウズムシにも保存されていると考えられます。
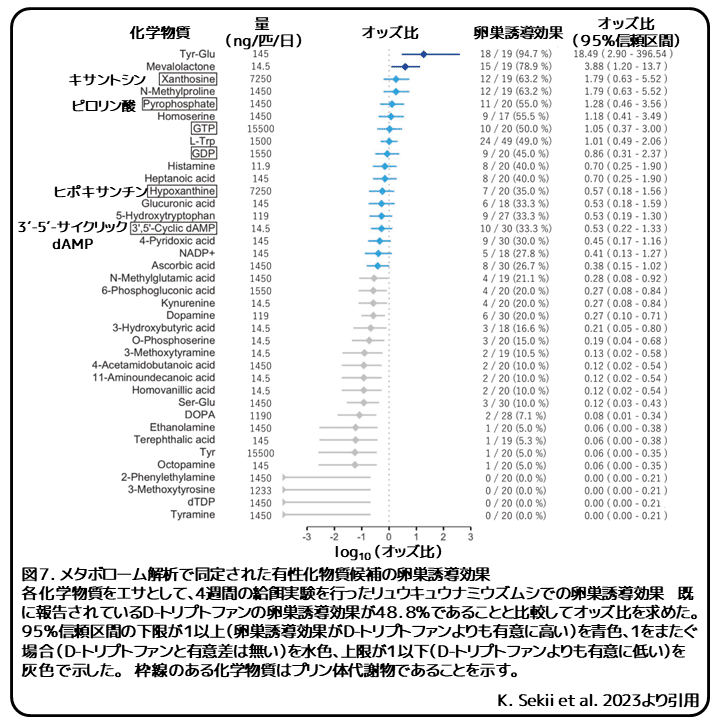
次に、無性と有性のリュウキュウナミウズムシと複合卵のメタボローム解析を比較しました。有性リュウキュウナミウズムシと複合卵は完全な有性化効果を示すため、無性リュウキュウナミウズムシと比較して有性リュウキュウナミウズムシと複合卵の両方もしくはどちらか一方で5倍以上ある代謝物を潜在的有性化物質として選択しました。注意すべきは、メタボローム解析に使用されたデータ数は1であることです。そのため、比較と選択に統計解析を使用しませんでした。これらの候補物質の有性化効果を複数濃度での給餌実験で調べました。調べた38種の化学物質のうち18種には、これまでに卵巣誘導物質として同定されているD-トリプトファンと同等の卵巣誘導効果がありました(図7)。同定された18種の卵巣誘導物質には、キサントシン、ピロリン酸、GTP、GDP、ヒポキサンチン、3′-5′-サイクリックdAMP の6つのプリン体代謝関連物質が含まれていました(図6、7)。キサントシン、ピロリン酸、GTP、GDPを給餌されたプラナリアでは、一部がまれに引返し限界点を超えましたが、対照群と比較して有意差はみられませんでした。完全な有性化効果を持つ有効物質は発見されませんでした。トランスクリプトーム解析とメタボローム解析の結果は適度に一致していたことから、第一遺伝子群と第二遺伝子群の活性化によって上昇した6つの代謝物を含むプリン体代謝が扁形動物の性成熟に重要な役割を担っていることが強く支持されました。
[考察]
本研究では、リュウキュウナミウズムシに対する完全な有性化効果は、内黄卵を産む非寄生性と寄生性の両方の扁形動物、つまり、非寄生性のプラナリア、寄生性の単生類と吸虫類で幅広く観察されました。オープンカラムと高速液体クロマトグラフィーを使用した新しい分画方法によって、双口吸虫に含まれる完全な有性化効果を担う物質がトリプトファンのすぐ近くに溶出されることが明らかとなり、これは、これまでのオオミスジコウガイビルの結果と一致しました。これらの結果から、完全な有性化効果を担う物質は、系統的に近縁ではなく、異なる生活様式であっても扁形動物の間で保存されている可能性が高いことが分かりました。さらに、同定された18種の卵巣誘導物質のうち、6種がプリン体代謝に関連していたことから、プリン体代謝が扁形動物の有性化に重要な役割を担っていることを明らかにしました。
注目すべきは、マンソン住血吸虫のオスもメスも完全な有性化効果を示したことです。これまでの研究で、オスのマンソン住血吸虫にはない卵黄細胞に有性化物質が含まれている可能性が高いことが分かっていたため、これらの結果は予想外でした。しかし、本研究結果は合理的であることから、有性化物質は雄性と雌性の両方の維持に必要である可能性があります。給餌実験に使われたマンソン住血吸虫は性成熟した雌雄の対として採集され、その後オスとメスに分けられたため、オスは卵黄腺を持つメスから有性化物質を得ていた可能性があります。一貫して、マンソン住血吸虫のオスから抽出したM10画分の完全有性化効果は雑種肝蛭や双口吸虫が示す効果と比較して中程度でした(図2)。これら2種の吸虫類は、それぞれのM10画分によって給餌されたプラナリアのそれぞれ97%と100%が引返し限界点を超えたことからわかるように、多数の卵黄腺を体中に持ち、多量の有性化物質を持っている可能性があります(図2)。番になったあと、マンソン住血吸虫のオスとメスは永遠にくっついたままになり、機能的には雌雄同体になります。しかし、メスの乾燥重量はオスよりも約3.8倍小さいです。他の2種の吸虫類と比較して、マンソン住血吸虫のオスの有性化物質の少なさは、マンソン住血吸虫のオスがメスから主に受け取っている可能性を反映しているかもしれません。または、マンソン住血吸虫のオスも有性化物質を生産する可能性もあります。興味深いことに、オスは時折精巣に加えて、疑似卵巣と卵黄腺を発達させます。そのため、雌雄同体の祖先の卵黄腺に由来する遺残として、マンソン住血吸虫のオスは有性化物質を生産する細胞を持つ可能性があります。(補足:遺残とは本来退化消失すべき組織が存在すること。)ほとんどの住血吸虫属では、メスはオスと番になるまでは性成熟することができませんが、オスはメスがいなくてもある程度まで精巣を発達させることができます。物理的な接触面で作用する化学的な要因は長い間謎に包まれていましたが、本研究によって、オスによって生産される有性化物質がそのような要因のひとつである可能性が浮かび上がりました。
条虫類も内黄卵を産みます。しかし、興味深いことに、本研究で使用した条虫では完全有性化効果を示さず、卵巣誘導効果のみでした。この理由として考えられるのは、吸虫類と比較して条虫類では卵黄腺の大きさと機能が縮小していることから、有性化物質が減少している可能性です。イズミオオウズムシのM0画分とM10画分の混合物の完全有性化効果が、5倍に希釈することで劇的に低下したことが、以前の研究で報告されています。そのため、本研究で使用された条虫類の有性化物質の量では、リュウキュウナミウズムシを有性化させるために十分ではなかった可能性があります。しかし、より可能性がある別の理由は、条虫類の性成熟に必要な化学物質セットがわずかに異なることです。この可能性は、有性化の第一段階である卵巣の発達を示すマーカー遺伝子の発現が上昇することがなくても、引返し限界点を超えた後に発達する生殖器官のマーカー遺伝子の発現が予想外に上昇したことを示した給餌実験および定量的RT-PCRの結果から支持されます。実際、組織切片の観察では、例外的でしたが、ヒメノレピスにM10画分を給餌されたプラナリアでは非常に小さな精巣が観察されました。条虫類を給餌されたグループでは組織化された精巣、卵黄腺、交接器官が観察されなかったことから、複合卵は形成されそうにはありませんでした。条虫類に含まれる物質は組織化された生殖構造の完全な発達必要なものとは異なる可能性がありますが、遺伝子発現や細胞レベルでのある種の変化を誘導するかもしれません。有性化物質は扁形動物のサブグループに特異的で、プラナリア類、単生類、そして吸虫類の間で保存されている可能性があります。しかし、単生類と吸虫類の約77%は寄生性扁形動物であることから、ヒトの健康および家畜・魚類生産に関連した多大な損害を引き起こします。そのため、本研究成果は多くの寄生性扁形動物に応用可能であることから、ある種の扁形動物の生殖生物学に関する知見を研究者が他に応用するための合理的な根拠を示しています。
以前の陸生プラナリアであるオオミスジコウガイビルの研究で、完全有性化効果は高速液体クロマトグラフィーで分画したトリプトファンのピークと続く溶出画分との混合画分で観察されました。しかし、さらなる分画によって、完全有性化効果は最終的に消失し、いくつかの画分で卵巣を誘導しただけでした。このことから、2つの可能性が考えられます。1つ目は、有性化物質は付加的かつ協調的に働く複数の卵巣誘導物質から構成されていることです。2つ目は、ただ一つの決定的な有性化物質が存在しますが、実験に使用された画分に含まれる量が卵巣誘導しかできないほどに少なかった可能性です。オオミスジコウガイビルの研究では、大量のトリプトファンから有性化物質を分けるための最初の抽出段階で酢酸エチルを使用したことにより、おそらく有性化物質の量が減少したと考えられるため、後者の可能性は高そうです。(補足:酢酸エチルを最初に使用することで、油に溶けやすい化合物が酢酸エチルに溶けてしまう。水と油のどちらにも溶けるような化合物であれば、一部が酢酸エチルに溶けてしまうと考えられる。)さらに、オオミスジコウガイビルは産卵期に野外で採集しなければならず、有性化効果はおそらく野外環境の違いによりバッチ間でばらつきがあり不安定です。本研究では、最初に水で溶出することで有性化物質を効率的に確保する新しい方法を確立しました。一緒に溶出された大量のトリプトファンはオープンカラムによって分けられました。付け加えて、双口吸虫を使用することにより、M30画分は、その後の高速液体クロマトグラフィーによる分画で活性を失わないほどに十分に強い有性化効果を持つことになったと考えられます。しかし、効率的な高速液体クロマトグラフィーによる分画では一貫したピークは観察されず、単離と構造予測のためにはまだ多くの量を確保する必要があります。反対に、これは、有性化物質は微量でも効果があることを意味しています。給餌実験で使用された濃度は、完全有性化効果を示す画分に大量に含まれるトリプトファンの濃度を基に決められましたが、その濃度範囲でさえ単一の有効物質が見つかりませんでした。このことから、有性化物質は、本研究のメタボローム解析では検出されない未知の物質である可能性が明らかになりました。しかし、現段階では有性化物質が同定されない限り1つ目の可能性を排除することはできません。本研究では、18種の卵巣誘導物質を同定しました。今後の重要な方向性は、本研究ではできませんでしたが、いくつかの物質を混合した時に有性化効果が増強されるかどうかを確認することです。給餌実験は簡単で実施しやすいですが、短所のひとつとして、複数物質の付加的かつ協調的効果を確かめることが難しいことが挙げられます。これは、おそらく味のバランスが変化するために、複数の物質を添加したエサを食べることをプラナリアが嫌がるためです。今後は、扁形動物に含まれるそれぞれの卵巣誘導物質の相対比を考慮した入念な給餌実験が、混合物質の協調的効果を確かめるためには必要になるでしょう。
18種の卵巣誘導物質のうち、6種はプリン体代謝に関連していました。トランスクリプトーム解析の結果と合わせると、プリン体代謝は紛れもなく重要な役割を担っており、この段階で、3つの潜在的な作用メカニズムが考えられます。1つ目として、完全有性化効果を担う決定的な物質との構造活性相関のために、代謝物は間接的に作用する可能性があります。(補足:構造活性相関とは化学物質の構造と生物学的な活性の間にある量的関係のこと。)2つ目として、代謝物は、プリン代謝のバランスを有性状態へと変化させることでプラナリアの生理に影響を与え、不十分ながらも決定的な物質を生産されるようになった可能性があります。これらの場合では、そのような決定的な物質は扁形動物のサブグループに特異的で、本研究で使用されたメタボロームデータベースからでは同定できない未知のプリン体代謝物である可能性があります。例えば、キサントシンは、カフェインやテオブルミンのようなプリン体アルカロイドの重要な前駆体です。これらはメッセンジャー分子としてアデノシンと構造的に似ておりアンタゴニストとして作用します。3つ目として、キサントシン、GTP、GDPは多くの生体内作用と関連しているため、そのような代謝物は性成熟のシグナル経路に直接関わる可能性があります。例えば、乳腺上皮細胞をキサントシンで処理すると、対称的細胞分裂が促進され、乳腺幹細胞の数が増加することが最近報告されましたが、その分子メカニズムはまだ分かっていません。GTPとGDPはGたんぱく質と結合し、活性状態と非活性状態を切り替え、Gたんぱく質共役受容体の機能に影響を与える重要な分子です。多様なリガンドと結合することで、Gたんぱく質共役受容体はシグナルを細胞外環境から顆粒のシグナルカスケードへ伝え、成長、分化、増殖などの多くの生体内作用で必須の役割を担っています。
完全有性化効果の欠如と矛盾なく、条虫類でのプリン体代謝の低さは、いくつかの寄生虫で高速液体クロマトグラフィーを使用してリボヌクレオチド量を比較した研究から推測されます。肝蛭のアデニン(AMP+ADP+ATP)とグアニン(GMP+GDP+GTP)の核酸量は生重量あたりでそれぞれ3.07 ± 0.50 mmol/gと 0.68 ± 0.18 mmol/gでしたが、条虫類の拡張条虫ではそれぞれ1.72 ± 0.41 mmol/gと0.34 ± 0.10 mmol/gでした。寄生性扁形動物の性成熟でのプリン体代謝の重要性は吸虫類の日本住血吸虫で示されています。日本住血吸虫の成長、発生、増殖は免疫不全の哺乳類の中では停滞し、産卵が低下することが知られています。重症複合免疫不全マウスから採取した日本住血吸虫と正常マウスから採取した日本住血吸虫の代謝プロファイルを比較することで、オスとメスの両方で相違のある代謝物のセットの一つとしてプリン体代謝が浮かび上がったことが報告されています。プラナリアでの有性化物質とプリン体代謝の機能を明らかにするために、例えば、生物活性物質に関連する遺伝子制御に注目して、今後の研究を進めることで、寄生性扁形動物の性成熟についてより深く、包括的に理解することに繋がります。
これまでの研究で、D-トリプトファン、L-トリプトファン、そしてセロトニンが卵巣誘導物質として同定されました。本研究で同定された5-ヒドロキシトリプトファンを合わせると、トリプトファン代謝から、計4種の卵巣誘導物質が同定されています。プラナリアの有性化でのプリン体代謝とトリプトファン代謝の間の直接的な関係はまだ分かっていません。これまでの有性と無性のプラナリアを比較したトランスクリプトーム解析では、有性化によって発現が変化した10,059の遺伝子配列群を同定し、トリプトファンが卵巣誘導物質として同定されていたことから主にトリプトファンを含むアミノ酸代謝に注目していました。しかし、実際には、注目されていませんでしたが、プリン体代謝経路も有性プラナリアで見つかっていました。本研究では、卵黄腺に注目して複合卵のトランスクリプトーム解析を行ったところ、プリン体代謝が唯一の代謝経路として絞り込まれたことから、有性化物質を考える上で重要であることが分かりました。本研究の高速液体クロマトグラフィーのデータ(図5B)とこれまでの研究から、大量のトリプトファンによって覆い隠されていた決定的な有性化物質の存在が考えられ、それは扁形動物のサブグループに特有な未知のプリン体代謝物である可能性があります。本研究の給餌実験によって、卵巣誘導物質として様々な代謝物が同定されていたことから、異なる物質が協調して有性化に作用しているようにみえ、トリプトファンはそのような物質の代表格であると考えられます。現状では、プリン体代謝とトリプトファン代謝との関係に2つの可能性が考えられます。1つ目として、トリプトファンの近くで溶出される決定的な有性化物質はプリン体アルカロイドのようなプリン体代謝物である可能性があります。2つ目として、プリン体代謝物もトリプトファン代謝物と協調して働いている可能性があります。興味深いことに、精神障害のある患者では、トリプトファン代謝とプリン体代謝に変化が起きることがあります。無性プラナリアの体構造は主に消化器系と脳を含む神経系からなり、単純です。地中海プラナリアでは、神経ペプチドのNPY8とその受容体であるGタンパク質共役受容体が性成熟の中心的制御因子であることが報告されているため、神経系の機能は有性化過程で重要であると考えられます。卵巣誘導物質であるセロトニンを含むトリプトファン代謝物と本研究で同定されたプリン体代謝物も、扁形動物の性成熟中の神経系でそのような役割を持っている可能性があります。
[限界]
プラナリアを有性化させる物質は寄生性扁形動物に存在しました。オオミスジコウガイビルと寄生性扁形動物である双口吸虫に含まれる有性化物質は、トリプトファンのすぐ近くに溶出し、これまでは大量のトリプトファンによって隠されていた同じ化合物であるように思えます。しかし、精製後にNMRなどによって構造を推定しない限り、プラナリアと寄生性扁形動物の間で保存されている同一物質であると結論することはできません。トランスクリプトーム解析とメタボローム解析によって同定された化合物はデータベースに登録されており既に知られている注釈付きのものに限られています。未知の化合物は本研究の方法では認識されることはありません。本研究で使用されたオープンカラムと高速液体クロマトグラフィーを組み合わせた方法に基づいたさらなる精製によって、考察で提案したように、独自に進化した扁形動物のサブグループ特異的な未知の有性化物質を単離、同定することが可能になると思われます。
また、寄生性扁形動物が自身の性成熟のために有性化物質を利用しているかどうかを確かめるためにさらなる研究が必要です。ビタミンCを添加することで、吸虫類のマンソン住血吸虫の培養液が改善されることが最近明らかになったように、ビタミンCを含む本研究で同定された18種の卵巣誘導物質は、試験管内での寄生性扁形動物の性成熟のための培養液を改善する可能性があります。これら卵巣誘導物質と本研究で分離された高速液体クロマトグラフィーによる画分が試験管内での性成熟にどのように影響するかについてのさらなる研究は、寄生性扁形動物の有性化物質の役割を明らかにするでしょう。
今後、有性化物質が同定されることで、これまでに報告されているプラナリアと寄生性扁形動物との間にある幹細胞の類似性に加えて、扁形動物間の生殖様式制御の類似性を裏付けることになるでしょう。このことから 、性発生を開始するための有性化物質を介して、幹細胞の挙動を制御する共通の分子メカニズムが存在すると考えられます。本研究は、例えば生理学的調節因子としての新規プリンアルカノイドに基づいた抗寄生虫薬の開発など、今後の寄生虫学研究とその活用のために、寄生性扁形動物の生活環を理解し制御するためのプラットフォームを提供します。
いいなと思ったら応援しよう!
