
ホウセキカナヘビの着色は、セルオートマトンによって生成されます

https://elementy.ru/novosti_nauki/432997/Okraska_glazchatykh_yashcherits_porozhdaetsya_kletochnym_avtomatom 2017年5月4日
TATYANA ROMANOVSKAYA

動物の色は奇妙で複雑です。細胞レベルで、その発生は微分方程式系を用いた反応拡散モデルによって記述されます。スイスの科学者チームが、ホウセキカナヘビの着色のメカニズムを詳細に研究しました。これは、個々のトカゲの皮膚の鱗がオートマトン細胞として機能する(個別のセルオートマトンに特徴的な規則に従う)ことが判明しました。数学的モデリングにより、反応拡散系が特殊な条件でセルオートマトンを生成できることがわかりました。この場合は、サイズも、トカゲの皮の厚さも適切な鱗境界にあります。
脊椎動物の皮膚の色は、しばしば奇妙に見え、縞模様、斑点、カール、迷路などの形の複雑なパターンを形成します。それは、誕生で形成される場合もあれば、生涯を通じ変化を続ける場合もあります。色の形成は、細胞膜の直接接触、または細胞間媒体を伝播する可溶性シグナルの助けを借りて起こり、短距離と長距離の両方で,細胞間の相互作用に依存します。
爬虫類(魚も)は、さまざまなパターンと色を示すため、皮膚着色のメカニズムを研究するための便利なモデル対象になります。これらの色は、いくつかの種類のセルの組み合わせによって生成されます。色素胞細胞には色素が含まれて:メラノフォアの黒茶色のメラニン、キサントフォアの黄色のプテリジン/カロテノイド、および、エリスロフォアの赤のプテリジン/カロテノイド。グアニンのナノ結晶の規則格子を含み、干渉により構造色を発するイリドフォア細胞もあります。皮膚の色の分布は、色素胞とイリドフォアがさまざまな部分でどのように組み合わされているかによって異なります。
ホウセキカナヘビ(Timon lepidus)は、背中の模様が生涯を通し変化し続けるため、科学者にとって興味深いものです。卵から孵化した直後、これらのトカゲは茶色で、数十個の白い斑点があります(図1)。その後、ほとんどの鱗は緑色になり、一部は黒色になります(主に白い斑点の周辺に沿って元々局在していたもの)。さらなる発展過程で、鱗が2つの状態の間で切り替わり、色が緑から黒に、またはその逆に変わることがあります。その結果、トカゲの背中に黒い背景に緑の迷路のような模様が形成されます。
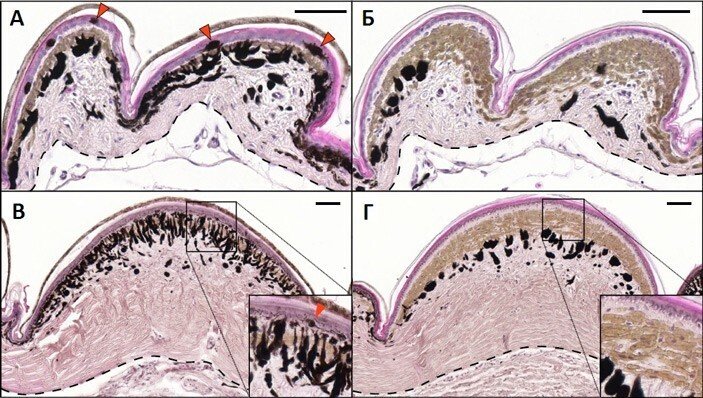
特に注目に値するのは、この種のトカゲの一つの鱗(背面)内で、異なる色の領域が決して融合しないという事実です。鱗は完全に緑または完全に黒で、鱗の全領域にわたって、色素胞の分布は完全に均一であり、隣接する鱗は、色が同じ、または、色が異なるのいずれかです。 図2は、断面における若い動物および成体動物の鱗の組織構造を示します。
このような珍しいパターンを生成するための内部機構と、鱗の色を変更するためのルールは何でしょうか?フィールズ賞を受賞した数学者スタニスラフ・スミルノフを含む研究者の注目を集めたのはこの疑問でした。
著者らは、成虫のトカゲの鱗の離散的な着色を決定する機構を解明するために、若年期から成体への着色移行を追跡しました。科学者たちは動物の写真を撮り、生後2週から3〜4歳で、数週間から3か月の間をあけて、10の時点で色素の分布の特徴を計算し、各トカゲの何千もの鱗を調べました!
トカゲが成長しても、鱗の数と相対的な位置は変化せず、各鱗のサイズは大きくなるだけです。これにより、科学者は個々の鱗の「色の履歴」を追跡することができました。観察期間(最長4年)の間に、各トカゲの約1500の鱗が色を変えました。
鱗が形成するパターンを説明するために、論文著者は確率的セルオートマトンのモデルを使用しました。鱗は六角形で、それぞれのまわりに緑または黒の6つの鱗があります。モデルは、鱗が1つの色または別の色を獲得する確率は、その隣にある同じ色の鱗の数に依存すると想定しています。モデルで得られたデータは、図3に示すように、トカゲの鱗の色がどのように変化したかと非常によく一致しました。 平均して、大人では、4つの黒と2つの緑の鱗が各緑の鱗の周りにあります。各時点で鱗の色が緑または黒から反対に変わる確率は、特定の方法で、前の時点で鱗に隣接した同じ色の鱗の数に依存します。鱗の色の変化は、徐々に頻度が少なくなるものの、動物の生涯を通じて継続し、鱗の色の変化の頻度のこの減少は、セルオートマトンモデルの予測とも一致していました。

動物の皮膚における色素細胞の分布は、遠距離と短距離での色素胞間の相互作用によって決定されます。 これらの相互作用のいくつかは活性化し、いくつかは抑制されています(図4)。 細胞は、アポトーシス、混合または分化によって、それら自身がシグナルを出し受信応答をします。

さらなる研究は、ホウセキカナヘビの背中で観察された色の分布がこれらの相互作用のシステムからどのように生まれるかを理解することを目的としていました。すなわち:
1)なぜ各鱗は厳密に均一な色となるのか?
2)隣接する鱗の色の違いの原因は何か?
3)各鱗の色を変える確率を決定する特定のセルオートマトンの規則を決定するものは何か?
4)最終的な迷路パターンは巨視的なスケールでどのように生成されるか?
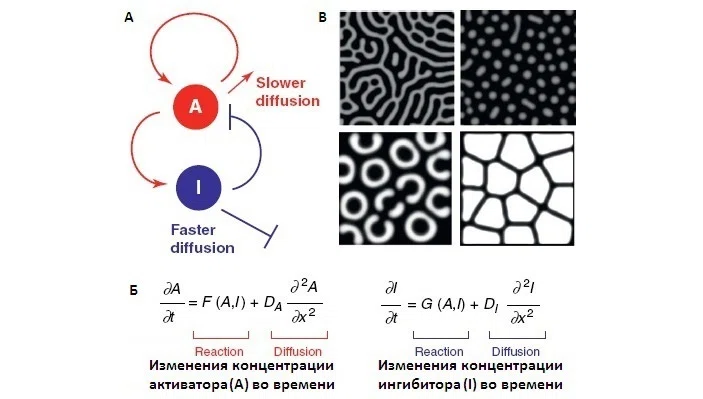
クロマトフォア間の相互作用の既知の構造に基づいて、チューリング反応拡散モデルのフレームワーク内でこのシステムの動作を記述する微分方程式系を構築することが可能です(反応拡散システムを参照)。この相互作用の構造(図4に示されている)と連立方程式は、ダニオ・レリオ(ゼブラフィッシュ)の腹部に縞模様が形成される機構を研究する過程で確立されていました。著者らは、これらのデータと方程式を使用して、ホウセキカナヘビの皮膚の色を解析しました。
チューリングのモデルは、シグナルの局所的な集中の変化によるパターンの形成を説明しています。シグナルの局所濃度は反応拡散方程式で表され、シグナルの出現率、劣化率、拡散率の比率に依存します。空間内のシグナルの不均一な分布は、異なる速度で伝播(拡散)する2つ(またはそれ以上)の反応生成物の相互作用を活性化および阻害するシステムによって引き起こされます。分布は、反応拡散方程式の定数のさまざまな比率に対してさまざまなパターンを示します(図5)。このモデルは、生物の発達中の形態形成の説明に際し、その価値を繰り返し証明してきました。この場合の活性化因子と阻害因子の役割は、細胞膜に固定されたシグナル分子(短距離接触相互作用)または拡散モルフォゲン分子(長距離シグナル)によって果たされます。
著者らは、鱗の色の分布が皮膚組織の形状によって決定され、1つの鱗内とその境界でのシグナル伝達率の違いの結果であるという仮定を、数学的モデリングの助けを借りて提唱し確認しました:第一に、皮膚の厚さの違いによるもの、第二に、鱗の端と分割領域での色素胞細胞の数の減少によるものです(これは図2に明瞭に見られます)。これらの2つの条件に基づいて、色素胞の接触相互作用を介して送信されるシグナルの絶対量と相対量は、境界領域で大幅に減少すると想定できます。これらの要因は、反応拡散方程式系に適切な修正を加えることによって考慮できます。その結果、係数の特定の値で、鱗の離散的な色がモデルに表示され(図6A)、鱗の面積が特定の閾値を超えると、この離散性は破れます。ホウセキカナヘビの尾にある大きな長方形の鱗は、背中の皮膚にある鱗よりもはるかに大きく、実際には個別の色を失っています。一部の尾の鱗は、部分的に黒と部分的に緑に着色されています(図6, Б, B)。

したがって、新しいモデル対象(ホウセキカナヘビ)の研究では、反応拡散チューリングモデルと環境の不均一性(組織の幾何学的パラメーター)の組み合わせにより、システムの動作が生成されることがわかりました。セルオートマトンの特徴。皮膚の鱗の分布そのものが、反応拡散関係の産物である可能性もあることにも注意してください。これは、セルオートマトンが、2つの反応拡散システムの重ね合わせの結果である可能性があることを意味します。卵形のトカゲの皮膚組織の形成。このスキームは、生物の形態形成中の他のパターン形成の場合にも起こり得ることが非常に期待されています。
出典:Liana Manukyan、Sophie A. Montandon、Anamarija Fofonjka、Stanislav Smirnov、MichelC.Milinkovitch。
皮膚の鱗でできた生きているメゾスコピックセルオートマトン
Nature. 2017. V. 544. P. 173–179. DOI: 10.1038/nature22031.
この記事が気に入ったらサポートをしてみませんか?