微生物と内皮のクロストークによる好中球の肝臓への動員を血管交通で制御する

記事|42巻5号、112507、2023年5月30日発行
微生物と内皮のクロストークによる好中球の肝臓への動員を血管交通で制御する
https://www.cell.com/cell-reports/fulltext/S2211-1247(23)00518-1?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS2211124723005181%3Fshowall%3Dtrue
アマンダ・Z・ズコロト
ジャレッド・シュレヒト
アライン・イグナシオ
ブライアン・G・イップ
マーク・R・ギルリー
ブレドン・マクドナルド6
すべての著者を表示する
脚注を表示する
オープンアクセス公開日:2023年5月16日DOI:https://doi.org/10.1016/j.celrep.2023.112507
PlumX メトリクス
ハイライト
腸内常在菌は全身感染時に肝臓への好中球の動員を促進する
GFマウスにおける好中球の動員障害は、腸内細菌のD-乳酸によって回復される
D-乳酸は肝臓内皮を活性化し、好中球の接着分子をアップレギュレートする。
D-乳酸産生を治療的に補正することで、感染症に対する防御力が向上した
概要
血流感染症では、好中球が血管内免疫反応の一環として肝臓に集まり、血液中の病原体を駆逐するが、この重要な反応を制御するメカニズムは不明であった。我々は、無菌マウスと無脊椎動物マウスの好中球輸送のin vivoイメージングを用いて、腸内細菌叢が微生物の代謝物であるD-乳酸を介した感染に応答して好中球の肝臓へのホーミングを導くことを証明した。腸内細菌由来のD-乳酸は、骨髄の顆粒球形成や血液中の好中球の成熟・活性化とは無関係に、肝臓での好中球の接着を増大させる。その代わりに、腸から肝臓へのD-乳酸シグナルは、感染に応答して接着分子発現をアップレギュレートし、好中球の接着を促進する肝臓内皮細胞を刺激する。抗生物質誘発性ディスバイオシスモデルにおいて、微生物叢のD-乳酸産生を標的として補正すると、好中球の肝臓へのホーミングが回復し、黄色ブドウ球菌感染モデルにおいて菌血症を減少させる。これらの知見は、微生物と内皮のクロストークによる肝臓への好中球の長距離交通制御を明らかにするものである。
図解抄録
大きな画像を見る
高解像度画像をダウンロードする
キーワード
宿主防御
好中球の動員
D-乳酸
ジペプチダーゼ1
血管内免疫
敗血症
研究テーマ(複数可)
CP:免疫学
はじめに
全身感染症では、血流を介した病原体の拡散に対する宿主の防御には、高度に調整された血管内免疫応答が必要である。
1
好中球は微小循環をパトロールし、肝マクロファージ(クッパー細胞)と連携して、循環する細菌を捕捉・除去するファイアウォールを形成します。
1
感染症や敗血症の際に好中球が肝臓にホーミングするためには、通常の細胞リクルートメントのパラダイムとは異なる臓器特異的な接着機構が必要です。
2
感染時の好中球の肝臓への誘導は、ローリングステップを伴わず、非古典的な接着分子(好中球リガンドに結合する内皮ジペプチダーゼ-1 [DPEP-1] やCD44-HA相互作用)を介した肝類洞内皮細胞への繋留と直接的な固着によって進行します。
3
,
4
,
5
肝臓の微小循環に入ると、好中球は血管系をパトロールし、抗微生物分子、好中球細胞外トラップ(NETs)、食作用などの武器を通じて、循環から病原体の除去に貢献する。
6
,
7
,
8
好中球の肝臓へのホーミング、あるいは肝臓での抗微生物エフェクター機能が阻害されると、血液からの病原体除去が破綻し、播種性感染症になる。
7
,
8
,
9
肝血管系における好中球の動員を媒介する接着分子については詳細に理解されているが、この重要な血管内免疫反応と全身感染時の好中球の肝臓へのホーミングを組織化する機構についてはほとんど知られていない。
腸内細菌叢は、腸管内腔に区分されているにもかかわらず、血液中や遠くの臓器の免疫細胞との長距離コミュニケーションに参加し、血流感染に対する血管内免疫応答を含む全身性免疫の発達と機能を指揮しています。
10
好中球を介した宿主防御は腸内細菌叢に大きく影響され、無菌状態(GF)や抗生物質による腸内細菌叢の異常は、感染症に対する好中球の反応を損ない、最終的には制御不能な病原体の拡散をもたらす。
10
骨髄における好中球の生産には、増殖と分化を促進する微生物群由来の産物による骨髄前駆細胞の刺激が必要です。
11
,
12
,
13
好中球の成熟と老化は、腸由来のToll様受容体(TLR)リガンドが循環好中球のMyD88依存性シグナルに関与することにより、腸内細菌叢からの長距離シグナルを通じて微調整されます。
14
血液から組織への好中球の移動も腸内常在菌によって指示される可能性があり、これは感染巣への好中球の移動がGF宿主で変化するという観察から示唆されている。
15
,
16
,
17
しかし、腸内細菌が好中球ホーミングの長距離交通制御を媒介するメカニズムは、好中球移動機能の障害を反映しているのか、血管ホーミングシグナルの障害を反映しているのかなどを含めて、依然として不明である。
肝免疫の制御において腸内細菌の重要な役割が確立され、腸と肝臓の長距離交通のメカニズムが明らかになりつつある。血液媒介感染症の効果的な制御に不可欠な肝ゾネーションの発達とクッパー細胞の門脈周囲への配置は、腸内微生物のTLRリガンドからの入力を必要とする。
18
さらに、ユニークな腸-肝臓軸は、微生物叢由来の代謝物であるD-乳酸によって媒介される。D-乳酸は門脈を通じて高濃度で肝臓に到達し、クッパー細胞を刺激して循環病原体を捕捉・除去する。
19
さらに、肝類洞内皮細胞(LSEC)の基本的なトランスクリプトームが、腸から届く微生物分子によって調律されることが、最近の証拠によって証明されている。
20
したがって、肝臓微小循環は、腸内常在微生物からの長距離入力によって独自の影響を受けるが、好中球輸送の制御を通じた全身感染に対する宿主防御における腸-肝臓クロストークの役割については、これまで検討されてこなかった。
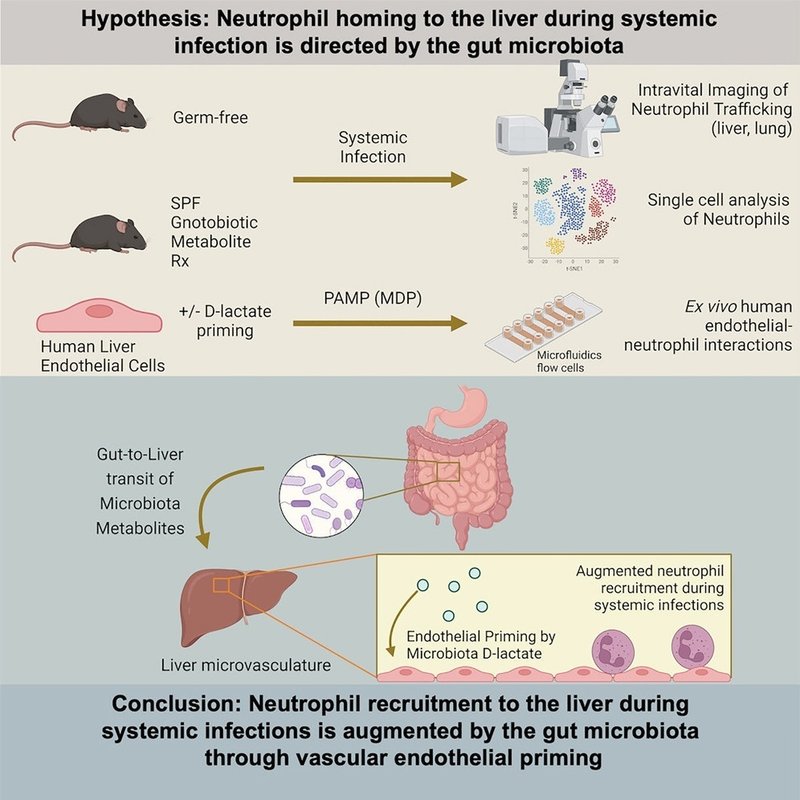
本研究では、全身感染時に腸内細菌叢が好中球の肝臓へのホーミングを指令するという仮説を検証し、肝臓微小循環への好中球ホーミングの長距離交通制御を媒介する常在細菌からのコミュニケーションの分子メカニズムを明らかにすることを目的としました。GFマウスおよびgnotobioticマウスにおける好中球輸送のin vivo(イントラビタル)イメージングを用いて、黄色ブドウ球菌による血流感染に応答して好中球が肝臓にホーミングするためには、腸内細菌叢が必要であることを明らかにした。腸内細菌叢由来の代謝物であるD-乳酸は、肝類洞内の好中球の接着を増強し、その効果は肝臓に限定され、肺への好中球のホーミングには影響を与えなかった。驚くべきことに、微生物のD-乳酸塩は、循環する好中球の産生、表現型景観、接着分子の発現に影響を与えなかった。その代わりに、微生物由来のD-乳酸による好中球輸送の制御は、感染に応答して接着分子発現をアップレギュレートし好中球を接着させるLSECのプライミングによって媒介されていた。抗生物質誘発性ディスバイオシスモデルにおいて、微生物叢のD-乳酸産生を標的的に補正すると、好中球の肝臓へのホーミングが回復し、全身性の黄色ブドウ球菌(S. aureus)感染後の菌血症を減少させた。この結果は、微生物と内皮のクロストークによって、好中球の肝臓微小循環への動員を長距離にわたって制御していることを示すものである。
研究成果
腸内細菌叢は全身性細菌感染時の好中球の肝臓への輸送を制御する
全身感染時に腸内常在菌が肝臓微小血管系への好中球輸送に影響を与えるかどうかを調べるため、特定病原体非感染マウス(SPF)とGFマウスにS. aureus USA300を静脈内投与し、既報のように共鳴走査共焦点顕微鏡(RS-IVM)を用いて肝臓での好中球輸送挙動を可視化しました。
8
,
19
SPFマウスでは、感染後12時間で大量の好中球が肝臓に集まり、洞内皮への好中球の動的な繋留と固着が見られた(図1A-1C;ビデオS1)。接着後、SPFマウスの好中球は平らになって伸び、血管内皮の内腔面に沿って這い始める(図1Dおよび1E;Video S1)。一方、GFマウスの肝臓における好中球の輸送は著しく調節され、繋留相互作用が著しく減少し、肝臓微小循環における付着好中球の量も減少した(図1A-1C)。好中球の動員における量的な違いに加えて、GFマウスの付着好中球の移動挙動もまた、調節されていた。肝臓に付着した好中球の形態と這い上がり速度のシングルセル解析から、GFマウスでは扁平化/伸展が損なわれ、這い上がり速度が低下していた(図1Dおよび図1E)。これらの結果から、腸内常在菌は、全身性の細菌感染に応答して、肝臓微小循環内における好中球の動員および血管内輸送挙動を増強することが示されました。
図1腸内細菌叢は、全身性細菌感染時に肝臓への好中球輸送を制御する
キャプションの全文を表示する
大きな画像を見る
高解像度画像をダウンロードする
.mp4のダウンロード (1.65 MB)
.mp4ファイルに関するヘルプ
ビデオS1. 肝臓における好中球の輸送は腸内細菌叢に支配されており、図1に関連する
SPFマウスと無菌マウスの肝臓微小循環のRS-IVMで、黄色ブドウ球菌感染から12時間後のBV421標識好中球(白色)の輸送挙動を示す。10分間のタイムラプス、スケールバー=70μm。
微生物叢由来のD-乳酸が肝臓への好中球の動員を制御する
腸内細菌叢は腸管に区分されているが、これまでの研究で、細菌叢由来の細菌産物や代謝物が血液やリンパ管を通じて腸管外の部位に移動し、肝臓やその他の臓器の免疫機能を制御することが示されている。
11
,
12
,
19
,
21
,
22
特に、微生物叢由来の代謝物であるD-乳酸は、門脈血液を介して肝臓に高濃度で輸送され(全身血流よりも約20倍高濃度)、肝臓に常駐するクッパー細胞の機能調節に関与している。
19
腸-肝臓軸におけるD-乳酸のユニークな役割を考慮し、この微生物代謝物が肝臓微小循環内の好中球流入に影響を与える可能性があると我々は仮定している。腸由来のD-乳酸の役割に直接言及するため(他の微生物や微生物の代謝物がない場合)、既述のように、GFマウスにD-乳酸を飲料水として3週間投与した、
19
,
23
そして、黄色ブドウ球菌感染に応答した肝臓での好中球の輸送を、RS-IVMを用いて可視化した。D-乳酸を投与したGFマウスは、ビヒクル飲料水を投与したGFコントロールと比較して、肝臓内皮への好中球の繋留および接着が有意に増加し、接着細胞の伸長も増加した(図2A-2C ; ビデオS2)。しかし、SPFマウスで増加した付着好中球のクロール速度(図1E)は、GFマウスのD-乳酸投与によって影響を受けなかったことから、肝臓の血管内輸送動態に寄与する微生物叢由来のシグナルがさらに存在すると考えられる(図2D)。肝臓への好中球輸送におけるD-乳酸の役割が黄色ブドウ球菌感染に特有であるかどうかを調べるために、グラム陽性(Enterococcus faecalis)とグラム陰性(Escherichia coli)の両方の院内感染の一般的な原因である追加の病原体の感染に反応して、D-乳酸およびビークル処理GFマウスの肝臓の好中球動態を画像化しました。黄色ブドウ球菌感染マウスと同様に、D-乳酸菌処理GFマウスでは、Enterococcus faecalis(E.フェカリス)およびEscherichia coli(大腸菌)の両感染後に、肝類洞における好中球付着量の増加が観察された(図S1A-S1H)。最後に、感染していない状態で肝類洞における基礎好中球輸送を調べたところ、SPFマウス、GFマウス、D-乳酸処理GFマウスの間で差が見られなかったことから、腸由来のD-乳酸存在下で肝臓への好中球輸送が増大するのは感染時にデノボで、肝臓の基礎好中球に差が生じた結果ではないことがわかりました(図S1I-S1L)。
図2微生物叢由来のD-乳酸は肝臓への好中球の動員を制御する
キャプションの全文を表示する
大きな画像を表示する
高解像度画像をダウンロードする
.mp4のダウンロード (0.44 MB)
.mp4ファイルに関するヘルプ
ビデオ S2. 腸由来のD-乳酸が肝臓への好中球輸送を制御する、図2に関連した図
GFマウスにD-乳酸またはビヒクル対照を飲水投与した後、S. aureus感染させ、肝臓微小循環のRS-IVMで好中球の輸送を可視化した(BV421-Ly6G.) スケールバー=50μm。
第二のアプローチとして、我々は、高レベルのD-乳酸を産生するラクトバチルス種を選択的にコロニー化したグノトビオティックマウスを用いて、善意の微生物コロニー化の設定におけるD-乳酸の役割を評価した(Lactobacillus intestinalis ASF360、 Lactobacillus intestinalis ASF360、LactateHI Gnoto)と、低レベルのD-乳酸を産生する乳酸菌(Lactobacillus reuteri I49、LactateLO Gnoto)でコロニー形成されたグノトバイオティクスマウスと比較する。
19
GFコントロールと比較して、LactateHI Gnotoマウスは、肝類洞内での好中球の繋留、接着、伸長、這い回り速度の増大を示したが、LactateLO Gnotoマウスの好中球運動はGFコントロールと変わらなかった(図2E-2H)。これらの結果から、腸内細菌が産生するD-乳酸は、全身性細菌感染時に肝臓微小循環への好中球の動員を促進することが示されました。
腸内細菌由来のD-乳酸による好中球輸送の臓器選択的制御
肝臓に加え、肺も全身性細菌感染や敗血症の際に大量の好中球が浸潤する。
24
我々は、腸由来のD-乳酸が、肝臓以外の肺微小循環内における好中球の輸送に同様の役割を果たすかどうかを検討した。RS-IVMを用いて、黄色ブドウ球菌の静脈内感染から12時間後の肺血管系における好中球輸送を可視化したところ、GFマウスでD-乳酸を補充しても、肺への好中球の動員には影響がないことがわかりました(図3A-3C )。これと一致して、肺組織におけるグローバルな好中球量の代用指標である肺組織のミエロペルオキシダーゼ(MPO)活性の測定は、感染後(図3D)または基礎(非感染)条件下(図S1M)、GFマウスの腸管D-乳酸投与によって影響を受けなかった。これらのデータは、微生物叢由来のD-乳酸が、肝臓への好中球の輸送を選択的に制御するが、肺への輸送は制御しないことを示している。
図3腸内由来D-乳酸による好中球輸送の臓器選択的制御
キャプションの全文を表示する
大きな画像を見る
高解像度画像のダウンロード
腸管由来D-乳酸による好中球輸送の制御は、顆粒球形成や循環好中球の機能調節とは無関係である。
腸内細菌由来のD-乳酸が好中球の肝臓への輸送を制御するメカニズムを明らかにするために、まずD-乳酸が循環好中球の量や表現型に影響を与えるかどうかを検討した。これまでの研究で、GFマウスの骨髄(BM)と末梢血において、恒常性維持下では好中球の量が減少し、微生物叢を持つマウスでは、BMの顆粒球形成を刺激する循環細菌産物によってこの量が回復することが報告されています。
11
,
25
これと一致して、GFマウスではSPFマウスと比較してBMおよび循環好中球の数が減少していることが観察された(図4A、4B、およびS2)。しかし、D-乳酸の経腸投与やLactateHI Gnotoマウスの腸管コロニー形成は、GF対照と比較してBMや循環中の好中球数に影響を与えなかったことから、D-乳酸やLactateHI Gnotoの他の代謝物や生成物は、顆粒球生成やBMから循環への排出を調節しないことがわかった(図4A~4D)。これらの所見は、常在性D-乳酸に応答した肝臓への好中球輸送の違いは、好中球生産または循環好中球の総数の違いに帰することはできないことを示している。
図4腸管由来のD-乳酸による好中球輸送の制御は、顆粒球の造血や循環好中球の機能調節に依存しない
キャプションの全文を表示する
大きな画像を見る
高解像度画像をダウンロードする
腸内D-乳酸に応答するBMや血液中の好中球の量には影響がないことが確認されたため、次に、腸内D-乳酸に応答する循環好中球の成熟、活性化、接着分子の発現に差が生じた結果、輸送量が増加するのではないかという仮説を立てました。この仮説を検証するため、D-乳酸投与およびビヒクル投与したGFマウスの血中好中球および肝臓から回収した好中球について、マスサイトメトリーを用いて高次元の単細胞解析を行った。まず、17の主要な好中球の表現型および機能マーカーに基づくFLOWSOMクラスタリング解析を行い、腸由来のD-乳酸が好中球の表現型多様性を調節するかどうかを調べた(図S3AおよびS3B)。黄色ブドウ球菌感染後の血液と肝臓の好中球を解析したところ、GFマウスでは9つの離散的な好中球のクラスターが確認され、主要マーカーの発現差に基づいてN1〜N9(任意のラベル)と名付けられた(図4E〜4H、S4A、およびS4B)。血液中の好中球は、主に成熟した好中球を示す3つのクラスターによって支配されていたが、肝臓で採取された好中球はより大きな表現型の異質性を示した(図4E-4H)。感染したD-乳酸処理GFマウスの血液と肝臓の両方において、個々の好中球クラスターの分布と量はGFコントロールと比較して変化せず、肝臓だけでなく血液中の好中球表現型の全体のランドスケープは、腸由来のD-乳酸によって影響を受けないことが示された(図4E-4H)。同様に、ナイーブな(感染していない)D-乳酸とビヒクル処理したGFマウスでは、好中球クラスターの分布に違いはなかった(図S4AおよびS4B)。プロトタイプ好中球集団の従来のゲーティングを用いてこれを確認するため、感染マウスまたはナイーブマウスの血液中の未成熟(CD11bintLy6Gint)、成熟(CD11bhiLy6Ghi)および老化(CD62LloCXCR4hi)集団の量に差がないことも判明し(図4I〜4K)、肝臓で観察された唯一の差は、D-乳酸投与GFマウスの高齢好中球がわずかに減少したことでした(図4I〜4K)。したがって、教師なし高次元解析とプロトタイプゲーティングの両方が、循環好中球の表現型に対するD-乳酸の影響を特定できず、肝臓では微妙な違い(老化好中球の減少)しか見つからなかった。最後に、D-乳酸が好中球の接着分子発現を調節し、肝臓への輸送に影響を与えるかどうかという、より焦点の定まった疑問について検討した。血液中および肝臓で採取した好中球のCD11a、CD11b、CD44、CD49dおよびCD62Lの発現に、D-乳酸を投与したGFマウスとビヒクル投与したGFマウスとの間に差は認められなかった(図S4C)。最後に、腸管由来のD-乳酸が生体内で肝採取好中球の抗微生物エフェクター機能に与える影響を評価した。RS-IVMを用いて、病原体(蛍光標識した黄色ブドウ球菌を静脈内投与)の好中球貪食とNET産生(Sytox Green+細胞外DNA)の可視化および定量化を行った(既報)。
8
グループ間の好中球の動員総量の違い(付着好中球100個あたりで正規化)を考慮すると、D-乳酸投与は好中球を介した黄色ブドウ球菌の貪食および肝類洞内の細胞外NETに影響を与えなかった(図S5A-S5C)。これらの結果から、微生物由来のD-乳酸による好中球輸送の制御は、顆粒造血や循環好中球の成熟、表現型、接着分子発現の機能的制御によって媒介されるものではないことが示された。
腸由来のD-乳酸は、血管内皮とのクロストークを通じて肝臓の好中球輸送を制御する
腸管由来のD-乳酸は、循環中の好中球数や活性化、肺の肝外血管系への好中球の動員には影響しないことから、好中球の動員に対するD-乳酸の臓器選択的作用は、肝臓の微細血管内皮を介するのではないかと考えられた。これを裏付けるように、細胞外乳酸シグナルに関与する受容体Gpr31の発現は、肝臓のマクロファージ(クッパー細胞)や肺の内皮細胞と比較して、LSECで顕著かつ差のある発現を示し、循環好中球では検出されないことがわかった(図5A)。Gpr31に加えて、もう一つの主要な細胞外乳酸受容体Gpr81もLSECで検出されたが好中球では検出されなかった(図5B)。次に、腸由来のD-乳酸が、感染時の肝臓における好中球のリクルートに関与する内皮接着分子の発現を変調させるかどうかを評価した。これにはICAM-1が含まれる、
26
,
27
TNFAIP6(CD44を介した好中球の接着経路に関与している)、
5
,
28
および最近発見されたDPEP-1は、敗血症時の肝臓における好中球の接着に重要である。
4
黄色ブドウ球菌を6時間感染させたD-乳酸菌投与GFマウスとビヒクル投与GFマウスからLSECを分離したところ、Tnfaip6やIcam1のLSEC発現はビヒクル投与GFマウスとD-乳酸菌投与GFマウスで差が見られなかったが、Dpep1の発現はD-乳酸菌処理GFマウスの肝臓内皮細胞で著しく上昇した(図5C〜図5E)。これらの結果から、生体内で肝臓内皮細胞がD-乳酸を感知することで、全身性の細菌感染に応答して、好中球のリクルートに必要なこの重要な接着分子であるDPEP-1の発現が上昇することが示唆されました。D-乳酸に応答した好中球の肝臓への動員におけるDPEP-1の役割を確認するために、D-乳酸感染マウスおよびビヒクル処理GFマウスにおいて、既述のように、スクランブルペプチドコントロールと比較して、定評あるペプチド阻害剤(LSALTペプチド)を用いてDPEP-1を阻害した。
4
実際、DPEP-1遮断は、D-乳酸塩を投与したGFマウスでは好中球のテザリングに影響を与えずに好中球の固着が減少したことから、腸由来のD-乳酸塩はDPEP-1依存的に肝臓での好中球固着を促進すると考えられた(図5Fおよび5G)。
図5腸由来のD-乳酸は、血管内皮とのクロストークを通じて肝臓での好中球の輸送を制御する
キャプションの全文を表示する
大きな画像を見る
高解像度画像をダウンロードする
微生物叢を持つマウスでは、肝内皮細胞は常に門脈血中の腸から届くD-乳酸にさらされているが、好中球が肝臓に集まるのは全身性の感染にさらされたときだけである。このことは、D-乳酸そのものが内皮を直接活性化して好中球の接着を誘導するのではなく、肝内皮が感染・炎症に反応するための素地を作っていることを示唆しています。このことを確認し、我々の知見をヒト系に拡張するために、D-乳酸によるhLSECプライミングの有無にかかわらず、マイクロ流体チャンバー内のフロー条件下でヒトLSEC(hLSEC)への好中球接着を検討した(図5H)。hLSECのD-乳酸プライミングは、SPFマウスの門脈血で観察される濃度(約10 mM)で実施した。
19
D-乳酸によるプライミングだけでは、フロー下で好中球の接着を誘導することができなかった(図5I)。しかし、ムラミルジペプチド(MDP;グラム陽性菌が産生するNOD2アゴニスト)で内皮を刺激すると、D-乳酸でプライミングしたhLSECはプライミングしなかったhLSECと比較して好中球の接着が劇的に増加した(図5I)。さらに、マウスで観察されたように、DPEP-1をLSALTペプチドでブロックすることにより、D-乳酸でプライミングしたLSECへの好中球の接着の増大が抑制された(図5I)。以上より、我々のin vivoマウスおよびin vitroヒトデータは、D-乳酸が肝類洞内皮を刺激し、感染症に反応して接着分子発現を上昇させ、好中球を接着させることを実証した。
抗生物質誘発性ディスバイオシス中の微生物叢のD-乳酸産生を治療的に補正すると、血流感染に対する好中球の血管内宿主防御が回復する。
私たちや他の研究者がマウスモデルで示したように、抗生物質(ABx)誘発性の腸内細菌叢のディスバイオシスは、好中球を介した血管内免疫の障害など、細菌感染に対する宿主防御の障害につながる。
12
,
19
,
29
,
30
ヒトにおいても、重篤な疾患の文脈で観察されるような重度のディスバイオシスが、有害な転帰や全身性感染症のリスク増加にも関連するという新たな証拠が得られている。
31
,
32
好中球が肝臓に集まる仕組みにおいて腸内細菌のD-乳酸が果たす役割を明らかにした我々は、抗生物質で誘発されたディスバイオシス後に微生物群のD-乳酸生成を補正することで、好中球の集まりとブドウ球菌による血流感染に対する宿主防御を回復できるかどうかを調べることを目的としました。SPFマウスに抗生物質を飲料水として3週間投与し、既述のように腸内環境のディスバイオシスを誘導した。
19
,
33
,
34
我々は以前、この抗生物質レジメンが腸内環境の異常を引き起こし、その結果、黄色ブドウ球菌を含む様々な細菌種によって引き起こされる血流感染に対する宿主防御が損なわれることを示している。
19
抗生物質投与および5日間の抗生物質洗浄期間の後、マウスは、ラクトバチルス・インテスティナリス(L. intestinalis)ASF360(高乳酸生産者、LactateHIと表記)またはラクトバチルス・ロイテリ(L. reuteri)I49(低乳酸生産者、LactateLOと表記)のいずれかの経口ガベージを(0日および3日に)2回受け取った(図6A)。両者とも抗生物質カクテルに感受性があったため、ラクトバチルス種の生着確立には抗生物質の洗浄期間が必要だった(図S6AおよびS6B)。両方の乳酸菌種による腸のコロニー形成は、摂取後7日までのすべてのマウスの糞便の培養によって確認されたが、PBS摂取のコントロールでは見られなかった(図6Bおよび図6C)。ABx処理したコントロールの小腸内容物中のD-乳酸レベルの定量化により、抗生物質処理により腸内のD-乳酸生成レベルが非常に低い(10匹中7匹で検出不能)ことが確認された(図6D)。LactateLO L. reuteri I49でコロニーを形成しても腸内D-乳酸レベルは上昇しなかったが、LactateHI L. intestinalis ASF360で処理すると腸内D-乳酸レベルが約17倍上昇し、抗生物質によるディスバイオシス環境において微生物群のD-乳酸生成を修正できることが確認された(図6D)。
図6抗生物質誘発性ディスバイオシスにおける微生物叢のD-乳酸産生の治療的補正は、血流感染に対する好中球の血管内宿主防御を回復させる
キャプションの全文を表示する
大きな画像を見る
高解像度画像をダウンロードする
微生物叢のD-乳酸産生を治療的に補正することで、血流感染に対する血管内宿主防御が改善するかどうかを調べるために、マウスにS. aureus USA300を静脈内感染させた。感染後12時間、RS-IVMを用いて肝臓微小循環における好中球の動員および輸送挙動を解析すると、LactateHI L. intestinalis ASF360でコロニされたマウスは肝臓洞底における好中球の繋留と接着が著しく増加している(図6Eと6F)。乳酸菌のコロニー形成は好中球の這い上がりに影響を与えず、付着した好中球の平坦化/伸長にも影響を与えなかった(図6Gおよび図6H)。最後に、微生物叢のD-乳酸産生の補正が血流における細菌播種に対する防御を改善するかどうかを決定するために、マウスに黄色ブドウ球菌を感染させ、感染後24時間後に採血して菌血症のレベルを定量化した。AbxマウスにLactateLO L. reuteri I49ではなく、LactateHI L. intestinalis ASF360を投与すると、感染後24時間の菌血症の負担が著しく減少した(図6I)。これらの結果から、抗生物質誘発性ディスバイオシス時に微生物群のD-乳酸産生を補正することで、肝臓への好中球の動員を改善し、全身感染時に循環からの細菌クリアランスの改善を促進することが実証されました。
考察
好中球輸送のin vivoイメージングと腸から肝臓へのクロストークのgnotobiotic解剖を組み合わせることで、腸内細菌叢が代謝物を介した微生物内皮クロストークを通じて、全身感染時の肝臓における好中球輸送の血管交通制御に寄与することを明らかにした。好中球輸送の制御は、腸内細菌によって産生され、門脈血液を介して肝臓に送られる細菌代謝物D-乳酸の腸-肝臓間輸送に依存していた。
19
GFマウスでは、微生物と内皮のクロストークが欠如しているため、全身感染に反応して肝臓にホーミングする好中球の数が減少した。D-乳酸の経腸投与、D-乳酸を産生する常在菌による腸管内コロニー形成、およびD-乳酸によるhLSECのex vivoプライミングにより、マウスにおける腸から肝臓へのD-乳酸輸送を再確立すると、感染に応じた肝臓内皮細胞への好中球の接着は回復した。注目すべきは、D-乳酸による好中球輸送の制御は肝微小循環に限定され、肺への肝外輸送には影響を与えず、循環中の好中球生産または成熟とは無関係であったことである。その代わりに、D-乳酸は、感染症に応答して内皮接着分子(DPEP-1)の発現を促進することにより、好中球の肝臓への選択的ホーミングを促進しました。抗生物質誘発性ディスバイオシスモデルマウスにおいて、D-乳酸を介した微生物と内皮のクロストークを治療的に補正することにより、肝臓への好中球の移動を回復し、黄色ブドウ球菌の血流感染に対する血管内宿主防御を改善した。これらの結果から、腸内細菌は血管内皮との長距離コミュニケーションを行い、好中球の肝臓へのホーミングを誘導することで血流に対する宿主防御をオーガナイズしていることが明らかになりました。
これまでの研究で、全身性感染症や敗血症の際に好中球が肝臓微小循環にホーミングすることが、血流を介した病原体の拡散に対する血管内防御に重要であることが立証されている。動員された好中球は、肝臓に常駐するクッパー細胞とともに血管内ファイアウォールを形成し、血液を介して運ばれる病原体や血液に移行する腸内常在菌を迅速に捕捉・除去し、播種性感染症から保護する機能を持つ。
35
,
36
好中球を肝臓の広大な微小循環内に配置し、洞道をパトロールしてNETを産生することで、全身感染時に血流から病原体を濾過する作用が増強されることが示されてきた。
7
,
8
今回のデータから、D-乳酸は腸内細菌叢と肝臓内皮細胞との間の直接的なコミュニケーションを仲介し、好中球のホーミングと肝臓の免疫ファイアウォールの強化を組織化することが明らかになりました。最近、微生物叢と肝臓内皮のクロストークは、クッパー細胞の肝臓微小循環内での位置決めとゾーニングのメディエーターとしても同定され、これは血液を介して感染する病原体に対する効果的な防御にも重要である。
18
これらのデータから、微生物と内皮のクロストークは、血液区画の最適な宿主防御のために、細胞輸送と血管内食細胞の位置決めを制御することによって、肝臓の免疫ファイアウォールを制御する重要なメカニズムであることが明らかになりました。
腸内細菌が肝臓への好中球のホーミングを調整するという我々の発見は、効果的な宿主防御の基本である微生物と好中球の相互作用の武装を高めるものである。これまでの研究で、常在細菌がBMでの好中球の発達と末梢循環での成熟を長距離にわたって制御していることが明らかになっている。腸内細菌叢の細菌産物は、腸内自然リンパ球とインターロイキン17A(IL-17A)および顆粒球コロニー刺激因子(G-CSF)の産生を含むTLR4/MyD88フィードバックループを介してBMの骨髄造血を促進します。
11
,
12
,
13
,
14
このように、腸内細菌は、病原体の侵入に対応できる末梢血中の好中球(および単球)の供給量を調節しています。さらに、末梢血区画内では、微生物叢由来の細菌産物が、循環好中球の老化やエフェクター機能の駆動に大きな役割を果たすなど、好中球の表現型景観を形成していることも分かっています。
14
,
37
,
38
私たちの発見は、血管内の宿主防御に重要な部位への好中球の動員を標的化、あるいは集中させる微生物叢の代謝物(D-乳酸)の役割を実証し、長距離の微生物叢-免疫制御の概念を構築した。しかし、このことは、微生物が顆粒造血や好中球の成熟に果たす役割から切り離されていることがわかりました。微生物由来のD-乳酸は、顆粒造血、循環好中球数、血液中の好中球の成熟や表現型に影響を与えないからです。微生物と好中球のクロストークを解除するメカニズムは、好中球の肝臓へのホーミングが、慢性炎症性肝障害を避けるために全身感染状況に限定され、血中の好中球の生産と発達に必要な構成的要件を確実に満たすために重要であると考えられる。興味深いことに、宿主由来(L-乳酸)と腸由来のBMにおけるGPR81を介した乳酸シグナルは、それぞれ好中球の動員および骨髄の細胞性/赤血球生成に影響を与えることが示されており、BMにおけるこの代謝物の多面的かつ文脈依存的な役割を示しています。
39
,
40
腸と肝臓をつなぐ門脈血液は、マイクロバイオームと腸管外(全身)免疫反応の長距離相互作用の文脈で、腸-肝臓軸をユニークなものにしています。門脈血は微生物由来の産物を豊富に含み、他の臓器と比較して不釣り合いなほど高濃度の微生物シグナルを肝臓に浴びせる。
35
,
41
D-乳酸の場合、門脈の濃度(小腸内容物の濃度に近い)は全身循環の10倍以上であることを示しました。
19
D-乳酸輸送の解剖学的コンパートメント化に加え、我々は、細胞外乳酸シグナル伝達に関与する受容体の遺伝子(Gpr31およびGpr81)が、LSEC(およびより少ない程度ではクッパー細胞)で劇的に不均衡に発現することを観察したが、これらの遺伝子は、以前の研究と一致して好中球で検出されなかった。
23
,
39
これらの知見を総合すると、好中球のホメオスタシス(末梢循環における量、成熟、活性化、または肺の肝外微小血管系への輸送)ではなく、肝臓における好中球輸送に対する微生物群D-乳酸の臓器選択的影響と一致する。
具体的には、腸由来のD-乳酸が肝臓内皮を刺激し、感染症に反応して血液中の好中球の接着と動員を開始するが、興味深いことに、その後の洞房内でのクロール/化学走性行動には影響しないことを見いだした。このことから、D-乳酸による内皮のプライミングは、肝臓での強固な接着に重要な内皮接着分子であるDpep-1の感染に応じたアップレギュレーションには必要だったが、Mac-1(CD11b/CD18)を介して肝臓での好中球のクロールと血管内パトロールに必要なIcam1の発現には変調を来さなかったことが判明した。
4
,
27
注目すべきは、SPF微生物群による腸内コロニー形成は、GF条件と比較して、接着だけでなく、その後のクロール/パトロール行動も増加させたことで、肝臓微小循環における好中球の移動行動には、微生物由来のシグナルがさらに寄与することが示唆された。実際、従来の微生物群による腸内コロニー形成は、GFマウスと比較して、LSECの基底トランスクリプトームにおいて、複数の化学誘引経路のアップレギュレーションや洞道における好中球のクロールに寄与すると考えられるメディエーターなど、複数の経路による変化を促すことが示されている。
20
集中治療室の重症患者に見られるような腸内細菌叢の重度の異常は、保護的な微生物と免疫の相互作用の崩壊につながり、院内感染のリスク上昇と関連している。
31
,
32
,
42
同様に、SPFマウスにおける抗生物質誘発性腸内異常症のマウスモデルでも、好中球応答の障害など、全身的な宿主防御機構の破綻が報告されており、糞便微生物叢移植(FMT)による微生物叢再構成によって救済されることがある。
12
,
43
しかし、脆弱な重症患者におけるFMTの安全性に関する新たな懸念から、腸内細菌叢が破壊された患者において、微生物と免疫の恒常性を再確立するための、より的を絞ったアプローチを開発する必要性が生じている。
44
そこで、我々は、抗生物質で治療したSPFマウスにおいて、乳酸産生常在菌による治療的腸内コロニー形成により、腸内のD-乳酸産生を標的として補正することで、黄色ブドウ球菌の菌血症時に肝臓への好中球の誘導が回復し、血流からの病原体の除去が改善されることを示す。全身的な宿主防御における微生物と免疫のクロストークを媒介するメカニズムの理解が進むにつれ、微生物叢の編集による免疫調節(微生物免疫療法)は、感染症を予防・治療する精密誘導型介入の有望な機会であることが示された。
本研究の制限事項
本研究では、敗血症時の好中球ホーミングと血管内宿主防御に重要な臓器として、主に肝臓と肺に焦点を当てました。本研究の結果は、肺ではなく肝臓への好中球輸送のプライミングにおける微生物由来のD-乳酸の役割を支持しているが、体内のすべての臓器を評価したわけではないので、他の臓器における好中球ホーミングの制御や内皮の調節におけるD-乳酸の役割の可能性を排除できない。さらに、本研究では、急性全身感染症に反応して肝臓に流入する好中球を調べたが、好中球(および他の免疫細胞)は、局所的(肝腫瘍微小環境や局所無菌性損傷など)だけでなく、虚血再灌流損傷や非感染性全身性炎症などの全身的な他の炎症刺激に反応して肝臓にも浸潤する。他の炎症病態に反応する肝臓内皮のプライミングと好中球輸送における腸内細菌叢とD-乳酸の役割は、本研究では扱われておらず、好中球輸送の分子機構が炎症刺激によって異なる可能性があることを考えると、異なる貢献があるのかもしれない。
26
,
27
最後に、好中球は、クッパー細胞などの肝微小循環にある他の免疫細胞と協力して血流から病原体を除去しますが、このクッパー細胞もD-乳酸を含む腸内細菌叢のシグナルによって機能的に調節されることが分かっています。
18
,
19
したがって、腸内細菌叢のD-乳酸産生を背景に血流病原体のクリアランスが改善されるという我々の発見は、肝臓における好中球および非好中球を介した反応の集大成であると考えられる。
STAR★メソッド
キーリソース表
REAGENTまたはRESOURCEIDENTIFIER抗体抗マウスCCR2 (clone 473201)R&DCat # MAB55381; RRID:AB_2749828 抗マウスCD11a (clone M17/4)BioXCellCat # BE0006; RRID: AB_1107578FITC-conjugated anti-mouse CD11b (clone M1/70)BD BiosciencesCat # 557396;RRID:AB_396679PerCP-Cy5. 5コンジュゲート抗マウスCD11b(クローンM1/70)BioLegendCat # 101227; RRID:AB_893233143Nd-conjugated anti-mouse CD11b (clone M1/70)FluidigmCat # 3143015B; RRID: AB_2811240Alexa Fluor 647-conjugated anti-human CD15 (clone W6D3)BioLegendCat # 323012; RRID:AB_756018209Bi-conjugated anti-mouse CD11c (clone N418)FluidigmCat# 3209005B; RRID:AB_2811244PerCP-Cy5. 5コンジュゲート抗マウスCD3e (clone145-2C11)BioLegendCat # 100327; RRID:AB_893320152Sm-conjugated anti-mouse CD3e (clone 145-2C11)FluidigmCat # 3152004B; RRID:AB_2687836PerCP-Cy5. 5コンジュゲート抗マウスCD19(クローン6D5)BioLegendCat # 115533; RRID:AB_2259869171Yb-conjugated anti-human/mouse CD44 (clone IM7)FluidigmCat # 3171003B; RRID: AB_2895121BV510-conjugated anti-mouse CD45 (clone 30-F11)BD BiosciencesCat # 563891; RRID:AB_273413489Y-conjugated anti-mouse CD45 (clone 30-F11)FluidigmCat # 3089005B; RRID:AB_2651152PerCP-Cy5. 5コンジュゲート抗マウスCD45R/B220 (clone RA3-6B2)BioLegendCat # 103235; RRID:AB_893356144Nd-conjugated anti-mouse CD45R/B220 (clone RA3-6B2)FluidigmCat # 3144011B; RRID: AB_2811239151Eu-conjugated anti-mouse CD49d/VLA-4 (clone R1-2)FluidigmCat # 3151016B160Gd-conjugated anti-mouse CD62L (clone MEL-14)FluidigmCat # 3160008B;RRID: AB_2687840159Tb-conjugated anti-mouse CD184/CXCR4 (clone L276F12)FluidigmCat # 3159030B164Dy-conjugated anti-mouse CXC3R1 (clone SA011F11)FluidigmCat # 3164023B;RRID: AB_2832247PE-conjugated anti-mouse F4/80 (clone BM8)BioLegendCat # 123109; RRID:AB_893498Anti-mouse IA/IE (clone M5/114. 15.2)BD BiosciencesCat # 556999; RRID:AB_396545165Ho-conjugated anti-mouse IFNg (clone XMG1. 2)FluidigmCat # 3165003B; RRID:AB_2925174158Gd-conjugated anti-mouse IL-10 (clone JES5-16E3)FluidigmCat # 3158002B; RRID:AB_2922918174Yb-conjugated anti-mouse IL-17A (clone TC11-18H10. 1)FluidigmCat # 3174002B; RRID:AB_2925175150Nd-conjugated anti-mouse Ly-6C (clone HK1.4)FluidigmCat # 3150010B; RRID:AB_2895118AF700-conjugated anti-mouse Ly6C (clone HK1.4). 4)BioLegendCat # 128023; RRID:AB_10640119141Pr-conjugated anti-mouse Ly-6G (clone 1A8)FluidigmCat # 3141008B; RRID:AB_2814678BV421-conjugated anti-mouse Ly6G (clone 1A8)BioLegendCat # 127627; RRID: AB_10897944162Dyコンジュゲート抗マウスTNFa(クローンMP6-XT22)FluidigmCat#3162002B;RRID:AB_2801437細菌およびウイルス株Enterococcus faecalis KB1Brugiroux et al.
45
N/AEscherichia coli Xen14Perkin ElmerCat # 119223Lactobacillus intestinalis ASF360Brugiroux et al.
45
N/ALactobacillus reuteri I49Brugiroux et al.
45
N/AStaphylococcus aureus USA300/LACSurwaard et al.
46
N/AChemicals、peptides、recombinant proteinsAmpicillinCayman ChemicalsCat # 14417Anti-CD31 MicroBeads, mouseMiltenyi BiotecCat # 130-097-418; RRID:AB_2814657Anti-F4/80 MicroBeads UltraPure, mouseMilteny BiotecCat # 130-110-443Anti-Ly6G MicroBeads UltraPure, mouseMiltenyi BiotecCat # 130-120-337; RRID AB_2858241Cytofix/Cytoperm™BDCat # 554722De Man, Rogosa and Sharpe (MRS) BrothSigma-AldrichCat # 69966EQ Four Element Calibration BeadsFluidigmCat # 201078LSALT peptide, C-term amidated (H-LSALTPSPSWLKYKAL-NH2)Choudhury et al.
4
N/ANeomycinFisher BioReagentsCat # BP2669RNAlaterInvitrogenCat # AM7021Sodium D-LactateSigma-AldrichCat # 71716Scrambled LSALT peptide, C-term amidated(H-LWLPLKSATPSYALSK-NH2)Choudhury et al.
4
N/AVancomycinCayman ChemicalsCat # 15327-5Critical commercial assaysCell-ID™ 20-Plex Pd Barcoding KitFluidigmCat # 201060iScript™ Reverse Transcription 5X SupermixBioRadCat # 1708841D- Lactate Colorimetric Assay KitBioVisionCat #K667Lactate Colorimetric Assay Kit IIBioVisionCat #K627Maxpar ® X8 Multimetal Labeling KitFluidigmCat # 201300PowerUp™ SYBR™ Green Master MixApplied BiosystemsCat # A25742QIAamp PowerFecal Pro DNA KitQiagenCat # 51804実験モデルについて: セルラインプライマリーヒト肝洞内皮細胞(hLSEC)Angio-ProteomieCat # cAP-0012実験モデル: 生物/系統C57BL/6マウス、雄および雌、SPFおよび無菌International Microbiome CenterN/AOオリゴヌクレオチドGapdh、3′-GGGTGTGAACCACGAGAAAT-5′、5′-CCTCCACAATGCCAAAGTT-3′Morita et al.
23
N/AIcam1、3′-TTCACACTGAATGCCAGCTC-5′、5′-GTCTGCTGAGACCCCTTG-3′Salim et al.
47
N/AGpr31、3′-AGTCTGACAAACAGCCCAGG-5′、5′-CACTAGCAGGAAGCACAGTC-3′Morita et al.
23
N/AGpr81 3′-GGTGGCACGATGTCATGTT-5′、5′-GACCGAGCAGAACAAGATGATT-3′Jeninga et al.
48
N/ADpep1、3′-TGCTCTTCCGTCTCCATCTT-5′、5′-TTCAGGAAAGGTGGTTC-3′本論文N/ATnfaip6、3′-ATACAAGCTCACCTACGCCGAA-5′、5′-ATCCATCCAGCAGCACAGAT-3′Diaz et al.
49
N/AL. intestinalis ASF360 3′- GGTGAGAGCTGGGAAC-5′, 5′- AAGCAATAGCCATGCAGC-3′Gomes-Neto et al.
50
N/AL. reuteri I49 3′-GATTGACGATGGATCACCAGT -5′,5′-CATCCCAGAGTGATAGCCAA-3′Kim et al.
51
N/ASoftware and algorithmsLeica Application Suite XLeica MicrosystemsVersion 3.5.6.21594PrismGraphPadVersion 9.0CytobankCytobankVersion 7.2
新しいタブで表を開く
リソースの有無
リードの連絡先
リソースや試薬に関する詳細な情報およびリクエストは、リードコンタクトであるBraidon McDonald (bamcdona@ucalgary.ca)までお願いします。
試薬の入手方法
この研究では、独自の試薬は生成していない。
実験モデルおよび被験者の詳細
マウス
8~12週齢の雄雌C57BL/6 WTマウスを飼育し、カナダ・カルガリー大学国際マイクロバイオームセンターのフレキシブルフィルムアイソレーターで無菌またはgnotobiotic条件下で飼育した。特定病原体不検出(SPF)動物は、カナダ・カルガリー大学動物資源センターの個別換気ケージ(IVC)内で繁殖・飼育された。無菌状態またはコロニー形成状態は、糞便グラム染色、Sytox green DNA染色、好気性および嫌気性培養、16S rRNA遺伝子アンプリコン配列決定の組み合わせによって確認された。SPFマウスには、GFマウスおよびgnotobioticマウスと同じオートクレーブ食を与えた。すべてのプロトコルは、カルガリー大学動物ケア委員会(AC19-0139)の承認を得ている。
メソッドの詳細
細菌培養条件と感染症
S. aureus USA300 LACをBrain Heart Infusion(BHI)液体培地で37℃、撹拌しながら一晩培養した。感染には、一晩培養したものを指数期(OD660nm=1)になるまで下培養し、遠心分離して洗浄し、滅菌1Xリン酸緩衝生理食塩水に1×108CFU/mLの濃度で再懸濁した。動物を、2×107CFUの単回静脈内(i.v.)注射で感染させた。E. faecalis KB1およびE. coli Xen14をLuria Broth(LB)培地で37℃、一晩撹拌しながら培養し、上記のようにサブカルチャーした。感染量が2×108CFU E faecalis KB1または2×107CFU E coli Xen14となるように、腹腔内(i.p.)注射を単回投与した。
ラクトバチルス属菌によるモノクローナル化
Lactobacillus intestinalis ASF360およびL. reuteri I49を、De Man, Rogosa and Sharper(MRS)ブロス中、37℃で48時間、嫌気的に増殖させた。無菌動物には、L. intestinalis ASF360またはL. reuteri I49の培養液300μLを単回経口投与した。モノコロン化は、グラム染色と便のMRS寒天培地で37℃、24時間の嫌気性培養により毎週確認した。実験は、経口投与から3週間後に実施した。
D-乳酸のインビボ処理
指示された場合、無菌マウスは、既述のように、飲料水に50mMのD-乳酸(Sodium D-lactate, Sigma-Aldrich)を3週間投与(毎週補充)した。
19
抗生物質処理と乳酸菌のコロニー形成
SPFマウスに抗生物質(アンピシリン1 mg/mL、バンコマイシン0.5 mg/mL、ネオマイシン0.5 mg/mL、毎週補充)を飲料水にてアドリブで投与した。
19
,
33
,
34
,
52
3週間後、抗生物質カクテルを滅菌水に置き換え、5日間のウォッシュアウト期間を設けた。その後、マウスはL. intestinalis ASF360またはL. reuteri I49の107 CFUを経口摂取し、これを3日目に1回繰り返した。実験は、1回目の投与から7日目に実施した。以下に詳述するように、種特異的なプライマーを用いたqPCRによってコロニー形成を確認し、定量化した。
便サンプルの定量的PCR
QIAmp PowerFecal Pro DNAキット(Qiagen)を用いて、製造者の説明書に従って便サンプルからDNAを抽出した。便中のL. intestinalis ASF360およびL. reuteri I49の菌量を推定するために、菌株特異的プライマーを使用し、主要リソース表に記載した。定量的PCRは、PowerUp SYBR Green Master Mix(Applied Biosystems)を用いて、QuantStudio 3 Real-Time PCR System(Applied Biosystems)で、メーカーの説明書に従って行った。コロニー形成モニタリングでは、SPFマウスの便から抽出したDNAを1反応あたり2ng使用し、qPCRサーモサイクラーを30サイクルに設定した。L. intestinalis ASF360およびL. reuteri I49の純粋培養物5 mLから抽出したDNAを用いて、標準曲線を作成した。同じ培養物からのアリコートをMRS寒天培地にプレーティングし、CFU/mLを定量した。結果はCFU/g of stoolとして表され、液体純粋培養物からのCFUあたりの抽出DNAのngに基づいて推定されたものである。
肝臓のイントラビタル顕微鏡検査
マウスを麻酔し(ケタミン200mg/kg、i.p.、Bayer Animal Health、およびキシラジン10mg/kg、i.p.、Bimeda-MT)、蛍光標識抗体および麻酔薬の送達のために頸静脈をカニュレーションした。肝臓顕微鏡検査のための外科的処置は、記載されたとおりに行った。
8
,
53
簡単に言うと、開腹手術を行い、皮膚と筋肉を除去して腹腔を露出させた。次に、鎌状靭帯を解放し、動物を温度制御されたステージ(37℃)に左側側臥位で置き、肝臓の左葉をカバースリップに外嵌した。肝微小血管のイントラビタルイメージングは、HC FLUOTAR 25x/0.95 NA VISIR水対物レンズ、4つの励起レーザーライン(405、488、552、638 nm)、5つのスペクトル検出器(3 hybrid HyDと2 conventional PMT)、8kHz tandem resonant scan head (Leica Microsystems) を備えたLeica TCS-SP8 倒立顕微鏡で行った。好中球は、Brilliant Violent 421コンジュゲートLy6G抗体(0.4 μg/animal i.v., clone 1A8, BioLegend)を用いて可視化した。一部の実験では、マウスにDPEP-1遮断合成ペプチド(LSALT、1mM溶液200μL、14.2mg/kg、i.v.)または対照ペプチド(スクランブルLSALT、200μLまたは1mM溶液、14.2mg/kg、i.v)を前記のようにS. aureus感染後5分してから投与した。
4
一部の実験では、イメージング時にマウスに5×107 CFU Syto60標識S. aureus USA300を注入し、蛍光標識細菌の貪食と細胞外DNA NETをそれぞれ可視化するために10μL Sytox Green(5μM溶液)を、前述のように注入しました。
8
肺のイントラビタル顕微鏡観察
マウスを麻酔し、上記のように蛍光標識抗体と麻酔薬を送達するために頸静脈をカニュレーションした。肺の IVM のための手術は、以前に説明したように行った。
54
気管切開を行い、PE90チューブを挿入し、外科用タイで固定し、機械的換気を行った。動物は、右側側臥位でカスタムステージに置かれた。皮膚、結膜組織、2本の前肋骨を慎重に切除し、肺の小さな開口部を露出させた。動きを最小限にするため、左葉は胸部吸引窓に取り付けた。肺微小血管のイントラビタルイメージングは、25x/0.95NA水対物レンズ、調整可能な白色光と紫外線(405nm)レーザー、5つのスペクトル検出器(ハイブリッドHyD3つとPMT2つ)を備えたLeica SP8顕微鏡で行った。好中球はBrilliant Violent 421コンジュゲート抗Ly6G抗体(0.4 μg/animal i.v., clone 1A8, BioLegend)を、内皮はAlexa Fluor 647コンジュゲート抗CD31抗体 (0.4 μg/animal, MEC13.3, BioLegend) で可視化された。
イントラビタルイメージング解析
すべてのビデオと画像は、Leica LASXソフトウェアを使用して取得した。画像解析は、LASXソフトウェア(Leica Microsystems)を用いて行った。好中球の動態は、マウス1匹につき10分間追跡した。付着性好中球は、肝臓の血管系で60秒以上停止したLy6G+細胞と定義し、繋留性好中球は、60秒未満で停止し、その後血液の流れに放出されたものと定義された。付着好中球と繋留好中球の数は、1動物あたり5視野(肝臓)または3視野(肺)にわたる1mm2あたりの好中球の平均数として表した。細胞這い上がり速度は、付着好中球について取得し、μm/sとして表した。好中球の極性を測定するために、個々の付着好中球について、最長軸(長さ)を最短軸(幅)で割ることによって伸長指数を求めた。
肺組織におけるミエロペルオキシダーゼ(MPO)活性の定量化
MPO活性は、以前に記述したように評価した。
55
簡単に言うと、肺サンプルは、50mgの組織/mLの濃度でK2HPO4緩衝液(50mM、pH6.0、HTAB 5mg/mL)中でホモジナイズした。ホモジネートを遠心分離した(5分、5000g、4℃)。上清を3枚ずつ96ウェルプレートに移し、サンプルに新しく作ったO-dianisidine溶液(50mMリン酸緩衝液、pH6.0、O-dianisidine 0.167mg/kg, 0.005% H2O2)を添加した。直後に、405nmで30秒間隔で3つの吸光度値を得て、組織1mgあたりのMPOの単位を得ることになる。
好中球のカウント
麻酔したマウスから心臓穿刺で得た血液サンプルにおいて、Türkの溶液(2%氷酢酸、0.01%クリスタルバイオレット)と光顕微鏡下で可視化した血球計数器を用いて総白血球数をカウントした。末梢血中の好中球を定量化するため、Wright-Giemsaで染色した血液塗抹標本で微分カウントを行った。結果は、全血1mLあたりの細胞数で表した。
血液および骨髄のフローサイトメトリー
大腿骨をFACSバッファー(1x PBS、2mM EDTA、10%不活性化ウマ血清)でフラッシュし、70μmのセルストレーナーで懸濁液をろ過することにより、骨髄の細胞懸濁液を得た。ACK溶解バッファー(155mM NH4Cl、10mM KHCO3、0.1mM EDTA)で赤血球を溶解した後、サンプルを遠心分離(400g、5分、4℃)してFACSバッファーに再懸濁しました。マウス1匹あたり100万個の細胞を蛍光標識抗体で染色し、BD FACSCanto II (BD Biosciences)で解析した。モノクローナル抗体の製造元、クローン、カタログ番号は、主要リソース表に詳述されている。血液中では、好中球は生きた(Fixable Viability Dye eFluor 780, eBioscienceでは陰性)、CD45+CD11b+Ly6G+細胞として同定された。骨髄では、好中球は生きたCD45+CD3ϵ-CD19-B220-CD11b+Ly6G+ 細胞として同定された。ゲーティングストラテジーを図 S2 に示す。
飛行時間型マスサイトメトリー
麻酔したマウスから心臓穿刺により全血を採取し、直ちにプロテオーム安定剤(PROT1、SmartTube)と1:1.4の割合で混合し、既述のように凍結保存を行った。
56
サンプルは、さらに処理するまで-80℃で保存した。その後、試料を室温で解凍し、解凍-溶解バッファー(SmarTube)を用いて赤血球を溶解した。
マウス肝臓の単細胞懸濁液は、以前に記載されたとおりに調製した。
57
簡単に説明すると、肝臓をはさみでミンチにし、消化バッファー(10%ヒスと100nM HEPESを含むRPMI、コラゲナーゼVII 1mg/mL、DNase 1 10U/mL含有)中で37℃、30分間、撹拌しながらインキュベートした。その後、細胞懸濁液を100μmおよび70μmのセルストレーナーで濾過し、次いで低速遠心分離(60g×5分)して肝細胞を沈殿させ、上澄み画分に非実質細胞を回収した。赤血球溶解後、白血球および肝細胞懸濁液を細胞染色培地(PBS中1%ウシ血清アルブミン)で洗浄し、赤血球破片および溶解バッファーを十分に除去した。次に、Cell-ID 20-Plex Pd Barcoding Kit (Fluidigm)を用いて、製造者の説明書に従ってサンプルのバーコード化を行った。その後の染色ステップのために、6つのサンプルを1つのチューブにプールした。次に、サンプルをFcブロックし、金属結合抗体で標識した(抗体パネルの詳細、金属結合体、供給者、カタログ番号については、主要リソース表を参照)。次に、細胞を100 nM cell-ID Iridium Intercalator(Fluidigm)、0.3%サポニン、1.6%パラホルムアルデヒドを含むPBS中で4℃、一晩インキュベートした。翌日、細胞を洗浄し、EQ Four Element Calibration Beads(Fluidigm)と混合して、Helios CyTOF IIマスサイトメーター(DVS Sciences)で取得しました。データは、内部のHelios CyTOF IIマスサイトメータービーズベースの正規化ソフトウェア(DVS Sciences)を使用して正規化した。
Helios CyTOFIIで生成された飛行時間型マスサイトメトリーFCSファイルをCytoBankにインポートし、生存CD45+シングルレットセルイベント、および好中球(CD3-CD19-CD11b+Ly6G+セル)の初期解析およびゲーティングを行い、CATALYSTパッケージを使用してRでクラスタリング解析するためにFCSファイルとしてエクスポートした。エクスポートされた好中球は、CATALYST v1.16.0により、FlowSOM関数を用いて、20メタクラスタのデフォルトパラメータを使用してRでクラスタリングした。非常に稀な集団(相対存在率が2%以下)を表すクラスターは除外された。主要な好中球マーカーの発現をマウス集団間で比較するため、アークサイン変換したシグナル強度の平均値(平均シグナル強度)を比較した。
マイクロフルイディック・フローチャンバーアッセイ
初代ヒト肝類洞内皮細胞(hLSEC, Angio-Proteomie, CAT# cAP-0012)は、内皮増殖培地(Lifeline Cell Tech)で培養し、継代3から5まで使用しました。マイクロ流体フローチャンバー実験は、既述のように実施した。
54
簡単に説明すると、hLSECをIbidi VI 0.1チャンバーに1x106/mLで播種し、実験中は連続振動流(約0.1Hz、5cm H2O 圧力勾配)下で培養した。合流後1-2日後、0日目に単層を10 mM D-乳酸またはビヒクル(滅菌水)で刺激した。1日目に、チップの各チャンバーを、ビヒクル、D-乳酸10mM、ムラミルジペプチド(MDP)20μg/mL(Invivogen)、またはD-乳酸とMDPの両者の組み合わせで処理した。2日目、すべての培地を直ちに吸引し、チャンバーをHBSSでフラッシュしてヒト全血灌流を準備した。同時に、健康なドナーから、低用量ヘパリン20U/mLの抗凝固を施した静脈穿刺を用いて、ヒト全血を調製した。100μLのヘパリン化血液をAlexa Fluor 647標識抗CD15(2.5μL、W6D3、BioLegend)で標識し、穏やかに攪拌しながらRTで15分間インキュベートした。DPEP1阻害実験のために、細胞を、全血灌流の前に、20μMのブロッキングペプチド(LSALT)または対照ペプチド(スクランブルLSALT)と30分間インキュベートした。標識した全血をさらにHBSSで1:10に希釈し、シリンジポンプ(Harvard Apparatus)を用いて、hLSECを含むIbidiチップのチャンネルを通して0.5dyn/cm2、12分間灌流した。チャンバーをLeica TCS-SP8共焦点顕微鏡(10×0.4NA対物レンズおよびLASXソフトウェア(Leica Microsystems)を用いて可視化し、CD15+好中球の接着をFOVごとに定量化した(既報の通りである)。
54
ヒト細胞および血液を含む実験は、Alberta Health Services および The University of Calgary の Conjoint Research Ethics Board による審査および承認を得た(REB20-0481)。
肝臓からのマウス内皮細胞およびマクロファージの単離
マウス肝臓の単細胞懸濁液は、既述のように調製した。
57
簡単に言うと、肝臓をはさみでミンチにし、消化バッファー(10%ヒスと100nM HEPESを含むRPMI、コラゲナーゼVII 1mg/mL、DNase 1 10U/mL含有)中で37℃、30分間撹拌しながらインキュベートした。その後、細胞懸濁液を100μmおよび70μmのセルストレーナーで濾過し、低速遠心分離(60g×5分)して肝細胞を沈殿させ、上澄み画分に非実質細胞を回収した。次に、非実質細胞画分を、10mM HEPESおよび2% hisを含む補充RPMIに再懸濁し、その後、CD31+細胞のポジティブセレクションのために、抗CD31 MicroBeads (Miltenyi Biotec) とともに、製造者の指示に従ってインキュベーションした。マクロファージの分離には、CD31-画分を抗F4/80 MicroBeadsとインキュベートし、F4/80+細胞のポジティブセレクションを行い、製造元の指示に従った。
肺からの内皮細胞単離
肺の細胞懸濁液は、以前に記載されたとおりに入手した
58
を得たが、以下の変更を加えた。肺血管を冷PBSで灌流し、ハサミでミンチにし、消化バッファー(コラゲナーゼIA 3mg/mL)中で37℃、45分インキュベートした。その後、細胞懸濁液を70μmセルストレーナーでろ過し、遠心分離(400g、5分、4℃)し、無血清DMEMに再懸濁した。内皮細胞は、CD31+細胞のポジティブセレクションのために、抗CD31 MicroBeads(Miltenyi Biotec)を用いて、製造者の説明書に従って単離した。
全血からの好中球の分離
マウスから心臓穿刺により全血を採取した。ACK溶解バッファー(155 mM NH4Cl, 10 mM KHCO3, 0.1 mM EDTA)で赤血球を溶解後、サンプルを遠心分離(400 g, 5 min, 4℃)し、RPMI(10 mM HEPES, 2% iHS)を加えて再懸濁し、製造元の指示に従って、抗Ly6G MicroBeads(ミルテニ・バイオテック)とインキュベーションしてLy6G+細胞をポジティブ選択しました。
RNAの単離
組織サンプルは、RNAlater安定化溶液(Invitrogen)中で4℃で一晩保存し、その後-80℃で長期保存した。RNA単離のため、組織をRNAlaterから取り出し、TRIzol Reagent(Invitrogen)でホモジナイズし、クロロホルムと混合し、12000g、15分、4℃にて遠心分離を行った。上相を得、分子グレードのエタノール70%と1:1の割合で混合し、Qiagen Rneasy Mini column(Qiagen社)に移した。次の工程は、製造元の説明書に従って行った。全RNAを30~60μLのRnaseフリー水で溶出した。肝臓および肺内皮細胞、肝臓マクロファージおよび血液好中球からのRNA単離は、Qiagen Rneasy Micro Kit(Qiagen)を用いて磁気選別後すぐに、製造者の指示に従って行い、全RNAを14μLのRnaseフリー水で溶出させた。
定量的(RT)-PCR
iScript Reverse Transcription 5X Supermix(BioRad)を用いて、製造者の指示に従ってRT-PCRを行い、qPCR用のcDNAを取得した。1回の反応につき、肝臓および肺の組織サンプルから抽出した2μgのRNAと、細胞分画から40〜200ngを使用した。QuantStudio 6 Real-Time PCR System(Applied Biosystems)上でPowerUp SYBR Green Master Mix(Applied Biosystems)を用いて定量的PCRを実施した。相対的な遺伝子発現は、比較2-(ΔΔCt)法を用いて測定した。Gapdh (glyceraldehyde 3-phosphate dehydrogenase) mRNAの発現は、データを正規化するための参照遺伝子として使用された。プライマー配列は、主要リソース表に記載されている。
D-乳酸の測定
D-乳酸レベルは、製造者の指示に従い、比色アッセイキットを用いて測定した(BioVision)。試験管内でのD-乳酸産生を測定するために、L. intestinalis ASF360およびL. reuteri I49をMRS中、37℃で48時間嫌気的に培養し、小腸内腔のD-乳酸を測定するために、小腸内容物ホモジネート(滅菌水中0.5mg/μL)をMRSブロスに接種して、純粋培養と同じ条件で培養した。遠心分離(10,000g、10min、RT)により培養上清を得、5μLの培養上清をアッセイに使用した。
定量化および統計解析
すべての実験について、生物学的複製の数(n)は、図の説明文に示されている。生物学的複製は個々のマウスで構成されている。正規分布データは、平均値±平均値の標準誤差(SEM)で示し、非対称T検定(2群比較)または一元配置ANOVAと多重比較のためのTukeyのポストホックテスト(2群以上比較)を用いて比較する。正規分布でないデータは、中央値と四分位範囲で示し、Mann-Whitney Uテスト(2群について)、またはKruskal-Wallisに続いてDunnの多重比較テスト(>2群について)を用いて比較した。2つの独立変数が存在する場合(例:感染と異なる臓器/区画、感染とDPEP1阻害剤治療)、Šídákの多重比較検定による二元配置ANOVAを使用しました。データ解析と可視化は、GraphPad Prism version 9.0 (GraphPad Software, Inc)を用いて行った。
データおよびコードの利用可能性
本論文では、オリジナルのコードは報告していません。この論文で報告されたデータは、要求に応じてリードコンタクトが共有する。本論文で報告されたデータを再解析するために必要な追加情報は、要求に応じてリードコンタクトから入手できます。
謝辞
Nicola Pett、Anowara Islam、Karen Poon博士、Michael Dicay、Christopher Richmond、およびInternational Microbiome Center(IMC)スタッフの本研究における技術サポート、およびBas Surewaard博士のS. aureus株の提供に感謝する。LSALTペプチドおよびスクランブルペプチドコントロール試薬を提供してくださったDaniel Muruve博士とAuthur Lau博士に感謝します。イラストはBioRender.com (http://biorender.com)を使用して作成しました。この研究は、カナダ保健研究所(CIHR)のプロジェクト助成金(助成番号173296)、CIHR Early Career Investigator Award in Infections and Immunity(助成番号170746)、カナダ革新財団JR Evans Leaders Fund Grant(助成番号40697)により、すべてB. M.A.Z.Z.は、CIHR Frederick Banting and Charles Best Canada Graduate Scholarship Doctoral Award(CGS-D)、Cumming School of Medicine Graduate Scholarship(カナダ、カルガリー大学)、Beverley Phillips Rising Star Graduate Scholarship(カナダ、カルガリー大学)から支援を受けています。
著者貢献
構想、A.Z.Z.およびB.M.、方法論、A.Z.Z.、K.D.M.、M.R.G、 B.G.Y. および B.M.; 調査、A.Z.Z., J.S., A.I., C.A.T., I.-l.Y., S.P., B.G.Y., M.R.G.. 、 およびB.M.、執筆-原案、A.Z.Z.およびB.M.、執筆-レビューおよび編集、A.Z.Z., J.S., A.I., M.B.G., K.D.M., B.Y., M.R.G., B.M. 、資金獲得、B.M、指導、 K.D.M. and B.M.
利害関係の宣言
著者は、競合する利害関係を宣言していない。
インクルージョンと多様性
私たちは、包括的で多様な、そして公平な研究の実施を支持します。
補足情報
.pdfをダウンロード (5.06 MB)
pdfファイルのヘルプ
資料S1. 図S1〜S6
参考文献
ヒッキー M.J.
クベスP.
血管内免疫:血管における宿主と病原体の出会い。
Nat. Rev. Immunol. 2009; 9: 364-375https://doi.org/10.1038/nri2532
記事で見る
Google Scholar
レイ・K.
ラウダンナC.
サイブルスキーM.I.
Nourshargh S.
炎症部位への到達:白血球接着カスケードが更新された。
Nat. Rev. Immunol. 2007; 7: 678-689https://doi.org/10.1038/nri2156
記事で見る
Google Scholar
ウォン J.
ジョンストン B.
リーS.S.
ブラードD.C.
スミスC.W.
ボーデA.L.
Kubes P.
炎症を起こした肝微小血管への白血球の動員におけるセレクチンの最小限の役割。
J. Clin. Invest. 1997; 99: 2782-2790https://doi.org/10.1172/JCI119468
記事で見る
Google Scholar
Choudhury S.R.
バベスL.
ラーンJ.J.
アーンB.Y.
ゴーリング K.A.R.
キングJ.C.
ラウ A.
ペトリ B.
ハオ X.
チョイナッキ A.K.
et al.
ジペプチダーゼ-1は、肺と肝臓における好中球のリクルートメントのための接着受容体である。
Cell. 2019; 178: 1205-1221.e17https://doi.org/10.1016/j.cell.2019.07.017
記事で見る
グーグル・スカラー
マクドナルド・B.
マカボイE.F.
ラム・F.
ギルV.
ド・ラ・モットC.
サヴァニ R.C.
Kubes P.
CD44とヒアルロン酸の相互作用は、炎症を起こした肝類洞における好中球の隔離の主要なメカニズムである。
J. Exp. Med. 2008; 205: 915-927https://doi.org/10.1084/jem.20071765
記事で見る
Google Scholar
マスバーグ S.
グラールL.
フォン・ブリューエルM.L.
マヌキアンD.
ファイラーS.
グースマンC.
ブリンクマン V.
ローレンツ M.
ビジェコフ K.
カンダガレA.B.
他
好中球のセリンプロテアーゼを介した凝固と自然免疫の相互結合
Nat. Med. 2010; 16: 887-896https://doi.org/10.1038/nm.2184
記事で見る
Google Scholar
クラーク S.R.
マーA.C.
タベナーS.A.
マクドナルドB.
グッダージZ.
ケリーM.M.
パテル K.D.
チャクラバルティ S.
マカボイ E.
シンクレアG.D.
他
血小板 TLR4 は好中球細胞外トラップを活性化し、敗血症の血液中の細菌を捕捉する。
Nat. Med. 2007; 13: 463-469https://doi.org/10.1038/nm1565
記事で見る
Google Scholar
マクドナルド B.
ウルティアR.
イップ B.G.
ジェンヌC.N.
クベスP.
血管内好中球細胞外トラップは、敗血症の際に血流から細菌を捕捉する。
Cell Host Microbe. 2012; 12: 324-333https://doi.org/10.1016/j.chom.2012.06.011
記事で見る
Google Scholar
コラツコウスカ E.
ジェンヌC.N.
スールワルドB.G.J.
タナバラスリアーA.
リーW.Y.
サンツM.J.
モウエン K.
オプデナッカーG.
Kubes P.
肝血管系におけるNETの形成と分解の分子機構をイントラビタルイメージングにより明らかにした。
Nat. Commun. 2015; 6: 6673https://doi.org/10.1038/ncomms7673
記事で見る
グーグル スカラー
シュレヒトJ.
スカロスキーI.
ゲウキングM.B.
マクドナルドB.
遠距離恋愛-腸内細菌叢による感染症に対する全身性宿主防御の制御。
Mucosal Immunol. 2022; 15: 809-818https://doi.org/10.1038/s41385-022-00539-2
記事で見る
グーグル・スカラー
バルマー M.L.
シュルヒC.M.
サイトウ ヨウ。
ゲウキングM.B.
リー・エイチ
クエンカ M.
コヴトニューク L.v.
マッコイ K.D.
ハプフェルマイヤー S.
オクセンバインA.F.
他
微生物由来の化合物は、MyD88/TICAMシグナルを介して定常的な顆粒球形成を促進する。
J. Immunol. 2014; 193: 5273-5283https://doi.org/10.4049/jimmunol.1400762
記事で見る
グーグル・スカラー
デシュムク・H.S.
リュウ・Y.
メンキチO.R.
Mei J.
ダイ N.
オリアリーC.E.
オリバー P.M.
コルスJ.K.
ワイザーJ.N.
Worthen G.S.
新生児マウスの好中球のホメオスタシスと大腸菌K1敗血症に対する宿主抵抗性を微生物叢が制御している。
Nat. Med. 2014; 20: 524-530https://doi.org/10.1038/nm.3542
記事で見る
グーグル・スカラー
ホスラビ A.
ヤーニェスA.
プライスJ.G.
チャウ A.
メラド M.
グッドリッジ H.S.
マズマニアン S.K.
腸内細菌は造血を促進し、細菌感染を制御する。
セルホストマイクロベ。2014; 15: 374-381https://doi.org/10.1016/j.chom.2014.02.006
記事で見る
グーグル スカラー
チャン・ディー.
チェンG.
マンワニD.
モルサ A.
シュー・C.
フェイス・J.J.
バーク・R.D.
クニサキ Y.
ジャン・J.E.
Scheiermann C.
ら。
好中球の老化はマイクロバイオームによって制御されている。
Nature. 2015; 525: 528-532https://doi.org/10.1038/nature15367
記事で見る
グーグル・スカラー
ファグンデス C.T.
アマラルF.A.
ヴィエイラA.T.
ソアレスA.C.
ピノ V.
ニコリ J.R.
ヴィエイラ・L.Q.
テイシェイラM.M.
ソウザ D.G.
一過性のTLR活性化により、生殖不能マウスの炎症反応と肺細菌感染を制御する能力が回復する。
J. Immunol. 2012; 188: 1411-1420https://doi.org/10.4049/jimmunol.1101682
記事で見る
Google Scholar
カンター M.
トムコビッチS.
シャオルン・S.
グロッサーM.R.
クー J.
フリンE.J.
ジョビン C.
ロールズ J.F.
腸内細菌は血清アミロイドAの誘導を介して全身性好中球の移動を刺激する。
Cell Microbiol. 2014; 16: 1053-1067https://doi.org/10.1111/cmi.12257
記事で見る
グーグル・スカラー
カルマルカー D.
ロック・K.L.
MyD88を介した微生物群のシグナル伝達は、全身の好中球性炎症反応に必要である。
Immunology. 2013; 140: 483-492https://doi.org/10.1111/imm.12159
記事で見る
グーグル・スカラー
ゴラ A.
ドリントンM.G.
スペランツァE.
サラC.
シーR.M.
ラドトケA.J.
ウォン・H.S.
バプティスタ A.P.
エルナンデス J.M.
カステラーニG.
他
肝臓の宿主防御を促進する宿主主導の免疫ゾーネーション。
ネイチャー(Nature)。2021; 589: 131-136https://doi.org/10.1038/s41586-020-2977-2
記事で見る
グーグル・スカラー
マクドナルド・B.
ズコロトA.Z.
ユウ・I.L.
ブルクハルトR.
ブラウン K.
ゲウキングM.B.
マッコイ K.D.
腸内細菌叢による血管内免疫ファイアウォールのプログラムにより、感染時の病原体の播種から保護される。
Cell Host Microbe. 2020; 28: 660-668.e4https://doi.org/10.1016/j.chom.2020.07.014
記事で見る
グーグル スカラー
フォルメス H.
ベルナルデスJ.P.
マンA.
バイエルF.
ポンタロロG.
キオウプツィK.
シェーファー K.
アッティグ S.
ニコロバ T.
ホフマン T.G.
ら。
腸内細菌叢は肝内皮細胞のトランスクリプトームを指示する。
iScience. 2021; 24: 103092https://doi.org/10.1016/J.ISCI.2021.103092
記事で見る
グーグル・スカラー
エルニーD.
フラビェ・デ・アンジェリスA.L.
ジャイティンD.
ヴィーゴファーP.
スタシェフスキ O.
デイビッド E.
ケレンショール H.
マフラコイヴT.
ヤコブシャゲン K.
ブッフ T.
et al.
宿主微生物が中枢神経系におけるミクログリアの成熟と機能を絶えず制御している。
Nat. Neurosci. 2015; 18: 965-977https://doi.org/10.1038/nn.4030
記事で見る
グーグル スカラー
オルツァック T.
アン・D.
ツァイシグ S.
ヴェラ M.P.
リヒターJ.
フランケ A.
グリックマンJ.N.
シーバートR.
バロンR.M.
カスパーD.L.
Blumberg R.S.
幼少期の微生物曝露はナチュラルキラーT細胞機能に持続的な影響を及ぼす。
サイエンス。2012; 336: 489-493https://doi.org/10.1126/science.1219328
記事で見る
Google Scholar
森田直樹
梅本英明
藤田 聡
林 敦
菊田淳一
木村伊織
羽田 毅
今井 崇
井上彰
三室秀樹.
他
細菌代謝物による腸管CX3CR1+細胞のGPR31依存的な樹状突起突出。
Nature. 2019; 566: 110-114https://doi.org/10.1038/s41586-019-0884-1
記事で見る
グーグル スカラー
イップ B.G.
キム・J.H.
リマ・R.
ズビトヌイックL.D.
ペトリB.
スワンランド N.
ホー M.
セト V.G.
タック T.
ケンダーマンL.
ら。
肺は即時型好中球を介した血管保護のための宿主防御ニッチである。
Sci. Immunol. 2017; 2: イーアム8929https://doi.org/10.1126/sciimmunol.aam8929
記事で見る
グーグル スカラー
イワムラ C.
ブーラドゥーN.
ベルカイドY.
シャー A.
ヤンコヴィッチD.
間葉系間質細胞におけるNOD1による微生物叢の感知がマウス造血を制御する。
Blood. 2017; 129: 171-176https://doi.org/10.1182/blood-2016-06-723742
記事で見る
グーグル スカラー
メネゼスG.B.
リー・W.Y.
Zhou H.
ウォーターハウスC.C.M.
カーラ D.C.
Kubes P.
内毒素血症肝微小循環における好中球Mac-1のIL-10による選択的ダウンレギュレーション。
J. Immunol. 2009; 183: 7557-7568https://doi.org/10.4049/jimmunol.0901786
記事で見る
Google Scholar
マクドナルド B.
ピットマンK.
メネゼスG.B.
ヒロタ・S.A.
スラバ I.
ウォーターハウス C.C.M.
ベック P.L.
ムルーヴェ D.A.
クベスP.
血管内危険信号が好中球を無菌性炎症部位に誘導する。
Science. 2010; 330: 362-366https://doi.org/10.1126/science.1195491
記事で見る
Google Scholar
マカボイ E.F.
マクドナルドB.
パーソンズS.A.
ウォン・C.H.
ランドマンR.
Kubes P.
内毒素血症時の肝微小循環内における好中球の動員におけるCD14の役割。
J. Immunol. 2011; 186: 2592-2601https://doi.org/10.4049/jimmunol.1002248
記事で見る
Google Scholar
ブルデットC.
サヤ=ジャンヌS.
グエン・T.T.
ヒューゴンP.
サブリエ=ガリスF.
サン・ルー N.
コーベル T.
フェレイラ S.
パルス・M.
ワイス・W.
ら。
Clostridium difficile感染症の動物モデルにおいて、抗生物質によるdysbiosisは死亡率を予測する。
Antimicrob. Agents Chemother. 2018; 62: 009255-18https://doi.org/10.1128/AAC.00925-18
記事で見る
グーグル スカラー
デセインR.
ボードゥアンM.
グランジャン T.
ル・ガーンR.
フィゲアック M.
ベーリー D.
フォール K.
ファベウ C.
グエリ B.
ゴッセ P.
キプニス E.
抗生物質による腸内細菌異常症は肺の免疫低下を誘発し、マウスの肺感染症を悪化させる。
Crit. Care. 2020; 24: 611-710https://doi.org/10.1186/S13054-020-03320-8/FIGURES/6
記事で見る
グーグル・スカラー
フリードバーグ D.E.
周M.J.
コーエンM.E.
アナバジャラM.K.
カーン S.
モスコーソ D.I.
ブルックス C.
ウィッティア S.
チョンD.H.
ウレマンA.C.
エイブラムス J.A.
集中治療室入室時の消化管マイクロバイオームの病原体コロニー形成とその後の死亡や感染症のリスク。
Intensive Care Med. 2018; 44: 1203-1211https://doi.org/10.1007/S00134-018-5268-8
記事で見る
グーグル・スカラー
シュー・R.
タン・C.
朱・J.
Zeng X.
Gao X.
Wu Q.
チェン Q.
Wang H.
Zhou H.
He Y.
et al.
神経難病患者における腸内細菌叢の異常と死亡リスク.
Crit. Care. 2019; 23: 195https://doi.org/10.1186/S13054-019-2488-4
記事で見る
グーグル・スカラー
プラット D.J.
ローレンス・D.
ロジャース・R.
シュリーファーL.
Qian W.
マイナー C.A.
メノスA.M.
ケネディ E.A.
ピーターソンS.T.
スティンソンW.A.
他
STING関連肺疾患に対する腸内細菌による移殖可能な保護。
セル・レップ 2021; 35: 109113https://doi.org/10.1016/j.celrep.2021.109113
記事で見る
グーグル スカラー
キョウ C.E.
キムD.H.J.
プセドゥM.M.
ノッツ・T.A.
ラバサG.
スラデック J.A.
シェーM.T.
ハニーカットM.
ブルスト=マスチャー I.
バルボーザ M.
ガロー M.G.
微生物叢-腸-脳軸の発達制御因子としてのミエリン。
Brain Behav. Immun. 2021; 91: 437-450https://doi.org/10.1016/J.BBI.2020.11.001
記事で見る
グーグル・スカラー
クベス・P.
ジェンヌC.
肝臓の免疫反応。
Annu. Rev. Immunol. 2018; 36: 247-277https://doi.org/10.1146/annurev-immunol-051116-052415
記事で見る
グーグル・スカラー
バルマー M.L.
スラックE.
デ・ゴッタルディ・A.
ローソンM.A.E.
ハプフェルマイヤーS.
ミーレL.
グリコ A.
ヴァン・ヴリエルベルヘH.
ファールナーR.
パトゥートN.
他
肝臓は、宿主と腸内常在菌の相互作用を媒介するファイヤーウォールとして機能する可能性がある。
Sci. Transl. Med. 2014; 6: 237ra66https://doi.org/10.1126/scitranslmed.3008618
記事で見る
グーグル・スカラー
オオクボ・タカ。
津田真一。
田村真理子
山村真理子
無菌ラットの末梢血好中球におけるスーパーオキシド産生の障害。
Scand. J. Immunol. 1990; 32: 727-729https://doi.org/10.1111/j.1365-3083.1990.tb03216.x
記事で見る
Google Scholar
クラーク T.B.
デイビス・K.M.
リセンコ E.S.
Zhou A.Y.
Yu Y.
Weiser J.N.
Nod1による微生物叢のペプチドグリカンの認識は、全身の自然免疫力を高める。
Nat. Med. 2010; 16: 228-231https://doi.org/10.1038/nm.2087
記事で見る
Google Scholar
カティブ・マッサルハ E.
バッタチャリヤS.
マサルハH.
ビラムA.
ゴラン K.
コレット O.
クマリ A.
アヴェマリア F.
ペトロビッチ=コピットマン E.
ガー-コーエンS.
et al.
炎症性骨髄好中球が放出する乳酸は、内皮GPR81シグナルを介してその動員を誘導する。
Nat. Commun. 2020; 11: 3547-3618https://doi.org/10.1038/s41467-020-17402-2
記事で見る
Google Scholar
リー Y.S.
キムT.Y.
キム・Y.
Kim S.
Lee S.H.
ソ・S.U.
Zhou B.O.
Eunju O.
キム・K.S.
Kweon M.N.
微生物叢由来の乳酸は、レプチン受容体+ニッチ細胞からの幹細胞因子産生を誘導することにより、造血および赤血球形成を促進する。
Exp. Mol. Med. 2021; 53: 1319-1331https://doi.org/10.1038/s12276-021-00667-y
記事で見る
グーグル スカラー
ソンG.
クレーマーM.
ハインズI.N.
肝臓病態に対する腸内細菌の寄与。
Gastroenterol. Res. Pract. 2010; 2010: 453563-453613https://doi.org/10.1155/2010/453563
記事で見る
Google Scholar
シュレヒト J.
ズコロト A.Z.
ユー I.L.
ドイグ C.J.
ダンバーM.J.
マッコイ K.D.
McDonald B.
重症患者における微生物-免疫メタシステムのディスバイオシスは、院内感染と関連している。
Nat. Med. 2023; 29: 1017-1027https://doi.org/10.1038/s41591-023-02243-5
記事で見る
Google Scholar
キム・S.M.
デファジオJ.R.
ヒョジュS.K.
サンガニ K.
ケイスキーR.
クレザレックM.A.
コダレフ・N.N.
サングワン N.
クリストリー S.
ハリス K.G.
ら
糞便微生物叢移植は、全身免疫を回復させることにより、ヒト病原体を介する敗血症からマウスを救う。
Nat. Commun. 2020; 11: 2354https://doi.org/10.1038/s41467-020-15545-w
記事で見る
グーグル スカラー
デフィリップZ.
ブルーム・P.P.
トーレス・ソトM.
マンスールM.K.
セイターM.R.A.
ハントリーM.H.
ターベット S.
チョン R.T.
チェン Y.-B.
Hohmann E.L.
糞便微生物叢移植により感染した薬剤耐性大腸菌による菌血症
N. Engl. J. Med. 2019; 381: 2043-2050https://doi.org/10.1056/NEJMoa1910437
記事で見る
グーグル・スカラー
ブリュギルーS.
ボイトラーM.
プファンC.
ガルゼッティD.
ルシェウィーH.J.
リング D.
ディール M.
ヘルプ S.
レッチェ Y.
フセイン S.
他
Salmonella Enterica serovar Typhimuriumに対するコロニー形成抵抗性を付与するマウスの微生物相をゲノム誘導で設計した。
Nat. Microbiol. 2016; 2: 16215https://doi.org/10.1038/nmicrobiol.2016.215
記事で見る
グーグル・スカラー
スールワルド B.G.J.
デニセットJ.F.
ゼンプF.J.
アムレインM.
オットーM.
コンリー J.
オムリ A.
イェーツ R.M.
クベスP.
生体内における黄色ブドウ球菌リザーバーの同定と治療。
J. Exp. Med. 2016; 213: 1141-1151https://doi.org/10.1084/jem.20160334
記事で見る
グーグル スカラー
サリム H.M.
フクダ・ディー.
ヤギ・S.
添木 崇
島袋雅彦
佐多 稔
新規選択的SGLT2阻害薬イプラグリフロジンによる血糖コントロールは、ストレプトゾトシン誘発糖尿病マウスの内皮機能不全を改善した。
Front. Cardiovasc. Med. 2016; 3: 43https://doi.org/10.3389/FCVM.2016.00043/XML/NLM
記事で見る
グーグル スカラー
ジェニンガ E.H.
ブッゲA.
ニールセンR.
ケルステンS.
ハマースN.
ダニ C.
ワビッチM.
ベルガーR.
シュトゥンネンベルグ H.G.
マンドラップ S.
カルクホーベン E.
ペルオキシソーム増殖剤活性化受容体γは、抗リポリンGタンパク質共役受容体81(GPR81/Gpr81)の発現を制御する。
J. Biol. Chem. 2009; 284: 26385-26393https://doi.org/10.1074/jbc.M109.040741
記事で見る
Google Scholar
ディアス F.J.
オブライエンM.J.
ウィグルスワースK.
エッピグJ.J.
マウス卵巣における前胸部顆粒膜細胞から卵丘細胞への移行:拡張を受ける能力の発達。
Dev. Biol. 2006; 299: 91-104https://doi.org/10.1016/j.ydbio.2006.07.012
記事で見る
Google Scholar
ゴメス-ネトJ.C.
マンツ・S.
ヘルドK.
シンハR.
セグラ・ムノスR.R.(Segura Munoz R.R.
シュマルツR.
ベンソン A.K.
ウォルター J.
Ramer-Tait A.E.
ノトバイオティックマウスにおけるAltered Schaedler Flora微生物叢の個々のメンバーの正確な定量化のためのリアルタイムPCRアッセイ。
J. Microbiol. Methods。2017; 135: 52-62https://doi.org/10.1016/j.mimet.2017.02.003
記事で見る
グーグル スカラー
キム・イー.
ヤン・S.-M.
リム・B.
パークS.H.
ラッカビーB.
Kim H.-Y.
96ウェルプレートで37種のLactobacillusを特異的に検出・同定するためのPCRアッセイの設計。
BMC Microbiol. 2020; 20: 96https://doi.org/10.1186/s12866-020-01781-z
記事で見る
グーグル・スカラー
ラコフ=ナホウム S.
パグリーノJ.
エスラミ=ヴァルザネF.
エドバーグS.
メジトフR.
Toll様受容体による常在細菌叢の認識は、腸のホメオスタシスに必要である。
Cell. 2004; 118: 229-241https://doi.org/10.1016/j.cell.2004.07.002
記事で見る
Google Scholar
マルケス P.E.
アントゥネスM.M.
デビッドB.A.
ペレイラ R. v.
テイシェイラM.M.
メネゼスG.B.
従来の共焦点顕微鏡を用いたin vivoでの肝臓生物学のイメージング。
Nat. Protoc. 2015; 10: 258-268https://doi.org/10.1038/nprot.2015.006
記事で見る
グーグル スカラー
リーE.K.S.
ギルリーM.R.
リーL.
アルナソンJ.W.
キム・J.H.
バベス・L.
ルー Y.
サナティ-ネズハドA.
カイ・S.K.
ケリーM.M.
他
ロイコトリエンB4を介した好中球の動員は、致死的な真菌性敗血症の際に肺毛細血管炎を引き起こす。
Cell Host Microbe. 2018; 23: 121-133.e4https://doi.org/10.1016/j.chom.2017.11.009
記事で見る
グーグル スカラー
ブラッドリーP.
クリステンセンR.
ロススタインG.
化膿性炎症における細胞および細胞外ミエロペルオキシダーデ。
血液。1982; 60: 618-622https://doi.org/10.1182/blood.V60.3.618.618
記事で見る
Google Scholar
オリンA.
ヘンケルE.
チェン Y.
ラクシュミカントT.
プー C.
マイクス J.
グスタフソン A.
ベルンハードソンA.K.
チャン・C.
ボーリン K.
ブロディン P.
新生児における定型的な免疫系の発達。
Cell. 2018; 174: 1277-1292.e14https://doi.org/10.1016/J.CELL.2018.06.045
記事で見る
グーグル スカラー
デイビッドB.A.
レゼンデ・R.M.
アントゥネスM.M.
サントスM.M.
フレイタス・ロペス M.A.
ディニーズA.B.
ソウサ・ペレイラR.V.
マルケージS.C.
アルヴァレンガ D.M.
ナカガキB.N.
他
マスサイトメトリーと画像解析の組み合わせにより、マウスにおける肝ミエロイド細胞の起源、位置、および機能的再増殖を明らかにした。
Gastroenterology. 2016; 151: 1176-1191https://doi.org/10.1053/j.gastro.2016.08.024
記事で見る
グーグル スカラー
ワン・J.
ニウ・N.
Xu S.
ジン Z.G.
マウス肺内皮細胞単離のためのシンプルなプロトコール。
Sci. Rep. 2019; 9: 1458-1510https://doi.org/10.1038/s41598-018-37130-4
記事で見る
グーグル・スカラー
記事情報
出版物の歴史
オンラインで公開されました: 2023年5月16日
受理された: 2023年4月28日
改訂版受理 2023年3月20日
受理された: 2022年11月21日
識別情報
DOI: https://doi.org/10.1016/j.celrep.2023.112507
著作権について
© 2023 The Author(s).
ユーザーライセンス
クリエイティブ・コモンズ 表示 - 非商用 - 非デリバリー (CC BY-NC-ND 4.0) |.
再利用の方法
サイエンスダイレクト
ScienceDirectでこの論文にアクセスする
図版
グラフの概要
図1腸内細菌叢は、全身性細菌感染時の肝臓への好中球輸送を制御する
図2腸内細菌叢由来のD-乳酸は肝臓への好中球の動員を制御する
図3腸管由来D-乳酸による好中球輸送の臓器選択的制御
図4腸由来D-乳酸による好中球輸送の制御は、顆粒球形成や循環好中球の機能調節とは無関係である
図5腸由来のD-乳酸は血管内皮とのクロストークを通じて肝臓の好中球輸送を制御する
図6抗生物質誘発性ディスバイオシス中の微生物叢のD-乳酸産生を治療的に補正すると、血流感染に対する好中球の血管内宿主防御が回復される
関連記事
本サイトのコンテンツは、あらゆる分野の医療従事者や研究者を対象としています。
研究ジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノミクス
細胞宿主と微生物
細胞代謝
セルレポート
セルレポートメディスン
セルレポートメソッド
セルレポート 物理科学
セルレポート サステイナビリティ
細胞幹細胞
細胞システム
化学
化学触媒
カレントバイオロジー
発生細胞
デバイス
ヘリオン
イミュニティ
アイサイエンス
ジュール
物質
医学
分子細胞
ニューロン
一つの地球
パターン
STAR プロトコル
構造
トレンドレビュー誌
生物化学
バイオテクノロジー
癌
細胞生物学
化学
認知科学
エコロジー&エボリューション
内分泌学・代謝学
遺伝学
免疫学
微生物学
分子医学
神経科学
寄生虫学
薬理学
植物科学
パートナージャーナル
AJHG
生物物理学雑誌
生物物理学レポート
HGGアドバンス
モレキュラープラント
分子療法ファミリー
ネクサス
植物通信
幹細胞レポート
イノベーション
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
コンソーシアムハブ
Nucleus Collections
スナップショット・アーカイブ
ジャーナルを越えて
Cellキャリアネットワーク
セルメンター
細胞シンポジューム
ラボリンク
ウェビナー
記事を進化させる
コミュニティレビュー
Figure360
スニークピーク
STARメソッド
サイエンス・イン・ソサイエティ
セル画展
セルプレスポッドキャスト
セルプレスビデオ
ぬりえ・コミック
リサーチアーク
コネクト
セルプレスについて
採用情報
お問い合わせ
ヘルプ&サポート
ニュースルーム
出版物アラート
アクセス
購読する
今すぐ読む
図書館に薦める
INFORMATION
広告主様向け
採用担当者様へ
図書館員の方へ
ご利用条件
個人情報保護方針
アクセシビリティ
当社は、サービスの提供・向上およびコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を変更するには、このサイトのクッキー設定にアクセスしてください。
著作権 © 2023 Elsevier Inc.ただし、第三者が提供する一部のコンテンツは除きます。
この記事が気に入ったらサポートをしてみませんか?