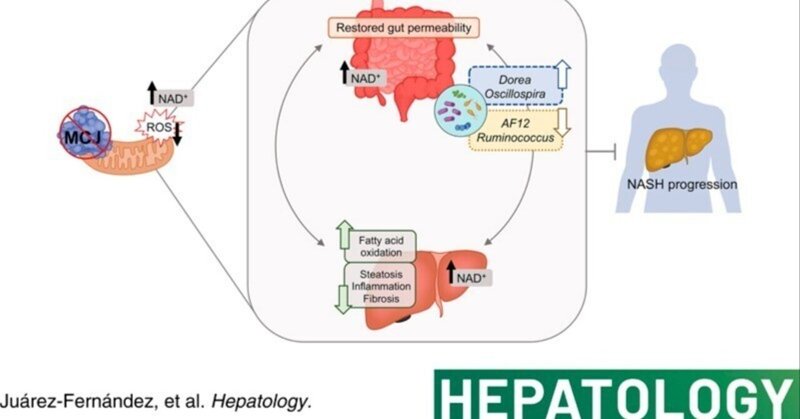
ミトコンドリア活性の亢進がNASHの進行を遅らせる腸内細菌叢プロファイルを再構築する

肝臓学(マサチューセッツ州ボルチモア)
肝臓学 2023 May; 77(5): 1654-1669.
オンライン公開 2023 Apr 17.
PMCID: PMC10113004
PMID: 35921199
ミトコンドリア活性の亢進がNASHの進行を遅らせる腸内細菌叢プロファイルを再構築する
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10113004/

マリア・フアレス-フェルナンデス1,2 ナロア・ゴイコエチア-ウサンディサガ3 ダビッド・ポラス1 マリア・ヴィクトリア・ガルシア-メディアヴィラ1,2 ミレン・ブラボー3 マリナ・セラノ-マシア3 ホルヘ・シモン2,3 テレサ・C. Delgado、3 Sofía Lachiondo-Ortega、3 Susana Martínez-Flórez、1 Óscar Lorenzo、4,5 Mercedes Rincón、6 Marta Varela-Rey、2,3 Leticia Abecia、7,8 Héctor Rodríguez、7 Juan Anguita、7,9 Esther Nistal、1,2 María Luz Martínez-Chantar、2,3 Sonia Sánchez-Campos1,2
著者情報 論文ノート 著作権およびライセンス情報 免責事項
関連データ
補足資料
データの利用可能性に関する声明
こちらへ
背景と目的
最近の研究から、ミトコンドリア機能障害が腸肝状態を悪化させることによりNASHへの進展を促進することが示唆されている。しかし、その根本的なメカニズムは不明である。ここでは、ミトコンドリア活性の亢進が特定の微生物叢シグネチャーを再構築し、それを無菌(GF)マウスに移植することでNASHの進行を遅らせることができるかもしれないという仮説を立てた。
アプローチと結果
野生型マウスおよびメチル化制御J蛋白ノックアウト(MCJ-KO)マウスに、対照食またはコリン欠乏L-アミノ酸定義高脂肪食(CDA-HFD)を6週間与えた。各群のマウス1匹は、同じくCDA-HFDモデルを3週間摂取したGFマウスへの腸内細菌叢のドナーとして機能した。その後、肝障害、腸管バリアー、腸内細菌叢、および関連する糞便メタボロームを調べた。6週間のCDA-HFDの後、ミトコンドリア複合体I活性阻害剤であるメチル化制御Jタンパク質の非存在は、攻撃的NASH食餌モデルにおいて肝障害を軽減し、腸-肝軸を改善した。この効果は、GFマウスへの腸内細菌叢移植によってもたらされた。我々は、Dorea属とOscillospira属の糞便中相対存在量の増加とAF12、Allboaculumおよび[Ruminococcus]の減少によって特徴づけられるMCJ-KOの特異的な微生物叢プロファイルが、短鎖脂肪酸、ニコチンアミドアデニンジヌクレオチド(NAD+)代謝およびサーチュイン活性を増強することによって保護作用を発揮し、その後、GFマウスにおいて脂肪酸酸化を増加させたことを示唆した。重要なことは、ドレア属がこの微生物叢依存的な保護表現型の主要な調節因子のひとつであることを同定したことである。
結論
全体として、我々は、NASHにおけるミトコンドリアと微生物叢の相互作用の関連性と、それを標的とすることが貴重な治療アプローチになりうるという証拠を提供した。
次へ
要旨
本文へ
はじめに
ミトコンドリアの機能障害は、酸化ストレスや腸内細菌叢の変化とともに、NASHへの進行を促進する1。実際、NAFLDでは腸-肝臓軸が障害されることが知られているが2、NAFLDやNASHに関連する特定のマイクロバイオームシグネチャーは完全には解明されていない3。
最近の研究で、ミトコンドリアの機能障害と腸内細菌叢の双方向の相互作用の可能性が明らかになった4。ミトコンドリアの機能障害とその結果生じる活性酸素種(ROS)の増加は、腸管バリアの完全性を調節することによって腸内細菌叢に影響を与え、それによって免疫反応を引き起こす可能性がある。反対に、腸内細菌叢はミトコンドリアの生合成を制御することが知られており、微生物の代謝産物もミトコンドリアの呼吸に影響を与える可能性がある5。したがって、ミトコンドリアの機能障害を標的とすることは、腸-肝軸を適切に維持するための防火壁となり、それによってNASHの進行を遅らせたり、あるいは予防したりする可能性がある。
メチル化制御Jタンパク質(MCJ)、またはDnaJC15は、ミトコンドリア複合体I6の内因性ネガティブレギュレーターである。MCJの欠損は呼吸とアデノシン三リン酸(ATP)合成を増加させ、呼吸性超複合体の形成を刺激し、酸化ストレスの産生を制限する。 -10さらに、MCJ欠損は潰瘍性大腸炎における宿主ミトコンドリアと腸内細菌叢の間の制御関係を変化させ、疾患の重症度に影響を及ぼす。したがって、Mcj欠損マウスは、NAFLDおよびNASHにおける腸内細菌叢の異常および腸-肝軸の変化に対する肝ミトコンドリアの寄与を研究するための信頼できるモデルである。
本研究では、NASHの食餌性モデルマウスにおいて、腸内細菌叢組成の変化を原動力として考え、MCJ欠損の効果を評価し、さらにメチル化制御Jタンパク質ノックアウト(MCJ-KO)マウスで観察された肝保護表現型が、セカル微生物叢移植(CMT)を通じて無菌(GF)マウスに移植可能かどうかを明らかにすることを目的とする。
次へ
方法
実験デザイン
すべての手順は、ARRIVEガイドラインおよびEuropean Research Council guidelines for animal careに従って行われ、CIC bioGUNEおよびUniversity of Leónのinstitutional animal care and use commitees(OEBA-ULE-003-2018 and CIC bioGUNE CBBA/IACUC [REGA 48/901/000/6106, P-CBG-CBBA-218] )により、管轄当局(それぞれJunta de Castilla y LeónおよびDiputación de Bizkaia)の承認を得た。CIC bioGUNEはAAALAC Intlの認定を受けている。
Aモデル:ドナーマウスの選択
野生型(WT)(n = 14)とMCJ-KO(n = 19)のC57BL/6バックグラウンドの6週齢の雄マウスをCIC bioGUNE動物施設で飼育した。対照食または脂肪60%kcal、メチオニン0.1%、コリン欠乏高脂肪食(CDA-HFD)(A06071302 Research Diets)を6週間摂取させた。その後、マウスをイソフルラン2.5%で麻酔し、心臓穿刺による失血で安楽死させた。嚢内容物は、還元剤としてシステイン(0.5 g/L)を添加したスキムミルク(10%)に直ちに保存し、液体窒素に浸漬し、微生物叢移植まで-80℃で保存した。モデルAの各実験群から1匹のマウスを、表S1に規定されたパラメーターに基づいて腸内細菌叢ドナーとして選択した。
Bモデル:腸内細菌叢の移植
6週齢のGF雄性C57BL/6Jマウス(n = 60)を、レオン大学の病原体フリーの特定動物施設で飼育した。すべてのマウスは温度、湿度、照度を一定に保ち、放射線照射した餌とオートクレーブ処理した水を自由摂取させた。馴化期間後、ドナーの糞便内容物100μlを1回経口胃内投与して移植した。マウスは、移植された4種類のドナーの微生物叢と食餌に基づいて8つのグループに分けられた(図S1)。3週間後、すべての動物をイソフルラン麻酔下で心臓穿刺により安楽死させた。
サンプル採取
モデルAおよびBのマウスの肝臓、腸、褐色脂肪組織および精巣上体白色脂肪組織(WAT)、ならびに糞便および糞便内容物を採取し、直ちに液体窒素に浸し、-80℃で保存して後日分析した。血液を採取し、6000rpmで10分間遠心して血漿を得た。
に進む:
結果
痩せ型NASHマウスモデルにおいて、肝ミトコンドリアは腸管バリアの完全性を改善し、特定の腸内マイクロバイオームシグネチャーを推測した。
WTマウスではCDA-HFDを6週間続けると腸管透過性が増加し、コントロールと比較して血清中のフルオレセインイソチオシアネート(FITC)-デキストラン濃度が上昇した(図1A)。興味深いことに、CDA-HFDを与えたMCJ-KOマウスではFITC-デキストラン透過性が有意に低下しており、保護効果の可能性が示唆された(図1A)。組織学的評価では、回腸組織の粘膜の厚さ、陰窩の深さ、および絨毛の高さに有意差は認められなかった(図S3A)。
図1
メチル化制御Jタンパク質ノックアウト(MCJ-KO)遺伝子型が腸-肝臓軸と微生物叢組成に及ぼす影響。(A)フルオレセインイソチオシアネート(FITC)-デキストラン血清レベル。(B)オクルジンとZonula occludens 1(Zo-1)の腸管メッセンジャーRNA(mRNA)の相対発現。(C)ZO-1染色回腸切片の定量と代表例。(D) 回腸におけるオクルディンレベルのデンシトメトリー解析。WT群(C/WT)に対する相対値で表した。(E)血漿中のリピドA濃度のデンシトメトリー解析。コリン欠乏、L-アミノ酸欠乏、高脂肪食(CDA-HFD)/野生型(WT)に対する相対値で表した。(F)IL-6およびTNF血漿中濃度。(G)NOD、LRR-およびピリン・ドメイン含有タンパク質3(Nlrp3)およびToll様受容体-4(Tlr-4)の肝mRNA相対発現。(H)操作分類単位(OTU)レベルでのBray-Curtisの非類似度指数に基づく主座標分析プロット。コントロール(I)とCDA-HFD(J)を与えたWTマウスとMCJ-KOマウスの属レベルでの相対存在量の違い。(K)WTマウスとMCJ-KOマウスの属および種レベルでの線形判別分析の効果量(閾値=2.0、p<0.05)。各実験群で少なくともn = 6を用いた。*p < 0.05, **p < 0.01, ***p < 0.001 vs C/WT; # p < 0.05, ## p < 0.01 vs CDA-HFD/WT
タイトジャンクションタンパク質の解析から、オクルディンおよびZonula occludens 1(Zo-1)のメッセンジャーRNA(mRNA)レベルの有意な減少(図1B)、クローディン-1陽性免疫染色(図S3B)、ZO-1陽性免疫蛍光(図1C)が明らかになった、 およびオクルジンタンパク質レベル(図1DおよびS3C)は、CDA-HFDを与えたWTマウスでは、コントロールおよびCDA-HFDを与えたMCJ-KOマウスの両方と比較して、MCJ欠損マウスでは接合部の完全性が改善されていることが確認され、細菌および微生物産物の移動を回避している可能性がある。
血清中のエンドトキシンおよび炎症性サイトカインを測定したところ、CDA-HFD/KO群ではWT群と比較して、リポ多糖の構成成分であるリピドA血漿タンパク質濃度が有意に低下し(図1EおよびS3D)、炎症性IL-6およびTNF血漿濃度も低下した(図1F)。
腸-肝臓軸のシグナル伝達に関しては、抗菌ペプチドの分泌を制御する核内胆汁酸受容体であるファルネソイドX受容体の腸内mRNA発現は、コントロールおよびCDA-HFD/KO群と比較して、CDA-HFDを与えたWTマウスで増加し、Gタンパク質共役型胆汁酸受容体-1の発現には変化が観察されなかった(図S3E)。さらに、CDA-HFD摂取により、腸内細菌叢由来の肝毒性または免疫刺激性化合物のセンサーであるToll様受容体-4(Tlr-4)およびNOD、LRR、およびピリンドメイン含有タンパク質3(Nlrp3)の肝臓mRNAレベルが上昇したが、Mcj欠乏はNlrp3のアップレギュレーションを抑制した(図1G)。
以上のことから、MCJの欠損は腸管バリアーの完全性の喪失に対抗し、細菌産物の移行を減少させ、NASHにおけるインフラマソーム反応を調節する。
我々のグループは、MCJの欠乏がNAFLDを改善することを以前に証明した9。以前の研究で、CDA-HFDはリーンNASHの攻撃的モデルであると報告されているので12、我々はこの食餌下での脂肪肝疾患におけるMCJの役割の解析を進めた。まず、CDA-HFD後の肝組織学的解析では、炎症巣を伴う高度の微小小胞および大小胞脂肪症が認められた(図S2A)。CDA-HFDは血清肝アミノトランスフェラーゼ値も上昇させた(図S2B)。重要なことは、CDA-HFDで飼育したMCJ-KOマウスでは、体重に対する肝臓重量およびWAT重量の比がWTマウスに比べてともに低かったことである(図S2C)。その上、MCJの欠乏は、C-Cケモカイン受容体5(Ccr5)、Il-1β、Il-6およびTnfのような主要な炎症性サイトカインの肝mRNA発現を有意に減少させた(図S2D)。肝線維化に関しては、MCJ-KOマウスでは、シリウスレッド染色によるコラーゲン含量の有意な減少(図S2E)と、プロフィブロゲンであるコラーゲンタイプ1α1鎖(Col1a1)とトランスフォーミング成長因子β(Tgf-β)の発現も、MCJ非存在下での線維化の顕著な減速を確認した(図S2F)。
腸内細菌叢もNAFLD感受性を決定する13。そこで、Aモデルにおける糞便中細菌叢組成を解析した。
操作分類単位(OTU)レベルでのBray-Curtis指数に基づく主座標分析(PCoA)により、細菌群集は食餌によって第1軸に沿って別々にクラスター化することが示された(F = 27.39; p < 0.0001)が、第2軸のスコアプロットでは遺伝子型によるクラスターの可能性が示唆され(F = 1.99; p = 0.082)、主に対照食摂取群で観察された(図1H)。
3週間後、CDA-HFD摂取により、シャノン多様性指数で算出した微生物叢の多様性は増加したが、MCJ欠乏では減少する傾向がみられた(図S4A)。門レベルでの腸内細菌叢プロフィールは、食餌と遺伝子型によって有意に変化した(図S4B)。食餌が腸内細菌叢組成を属レベルでも大きく変化させたにもかかわらず、MCJ-KO遺伝子型に関連した特異的プロファイルが同定された。AF12、Anaeroplasma、Christensenella、Coprococcus、[Ruminococcus]、Turicibacterの相対量は、対照食を与えたMCJ-KO群で減少したが、DoreaとOscillospiraはWTマウスと逆のパターンを示した(図1I)。さらに、CDA-HFD給餌によるMCJ-KO遺伝子型に関連した変化も観察され、Bacteroides属の増加とBilophila、Lactobacillusおよび[Ruminococcus]の減少が指摘された(図1J)。興味深いことに、MCJ欠損マウスでは、Doreaが増加し、AF12、Allobaculumおよび[Ruminococcus]が減少するというように、食餌に関係なく遺伝子型依存的な影響を示す細菌もあった(図1K)。MCJ-KO遺伝子型に関連するOTUレベルでの差異を線形判別分析の効果量分析によって調べた(図S4C)。
NASH発症におけるMCJ-KO遺伝子型に関連するこの腸内細菌叢プロフィールは、線維化の進行を回避し、腸管バリアの完全性を維持する、肝保護効果において重要な役割を果たす可能性がある。
MCJ-KOマウスからCDA-HFD飼育GFマウスへの腸内細菌叢移植は肝障害を改善する
CDA-HFD飼育MCJ-KOマウスで観察された保護効果が特定のマイクロバイオームシグネチャーによるものであり、したがって移植可能かどうかを調べるために、GFマウスにCMTを施し、CDA-HFD食を3週間与えた(図S1)。選択したドナーの個体値を表S1に示す。
CDA-HFD食は、組織学的評価によると、肝脂肪沈着、初期炎症、バルーニングを伴うNAFLDの初期段階を移植GFマウスに引き起こした(図2A)。MCJ-KOのCDA-HFDを投与したドナーマウス(dCDA-HFD/KO)を移植したGFマウスでは、炎症が抑制された(図2A)。その上、WT CDA-HFD-fedドナー群と比較して、dCDA-HGF/KO移植マウスでは肝トリグリセリド含量の減少(図2B)および血清アミノトランスフェラーゼ値の低下(図2C)も認められた。Ccr5、Il-1β、Il-6およびTnfの発現は、MCJ-KOドナーからのCMT後に有意にダウンレギュレートされた(図2D)。さらに、MCJ-KO対照食餌ドナー(dC/KO)群およびdCDA-HFD/KO受容体群のいずれにおいても、線維化の進展は遅延し、WT微生物叢移植群と比較して、α-平滑筋アクチン染色(図2E)およびCol1a1、マトリックスメタロペプチダーゼ9(Mmp9)、Tgf-βの発現(図2F)が有意に減少した。
図2
選択されたドナーからの無菌移植マウスにおけるNAFLDに関連した特徴の発現。(A)ヘマトキシリン・エオジン染色した肝臓切片(100倍)とNAFLD活性スコア(脂肪症、小葉の炎症、バルーニングの各スコアから算出)。(B)肝内トリグリセリド含量。(C)アラニンアミノトランスフェラーゼ(ALT)およびアスパラギン酸アミノトランスフェラーゼ(AST)の形質レベル。(D)炎症性C-Cケモカイン受容体5(Ccr5)、Il-1β、Il-6およびTnfの肝臓メッセンジャーRNA(mRNA)相対発現。 E)α-平滑筋アクチン(α-SMA)染色した肝臓切片の定量と代表的なもの。(F)線維化マーカーであるコラーゲンタイプ1α1鎖(Col1a1)、マトリックスメタロペプチダーゼ9(Mmp9)、トランスフォーミング成長因子β(Tgf-β)の肝臓mRNA相対発現。各実験群には少なくともn=6を用いた。p < 0.05、***p < 0.01、***p < 0.001は野生型(WT)対照食摂取ドナー(dC/WT)(対照食)に対して;# p < 0.05、# p < 0.01、## p < 0. 001対dC/WT(コリン欠乏・L-アミノ酸規定・高脂肪食[CDA-HFD]);a p < 0.05、aa p < 0.01、aaa p < 0.001対WT CDA-HFD食供与体(dCDA-HFD/WT)(CDA-HFD)
総じて、MCJ-KOドナーからのCMTを受けたCDA-HFD飼育GFマウスは、NASHの進行を抑制した。このように、MCJ-KOマウスで観察された肝保護効果は、腸内細菌叢移植によって移植可能である。
MCJ-KO微生物叢シグネチャーはCMTを通じて移植可能であり、CDA-HFD飼育GFマウスにおいて腸管バリアの完全性を維持する。
MCJ-KOドナーからのCMTが腸管バリアの完全性および腸-肝臓軸に及ぼす影響を明らかにするために、CDA-HFDを与えたGFマウスで組織学的および遺伝子発現試験を行った。回腸組織学的解析では有意差は認められなかった(図3A)。しかしながら、MCJ-KOドナーのGFマウスでは、Nlrp3およびTlr-4の肝臓mRNAレベルの低下とともに、Claudin-1およびZo-1の相対発現が増加した(図3B)(図3C)。このように、Aモデルで示されたこれらの遺伝子のアップレギュレーションを打ち消すMCJ欠損の能力は、CMT後にも観察された。
図3
メチル化制御Jタンパク質ノックアウト(MCJ-KO)遺伝子型のコリン欠乏L-アミノ酸規定高脂肪食(CDA-HFD)無胚芽(GF)マウスへの移植が腸-肝軸および腸内細菌叢組成に及ぼす影響。(A)ヘマトキシリン・エオジン染色腸切片(100倍)、組織学的評価。(B)クローディン-1とZonula occludens 1(Zo-1)の腸管メッセンジャーRNA(mRNA)の相対発現。(C)NOD、LRR、およびピリン・ドメイン含有タンパク質3(Nlrp3)およびToll様受容体-4(Tlr-4)の肝mRNA相対発現。(D)GFマウスの各ドナー群から各レシピエント群への操作分類学的単位(OTU)レベルのベン図(相対存在量>0.01%)。(E)CDA-HFD投与GFマウスのOTUレベルでのBray-Curtis非類似度指数から求めた主座標分析プロット。Aモデル(F)とCDA-HFD投与無菌マウス(Bモデル)(G)における属レベルでの相対存在量の違い。各実験群で少なくともn = 7を用いた。p<0.05、***p<0.01、***p<0.001 対 野生型(WT)対照食摂取ドナー(dC/WT)(CDA-HFD); # p<0.05、## p<0.001対WT CDA-HFD摂取ドナー(dCDA-HFD/WT)(CDA-HFD)。
ベン図を用いて、GFマウスの各ドナー群から各レシピエント群へのOTUレベルでの糞便微生物叢の移行を描いた。すべてのOTUの約50%は、すべてのドナーと対応する両方のレシピエント間で共有されたが、他のOTUは少なくとも一方のレシピエントグループで共有された(図3D)。CDA-HFDを投与したレシピエントは、対照を投与したレシピエントと比較して、シャノン指数に基づくα多様性がわずかに減少したが、この差はWTドナーの微生物叢を移植したグループでのみ有意になった(図S5A)。さらに、CDA-HFDを与えたGFマウスのOTUレベルでのBray-Curtis指数に基づくPCoAでは、第1軸に従ってドナーの遺伝子型を考慮すると、糞便微生物叢が明確に分離することが明らかになった(Fdonor_genotype = 8.29; p < 0.0001)。しかし、ドナーの食事は細菌群集を識別する別の因子として作用した(Fdonor_diet = 7.44; p < 0.0001)(図3E)。さらに、すべての実験群を比較するBray-Curtis指数に基づくPCoAも行った(図S5B)。
移植GFマウスにおける門レベルでの微生物叢組成分析は、食餌またはドナーの食餌および遺伝子型に影響された(図S5C)。Aモデルと同様に、食餌および遺伝子型は、門レベルでの腸内微生物群集構造に広範な変化を誘導した(図S4C)。属レベルでは、MCJ-KO遺伝子型(図3F)に関連する腸内細菌叢プロフィールは、CMTを介してCDA-HFDを与えたGFマウスに移行した(図3G)。Aモデルでは、AF12と[Ruminococcus]の相対量はMCJ-KO遺伝子型とともに減少パターンを示したが、DoreaとOscillospiraは逆の傾向を示した(図3F)。CDA-HFD(図3G)および対照食(図S5D)を与えたGFマウスでは、これらの属の相対存在量において同じ腸内細菌叢プロファイルが観察された。
MCJの欠乏は、CMTを介して伝達可能な肝NAD+利用可能性を増加させ、脂肪酸酸化の増強を可能にする
MCJ欠乏は脂質βの酸化を促進し、食餌誘導NAFLDモデルにおける肝脂肪症を改善する9。したがって、我々はMCJ-KOマウス(Aモデル)でも同様の効果を確認することを目的とした。CDA-HFDを投与したMCJ-KOマウスでは、脂肪酸輸送タンパク質2(Fatp2)とアシル-CoAデヒドロゲナーゼ長鎖(Acadl)の肝発現が有意に増加し(図4A)、脂質β酸化活性の亢進が認められた。さらに、還元型ニコチンアミドアデニンジヌクレオチド(NADH)と酸化型ニコチンアミドアデニンジヌクレオチド(NAD+)の比率が大きく増加すると、脂肪酸酸化が阻害されるため、ニコチンアミドアデニンジヌクレオチド(NAD+)の利用可能性は脂肪酸酸化にとって不可欠である。NAD代謝に関与する酵素の解析から、CDA-HFD MCJ-KOマウスではWTマウスに比べて肝ニコチンアミドホスホリボシルトランスフェラーゼ(Nampt)とサーチュイン(Sirt)1の相対発現が増加していることが示された(図4B)。興味深いことに、肝NAD+、NADH、およびそれらの比の測定は、CDA-HFD食MCJ-KOマウスにおいてNAD+の利用可能性が有意に増加することを示した(図4C)。
図4
AおよびB両モデルにおける肝脂肪酸酸化およびニコチンアミドアデニンジヌクレオチド(NAD+)産生。Aモデル:(A)AcadIと脂肪酸輸送タンパク質2(Fatp2)の肝臓メッセンジャーRNA(mRNA)相対発現。(B)NAD+合成酵素ニコチンアミドホスホリボシルトランスフェラーゼ(Nampt)およびサーチュイン(Sirt)1の肝臓mRNA相対発現。 C)総NAD(NADt)、還元型ニコチンアミドアデニンジヌクレオチド(NADH)、およびNAD+/NADH比の肝臓レベル。Bモデル:(D)NAD+合成酵素NamptとSirt1の肝mRNA相対発現。 E)NADt、NADH、NAD+/NADH比の肝レベル。(F)脂肪酸酸化アッセイ。(G)AcadIとFatp2の肝mRNA相対発現。各実験群で少なくともn = 6を用いた。*p < 0.05; **p < 0.01 vs コリン欠乏、L-アミノ酸定義、高脂肪食(CDA-HFD)/野生型(WT)。ピアソンの相関係数、p値、およびAモデルにおけるDoreaの相対存在量と(H)肝NAD+/NADH比および(I)Sirt1の相対mRNA発現量との間の直線関係(黒線および四角)。(J)BモデルにおけるDoreaの相対存在量とNamptおよびSirt1の肝相対mRNA発現量とのピアソンの相関係数、p値、および直線関係(青線と点)。
次に、このNAD+利用能の増加がCMTを介してCDA-HFDを与えたGFマウスにも移行したかどうかを調べることを目的とした。MCJ-KO微生物叢を摂取したGFマウスでは、NamptとSirt1の肝発現が有意に高く(図4D)、NAD+利用能も有意に増加した(図4E)。そこで、肝臓における脂肪酸酸化活性とFatp2およびAcadlの発現を測定したところ、MCJ-KO微生物叢を移植したGFマウスでは、活性が有意に上昇していることが確認された(図4F,G)。
MCJ-KOメタゲノムが肝保護効果を発揮する具体的な機序について可能性のある知見を得るために、肝、メタゲノム、メタボロミクスの結果に基づいて相関解析を行った。興味深いことに、Aモデルでは、Dorea属の相対存在量と肝の(1)NAD+/NADH比および(2)Sirt1発現量との間に正の相関が観察された(図4H,I)。その上、肝Sirt1およびNampt発現レベルもまた、BモデルにおけるDorea属の相対存在量と正の相関を示した(図4J)。
したがって、MCJ-KOマウスの肝臓で見られるNAD+の利用可能性の増加は、CMTを介して伝達可能であり、肝脂肪酸酸化を増加させ、NASスコアと肝内脂質蓄積を減少させる。
MCJ-KOマウスの特異的マイクロバイオームシグネチャーは、具体的なメタボロームプロフィールとリンクしており、腸内のNAD+生合成を促進する。
両モデル間で共通のマイクロバイオームシグネチャーを確認した後、腸内細菌叢がこのような肝保護効果を発揮している可能性のあるメカニズムを同定するために、糞便メタボロームを解析した。
Aモデルでは、主成分分析(PCA)により、第1軸(38.6%)に基づく食餌によって代謝物が明確に分離され、遺伝子型(12%)によってもわずかに分離された(図5A)。さらに、部分最小二乗判別分析を用いて、コントロール(図S6A)とCDA-HFD(図S6B)を与えたWTマウスとMCJ-KOマウスを識別する代謝物を同定したところ、MCJ欠乏症に関連する特定の代謝物プロファイルが示された。各群に対する特定の代謝物の寄与をVIP(Variables Importance in Projection)スコアで示し、コントロール食を与えた動物(図S6C)とCDA-HFDを与えた動物(図S6D)でVIPスコアの高い上位15代謝物を示した。MCJ-KOとWTの遺伝子型を識別できる代謝物のいくつかは、コントロールとCDA-HFDを与えた動物で異なっていたが、リボフラビン、イソバレリルカルニチン、ヘキソース、インドールのような他の代謝物は共通していた。食餌とは無関係にMCJ欠乏症と有意に関連した代謝物はシチジンとリボフラビンであったが、アデノシン、カルニチン、ゲンチジン酸、ヒオデオキシコール酸のような他の代謝物も遺伝子型によって変化したが、有意にはならなかった(図5B)。さらに、コントロールマウスにおけるMCJ欠損は、デオキシコール酸の有意な減少、アラントイン、カルノシン、デスチオビオチン、ソルビトールの有意な増加と関連していた(図5C)。しかし、CDA-HFD飼育マウスにおけるMCJ欠乏では、ベタイン、ビシンおよびN-アセチルノイラミン酸のわずかな増加が観察されたものの、検出された代謝物に有意な差は報告されなかったが、パントテン酸および4-アミノ安息香酸は逆の傾向を示した(図5D)。
図5
Aモデルの糞便メタボローム解析。(A)代謝物プロファイルの主成分分析。PC1とPC2の値を括弧内に示す。網掛け部分は食餌と遺伝子型によるサンプルクラスターを示す。(B)野生型マウス(WT)とメチル化制御Jタンパク質ノックアウトマウス(MCJ-KO)の食餌に依存しない代謝物の違い。(C)対照食を与えたWTマウスとMCJ-KOマウスの代謝物の違い。(D)コリン欠乏・L-アミノ酸定義・高脂肪食(CDA-HFD)を与えたWTマウスとMCJ-KOマウスの代謝物の違い。*p < 0.05. (E)主な短鎖脂肪酸(酢酸、酪酸、プロピオン酸、イソ酪酸)の糞便中濃度。(F) Doreaの相対量とアデノシン、ベタイン、リボフラビンのピーク面積のピアソンの相関係数、p値、および直線関係。各実験群で少なくともn=5を用いた。
さらに、腸内細菌叢の機能性に関連し、腸管バリアーの維持、肝脂質代謝、炎症に関与する主な代謝産物のひとつである糞便中の短鎖脂肪酸(SCFAs)を測定したところ14、MCJ-KOマウスは食餌によってその含有量が有意に増加したのに対し、CDA-HFDを与えたWTマウスはコントロールと比較してSCFAsが減少した。驚くべきことに、CDA-HFDで飼育したMCJ-KOマウスは、CDA-HFDで飼育したWTマウスと比較して、プロピオン酸およびイソ酪酸濃度が有意に増加した(図5E)。
さらに、MCJ-KO腸内細菌叢シグネチャーを特徴づける主要属の1つであるDorea属の相対存在量と、MCJ-KO遺伝子型に直接関連する代謝産物であるアデノシン、ベタイン、リボフラビン濃度との間に正の相関が認められた(図5F)。
MCJ欠損に関連する特異的なメタボローム・プロファイルが、CMTを介してBモデルにも移行可能かどうかを調べるために、移植したGFマウスの糞便メタボロームを解析した。PCAの結果、代謝物は2番目の軸(19.4%)に基づく食餌によってクラスター化したが、ドナーの食餌に関連する3番目の軸(8.8%)に沿って腸内細菌叢に起因する分布も観察された(図S7A)。さらに、CDA-HFDのメタボロームクラスタリングを同定するために2回目のPCAを行い、ドナーの食事と遺伝子型の影響を観察した(図S7B)。さらに、アデノシン、インドール、イソバレリルカルニチン、リボフラビンはdC/KOレシピエントで部分的に増加し、MCJ-KOマウスで観察されたプロファイルに類似しており(Aモデル)、CMTを介したわずかな移行効果が示唆された(図S7C)。
これらの結果から、Aモデルで観察されたMCJ欠損に関連する特異的なメタボロームは、GFマウスでは完全には移行・維持されていないことが示唆されたが、アデノシンやリボフラビンのようないくつかの代謝物パターンは依然としてすべてのGFマウス群で検出され、NAD+代謝の亢進に関与している可能性がある。
MCJ-KO微生物叢を投与したCDA-HFD飼育GFマウスで認められた肝NAD+利用能の増加が、MCJ欠乏症の腸内細菌叢シグネチャーによるものであることを確認するために、NAD+代謝に関連する主要酵素の発現と腸内のNAD+レベルを測定した。興味深いことに、MCJ-KO微生物叢を移植したGFマウスでは、NamptとSirt1の腸内発現が、腸内総NADレベルとNAD+/NADH比(図6B)とともに有意に増加した(図6A)。実際、dCDA-HFD/KO移植群では腸内のNAD+/NADH比が最も高く、NAD+の利用可能性が増加したことが確認された。興味深いことに、Doreaの相対存在量と腸内NAD総量との間に正の相関が観察された(図6C)。腸内Sirt1発現もまた、肝Sirt1レベルと同様に、Dorea属の相対存在量と正の相関が見られた(図6D)。その上、同じパターンがCDA-HFDを与えたMCJ-KOマウスのAモデルでも観察され(図6E-G)、NADシグナル伝達経路におけるこの細菌の役割の可能性を示唆した。
図6
BモデルとAモデルにおける腸内ニコチンアミドアデニンジヌクレオチド(NAD+)代謝。Bモデル、腸:(A)NAD+合成酵素ニコチンアミドホスホリボシルトランスフェラーゼ(Nampt)およびサーチュイン(Sirt)1の相対的メッセンジャーRNA(mRNA)発現。(C,D)コリン欠乏、L-アミノ酸定義、高脂肪食(CDA-HFD)を与えた動物におけるDoreaの相対存在量と(C)総NADおよび(D)Sirt1の相対mRNA発現量のピアソンの相関係数、p値および直線関係(青線および点)。モデル、腸:(E)NAD+合成酵素NamptとSirt1のmRNA相対発現。 F)NADt、NADH、NAD+/NADH比のレベル。各実験群で少なくともn = 6を用いた。*p < 0.05; **p < 0.01, ***p < 0.001 vs CDA-HFD/野生型(WT)。(G)CDA-HFDを摂取した動物におけるDoreaの相対存在量とSirt1の相対mRNA発現量のピアソンの相関係数、p値、および直線関係(黒線と四角)。
以上より、我々の研究は、Mcj欠損マウス由来の特異的マイクロバイオームシグネチャーとその特異的代謝が、食餌誘発NAFLDモデルにおいてNASHの進行を遅らせることができること、そしてこの肝保護作用がCMTを介して伝達可能であることを示している。
ヒトNAFLD患者コホートにおけるドレア属の役割の検証
さらに、Dorea属は、MCJ欠乏によってもたらされるNAFLD発症に対する保護効果における腸内細菌叢調節の役割を理解する上で、最も適切な属の一つとして際立っている。これは、MCJ-KOマウスにおけるその高い存在量とGFマウスへの完全な伝播によるだけでなく、相関解析によってこの属が観察された効果の主要な寄与者の1つであることが指摘されたためである。そこで我々は、ヒト疾患におけるこの属の関連性を検証するために、公開されているヒト情報の解析を行った。双生児と肝硬変患者のコホート15からのデータを、Leeらのアプローチに従って解析した16。患者は肥満度(BMI)により肥満(BMI≧30;n=71)と非肥満(BMI<30;n=121)に分けられ、興味深いことに、非肥満のサブセットでは、ドーリアの糞便量はNAFLDの存在により有意に低かった(p=0.026)が、肥満患者では有意差は認められなかった(p=0.636)(図S8)。これらの所見は、Dorea属の減少を特徴とする、痩せ型NASH患者における腸内細菌叢組成の特異的な変化を示唆している可能性がある。
次へ
考察
ミトコンドリア機能障害、酸化ストレス、および腸内細菌叢の変化は、その移行において重要な役割を果たす可能性がある。最近のデータによると、ミトコンドリア複合体Iの内因性阻害因子であるMCJの欠乏は、NASHにおけるミトコンドリア活性を高めることが示されている9。さらに、潰瘍性大腸炎における腸内細菌と宿主の相互作用にMCJが関与していることが以前に報告されている11。したがって、我々は、NASHにおける腸-肝臓軸のミトコンドリア活性の増加が及ぼす影響について研究し、MCJ-KOマウスで観察された肝保護表現型が微生物に関連しており、CMTを介してGFマウスのレシピエントに移行する可能性があるかどうかを調べることを目的とした。
我々の以前の知見9と同様に、6週間のCDA-HFD後にMCJを欠乏させると、炎症と線維化が抑制され、NAFLD発症におけるMCJ欠乏の潜在的な保護作用が強化された。NASHの進行における腸-肝軸の役割は深く報告されているが2、腸の状態および腸内細菌叢組成におけるMCJ-KOの影響については調べられていなかった。我々の研究において、MCJ欠損マウスは、脂肪性条件下で、主要なタイトジャンクションタンパク質の発現を増加させ、血漿中のエンドトキシン(リピドA)および炎症性サイトカイン濃度を減少させた。興味深いことに、MCJ-KOマウスは、潜在的な腸内細菌叢由来の肝毒性または免疫刺激性化合物のセンサーである肝Tlr-4およびNlrp3の発現が低下し、腸-肝軸の状態が改善した18。したがって、MCJ欠損によって改善されたミトコンドリア活性は、NAFLDの進行を特徴づける腸肝軸の変化を打ち消した。それは、腸バリアを強化し、肝炎を悪化させ、疾患の進行を加速させる細菌産物の門脈血液循環への移行を減少させたからである19。
さらに、MCJ-KOマウスは食餌とは無関係に特異的な腸内細菌叢組成を示し、Dorea属の増加とAF12、Allobaculum、[Ruminococcus]の減少を特徴とした。このような観点から、NAFLDにおけるDorea属の関与を示した研究もある3,20。しかし、いずれの場合も、肥満のある被験者を対象とした研究であり、Dorea属とBMIとの間に正の相関関係があることが示された。実際、NAFLD患者の公的コホートの解析では、非肥満のNAFLD患者では、NAFLDでない患者と比較してDoreaが有意に減少していることが示された。さらに、CDA-HFDはリーンNASHの積極的な食事モデルとして報告されており、肥満を促進することなく病態を進行させる12。したがって、我々の研究では、CDA-HFD/KOマウスにおけるDorea属の増加が観察された保護効果に関与している可能性がある。さらに、AllobaculumはNAFLDモデルマウスの高脂肪食飼育と関連していた13,24。また、RuminococcusはNAFLD発症と広く関連していた3,20。しかしながら、我々の研究において、遺伝子型に起因する特定の微小環境とその結果としての特異的代謝が、MCJ-KO依存的な方法で腸内細菌叢25を形成し、その結果、微生物叢シグネチャーを生み出している可能性があることは注目に値する。
MCJ-KOマウスの特異的マイクロバイオームシグネチャーに関連して、糞便メタボローム解析では、アデノシン、シチジン、リボフラビンが食餌とは無関係に増加したのに対し、ベタインなどの他の代謝物はCDA-HFDを与えたMCJ-KOマウスで増加する傾向がみられた。アデノシンは抗炎症作用を発揮し、腸管バリア機能の制御に関与している26。また、さまざまな研究により、活性酸素を消去し、ミトコンドリア複合体IIを保護し、ミトコンドリアグルタチオンレベルを再生するベタインの抗酸化特性とNAFLD発症における有益な役割が実証されている27,28。興味深いことに、CDA-HFDを投与したMCJ-KOマウスでは、プロピオン酸とイソ酪酸の糞便中濃度が有意に増加した。実際、SCFAによる治療は、抗酸化作用、炎症性メディエーターのダウンレギュレーション、腸管バリアの完全性の維持、ミトコンドリア呼吸能力の増加などを示している14,30,31。
重要なことに、我々の解析では、アデノシンおよびベタインと、MCJ-KO遺伝子型に関連する主要な分類群の1つであるDorea属の存在量との間に正の相関が認められた。
MCJ-KOマウスからCDA-HFD飼育GFマウスへのCMTは、肝障害を改善し、腸管バリアの完全性と腸肝軸の状態の部分的な回復を誘導し、遺伝子型の保護効果が微生物叢によって伝達されることを実証した。さらに、Aモデルで観察されたAF12、Dorea、Oscillospira、および[Ruminococcus]の相対量と同じパターンがBモデルでも検出され、肝保護効果がこの特異的なマイクロバイオームシグネチャーによって媒介されていることが裏付けられた。
次に、MCJ-KOに依存したメタボロームによって形成されたこの細菌群集が、食餌性NASHモデル後の腸と肝臓の両方において、どのように保護効果を発揮するかを理解しようとした。MCJの欠損がミトコンドリア活性とATP産生を増加させ、NAD代謝に影響を及ぼすことはすでに述べた8。MCJ-KOマウスでは、肝臓と腸管組織の両方でNAD+/NADH比の有意な増加が観察され、興味深いことに、MCJ-KO遺伝子型からCMT後のCDA-HFD飼育GFマウスでも観察された。NAD+はエネルギー代謝やミトコンドリア機能の燃料として重要な役割を果たすだけでなく、抗酸化防御システムの重要な構成要素であるNADPHの前駆体でもある37。実際、NAD+はNAFLDを予防・回復させる潜在的な標的として提案されている38。NAD+前駆体は、NAFLDの前臨床モデルにおいて、肝ミトコンドリア機能を改善し、酸化ストレスを減少させることが判明しており39、その補充はNASHへの進行も予防する40。これらの知見と一致して、CDA-HFDを投与したMCJ-KOマウス(Aモデル)およびGFマウス(Bモデル)では、肝臓のミトコンドリア機能が改善し、脂質β酸化活性が上昇した。さらに、NAD+の生合成と調節における腸内細菌叢の寄与についてもわずかに報告されている41。本研究では、肝臓と腸で観察されたNAD+レベルの増加は、MCJ-KOマウスだけでなくGFマウスでもDorea属の存在量の増加と関連していた。これまでの研究から、CDA-HFDで飼育したMCJ-KOマウスで認められたSCFAsレベルの増加が腸内代謝を促進し、その結果NAD+レベルの増加を可能にしている可能性があると考えられる。
さらに、NAD+は、SIRT38を含む様々な酵素の基質としても機能する。肝SIRT1活性は、de novo脂肪生成を抑制し、β酸化を促進する42。我々は、CDA-HFDで飼育したMCJ-KOマウス(Aモデル)、およびMCJ-KOの微生物叢を移植しCDA-HFDで飼育したGFマウス(Bモデル)において、Sirt1の肝および腸mRNAレベルが有意に高いことを観察した。実際、NAD+/NADH比の場合と同様に、2つのモデルにおいて、Sirt1発現とDorea存在量の間には、肝臓と腸組織の両方で正の相関が観察された。水素は、ミトコンドリアのNAD+産生を回復させるだけでなく、サーチュインのダウンレギュレーションを回避することができる45。
現在のところ、リーンNASHを阻止または逆転させる有効な治療法は承認されていない。リーンNASH患者の重症度とともに、その有病率の増加を考慮すると、代替治療が強く求められている。ここで、MCJ欠損とその結果改善されたミトコンドリア活性は、攻撃的なNASHリーン食モデルにおいて疾患進行を遅延させることができる特異的で保護的なマイクロバイオームのシグネチャーを再構築した。全体として、これらの結果は、NASHにおけるミトコンドリアと微生物叢のクロストークの重要性を浮き彫りにし、微生物叢移植に基づく治療戦略を指し示すものであり、異なるアプローチへの道を開くものである。
次へ
補足資料
補足資料
こちらをクリック(3.2M, docx)
閲覧はこちらから(24K, docx)
閲覧はこちら(44K, docx)
こちらへ
著者貢献
作品の構想またはデザイン: María Juárez-Fernández、Naroa Goikoetxea-Usandizaga、María Luz Martínez-Chantar、Sonia Sánchez-Campos。データの取得、分析、解釈: María Juárez-Fernández、Naroa Goikoetxea-Usandizaga、David Porras、Maria Victoria Garcia-Mediavilla、Héctor Rodríguez、Esther Nistal、Miren Bravo、Marina Serrano-Maciá、Jorge Simón、Teresa C.Delgado, Sofía Lachiondo-Ortega, Susana Martínez-Flórez, Óscar Lorenzo, Mercedes Rincón, Marta Varela-Rey, Leticia Abecia, Juan Anguita, María Luz Martínez-Chantar, Sonia Sánchez-Campos. 原稿執筆: María Juárez-Fernández、Naroa Goikoetxea-Usandizaga、María Luz Martínez-Chantar、Sonia Sánchez-Campos。実質的な改訂を行った: María Juárez-Fernández、Naroa Goikoetxea-Usandizaga、Esther Nistal、María Luz Martínez-Chantar、Sonia Sánchez-Campos。
資金提供
本研究は、Ministerio de Ciencia e Innovación, Programa Retos-Colaboración RTC2019-007125-1(Jorge SimónおよびMaria Luz Martínez-Chantar)より助成を受けた; Instituto de Salud Carlos III, Proyectos de Investigación en Salud DTS20/00138 (for Jorge Simón and María Luz Martínez-Chantar); Departamento de Industria del Gobierno Vasco (for María Luz Martínez-Chantar); Ministerio de Ciencia, Innovación y Universidades MICINN: PID2020-117116RB-I00、国家科学技術・イノベーション研究計画(Plan Estatal de Investigación Científica y Técnica and de Innovación)に組み込まれ、FEDER基金と共同出資(Maria Luz Martínez-Chantarのため)、Ayudas Ramón y Cajal de la Agencia Estatal de Investigación RY2013-13666(レティシア・アベシアのため)、BIOEF(バスク・イノベーション・健康研究財団); Asociación Española contra el Cáncer (for María Luz Martínez-Chantar); Fundación Científica de la Asociación Española Contra el Cancer (AECC Scientific Foundation) Rare Tumor Calls 2017 (for María Luz Martínez-Chantar); La Caixa Foundation Program (for María Luz Martínez-Chantar); Proyecto Desarrollo Tecnologico CIBERehd (for María Luz Martínez-Chantar). 本研究は、経済産業省/FEDER(BFU2017-87960-R; PID2020-120363RB-I0)(ソニア・サンチェス-カンポス)およびカスティーリャ・イ・レオン州(FEDER)(GRS2126/A/2020; LE063U16; LE017P20)(J.G.-Gおよびソニア・サンチェス-カンポス)からも助成を受けた。María Juárez-FernándezはMinisterio de Ciencia, Innovación y Universidades (FPU18/06257)から、David PorrasはJunta de Castilla y Leónから欧州社会基金の助成を受けている。CIBERehdはスペインのInstituto de Salud Carlos IIIから資金援助を受けている。メタボローム解析を行ってくれたMS-Omics社のDanielsen博士、メタゲノム解析を行ってくれたバレンシア大学のSCSIE Genómica Unidadに感謝する。
利益相反
Rincón博士はMitotherapeutix LLCの科学的創設者であり、Martínez-Chantar博士はMitotherapeutix LLCの顧問である。
データ利用声明
データは合理的な要求があれば入手可能である。本研究の16S配列データは、EMBL-EBIのEuropean Nucleotide Archive (ENA)にアクセッション番号PRJEB55379で寄託されている。
次へ
脚注
資金提供情報 Asociación Española contra el Cáncer; Ayudas Ramón y Cajal de la Agencia Estatal de Investigación, Grant/Award Number: RY2013-13666; BIOEF (Basque Foundation for Innovation and Health Research); Departamento de Industria del Gobierno Vasco; Fundación Científica de la Asociación Espaóola Contra el Cancer (AECC Scientific Foundation) Rare Tumor Calls 2017; Instituto de Salud Carlos III, Proyectos de Investigación en Salud, Grant/Award Number: DTS20/00138; Junta de Castilla y León (FEDER), Grant/Award Number: GRS2126/A/2020, LE017P20 and LE063U16; Junta de Castilla y León cofunded by the European Social Fund; La Caixa Foundation Program; Ministerio de Ciencia e Innovación, Programa Retos-Colaboración, Grant/Award Number: RTC2019-007125-1; 科学技術・イノベーション・大学省, 助成金/賞番号: FPU18/06257、科学技術・イノベーション・大学省MICINN統合科学技術・イノベーション国家計画、FEDER基金との共同助成、助成金/賞番号: PID2020-117116RB-I00;経済産業省/FEDER, Grant/Award Number: BFU2017-87960-R and PID2020-120363RB-I0; Proyecto Desarrollo Tecnologico CIBERehd
略語 BMI、肥満指数;Ccr5、C-Cケモカイン受容体5;CDA-HFD、コリン欠乏L-アミノ酸定義高脂肪食;CMT、セカル微生物叢移植;Col1A1、コラーゲンタイプ1α1鎖; dCDA-HFD/KO、MCJ-KO CDA-HFD投与ドナー;Fatp2、脂肪酸輸送タンパク質2;GF、無菌;MCJ、メチル化制御Jタンパク質;MCJ-KO、メチル化制御Jタンパク質ノックアウト;mRNA、メッセンジャーRNA; NAD、ニコチンアミドアデニンジヌクレオチド;NADH、還元型ニコチンアミドアデニンジヌクレオチド;Nampt、ニコチンアミドホスホリボシルトランスフェラーゼ;Nlrp3、NOD、LRR-およびピリンドメイン含有タンパク質3;OTU、操作分類単位; PCA、主成分分析;PCoA、主座標分析;SCFA、短鎖脂肪酸;SIRT、サーチュイン;Tgf-β、トランスフォーミング成長因子β;Tlr-4、Toll様受容体-4;WT、野生型;ZO-1、閉塞性ゾンデュラ1。
María Juárez-FernándezおよびNaroa Goikoetxea-Usandizaga共同筆頭著者。
この論文には補足デジタルコンテンツがあります。直接のURL引用は、印刷された本文に掲載されているほか、本誌ウェブサイトwww.hepjournal.com、本論文のHTML版およびPDF版で提供されている。
こちらへ:
参考文献
Ballard JWO、Towarnicki SG。ミトコンドリア、腸内マイクロバイオームと活性酸素。Cell Signal. 2020;75:109737. [PubMed] [Google Scholar].
また、「腸内細菌叢の動態的変化」と「腸内細菌叢と活性酸素の関係」についても研究している。メチオニン-コリン欠乏食誘発非アルコール性脂肪肝炎の発症における腸内細菌叢とメタボロームの動的変化。World J Gastroenterol. 2018;24:2468-81. [PMC free article] [PubMed] [Google Scholar].
Boursier J, Mueller O, Barret M, Machado M, Fizanne L, Araujo-Perez F, et al. 非アルコール性脂肪性肝疾患の重症度は、腸内細菌叢の代謝機能の腸内異常およびシフトと関連している。Hepatology. 2016;63:764-75. [PMC free article] [PubMed] [Google Scholar].
運動中の腸内細菌叢とミトコンドリアのクロストーク。Front Physiol. [PMC free article] [PubMed] [Google Scholar].
Microbiota-mitochondria inter-talk: consequence for microbiota-host interaction. Pathog Dis. 2016;74:ftv096. [PubMed] [Google Scholar].
(注1)この論文では、(注2)ミトコンドリアとミトコンドリアとの相互作用の解明、(注3)ミトコンドリアとミトコンドリアとの相互作用の解明を目的とした。MCJ/DnaJC15、代謝変化を制御する呼吸鎖の内因性ミトコンドリア抑制因子。2013:33:2302-14。[PMCフリー記事] [PubMed] [Google Scholar].
ナバサ N、マルティン I、イグレシアス-ペドラズ JM、ベラザ N、アトンド E、イザディ H、他。メチル化制御Jタンパク質による酸化ストレスの制御は、炎症性傷害に対するマクロファージ応答を制御する。J Infect Dis. 2015;211:135-45. [PMC free article] [PubMed] [Google Scholar].
バルビエ-トレスL、Iruzubieta P、Fernández-Ramos D、Delgado TC、Taibo D、Guitiérrez-de-Juan V、et al. ミトコンドリア陰性調節因子MCJはアセトアミノフェン誘発性肝障害の治療標的である。Nat Commun. 2017;8:2068. [PMC free article] [PubMed] [Google Scholar].
バルビエ-トレスL、フォートナーKA、Iruzubieta P、デルガドTC、ギディングスE、チェンY、ら。肝MCJのサイレンシングは、ミトコンドリアの脂肪酸酸化を増加させることにより、非アルコール性脂肪性肝疾患(NAFLD)を抑制する。Nat Commun. 2020;11:3360. [PMCフリー論文] [PubMed] [Google Scholar].
Iruzubieta P, Goikoetxea-Usandizaga N, Barbier-Torres L, Serrano-Maciá M, Fernández-Ramos D, Fernández-Tussy P, et al. MCJのサイレンシングによるミトコンドリア活性の増強は、胆汁うっ滞による肝障害を克服する。JHEP Rep.2021;3:100276。[PMC free article] [PubMed] [Google Scholar].
Pascual-Itoiz MA, Peña-Cearra A, Martín-Ruiz I, Lavín JL, Simó C, Rodríguez H, et al. ミトコンドリア陰性調節因子MCJは、潰瘍性大腸炎における微生物叢と宿主の相互作用を調節する。サイRep.2020;10:572。[PMC free article] [PubMed] [Google Scholar].
松本昌彦、羽田直樹、坂巻康弘、宇野亜由美、志賀俊一、田中千尋、他.非アルコール性脂肪性肝炎の線維化を急速に進行させる改良モデルマウス。Int J Exp Pathol. 2013;94:93-103. [PMCフリー論文] [PubMed] [Google Scholar].
ルロワT、ロピM、ルパージュP、ブルノーA、ラボS、ベビラクアC、ら。腸内細菌叢はマウスの非アルコール性脂肪性肝疾患の発症を決定する。Gut. 2013;62:1787-94. [PubMed] [Google Scholar].
アルコール誘発性肝障害のドライバーとしての腸内細菌叢異常。JHEP Rep.2020;3:100220。[PMC free article] [PubMed] [Google Scholar].
CaussyC、Hsu C、Singh S、Bassirian S、Kolar J、Faulkner C、他。血清胆汁酸パターンは、双子におけるNAFLDの存在と関連しており、生検で証明されたNAFLD患者における線維化ステージの増加に伴う用量依存的変化。Aliment Pharmacol Ther. 2019;49:183-93. [PMC free article] [PubMed] [Google Scholar].
李G、You HJ、Bajaj JS、Joo SK、Yu J、Park S、et al. 非肥満性NAFLDにおける有意な線維化と関連する腸内細菌叢と代謝産物の特徴的なシグネチャー。Nat Commun. 2020;11(1):4982. [PMC free article] [PubMed] [Google Scholar].
非アルコール性脂肪性肝疾患:NAFLDからNASHへの進行における基本的な病態機序。Transplantation. 2019;103:e1-e13. [PubMed] [Google Scholar].
Meli R, Raso GM, Calignano A. Role of innate immune response in non-alcoholic fatty liver disease: metabolic complications and therapeutic tools. Front Immunol. 2014;5:177. [PMC free article] [PubMed] [Google Scholar].
Hu H, Lin A, Kong M, Yao X, Yin M, Xia H, et al. 腸内マイクロバイオームとNAFLD:分子的洞察と治療の展望。J Gastroenterol. 2020;55:142-58. [PMC free article] [PubMed] [Google Scholar].
Velázquez KT, Enos RT, Bader JE, Sougiannis AT, Carson MS, Chatzistamou I, et al. 高脂肪食の長期摂取は非アルコール性脂肪性肝疾患を促進し、マウスの腸内細菌叢を変化させる。World J Hepatol. 2019;11:619-37. [PMC free article] [PubMed] [Google Scholar].
Gallè F, Valeriani F, Cattaruzza MS, Gianfranceschi G, Liguori R, Antinozzi M, et al. 地中海食、身体活動と腸内細菌叢組成:イタリアの健康な若い成人の間で横断的研究。栄養素。2020;12:2164. [PMCフリー記事] [PubMed] [Google Scholar].
また、「腸内細菌叢」は、「腸内細菌叢」と「腸内細菌叢」との関連性が示唆されている。肥満げっ歯類におけるインスリン抵抗性と全身性炎症に腸内細菌叢が関与している可能性:メタアナリシス。Physiol Genomics. 2018;50:244-54. [PubMed] [Google Scholar].
Ahn SB, Jun DW, Kang BK, Lim JH, Lim S, Chung MJ. Randomized, double-blind, placebo-controlled study of a multispecies probiotic mixture in nonalcoholic fatty liver disease. Sci Rep.2019;9:5688。[PMC free article] [PubMed] [Google Scholar].
サファリZ、Monnoye M、Abuja PM、Mariadassou M、Kashofer K、Gérard P、et al. 高脂肪食によって誘導されたステトーシスと腸内細菌叢異常は、1週間のチャウ食投与によって回復する。Nutr Res.2019;71:72-88。[PubMed] [Google Scholar].
Gomaa EZ. 健康と疾患におけるヒト腸内細菌叢/マイクロバイオーム:総説。Antonie van Leeuwenhoek. 2020;113:2019-40. [PubMed] [Google Scholar].
D'Antongiovanni V, Fornai M, Pellegrini C, Benevenuti L, Blandizzi C, Antonioli L. The adenosine system at the crossroads of intestinal inflammation and neoplasia. (1)腸の炎症と新生物の岐路におけるアデノシン系. [PMC free article] [PubMed] [Google Scholar].
ベタインはミトコンドリア複合体IIとグルタチオンの利用可能性を介しておそらくアセトアミノフェン肝毒性からマウスを保護する。Biomed Pharmacother. 2018;103:1436-45. [PubMed] [Google Scholar].
ベタインはミトコンドリア含量を増加させ、肝脂質代謝を改善する。Food Funct. 2019;10:216-23. [PubMed] [Google Scholar].
陳Q、王Y、焦F、石C、培M、王L、ら。ベタインは急性肝不全マウスのToll様受容体4反応を抑制し、腸内細菌叢を回復させる。(注1)本論文は、本論文の一部である。[PMC free article] [PubMed] [Google Scholar].
González-Bosch C, Boorman E, Zunszain PA, Mann GE. 健康および疾患における酸化還元シグナル伝達の調節因子としての短鎖脂肪酸。レドックスバイオル2021;47:102165。[PMC free article] [PubMed] [Google Scholar].
レオンC、リベラL、ファーネスJB、アンガスPW。NAFLDにおける腸内細菌叢の役割。Nat Rev Gastroenterol Hepatol. 2016;13:412-25. [PubMed] [Google Scholar].
Liu Y, Du T, Zhang W, Lu W, Peng Z, Huang S, et al. 黄連解毒湯は、アルツハイマー病モデルマウスにおけるAβシナプス毒性を改善する。Oxid Med Cell Longev. 2019;2019:8340192. [PMC free article] [PubMed] [Google Scholar].
Du J, Zhang P, Luo J, Shen L, Zhang S, Gu H, et al. 食餌性ベタインは、腸内細菌叢に依存したマイクロRNA-378aファミリーを介して肥満を予防する。Gut Microbes. 2021;13:1862612. [PMC free article] [PubMed] [Google Scholar].
Liao L, Schneider KM, Galvez EJC, Frissen M, Marschall HU, Su H, et al. 原発性硬化性胆管炎のマウスモデルにおいて、腸内細菌異常症はNLRP3を介して肝疾患の進行を増強する。Gut. 2019;68:1477-92. [PubMed] [Google Scholar].
Soderborg TK, Clark SE, Mulligan CE, Janssen RC, Babcock L, Ir D, et al. 肥満の母親の乳児の腸内細菌叢は、炎症とNAFLDへの感受性を増加させる。Nat Commun. 2018;9:4462. [PMC free article] [PubMed] [Google Scholar].
Wen L, Ley RE, Volchkov PY, Stranges PB, Avanesyan L, Stonebraker AC, et al. 1型糖尿病の発症における自然免疫と腸内細菌叢。Nature. 2008;455:1109-13. [PMC free article] [PubMed] [Google Scholar].
Katsyuba E, Mottis A, Zietak M, de Franco F, van der Velpen V, Gariani K, et al. デノボNAD +合成は、ミトコンドリアの機能を高め、健康を改善する。Nature. 2018;563:354-9. [PMC free article] [PubMed] [Google Scholar].
Dall M, Hassing AS, Treebak JT. NAD+とNAFLD-注意、因果関係と慎重な楽観論。J Physiol。2021;600:1135-54。[PubMed] [Google Scholar].
Ganji SH, Kukes GD, Lambrecht N, Kashyap ML, Kamanna VS. 非アルコール性脂肪性肝疾患のラットモデルにおける肝脂肪症の予防と退縮におけるナイアシンの治療的役割。2014;306:G320-7。[PubMed] [Google Scholar].
Dall M, Hassing AS, Niu L, Nielsen TS, Ingerslev LR, Sulek K, et al. マウスにおけるNAD+生合成経路の肝細胞特異的摂動は、可逆的非アルコール性脂肪肝炎様表現型を誘導する。J Biol Chem. 2021;297:101388. [PMCフリー論文] [PubMed] [Google Scholar].
哺乳類宿主のNaを増加させる細菌。細菌は、脱アミド化生合成経路に関与することにより、哺乳類宿主のNAD代謝を高める。Cell Metab. 2020;31:564-79.e7. [PMCフリー記事] [PubMed] [Google Scholar].
李Y、ウォンK、ジャイルズA、江J、リーJW、アダムスAC、ら。肝SIRT1は、線維芽細胞増殖因子21を誘導することにより、マウスの肝脂肪症を抑制し、エネルギーバランスを制御する。Gastroenterology. 2014;146:539-49.e7. [PMC free article] [PubMed] [Google Scholar].
肥満による炎症と腸機能障害における腸内細菌叢とサーチュイン。J Transl Med. 2011;9:202. [PMC free article] [PubMed] [Google Scholar].
(1)肥満と腸機能障害におけるサーチュイン1(Sirtuin 1)の役割. Sirtuin 1は潰瘍性大腸炎における小胞体ストレスを介した腸上皮細胞のアポトーシスを緩和する。World J Gastroenterol. 2019;25:5800-13. [PMC free article] [PubMed] [Google Scholar].
Niu Y, Nie Q, Dong L, Zhang J, Liu SF, Song W, et al. 水素はエネルギー代謝経路のスイッチを逆転させることでアレルギー性炎症を抑制する。Sci Rep.2020;10:1962。[PMC free article] [PubMed] [Google Scholar].
川崎病急性期における腸内細菌叢の変化の影響。PeerJ. 2020;8:e9698. [PMC free article] [PubMed] [Google Scholar].
Hepatology (Baltimore, Md.)の記事はWolters Kluwer Healthの好意によりここに提供される。
ama apa mla nlm
NCBIをフォローする
NLMとつながる
国立医学図書館
8600 Rockville Pike
ベセスダ、メリーランド州20894
ウェブポリシー
情報公開
HHS脆弱性情報公開
ヘルプ
アクセシビリティ
採用情報
NLM
NIH
保健省
USA.gov
この記事が気に入ったらサポートをしてみませんか?