乳児期の微生物と代謝産物が小児期の神経発達障害を指し示す

メインコンテンツへスキップ
セル
ログイン
検索...
記事|今すぐオンライン
乳児期の微生物と代謝産物が小児期の神経発達障害を指し示す
https://www.cell.com/cell/fulltext/S0092-8674%2824%2900238-1

アンジェリカ・P・アーレンス
トゥーリア・ヒョーティライネン
ジョセフ-R-ペトローネ
マテイ・オレシッチ
エリック・W・トリプレット
ジョニー・ルドヴィグソン
すべての著者を表示する
脚注を表示オープンアクセス掲載:2024年04月03日DOI:https://doi.org/10.1016/j.cell.2024.02.035
ハイライト
乳児の微生物と代謝産物が対照群と将来のNDを区別する
早期の耳炎は将来のNDにおいてコプロコッカスを減少させ、シトロバクターを増加させる。
早産、感染症、ストレス、親の喫煙、HLA DR4-DQ8はNDリスクを増加させる。
将来のASDの臍帯血清ではリノレン酸が低く、PFDA毒素が高い。
研究概要
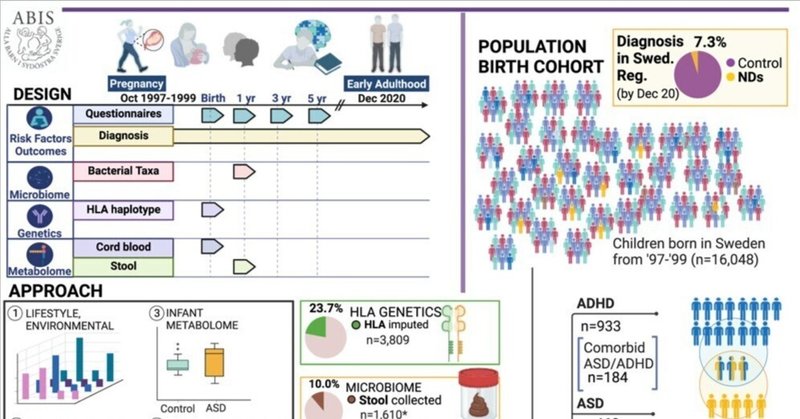
本研究は、神経発達障害(ND)の診断に関連する因子を見つけるために、出生コホートを20年以上追跡した。詳細な早期からの縦断的質問票により、感染症や抗生物質のイベント、ストレス、出生前因子、家族歴などを把握した。臍帯血清のメタボロームとリピドーム、ヒト白血球抗原(HLA)遺伝子型、乳児の微生物叢、便のメタボロームなどのバイオマーカーが評価された。16,440人のスウェーデン人小児のうち、1,197人がNDを発症した。将来のND診断全般と、知的障害、言語障害、注意欠陥・多動性障害、自閉症にまたがる特定のNDサブタイプに有意な関連が現れた。この調査により、将来的な診断だけでなく、早期に出現する気分障害や胃腸障害とマイクロバイオームとの関連が明らかになった。調査結果は、ストレス、早期感染、抗生物質によって複合的にもたらされる免疫調節や代謝との関連を示唆している。大規模集団を対象としたこの前向き縦断研究における乳児バイオマーカーとリスク因子の収束は、神経発達における早期の予測および介入の基盤を確立するものである。
図抄録
図サムネイルfx1
大きな画像を見るダウンロード 高解像度画像
キーワード
アッカーマンシア
抗生物質
ASD菌
コプロコッカス
エクオール
炎症
耳炎
小児科
PFOA
トリプトファン
はじめに
神経発達障害(ND)は中枢神経系の成熟に重大かつ永続的な影響を及ぼし、小児期に発症することが多い。神経発達障害には、自閉症スペクトラム障害(ASD)、注意欠陥・多動性障害(ADHD)、知的障害、コミュニケーション障害などがある。ASDは、社会的な困難や制限的で反復的な行動や興味を特徴とし、人口の1~2%が罹患している。一方、ADHDの有病率は小児で7%1、成人で3.4%に達し2、身体的・精神的な健康負担が大きい3。
NDの病因には様々な出生前および早期生活要因6,7が関与している可能性があり、特異的なバイオマーカーがないために早期診断が困難である。遺伝的および環境的影響は、神経適応性を乱し、神経細胞のホメオスタシス異常を引き起こす可能性がある一方で8、免疫調節異常、炎症、自己抗体も関与している可能性がある9,10。ASD患者のかなりの部分は、特に胃腸(GI)症状を併発しており、腸と脳の関連を示唆している11,12,13。
腸内細菌は、代謝、消化管の健康、神経の健康、14,15,16、免疫機能において重要な役割を果たしており、17,18,19,20,21はNDへの関与の可能性を示唆している。腸と脳のコミュニケーションは、迷走神経を介したり、短鎖脂肪酸(SCFA)、サイトカイン、アミノ酸、神経伝達物質前駆体の輸送など、様々な方法で行われている22。微生物は神経伝達物質を局所的に合成することができ23,24、腸内分泌細胞は免疫とミクログリアの機能を調節するホルモンと樹状細胞を放出し、最終的には行動に影響を与える神経活性代謝産物に影響を与える25,26。ASDと診断された小児を対象とした小規模なパイロット研究では、マイクロバイオーム移入療法後に、消化管症状およびASD関連症状の改善が観察され29、2年後まで効果が持続することも確認された30。しかし、マイクロバイオームの乱れがNDの病因や病態生理に正確に寄与しているかどうかは、ASD31とADHDに関する横断的研究において所見が分かれており、依然として不明である32。
初期の腸内細菌叢と認知との関連はまだ十分に理解されておらず、既存の研究は3件のみで、医学的に記録されたNDを対象とした前向きマイクロバイオーム研究はまだない。これらの研究では、マイクロバイオームの多様性とAges and States Questionnaires(ASQ-3)スコア33、定型発達児の認知機能34、3歳児の社会的行動35との相関が検討されている。
このギャップを解決するために、私たちの研究は、All Babies in Southeast Sweden(ABIS)コホートにおいて、臍帯血や便などの生物学的サンプルを収集し、早期生活因子を調査することを目的としている。出生時からの健康記録と包括的な早期生活データを分析し、NDリスクと関連するマイクロバイオームと代謝産物のシグネチャーを同定した。
結果
将来のND診断に関連する因子を特定するために、一連のバイオマーカーと質問票を分析した。いくつかの因子はNDのリスク上昇と関連していたが、他の因子はリスク低下と関連していた。質問票の結果が示され、バイオマーカーの結果およびそれらの相互作用を考慮した統合結果が続く。
アンケートでは、感染症や抗生物質によるイベント、化学物質への暴露、家族歴、その他の医学的問題、重大なライフイベントなど、多くのNDリスク因子が生後早期に明らかになった。
ABISの質問票データは、妊娠期から小児期を通じて、いくつかの時期に参加児童の両親から得られた。ここでは、より広範なABIS資料の中から5歳までのデータを分析した(コホートと診断日については図1Aと1Bを参照)。質問票データの範囲は詳細である。図1CとS1は、それぞれ複合的および個別的なNDの転帰と、以下に述べる危険因子との関連を示している。
図サムネイルgr1
図1神経発達障害(ND)に関与する早期生活環境および医学的危険因子
キャプション
大きな画像を見るダウンロード 高解像度画像
感染と抗生物質投与
幼児期(出生から5歳まで)の感染症は、ADHDまたはASDのリスク上昇と有意に相関していた。この時期にペニシリンを必要とする感染症を3回以上経験した人は、将来ND、例えば言語障害(オッズ比[OR]=3.89[2.14-7.05、95%信頼区間、CI])、ADHD(OR=3.27[2.29-4.67])、または知的障害(OR=2.44[1.18-5.06])になりやすかった。1~2.5年の間にペニシリンを使用した場合、将来のASDの可能性が1.6倍(1.2~2.1、95%CI)高くなった(p = 0.0030)。また、将来ASDになる可能性のある子どもは、この時期にペニシリン以外の抗生物質を使用していた可能性が高く(OR = 1.5 [1.0-2.1], p = 0.0292)、将来知的障害になる可能性のある子どもの23.8%は、ペニシリン以外の抗生物質を使用していた(OR = 2.2 [1.1-4.4, 95%CI]、p = 0.0337)。頻回の耳炎エピソード(1~2.5歳の間に3回以上)を経験した小児は、後に知的障害、ASD、ADHDと診断される可能性がそれぞれ2.13倍(1.1~4.13、95%CI)、1.74倍(1.21~2.51、95%CI)、1.75倍(1.33~2.30、95%CI)高かった。2.5年から5年の間、パラセタモール解熱剤の使用回数が増加(6回以上)するとASDリスクが上昇した(OR = 1.82 [1.16-2.88, 95%CI])。この時期のペニシリンの使用はADHDのリスクを1.54倍、ASDのリスクを1.76倍上昇させた。抗生物質を必要とする感染症が少なくとも3~6回あった小児は、ND、特にADHDを発症する可能性が1.58~2.39倍高かった(OR = 1.62-2.9)。将来言語障害になる可能性のある子供は、3回以上の耳炎を経験すると1.85-2.27倍高かった。これは将来の知的障害でもみられた。2.5~5年間に頻繁に胃腸炎(3回以上)を起こしたのは、対照群の26.9%に対して、将来のNDの33.8%、将来のADHDの35.8%であった。
化学物質曝露
妊娠中の母親の喫煙は、NDの累積リスク(OR=3.0[2.33-3.87、95%CI])だけでなく、ASD(OR=3.72[1.92-7.21、95%CI])およびADHD(OR=3.31[2.52-4.34、95%CI])の個別リスク、特に1日10本以上の喫煙リスクをもたらした。同様に、妊娠中の母親の鎮痛薬の使用は、ADHD(OR=1.41[1.23-1.62、95%CI])とASD(OR=1.46[1.19-1.78、95%CI])のリスクを増加させた。1~2.5年間の親の喫煙への曝露は、言語障害を除くすべてのNDにおいて用量依存的にリスク上昇と関連していた。最も顕著であったのは、母親が15本/日以上喫煙していた子どもにおけるADHD(OR = 4.88[3.23-7.36])、父親が15本/日以上喫煙していた子どもにおけるASD(OR = 3.47[2.01-6.01])であった。両親の喫煙または家庭内での喫煙全般に曝された幼児は、言語障害を除くすべての診断群において、用量依存的なNDリスクの上昇を示した。例えば、対照群の8.8%と10.2%しか父親または母親が喫煙していなかったが、将来の知的障害を持つ幼児の20.5%に父親が喫煙しており、30%に母親が喫煙していた。最もリスクが高かったのは将来のADHDで、母親が1日15本以上喫煙していた場合、ADHDの可能性が6.05倍(3.68-9.94、95%CI)増加した。
家族歴
喘息、セリアック病(CD)、1型糖尿病(T1D)の家族歴のある両親から生まれた子どもは、NDのリスクが高いことが示された(OR = 1.28-1.46)。父親の喘息は、ASD(OR = 1.71、[1.2-2.42、95%CI])やADHD(OR = 1.56、[1.20-2.02、95%CI])のリスクと最も強い関連を示した。しかし、これらの因子は言語障害や知的障害には有意な関連を示さなかった。
その他の医学的問題
初期の消化器系の問題、および気分の問題は、将来のNDを持つ小児に顕著であった。2.5歳および5歳時の症状(図2Aおよび2B)は、主成分分析(PCA)により「気分と不安」、「子どもの成長」、「GI問題」成分に縮小され、2.5歳時で57.7%、5歳時で58.3%の累積分散を説明した。2.5歳では、気分と不安の症状は将来のNDと相関し(OR = 2.05-3.46、95%CI [1.35-1.74 to 2.90-6.91])、特に将来の知的障害において顕著であった(OR = 3.46、95%CI [1.74-6.91]、症例17.9%対対照5.9%;図2C)。GI問題クラスターの中では、胃痛、腹部膨満感またはガス状胃、便秘のような症状は、将来の知的障害の21.3%にみられた(OR = 2.4、95%CI [1.2-4.9]、p = 0.013)。将来のASDについては有意ではなかったが、5.6%が2つ以上の症状を有していたのに対し、対照群では3.5%であった(p = 0.077)。5歳の時点で、「胃痛」と「腹痛」は、対照群(8.0%-8.9%)に比べ、将来のNDでより多く(13.5%-24.6%)みられ、将来のASD/ADHDの併存で特に顕著であった(OR = 3.39-3.45、p < 0.0001)。過敏症や不機嫌症は、将来のASDやADHDでより一般的であったが(将来の症例では5.0%-7.7%、対照群では2.4%-2.5%)、GI症状よりは頻度が低かった。
図サムネイルgr2
図22.5年および5年時の症状クラスターと1年目の微生物種の豊富さとの関連
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
重大なライフイベント
出生から5年までの深刻なライフイベント(例えば、別居/離婚、家族の死、重篤な病気/事故、失業)は、将来のNDの可能性を1.98倍(1.6-2.44、95%CI)増加させ、対照群の19.8%に比べて32.8%の小児に影響を与えた。妊娠中の深刻なライフイベントも将来のADHDと関連していた(OR=1.62[1.33-1.98、95%CI])。驚くべきことに、母親が妊娠中に喫煙していた子ども群では、将来のASDの85%が5歳までにそのような出来事を経験していたのに対し、対照群では40%にすぎなかった。早産児は、将来のND診断(ASDを除く)の可能性が1.4倍(1.12-1.87、95%CI)増加した。また、知的障害群では早産との関連がみられ、対照群の4.2%に対して15.2%にみられた(OR = 4.13[2.51-6.77、95%CI])。言語障害または知的障害のある小児は、帝王切開(C-section)で出産する確率が1.93~2.02倍高かった。
バイオマーカーデータにより、臍帯血清メタボローム、HLA遺伝子型、乳児腸内細菌叢、便メタボロームにおける有意差が明らかになった。
バイオマーカーデータは、出生時の臍帯血清と平均1歳時の便サンプルから収集された。HLAと乳児腸内細菌叢には、ND診断の集計とサブタイプによる違いが認められた。ASDにおいてのみ、臍帯血清メタボローム、臍帯血清リピドーム、便メタボロームを検討した。
臍帯血清メタボロームとリピドーム
臍帯血清リピドームとメタボロームの違いを評価するために(図3A-3K)、母親の感染症37、喫煙38、ストレス/深刻なライフイベント39、食事40,41、ビタミン/サプリメント、コーヒーの摂取42、さらに両親の学歴と妊娠年齢を含む出生前のリスク因子を、傾向スコアマッチングを用いてコントロールし、将来のASD症例(n=27)に対してマッチした対照(n=27)を選択した。
図サムネイルgr3
図3将来のASDにおけるコード血清メタボロームと便脂肪酸の違い(ABISASD)
キャプション
大きな画像を見るダウンロード 高解像度画像
リノレン酸(LA)は出生前の因子でマッチングした後、将来のASDと最も強い逆相関を示した(p = 0.00288;図3A)。逆に、ペルフルオロデカン酸(PFDA)は将来のASDと最も有意な正の相関を示した(p = 0.0087;図3B)。ゼアロンも将来のASDで高かった(p = 0.01;図3C)。対照群で高かったのは、3-カルボキシ-4-メチル-5-プロピル-2-フランプロパノエート、C16:1、デカン酸、パルミチン酸であった(図3D-3G)。有意に近いが、オクタン酸(p = 0.054)、オレイン酸(p = 0.10)、ウルソデオキシコール酸(UDCA)は一致した対照群で高く(図3H、3J、3K)、タウロ-β-ムリコール酸(TbMCA)は将来の症例で高かった(図3I)。リピドミクスの結果、コントロールの臍帯血清には8種類のトリグリセリド(TG)が濃縮されていた(ps < 0.05): TG(14:0/18:2/18:2)、質量電荷比(m/z)822.75;TG(46:2)/(16:1/14:0/16:1);TG(48:3);TG(49:1);TG(49:2);TG(51:1);TG(51:2)、m/z862. 79;TG(51:3);TG(53:2);TG(53:3)、m/z 888.80;TG(53:4);およびTG(58:1)。
HLA遺伝子型
家族性自己免疫疾患は、ND(特にASD43,44,45)を持つ人に多くみられ、その大部分は、免疫を媒介するHLA遺伝によって引き起こされる。HLAはまた、宿主と微生物の相互作用においても重要な役割を果たしている。そこでわれわれは、ABISの3,809人の小児(将来のND327人を含む)についてHLAクラスIIハプロタイピングを行った。DR4-DQ8ホモ接合体(対照群の2.9%にみられた)は、将来のND(5.1%;OR = 1.8[1.03-3.13]、p = 0.039)と将来のASD(7.6%;OR = 2.8[1.44-5.25]、p = 0.0021)で特に顕著であった。DR4-DQ8対立遺伝子は、古典的に自己免疫疾患46、特にT1DとCDの発症率と重症度と関連している。
乳児腸内細菌叢
11.9±2.9ヵ月の乳児1,748人から便が採取され、平均61,994リード/サンプルであった。便採取時の年齢、便1グラムあたりの16S rRNAのコピー数は、累積的にもNDサブタイプによっても、症例と対照の間で差がなかった(p's > 0.62)。われわれは、将来のNDの状態に基づいて、存在量の異なる細菌を同定しようとした。一般化可能性を考慮し、まず、入手可能なすべての対照群と将来のNDを比較し、次にリスク因子とマイクロバイオームの多様性の交絡因子、すなわち分娩様式(p = 0.003)、地理(p = 0.001)、初年度の抗生物質曝露(p = 0.005)、初年度の耳炎(p = 0.006)についてマッチングを行った。心理社会的脆弱性指数(p=0.08)と生物学的性別(p=0.10)は有意に近づいていたが、その他の感染症と妊娠中の喫煙は関連を示さなかった(表S1)。
将来のND状態は、個々の分類群(図4A)と共通の細菌コア47(図S2)の両方によって区別可能であった。対照群では、Akkermansia muciniphila、Roseburia hominis、Erysipelotrichiaceae UCG-003属、Adlercreutzia equolifaciens、Alistipes putredinis、Phascolarctobacterium、Coprococcus、Bifidobacterium sp.などの細菌が濃厚であった(図4A)。Veillonella parvula、Megamonas funiformis、ASV-77 Bacteroides sp.、およびASV-103 Klebsiella sp.は、将来の症例で高くなった。表S2に最も大きな差異を示す。いくつかの中核種は安定していたが(図S2)、Bacteroides uniformisとBacteroides vulgatus(4つのNDすべて)、Veillonella parvula(ASDとADHD)、Escherichia-Shigella(ASD、ADHD、知的障害)、Enterobacter(ASDと知的障害)など、特異的なものもあった。違いはシーケンスの深さによるものではなく、希薄なデータセットでも希薄でないデータセットでも持続した。
図のサムネイルgr4
図4微生物交絡因子とNDリスク因子のマッチング(ABISND-Match)の前後における、全ABISControls(n = 1,456)と将来のNDであるABISND(n = 116)の分類学的およびコアマイクロバイオームの差異
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
サムネイル図1
図S1図1に関連する、ADHD、言語障害、知的障害に関与する早期の生活環境および医学的危険因子
キャプション
画像を拡大する 高解像度画像
サムネイル図2
図S2図2に関連した、2.5歳時の症状と1歳時の微生物量との関連
キャプションをすべて表示
画像を拡大する 高解像度画像
交絡因子調整後の微生物群集の違い(図4Cおよび4D)により、ABISND-Match(n=82)とマッチさせた対照(n=163)では、中核となる微生物群集が異なっていることが示された。微生物有病率60%では、マッチ群間の分離が観察された(図4C;表S4)。乳酸桿菌科、Akkermansiaceae、Christensenellaceae、Erysipelotrichaceae UCG-003、AV-81 Bifidobacterium breve、Phascolarctobacterium faeciumなどがマッチドコントロールで濃縮されたのに対し、ABISND-MatchではBacteroidota、Enterobacteriaceae、Prevotella 9、Parabacteroides merdae、Barnesiellaceaeが濃縮された(図4D)。
乳児腸内細菌叢(NDサブタイプ別)
NDの重症度のスペクトラムと多様な特徴を考慮して、サブタイプ、すなわちASD、ADHD、言語障害、および複数のNDによる違いを検討した。コアマイクロバイオームはサブタイプによって有意に異なっていたが、いくつかの違いは共有されていた(図4E)。主成分でサブタイプ(n = 114)を分離した結果、将来複数のND(ABISmultiple)を発症する乳児のコア微生物叢は将来ASD(ABISASD)と重なる一方、ADHD(ABISADHD)と言語障害(ABISSpeech)はともに有病率が65%と大きく異なっていた。ND間で有意に多かった分類群には、Megamonas funiformis(全ND、ABISADHD)とASV-86 Enterobacter sp.(ABISASD)が含まれ、ABISSpeechではSutterella、ASV-252 Clostridium sensu stricto 1 neonatale、ASV-184 Blautia sp.、ASV-155 Bacteroides sp.が含まれた(表S5)。
コアのばらつきに対処するため、対照群を将来のASD、ADHD、言語障害、または複数のNDと別々に比較した(図5A-5D)。すべてのサブタイプにおいて、178種類の細菌(主にClostridia)が対照群でより豊富であった(図S3)。特に、Akkermansia muciniphila、Phascolarctobacterium faecium、Roseburia hominis、Coprococcus eutactus、Coprococcus comes、Bacteroides ovatus、Bifidobacterium breve、Alistipes putredinisは、サブタイプにかかわらず、将来のNDでは一貫して発現量が少なかった(図S3)。ABISSpeechではASV-184 Blautia sp.とASV-155 Bacteroides sp.が多かった。ABISASDではCarnobacteriaceaeとASV-86 Enterobacter sp.が、ABISADHDではMegamonas funiformisとASV-77 Bacteroides sp.が高かった。ABISSpeechではAdlercreutziaもChristensenellaも観察されなかった。Coprococcus eutactusとBacteroides stercorisは、すべてのABISで有病率が低かったが、ABISASDやASD/ADHD合併例では観察されなかった。興味深いことに、ビフィドバクテリウムの有病率は将来の言語障害で異なり、対照群の98.1%に対しABISSpeech群では85.7%に認められたが(OR = 0.11、p = 0.0059)、他のNDでは認められなかった。
図サムネイルgr5
図5将来の神経発達障害(ND)サブグループ間およびHLAリスクハプロタイプ間の腸内細菌叢の違い
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
サムネイル図3
図S3図4および図5に関連する、対照群と神経発達障害(ND)にまたがるコア微生物叢
キャプションを表示
画像を拡大する 高解像度画像
ABISASDは、重症度の潜在的な差と男性におけるASDの高い有病率を考慮してさらに細分化された49(図S4)。晩期診断の男性(n = 12)は、早期診断の男性(n = 11)と比較して、Veillonellales-Selenomonadales、Coprococcus、Akkermansia muciniphila、Ruminococcus gauvreauiiが高かった。早期診断の男性では、Parabacteroides distasonis、Sutterella wadsworthensis、Prevotella 9 copri、ASV-18および82 Bacteroides sp.、ASV-86 Enterobacter sp、 女性では、ASDの早期診断(n = 8)はBlautia、Bacteroides sp.、Citrobacter、Enterococcus sp.、Veillonella sp.、Subdoligranulum sp.に属するASVの増加と関連しており、一方、晩期診断(n = 8)はBacteroidesのASV 77、82、155の増加と関連していた。しかし、ここでの相違はサンプル数が少ないためと考えられ、過大解釈すべきではない。
サムネイル図4
図S4ABIS対照群ではすべてのNDカテゴリーと比較して腸内細菌叢が増加している(図5に関連)。
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
乳児便メタボローム
生物学的性別と市町村の傾向スコアマッチングによって選択された、将来のASD(n=23)対対照(n=23)のサブセットにおいて、便のメタボロームの違いが観察された(図6A-6C)。ポジティブモードでは19の有意差のある代謝物が同定され、ネガティブモードでは124の代謝物が同定された(p's < 0.05)。将来のASDは3-イソプロピルマレイン酸(FC = 6.3, p = 0.0024)とキナ酸(FC = 4.0, p = 0.035)の高いレベルを示し、対照は2-ヒドロキシフェニル酢酸(FC = 0.41, p = 0.032)、L-リジン(FC = 0.69, p = 0.016)、グルタル酸(FC = 0.4, p = 0.032)の高いレベルを示した。その他の有意差は表S6(p's < 0. 05)、最も注目すべきは、グルタル酸とL-アルギニン、L-セリン、L-システイン-S-硫酸、ニコチン酸、ピコリン酸の様々なイオン形成(対照群で高い)、および3-デヒドロシキメートである、 ピリドキサミン、N-アセチル-DL-セリン、グルタル酸、α-アミノアジピン酸-N-メチル-L-グルタミン酸、(R,R)-酒石酸、プロリン、N-BOC-L-アスパラギン酸、5-ヒドロキシメチル-2-フルアルデヒド(将来のASDで高い)。
図サムネイルgr6
図6便代謝産物は将来のASDおよび1年後のアッカーマンシアおよびコプロコッカスの存在量と強い相関がある。
キャプション
大きな画像を見るダウンロード 高解像度画像
主要な化合物の存在量を調査したところ、エクオールと酪酸産生の明確な傾向を強調する興味深いパターンが観察された。エクオール(抗酸化物質として知られている)のいくつかの産生量が対照群で一貫して高かったことから、潜在的なエクオールのシグナルを同定しようとした。エクオールの潜在的なm/zが同定された(m/z 241.0866)。シグナルは低いものの、ppmの質量誤差(1.66)は装置の精度と一致していた。対照群と比較して、将来の症例からのこの化合物の減少は有意であった(4,168.98±743.38と4,814.55±984.03、p = 0.008)。
SCFAと関連代謝物に焦点を移すと、酪酸産生者の増加にもかかわらず、酪酸シグナルは群間で実質的な差はなかった(p = 0.38)。このことは、観察されたエクオールの傾向と対照的であり、酪酸生産者の増加が酪酸レベルの上昇に対応しなかったことを示唆している。
部分最小二乗-判別分析(PLS-DA)では、正イオンモードと負イオンモードでそれぞれ24個と75個の有意な特徴が同定され、投影における重要度(VIP)スコア≧2であった(図S5)。ポジティブモードで対照群で高値を示した注目すべき代謝物には、m/z 171.1490、m/z 144.1018(N-メチルピペコール酸と思われる)、m/z 185.1283(N-(3-アセトアミドプロピル)ピロリジン-2-オンと思われる)、m/z 167. 0563(1-メチルキサンチンと思われる)、m/z 148.0967(N-(2-ヒドロキシエチル)-モルホリンN-オキシドと思われる)、m/z 144.1018(N-メチルピペコール酸と思われる)、m/z 159.0440(1,2-ナプトキノンと思われる)。逆に、m/z 149.0446(3-ヒドロキシグルタル酸と思われる)、Ser-Pro(m/z 203.1026)、m/z 253.093(ガラクトシルグリセロールと思われる)は将来のASDで高くなった。コントロールで上昇したネガティブモード代謝物としては、m/z 241.1193(ピログルタミルロイシンの可能性が高い)、His-Pro(m/z 251.1147)、m/z 132.0666(N-メチルスレオニンまたは3-ヒドロキシバリンの可能性が高い)、m/z 182.0126(ホモシステイン酸または8-ヒドロキシチオグアニンの可能性が高い)が注目される。グルタル酸、L-グルタミン、リボフラビンは対照群で高く、N-メチル-L-ヒスチジン、3-アミノイソブタン酸、3-イソプロピルマレイン酸、N-アセチル-DL-グルタミン酸は将来のASDで高かった。
サムネイル図5
図S5将来の自閉症スペクトラム障害(ASD)における診断年齢による腸内細菌叢の違い(N=39)、図5に関連する
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
検出された22種類の脂肪酸のうち、2つの顕著な違いが観察された。ABISASDはパルミトレイン酸(16:1)を欠く傾向が強く、X2(1、n=46)=3.9、p=0.049であり、対照群では43.5%に認められたが、将来のASDでは87.0%に認められなかった。逆に、便中のパルミチン酸(16:0)のモル割合は、ABISASDでは42.6%高く(27.4%±2.2%ABISASD;15.8%±16.5%対照;p = 0.056;図3L)、これは臍帯血清では低かったので興味深い。
質問票データとバイオマーカーデータの統合により、早期発症の消化器系および気分の問題、環境因子、およびHLA遺伝子型と細菌との関連が明らかになった。
次に、初期症状、危険因子、および観察されたメタボロミクスの違いに関連する微生物の相互作用を調べた。
初期のGIおよび気分の問題
プロテオバクテリアは、2.5歳時点でGI問題を有する小児において2.0-2.2倍高かった(q = 0.011, q = 0.048)。CoprococcusとSlackia isoflavoniconvertensは保護傾向を示し(図2D)、log2 FCは4.22~6.10で、GI問題クラスターおよび気分・不安クラスターと逆相関していた。Akkermansia muciniphilaとAlistipes fingoldiiは気分や不安の症状と逆相関を示し、Coprococcus eutactusは消化器系の問題クラスターと負の相関を示した。将来腹痛や下痢を起こさない乳児では、Adlercreutzia equolifaciens、Erysipelotrichaceae UCG-003 spp.、Roseburiaが高く、Robinsoniella peoriensisとMegasphaera micronuciformisが低かった。
2.5歳時の気分との関連と同様に、Akkermansia属、Adlercreutzia属、Coprococcus属、およびRoseburia属は、5歳時に気分や消化器症状のない乳児でより多かった(図6E)。Akkermansia muciniphila(FC = 2.9)、Blautia obeum(FC = 3.0)、およびTuricibacter sanguinis(FC = 1.6)は、全般的な苛立ちや不機嫌と逆相関していた。Coprococcus comesとCoprococcus eutactusは、胃痛クラスターと気分症状の両方に逆相関した(FC = 2.2-2.7)。その他の分類群では、Anaerostipes caccae、Adlercreutzia equolifaciens、Roseburia inulinivoransが両クラスタと逆相関していた。
環境要因
次に、将来のNDリスク減少(log2 FC ≥ 2;図4B;表S3)または増加(図7B;表S3)をもたらす早期生活因子との関連において、発現量の異なる細菌を調べた。保護的な早期生活因子は対照群で高い細菌と相関し、ABISNDで高い細菌は帝王切開出産、抗生物質の使用、感染症、乳児の食事と相関した。母親が妊娠中に喫煙しなかった乳児では、Akkermansia muciniphila、Bacteroides ovatus、Phascolarctobacterium faecium、Alistipes、Bifidobacterium breveが高かった。Coprococcus comes、Adlercreutzia equolifaciens、Lacticaseibacillus、Bifidobacteriumが高いほど、心理社会的脆弱性スコアが低かった。抗生物質に曝露されたことのない乳児では、対照群と関連する5つの分類群のみが高く、Ruminococcus CAG-352、Clostridia UCG-014、Coprococcus eutactusが含まれていた。また、Coprococcus euctatusは、Coprococcus comesやRuminococcus gauvreauiiと同様に、耳炎を起こしたことのない乳児で高かった。Bacteroides eggerthiiとASV-81 Bifidobacterium breveは胃腸炎を起こしたことのない乳児で高かった。少なくとも5ヵ月以上母乳で育てられた乳児では、LacticaseibacillusとAnaeroglobusのメンバーが高かった。
図サムネイルgr7
図7Citrobacter属およびCoprococcus属の有病率およびPhascolarctobacterium属の存在量と早期耳炎感染および将来の神経発達障害(ND)の転帰との関連性
キャプション
大きな画像を見るダウンロード 高解像度画像
いくつかの分類群はリスク因子と関連していた。帝王切開による分娩児に多かったのは、Megamonas funiformis、Parabacteroides merdae、Prevotella 9、Sutterella、ASV-47、-77、-155 Bacteroides、ASV-86 Enterobacterであった。ASV-77 Bacteroides sp.は心理社会的脆弱性スコアが最も高い乳児に多く、ASV-25 Bacteroides fragilisは母親が妊娠中に喫煙した乳児に多かった。Anaerostipes caccae、Oscillospiraceae UCG-002 spp.、およびClostridium sensu stricto 1 neonataleは、1年目の食事に大きく左右されるようであり、特にフライドポテト/フライドポテトやチップスの頻繁な摂取と負の関連があった(FC = 4.0-5.0)。Anaerostipes caccaeとSutterella wadsworthensisは、最も短い期間(1~4ヵ月)母乳のみで育てられた乳児に多くみられた。
HLA遺伝子型
コントロールに関連する細菌は、DR4-DQ8またはDR3-DQ2を持たない人でより豊富であった。DR3-DQ2/DR4-DQ8ヘテロ接合体では、AcidaminocaccaeaeとChristensenellaceaeが多かった。詳細な種レベルの解析(図5E)では有意差が認められ、注目すべき種として、Adlercreutzia equolifaciens(FC = 4.2)、Phascolarctobacterium faecium(FC = 2.4)、Blautia obeum(FC = 2.0)、Coprococcus comes(FC = 1.0)が挙げられた。同様に、DR4-DQ8ホモ接合体では、Roseburia spp.、Phascolarctobacterium faecium、Coprococcus eutactusおよびCoprococcus comes、Alistipes putredinis、Alistipes finegoldii、Adlercreutzia equolifaciensなど、コントロールに関連する細菌の存在量が低いことが観察された(図5F)。
質問票データとバイオマーカーデータの統合により、細菌有病率の有意差は抗生物質の使用や化学物質曝露とも関連していた。
2つのKlebsiella michiganensis株(ASVs-120および-318)は、ABISmultiple株(48.3%)およびABISASD株(43.6%)では対照株(21.7%)より多く、特に抗生物質を頻繁に使用する対照株(27.1%)に多かった。ASVのいずれかを有する乳児は、ASDまたは多発性NDを発症する確率が高く(OR = 2.8 [95% CI, 1.46-5.30], p = 0.0019、およびOR = 3.4 [95% CI, 1.71-6.44], p = 0.0004、それぞれ)、これは、少なくとも8ヵ月間母乳で育てられ、抗生物質の使用が最小限(なしまたは1~2レジメン)であった乳児と比較した場合に増加した。ASVのいずれかが存在することは、ASD/ADHDを併発するリスクが3.60倍(1.60-8.08、95%CI)高いことと関連しており、抗生物質(OR = 5.52 [95%CI, 1.74-17.54])またはニコチン/アルコール(OR = 5.69 [95%CI, 2.20-14.71])に暴露されることでさらに高まった。両方のASVの存在は、ASD/ADHDの併存リスクを増加させた(ABISASD 23.1%、ABISmultiple 24.1%、対照8.3%)。サルモネラ菌に関連した腸内細菌は、将来のNDにおいて顕著に高かった(ASD/ADHD併存21%対対照3%、p = 1.07e-7;図5G)。アッカーマンシアはコントロールの48.7%にみられたが、将来ASD/ADHDを合併する25.0%(OR = 2.84, 1.12-7.20 95%CI、p = 0.027)と将来ASDを合併する28.2%(OR = 2.41, 1.19-4.88 95%CI、p = 0.014)にのみみられた。これらの相関は、親の学歴と妊娠中の母親の喫煙を考慮したロジスティック回帰でも維持された(調整後のORはそれぞれ3.2と0.55)。環境暴露と遺伝的要因はさらにリスクを増加させた。
早期耳炎とND転帰との関連を検討したところ、Citrobacter、Coprococcus、Phascolarctobacteriumは、多重発見補正後でも顕著な格差を示した。Citrobacterが検出された乳児は、耳炎のない対照と比較して、早期耳炎を伴うABISNDである可能性が2.35倍(1.12-4.94、95%CI)高かった(p = 0.0235;図7A)。逆に、コプロコッカスは耳炎を有する将来のNDでは10%しか存在しなかったが、耳炎を有さない対照では34%に存在した(図7A)。さらに、コプロコッカスはABISND全体ではわずか15.7%で観察され、これらの小児の78.6%は1年目に耳炎を経験していた。分娩様式、出生前の抗生物質および喫煙曝露、母乳育児などの変数で調整しても、これらの関連は確認された(図7Aおよび7C-7E)。
同様に、シトロバクターに感染した乳児は、耳炎のない対照群と比較して、早期耳炎を伴うABISNDである可能性が3.99倍(1.66-9.61、95%CI)高かった(p = 0.002;図7A)。将来のND群では、過去に耳炎を起こした人の52%がシトロバクターに感染していたのに対し、感染していない人では31%に過ぎなかった(OR = 2.71 [1.08-6.80]、p = 0.0336)。対照群では、1年目の耳炎はコプロコッカスの有病率が低かった(17%対28%)。興味深いことに、シトロバクターは1年目に耳炎を起こしたABISNDの61%に認められたが、ABISControlでは28%~34%にしか認められなかった(図7E)。さらに、Phascolarctobacterium(図7D)とCoprococcus(図7F)の存在量は、耳炎がない人の方が有意に高かった(それぞれμmean = 0.60対μmean = 0.10、μmean = 0.34対μmean = 0.10)。
バイオマーカーのメタボロームデータとマイクロバイオームデータの統合により、出生時のTGおよび極性代謝物、ならびに乳児の便中のビタミンおよび神経伝達物質前駆体との有意な関連が明らかになった。
臍帯血清
微生物と臍帯血清メタボロームとの関連を明らかにするため、臍帯血清とマイクロバイオームデータを持つ全サンプル(n = 114)の相関解析を用いて、極性代謝産物およびNDと最も一貫して関連する分類群の相対量を調べた。ASV-81 Bifidobacterium breveはLA、オレイン酸、UDCAと相関し、ASV-17 Anaerostipes hadrusは3-カルボキシ-4-メチル-5-プロピル-2-フランプロパノエートと相関した。PFDAとゼアラロンは、将来のASDの臍帯血清に多く含まれ、それぞれシトロバクター(p = 0.007)とASV-86エンテロバクター(p = 0.016)と正の相関を示した。Lacticaseibacillusは対照群で高濃度であった15種類のTGのうち11種類と、ASV-81 Bifidobacterium breveは4種類と相関していた。TG(46:2)/(16:1/14:0/16:1)レベルはASV-31ビフィドバクテリウム・ブレーベと正の相関を示したが、ASV-120およびASV-318エンテロバクター属細菌とも相関を示し、これらは将来のASDで高値を示した。Roseburia inulinivoransは4つのTGと負の相関を示した。
乳児の便のメタボローム
将来ASDになる乳児とならない乳児の便のメタボロームプロファイルも同様に解析した。AkkermansiaおよびASV-88 Akkermansia muciniphilaの16S rRNA総コピーは、グルタル酸、アラビノース、ピコリン酸、L-フェニルアラニン、L-イソロイシン、L-セリン、ロイシン、およびセロトニンとカテコールアミンの前駆体であるD-トリプトファンとL-チロシンを含む便の代謝物と正の相関があった(図6Dおよび6E)。コプロコッカスはリボフラビン、キサンチン、ウラシル、アラビノース、チミン、4-ヒドロキシ安息香酸塩と正の相関を示し、3-イソプロピルマレートとデヒドロアスコルビン酸塩と負の相関を示した。
便から得られたメタボロームと微生物の違いを関連付けるために、機械学習を用いて、今回の調査で最も顕著だった5つの属の細菌量を予測する負イオンモードで上位20代謝物を抽出し、相対存在量との相関を決定した(Bifidobacterium、Roseburia、Faecalibacterium、Akkermansia、Coprococcus、図6G-6K)。ビフィドバクテリウムには、2-デオキシ-D-ガラクトース・フルクトース、3,4-ヒドロキシ-フェニル乳酸、3-ヒドロキシフェニル酢酸、N-アセチル-D-ガラクトサミン、L-アラビトール、2-デオキシ-D-グルコース、6-デオキシ-L-ガラクトースなど多くの正の相関が観察され、キサノトシンとモノ-エチル-マロネートは負の相関を示した。L-アルギニンはRoseburiaと正の相関を示し、マロン酸は負の相関を示した。シキメイトはFaecalibacteriumおよびCoprococcusと負の相関を示した。また、Faecalibacteriumはウリジンと正の相関があり、3-suflino-L-alanine、タウリン、5-hydroxyindoleacetate、クレアチン、L-カルニチンと負の相関があった。Akkermansiaは3,4-ジヒドロキシ-フェニル-プロピオン酸と正の相関を示した。
考察
我々の研究では、特定の常在細菌とNDの進行、早期の消化器症状、気分の問題、および自己免疫疾患に関連するHLA対立遺伝子との逆相関が観察され、神経発達、腸管バリア機能、および免疫系との間に強い関連があることが示唆された。これらの知見を総合すると、腸内細菌が介在する炎症段階が、人生のごく初期にNDリスクに関与している可能性が示唆される。
出生時と1歳時に観察された微生物とメタボロームの不均衡は、いくつかの重要な意味を持つ。第一に、臍帯血清のメタボロームの違いは、免疫刺激イベントや心理社会的ストレス因子をコントロールした後も持続することから、外的影響というよりもむしろ、出生時に固有の生物学的変動があることが明らかになった。注目すべきは、将来ASDになる新生児において、LAやα-リノレン酸(ALA)、胆汁酸、TGのような重要な脂質が減少していることで、出生時に炎症性イベントが存在することを示唆している。LAとALAはドコサヘキサエン酸(DHA、22:6n-3)のような長鎖多価不飽和脂肪酸(PUFA)の前駆体であり、脳に対して抗炎症作用を持ち50,51、オートファジー、神経伝達、神経新生を調節する52。エンドカンナビノイド系の調節は、γ-アミノ酪酸(GABA)などの神経伝達物質放出の抑制を通じて、シナプス機能と可塑性に影響を与える53。私たちは、将来ASDになる乳児の臍帯血清TG枯渇を観察し、これらのレベルは1歳時のビフィズス菌量と相関していることがわかった。55,56胆汁酸に関しては、UDCAは将来のASDでも低値であるが、代謝性疾患、自己免疫疾患、慢性炎症性疾患、神経病理学にまたがる病態において治療効果が期待されている57,58,59,60。その血液脳関門を通過する能力は注目に値する61。抗炎症および抗アポトーシス機序は、おそらくドーパミンやミトコンドリアの調節に関連している58。さらに、環境汚染物質PFDAは将来のASDで有意に高かった。PFDAを含むペルフルオロアルキル物質やポリフルオロアルキル物質は、免疫反応に大きな影響を及ぼす62。これらの物質は、慢性炎症、酸化ストレス、免疫抑制63に関連し、自己免疫疾患に関与する可能性があることが知られている64。
第二に、腸内細菌異常症の発症は、ごく早期(乳児期)に起こり、必須代謝産物(ビタミン、脂肪酸、神経伝達物質前駆体など)の破壊と有意な相関がある。急性、慢性の消化管炎症と腸管透過性の亢進はASD65,66と一親等の近親者で観察されているが、65、ABISでは診断の数年前から微生物の違いが明らかになり、抗炎症微生物と腸管バリアの強化因子の枯渇が特徴的であった。最近の研究では、アッカーマンシア・ムチニフィラが腸の健康を促進する顕著な可能性が示されているが67、神経発達については研究されていない。アッカーマンシア(Akkermansia muciniphila)は、後にASDまたはASD/ADHD併存と診断されたABIS乳児には認められず、幼児期の消化器症状および気分症状と逆相関していた。興味深いことに、アッカーマンシアは将来のADHDとは関連しておらず、ムチンの健康状態の乱れがASDとより強固な関係があることを示唆している。微生物由来の3,4-ジヒドロキシ-フェニル-プロピオン酸は、インターロイキン-6(IL-6)サイトカイン産生をダウンレギュレートするエピジェネティック修飾因子である73。ビフィズス菌もまた、将来のNDでは、条件を問わず減少していた。ビフィズス菌は健康的な免疫反応を促進し、フェニルアラニンを増加させることによってドーパミンの産生を促進する32。便中のビフィズス菌の量は、認知機能に関連するカテコールアミン前駆体であるチロシンの代謝物である4-ヒドロキシフェニル乳酸と相関していた74,75。コプロコッカス(Coprococcus)には強力な抗炎症作用があり、ABISではNDsと逆相関し、脆弱性スコアの低さ、抗生物質の少なさ(最初の1年間はなし)、間食の少ない幼児食などの保護因子と関連していた。Roseburiaはまた、強力な抗炎症作用を有し、大腸運動や免疫系に関与している77。Turicibacter sanguinisは、セロトニン利用78やステロイドおよび脂質代謝経路に関与している。
いくつかの主要な酪酸産生菌69は将来のNDと逆相関を示したが(Faecalibacterium prausnitzii、76,79 Roseburia、69,80 Anaerostipes、80 Acidaminococcales)、便中の酪酸濃度に顕著な差は認められなかった(p = 0.38)。しかし、酪酸の回転率が対照群と未来型群で異なるかどうかは不明である。クエン酸レベルは将来のASD症例で高く(p = 0.038)、尿中のコハク酸およびクエン酸の増加を認めたASD症例の小児を対象とした横断的研究81と一致している一方、便中のウリジンレベルはFaecalibacteriumと正の相関があった。ウリジンによる治療は、代謝適応による組織の再生と修復を促進し、ミトコンドリア活性を改善する82。
SlackiaやAdlercreutzia equolifaciensなど、いくつかの既知のエクオール産生菌は、対照群で一貫して高かった。脂質代謝85とエクオール産生に関与するコリオバクテリウム科の細菌もまた、将来のNDの乳児では減少していた。EggerthellaとSlackiaは、宿主の脂質と異種物質代謝にプラスの影響を与える86。将来のASD症例では、潜在的なエクオールシグナルの有意な減少が観察された。エクオールは抗炎症作用やエストロゲン活性について研究されてきた。前臨床モデルでは、エクオールは血液脳関門透過性能力87、抗神経炎症特性87、神経保護効果を示し、リポ多糖(LPS)誘発炎症87や神経毒からミクログリアを保護している88。
第三に、乳児の食事、心理社会的脆弱性(妊娠中の母親の喫煙など)、抗生物質の使用などの環境因子を調整した後でも、我々が観察したディスバイオーシスは持続する。母乳から固形食への移行期には食生活が変化する可能性があるが、微生物DNAは1歳児由来のものであり、食生活の違いに照らしても所見が持続したことから、食生活だけで観察された大きな違いを説明できる可能性は低い。このように、今回の研究結果は、NDと診断された小児を研究する文献における重要なギャップを解決するものである89。これらの研究結果は、微生物異常が診断後の食事変化のみによるものではなく、むしろ診断前から存在していたことを示しており、これらの疾患の早期発症に関する貴重な知見を提供するものである。多くの場合、乳児期のマイクロバイオームの違いは、2.5〜5歳における早期の気分や消化器症状も示唆していた。調整アプローチと非調整アプローチの両方で、対照(例えば、Coprococcus eutactus、Akkermansia muciniphila、Blautia obeum)と症例(例えば、Corynebacterium variabile)でいくつかの細菌が濃縮されていることが示された。コリネバクテリウム属細菌は一般に病原性細菌と考えられており、バンコマイシンまたはアミノグリコシドにのみ感受性を示すものもある90。ある種のビフィドバクテリウム・ブレーベ(Bifidobacterium breve)ASV、ならびにルミノコッカス科CAG-352、アドレルクロイツィア・エクオリファシエンス(Adlercreutzia equolifaciens)、およびローズブリア属細菌は、調整後に有意性を失ったことから、NDにおけるこれらの微生物の役割は環境要因に大きく介在していることが示唆される。
第四に、ABISの乳児にみられる微生物異常症の程度は、HLA、ストレス、小児期の感染症/抗生物質の反復の影響と関連している可能性がある。免疫賦活事象と心理社会的ストレス因子は、胎児の健康、発育、免疫に永続的な影響を及ぼし、前者は細胞の分化、移動、シナプスの成熟を阻害する。前臨床モデルでは、ストレスによって誘発される出生前の出来事は、炎症性サイトカインIL-6を介するGABA作動性遅延を活性化し、成体ミクログリアに影響を与える。帝王切開は言語障害と知的障害のリスクであった。ABISには分娩中の抗生物質に関するデータがないが、すべての帝王切開による出産(ABISにおける出産の11.9%)には抗生物質も含まれていたと思われる。シナプス機能、中枢神経系の発達、神経疾患に対するHLAの寄与は、次第に明らかにされつつある: DR4-DQ8ホモ接合体(CDやT1Dを含む自己免疫疾患と関連している)は、NDとASDのリスクを1.8倍と2.8倍増加させた。DR3-DQ2/DR4-DQ8ヘテロ接合体またはDR4-DQ8ホモ接合体の小児は、Roseburia属、Phascolarctobacterium faecium属、Coprococcus属の欠損を共有しており、将来NDになる小児も同様であった。
ABISでは、1年目に耳炎を起こした乳児は、コプロコッカスが検出されないか、シトロバクターを保有している場合、NDを発症しやすいことがわかった。対照群では耳炎に関係なく同レベルであったにもかかわらず、コプロコッカスが検出されなかったことから、微生物群集の回復について疑問が呈された。このような事象の後にマイクロバイオームが回復しない可能性は、中耳炎とNDリスクを結びつけるメカニズムとして機能するかもしれない。さらに、これらの乳児では抗生物質耐性のシトロバクター101がより一般的であった。サルモネラやシトロバクターに関連する菌株(この調査ではSREBと表示)の存在は、対照群(3%)と比較して、後にASD/ADHDを併発した乳児(21%)で有意に高かった。この混乱は、重要な時期の神経発達に影響を及ぼす可能性がある。サルモネラとシトロバクターは、ウィングレス(Wnt)シグナル伝達をアップレギュレートする能力を示した。Wnt経路は免疫異常と脳の発達に不可欠であり、その破綻がASDの発症に関与している102,103,104,105。
最後に、初期の腸内細菌叢異常は、アミノ酸、脂肪酸、ビタミン、神経伝達物質前駆体など、便中のいくつかの代謝産物の破壊を示唆している。ABISでは、将来ASDになる可能性のある乳児において、準必須アミノ酸であるL-アルギニンと必須アミノ酸であるリジンの有意な減少が観察され、すでに診断されている小児におけるアミノ酸の乱れと一致していた106,107。108。リジンとアルギニンはともに、成長代謝、免疫機能、ヒストン修飾、一酸化窒素の産生に重要である。リジンは脂肪酸代謝、カルシウム吸収、免疫機能、タンパク質沈着に関与しており、脾臓組織での代謝率が高い。110 リジンの分解経路は、重要な神経伝達物質の前駆体であるトリプトファンの分解経路と密接に関連している。アルギニンの欠乏は、特に細菌チャレンジの際に、新生児が適切な免疫応答を生成する能力を大幅に阻害し、それによって感染症、特に消化管に由来する感染症に対する感受性を上昇させる。
将来のASDの便では、対照群と比較して2つの脂肪酸の違いが顕著であった:オメガ7モノ不飽和パルミトレイン酸、(9Z)-ヘキサデック-9-エン酸(将来のASDの87.0%では検出レベル以下であったが、対照群では43.5%に存在)、パルミチン酸(将来のASDでは上昇)。パルミトレイン酸は、膵島および原発性インスリン自己免疫のリスク低下と関連している116。逆に、飽和脂肪酸であるパルミチン酸は、神経細胞のホメオスタシスの障害と関連している117。
将来ASDになる可能性のある乳児の便で高値を示した代謝産物はほとんどなかったが、注目すべき例がいくつかある: α-d-グルコース、ピルビン酸、3-イソプロピルマレートなどである。コプロコッカスは3-イソプロピルマレートと逆相関しており、腸と脳のつながりを示唆している118。また、ロイシンとイソロイシンの生合成における3-イソプロピルマレートデヒドロゲナーゼの役割を考えると、分岐鎖アミノ酸(BCAA)経路のバランスが崩れている可能性がある119。デヒドロアスコルビン酸の増加は、神経伝達物質の合成や抗酸化防御に重要なビタミンC代謝の乱れの可能性を示唆し、ピルビン酸の増加は、神経伝達物質の合成やエネルギー産生の初期段階での乱れを示唆する。脂肪酸酸化障害でみられるピメリン酸の上昇は、脂肪酸酸化のためのミトコンドリア経路の障害を示唆している120。
アッカーマンシアとコプロコッカスは、将来NDを発症する乳児では欠乏または減少しており、便中の神経伝達物質前駆体および必須ビタミンを表す便中シグナルと正の相関がみられた。具体的には、Akkermansiaはチロシンおよびトリプトファン(それぞれカテコールアミンおよびセロトニンの前駆物質)と、Coprococcusはリボフラビンと相関していた。ASDにおけるBCAA代謝の障害は、大アミノ酸トランスポーター(LAT)のコード変異や、トリポタンファンや大芳香族アミノ酸の利用低下121、グルタミン酸の増加、チロシン、イソロイシン、フェニルアラニン、トリプトファンの減少に関与している121,122。酸化ストレス、効率的なエネルギー輸送能力の低下122、神経伝達物質の合成や神経細胞の維持に不可欠なビタミン(ビタミンB2など)の欠乏が関与している123。抗酸化物質であるリボフラビンは酸化ストレスや炎症を軽減し124、神経疾患における神経保護効果を示す125が、おそらくグルタミン酸からグルタミンへの変換や5-ヒドロキシトリプトファンからセロトニンへの変換に必要なビタミンB6の維持126を介しているのだろう。
これらの知見を総合すると、腸内細菌叢を介したNDの早期発症という仮説が支持される。このことは、研究およびNDに対する早期介入を開発するための基盤となる。
研究の限界
神経心理学的検査は実施されていないが、国際疾病分類(ICD)-10基準を用いた医師の評価に基づくグループ分けは強固である。参加者が1997年から1999年にスウェーデンで生まれた子供に限定されているため、一般化可能性は不明である。1歳時の微生物DNAはABISコホートの10.8%しか分析されておらず、代表性に影響を及ぼす可能性がある。変数をコントロールする努力にもかかわらず、未測定の交絡因子が結果に影響を与える可能性がある。ABISは広範なアンケートを通じて豊富なメタデータを提供しているが、分娩中に投与された抗生物質など、いくつかの重要なデータは入手できなかった。細菌種の株は生理学的特性において著しく異なることがある。炎症との関連におけるディスバイオージスの時間的動態、微生物の特徴の連続性、そしてディスバイオージスが引き金として作用するのか、あるいは結果として作用するのかは、依然として不明である。本研究は、最適な介入法を開発し、因果関係を理解するために重要な、幼児期におけるディスバイオーシスの持続性と機能的影響に関するさらなる研究の必要性を強調するものである。
STAR★方法
主要リソース表
試薬またはリソースソースの識別子
生物学的サンプル
ABIS参加者の臍帯血 All Babies in Southeast Sweden, Linköping, Sweden https://www.abis-studien.se/hem/english-11100423
1歳児ABIS参加者の便サンプル All Babies in Southeast Sweden, Linköping, Sweden https://www.abis-studien.se/hem/english-11100423
化学物質、ペプチド、組換えタンパク質
16S rRNA増幅プライマー(Triplett lab127でカスタマイズ) Invitrogen, USA 341F: NNNNCCTACGGGAGGCAGCAG; 806R: GGGGACTACVSGGGTATCTAAT
再懸濁バッファー イルミナ Cat#15026770
水酸化ナトリウム溶液、5N Fisher Scientific Cat#SS256-500
2-ジヘプタデカノイル-sn-グリセロ-3-ホスホエタノールアミン (PE(17:0/17:0)) Avanti Polar Lipids Cat#830756
1-ステアロイル-2-ヒドロキシ-sn-グリセロ-3-ホスホコリン (LPC(18:0)) Avanti Polar Lipids Cat#855775
N-ヘプタデカノイル-D-エリスロスフィンゴシルホスホリルコリン (SM(d18:1/17:0)) Avanti Polar Lipids Cat#121999-64-2
2-ジヘプタデカノイル-SN-グリセロ-3-ホスホコリン (PC(17:0/17:0)) Avanti Polar Lipids Cat#850360
1-ヘプタデカノイル-2-ヒドロキシ-sn-グリセロ-3-ホスホコリン (LPC(17:0)) Avanti Polar Lipids Cat#855676
2-ジオクタデカノイル-sn-グリセロ-3-ホスホコリン (PC(18:0/18:0)) Avanti Polar Lipids Cat#850333
1-ヘキサデカノイル-2-オレオイル-sn-グリセロ-3-ホスホコリン (PC(16:0/18:1)) Avanti Polar Lipids Cat#850457
1-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホエタノールアミン (LPE(18:1)) Avanti Polar Lipids Cat#850456
1-パルミトイル-2-ヒドロキシ-sn-グリセロ-3-ホスファチジルコリン (LPC(16:0)) Avanti Polar Lipids Cat#846725
トリヘプタデカノイルグリセロール (TG(17:0/17:0/17:0)) Larodan Cat#33-1700
トリヘキサデカノイルグリセロール (TG(16:0/16:0/16:0)) ラロダン Cat#33-1610
1-stearoyl-2-linoleoyl-sn-glycerol (DG(18:0/18:2)) Avanti Polar Lipids Cat#855675
3-トリオクタデカノイルグリセロール (TG(18:0/18:0/18:0)) Larodan Cat#33-1810
3β-ヒドロキシ-5-コレステン-3-リノール酸 (ChoE(18:2)) ラロダン Cat#64-1802
1-ヘキサデシル-2-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホコリン (PC(16:0e/18:1(9Z))) Avanti 極性脂質 Cat#800817
1-(1Z-オクタデカニル)-2-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホコリン (PC(18:0p/18:1(9Z)) Avanti 極性脂質 Cat#878112
1-オレオイル-2-ヒドロキシ-sn-グリセロ-3-ホスホコリン (LPC(18:1)) Larodan Cat#38-1801
1-パルミトイル-2-オレオイル-sn-グリセロ-3-ホスホエタノールアミン (PE(16:0/18:1)) Avanti Polar Lipids Cat#852467
3β-ヒドロキシ-5-コレステン-3-ステアリン酸 (ChoE(18:0)) Larodan Cat#64-1800
1-palmitoyl-d31-2-oleoyl-sn-glycero-3-phosphocholine (PC(16:0/d31/18:1)) Avanti Polar Lipids Cat#850757
2-ジヘプタデカノイル-sn-グリセロ-3-ホスホエタノールアミン (PE(17:0/17:0)) Avanti Polar Lipids Cat#830756
N-ヘプタデカノイル-D-エリスロスフィンゴシルホスホリルコリン (SM(d18:1/17:0)) Avanti Polar Lipids Cat#860585
1-stearoyl-2-hydroxy-sn-glycero-3-phosphocholine (LPC(18:0)) Avanti Polar Lipids Cat#855775
β-ムリコール酸ステラロイド Cat#C1895-000
チェノデオキシコール酸 Sigma-Aldrich Cat#C1050000
コール酸 Sigma-Aldrich Cat#C2158000
デオキシコール酸 Sigma-Aldrich Cat#700197P
グリコチェノデオキシコール酸 Sigma-Aldrich Cat#700266P
グリココール酸 Sigma-Aldrich Cat#700265P
グリコデヒドロコール酸 ステラロイド Cat#C2020-000
グリコデオキシコール酸 Sigma-Aldrich Cat#361311
グリコヒドロコール酸 ステラロイド Cat#C1860-000
グリコヨデオキシコール酸 ステラロイド Cat#C0867-000
グリコリトコール酸 Sigma-Aldrich Cat#700268P
グリコルデオキシコール酸 Sigma-Aldrich Cat#06863
ヒオコール酸ステラロイド Cat#C1850-000
ヒヨデオキシコール酸ステラロイド Cat#C0860-000
リトコール酸 Sigma-Aldrich Cat#700218P
α-ムリコール酸ステラロイド Cat#C1891-000
タウロ-α-ムリコール酸ステラロイド Cat#C1893-000
タウロ-β-ムリコール酸ステラロイド Cat#C1899-000
タウロヘノデオキシコール酸 Sigma-Aldrich Cat#700249P
タウロコール酸 Sigma-Aldrich Cat#T9034
タウロデオキシコール酸 シグマ・アルドリッチ Cat#700242P
タウロデオキシコール酸 シグマ・アルドリッチ Cat#700250P
タウロデオキシコール酸 シグマアルドリッチ Cat#700248P
タウロリトコール酸 Sigma-Aldrich Cat#700252P
タウロオメガムリコール酸 ステラロイド Cat#C1889-000
タウルソデオキシコール酸 Sigma-Aldrich Cat#580549
トリヒドロキシコレスタン酸 Avanti Polar Lipids Cat#700070P
フマル酸 Sigma-Aldrich Cat#47910
グルタミン酸 Sigma-Aldrich Cat#G0355000
アスパラギン酸 Sigma-Aldrich Cat#A1330000
セリン Sigma-Aldrich Cat#S4500
スレオニン シグマ・アルドリッチ Cat#PHR1242
グルタミン Sigma-Aldrich Cat#G3126
プロリン シグマ・アルドリッチ Cat#V0500
バリン シグマ・アルドリッチ Cat#PHR1172
リジン Sigma-Aldrich Cat#L5501
メチオニン シグマ・アルドリッチ Cat#M0960000
シリング酸 Sigma-Aldrich Cat#63627
イソロイシン シグマ・アルドリッチ Cat#I2752
ロイシン シグマ・アルドリッチ Cat#L8000
リンゴ酸 Sigma-Aldrich Cat#PHR1273
フェニルアラニン Sigma-Aldrich Cat#P2126
フェルラ酸 Sigma-Aldrich Cat#Y0001013
クエン酸 Sigma-Aldrich Cat#C7129
トリプトファン Sigma-Aldrich Cat#93659
3-インドール酢酸 Sigma-Aldrich Cat#I3750
3-ヒドロキシ酪酸 Sigma-Aldrich Cat#52017
イソ吉草酸 Sigma-Aldrich Cat#78651
インドール-3-プロピオン酸 Sigma-Aldrich Cat#57400
サリチル酸 Sigma-Aldrich Cat#247588
イソカプロン酸 Sigma-Aldrich Cat#277827
デカン酸 Sigma-Aldrich Cat#C1875
ミリスチン酸 Sigma-Aldrich Cat#70079
リノレン酸 Sigma-Aldrich Cat#62160
パルミトレイン酸 Sigma-Aldrich Cat#76169
リノール酸 Sigma-Aldrich Cat#62230
エイコサペンタエン酸 Sigma-Aldrich Cat#44864
パルミチン酸 Sigma-Aldrich Cat#P0500
オレイン酸 Sigma-Aldrich Cat#75090
ステアリン酸 Sigma-Aldrich Cat#S4751
アラキジン酸 Sigma-Aldrich Cat#39383
[D4]-グリコソデオキシコール酸 Bionordica Cat#31309
[D4]-グリココール酸 Bionordica Cat#21889
[D4]-ウルソデオキシコール酸 Bionordica Cat#21892
[D4]-グリコヘノデオキシコール酸 ビオノーディカ Cat#21890
[D4]-コール酸 ビオノーディカ Cat#20849
[D4]-グリコリトコール酸 Bionordica Cat#31308
[D4]-チェノデオキシコール酸 Bionordica Cat#20848
[D4]-デオキシコール酸 ビオノーディカ Cat#20851
[D4]- リトコール酸 Bionordica Cat#20831
バリン-d8 Sigma-Aldrich Cat#486027
グルタミン酸-d5 Sigma-Aldrich Cat#631973
コハク酸-d4 Sigma-Aldrich Cat#293075
ヘプタデカン酸 Sigma-Aldrich Cat#H3500
乳酸-d3 Sigma-Aldrich Cat#616567
クエン酸-d4 Sigma-Aldrich Cat#485438
アルギニン-d7 Sigma-Aldrich Cat#776408
トリプトファン-d5 Sigma-Aldrich Cat#615862
グルタミン-d5 Sigma-Aldrich Cat#616303
重要な市販アッセイ
QuantStudio 3 システム用 qPCR アッセイ Applied Biosystems, Life Technologies, USA Cat#A28567
qPCR 用反応混合物(PowerUp SYBR Green 2X Master Mix) Applied Biosystems, Life Technologies, USA Cat#25742
E.Z.N.A 便 DNA キット Omega Bio-Tek、USA Cat#D4015-02
E.Z.N.A Cycle-Pure Kit Omega Bio-Tek、USA Cat#D6492-02
Nextera XT DNAライブラリー調製キット イルミナ Cat#FC-131-1096
MiSeq Reagent Kit v3(600サイクル) イルミナ Cat#MS-102-3003
PhiXコントロールv3 イルミナ Cat#FC-110-3001
寄託データ
処理済みデータ 本論文 Dryad: https://doi.org/10.5061/dryad.ghx3ffbwj
新しいタブで表を開く
リソースの有無
連絡先
詳細情報およびリソースのリクエストは、リードコンタクトであるProf. Eric Triplett (ewt@ufl.edu)までお願いします。
資料の利用可能性
本研究では、新規の試薬は使用していない。
データおよびコードの利用可能性
本論文で報告された処理されたデータは、"All Babies in Southeast Sweden - Neurodevelopment and Early Life Factors "データセット(Dryad: https://doi.org/10.5061/dryad.ghx3ffbwj)としてDryadに寄託され、発表日現在公開されている。キュレーションされたデータは、子どもの参加者に必要な、倫理的な被験者保護に従って選択された。
本論文はオリジナルのコードを報告していない。
本論文で報告されたデータを再分析するために必要な追加情報は、要求があれば、主担当者から入手可能である。
実験モデルと研究参加者の詳細
ヒト参加者
ABISは、1997年10月1日から1999年10月1日の間にスウェーデン南東部で生まれたすべての子供の親を対象とした、前向き一般集団ベースのコホート研究である128。ABISの当初の動機は、免疫介在性疾患の病因と環境要因および遺伝要因の影響を前向きに研究することであった。子供たちは出生時から追跡されている。参加した親は、出生時、1歳、2-3歳、5歳、8-10歳、14-16歳、17-19歳の時点でアンケートに答え、生後1年間は日記をつけた129。データには、家族の病歴、抗生物質の使用、投薬、食事、ライフスタイル、気分と体質、家庭環境、環境暴露、心理社会的脆弱性などが含まれるが、これらに限定されるものではない。
今回の調査では、1歳、3歳、5歳のABIS児の両親から得た最も古い質問票を分析した。この調査で考慮した最新の診断は、子どもたちが21~23歳のときに得られたものである。ABISコホートの生物学的性別は比較的バランスのとれた分布であった(女性48.2%、男性51.8%)。人種、祖先、民族に関するデータは収集されていないが、ABISの子供の89.2%が両親ともスウェーデン生まれ、7.7%が片方の親がスウェーデン以外の国で生まれ、両親ともスウェーデン以外の国で生まれたのはわずか3.1%であることから、同質性があると予想される。1990年代後半には、人口の大半をスウェーデン民族が占め、次いでフィンランド人が多かった。
社会経済面では、ABISの子どもの6.6%が最大の心理社会的脆弱性(脆弱性指数>2)を示した。2000年、2006年、2012年のABIS家族の可処分世帯収入は、それぞれ305,360.7 SEK(95%信頼区間:297,279-313,462.5)、411,253.2 SEK(408,306.9-414,199.5)、554,889.7 SEK(549,889.1-559,890.3)と報告された。
診断における生物学的性別の影響、特に男児における影響を考慮し、性別と診断年齢(ASDのみ)で層別化したマイクロバイオームとの関連を検討した。この層別化後のサンプルサイズは限られているため注意が必要である。コホート全体のリスク因子に対する生物学的性別の複合的な影響は明確に考慮されておらず、一般化可能性の制限となる可能性がある。設計上、ABISコホートはスウェーデン出身者で構成されているため、この知見はより多様な、あるいはヨーロッパ以外の集団には直接適用できない可能性があることに注意することが重要である。これらの知見の外部妥当性を高めるためには、さまざまな人口統計を持つ集団におけるさらなる研究が必要である。
診断
国際疾病分類の診断(ICD-10)はNational Patient Register(NPR)130から得たもので、2020年12月までの診断が収録されている。精神疾患診断は、診断日によりDSM-IV131(Diagnostic and Statistical Manual of Mental Disorders, fourth edition)またはDSM-V132(Diagnostic and Statistical Manual of Mental Disorders, fifth edition)に従って設定された(図1B)。対象となる診断コードには、ASD(F84.0、F84.1、F84.2、F84.3、F84.4、F84.5、F84.8、F84.9)、ADHD(F90.0、F90.0A、F90.0B、F90. 0C、F90.0X)、言語障害(F80.0、F80.0A、F80.0B、F80.0C、F80.1、F80.1A、F80.1B、F80.1C、F80.2、F80.2A、F80.2B、F80.2C、F80. 3、F80.8、F80.8A、F80.8B、F80.8C、F80.8D、F80.8W、F80.9)、知的障害(F70.0、F70.1、F70.8、F70.9、F71.0、F71.1、F71. 8、F71.9、F72.0、F72.1、F72.8、F72.9、F73.0、F73.1、F73.8、F73.9、F78.0、F78.1、F78.8、F78.9、F79.0、F79.1、F79.8、F79.9、F83.9)。
倫理承認と参加同意
すべての家族は、本研究に参加することにインフォームド・コンセントを提供した。ABIS研究は、リンシェーピン大学およびルンド大学の研究倫理委員会(承認番号Dnr 03-092、LiU 287.96、LU 83-97、LiU 321-99.およびリンシェーピン大学Dnr 36287、Dnr 03-513、Dnr 2018/380-32)により承認された。本研究で使用された非識別化生物試料の解析は、フロリダ大学の施設審査委員会(IRB)の承認の下で実施された。本研究はIRB201800903およびIRB202301239のもとで承認され、参加者へのリスクが最小であるという基準を満たしていることが確認された。
方法の詳細
臍帯血清代謝物の取得
臍帯血清サンプルは母親からの提供で入手し、乳児が1歳時に便も提供した母親から合計120サンプルを得た。分析にはリピドミクスと親水性(水溶性)代謝物プロファイリングの2つの方法を用いた。後者では遊離脂肪酸、胆汁酸、アミノ酸を測定した。 グリコチェンデオキシコール酸、グリココール酸、グリコヒドロコール酸、グリコデオキシコール酸、グリコヒオコール酸、グリコヒオデオキシコール酸、グリコリトコール酸、グリコルデオキシコール酸、ヒオコール酸 ヒオデオキシコール酸、リトコール酸、オメガ/アルファ-ムリコール酸、パーフルオロオクタン酸、タウロ-アルファ-ムリコール酸、タウロ-ベータ-ムリコール酸、タウロヘノデオキシコール酸、タウロコール酸、 タウロデヒドロコール酸、タウロデオキシコール酸、タウロヒオデオキシコール酸、タウロリトコール酸、タウロオメガムリコール酸、タウロルソデオキシコール酸、トリヒドロキシコレスタン酸、ウルソデオキシコール酸。遊離脂肪酸には、デカン酸、ミリスチン酸、リノレン酸、パルミトレイン酸、リノール酸、エイコサペンタエン酸、パルミチン酸、オレイン酸、ステアリン酸、アラキジン酸が含まれた。極性代謝物としては、フマル酸、グルタミン酸、アスパラギン酸、リンゴ酸、フェニルアラニン、フェルラ酸、クエン酸、トリプトファン、3-インドール酢酸、3-ヒドロキシ酪酸、イソ吉草酸 インドール-3-プロピオン酸、サリチル酸、イソカプロン酸、コハク酸、セリン、スレオニン、グルタミン、プロリン、バリン、リジン、メチオニン、シリンギン酸、イソロイシン/ロイシン。
臍帯血清のリピドミクス解析
合計360の臍帯血清サンプルをランダム化し、以下に記述するように分析した。10μlの血清を10μlの0.9%NaClと混合し、120μlのCHCl3で抽出した: 内部標準混合物を含むMeOH(2:1、v/v)溶媒混合物(c=2. 5 μg/ml;1,2-ジヘプタデカノイル-sn-グリセロ-3-ホスホエタノールアミン(PE(17:0/17:0))、N-ヘプタデカノイル-D-エリスロスフィンゴシルホスホリルコリン(SM(d18:1/17:0))、N-ヘプタデカノイル-D-エリスロスフィンゴシン(Cer(d18:1/17: 0)、1,2-ジヘプタデカノイル-sn-グリセロ-3-ホスホコリン(PC(17:0/17:0))、1-ヘプタデカノイル-2-ヒドロキシ-sn-グリセロ-3-ホスホコリン(LPC(17: 0)、1-パルミトイル-d31-2-オレオイル-sn-グリセロ-3-ホスホコリン(PC(16:0/d31/18:1))およびトリヘプタデカノイルグリセロール(TG(17:0/17:0/17:0))。サンプルをボルテックスし、氷上で30分間放置した後、遠心分離した(9400rcf、3分間)。60μlの下層を集め、60μlのCHCl3: MeOHで希釈した。サンプルは分析まで-80℃で保存した。
サンプルは、超高性能液体クロマトグラフィー四重極飛行時間型質量分析計(Agilent Technologies社製UHPLC-QTOFMS;米国カリフォルニア州サンタクララ)を用いて分析した。分析は、Waters(米国ミルフォード)のACQUITY UPLC BEH C18カラム(2.1 mm×100 mm、粒子径1.7 μm)を用いて行った。品質管理は、ブランク、純粋な標準試料、抽出標準試料、コントロール血漿試料を含むデータセット全体で実施した。溶離液システムは、(A) 10 mM NH4Ac in H2Oおよび0.1%ギ酸、(B) 10 mM NH4Ac in ACN: IPA (1:1)および0.1%ギ酸で構成された。グラジエントは以下の通りであった: 0-2分、35%溶媒B; 2-7分、80%溶媒B; 7-14分、100%溶媒B。
脂質の定量は、7点内部検量線(0. 1-5μg/mL)を用い、以下の脂質クラス特異的な真正標準物質を用いた:1-ヘキサデシル-2-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホコリン(PC(16:0e/18: 1(9Z))、1-(1Z-オクタデセニル)-2-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホコリン(PC(18:0p/18:1(9Z))、1-ステアロイル-2-ヒドロキシ-sn-グリセロ-3-ホスホコリン(LPC(18: 0)、1-オレオイル-2-ヒドロキシ-sn-グリセロ-3-ホスホコリン(LPC(18:1))、1-パルミトイル-2-オレオイル-sn-グリセロ-3-ホスホエタノールアミン(PE(16:0/18: 1)、1-(1Z-オクタデセニル)-2-ドコサヘキサエノイル-sn-グリセロ-3-ホスホコリン(PC(18:0p/22:6))および1-ステアロイル-2-リノレイル-sn-グリセロール(DG(18:0/18:2))、1-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホエタノールアミン(LPE(18: 1))、N-(9Z-オクタデセノイル)-スフィンガニン(Cer(d18:0/18:1(9Z))、1-ヘキサデシル-2-(9Z-オクタデセノイル)-sn-グリセロ-3-ホスホエタノールアミン(PE(16:0/18: Avanti Polar Lipids製)、1-パルミトイル-2-ヒドロキシ-sn-グリセロ-3-ホスファチジルコリン(LPC(16:0))、1,2,3-トリヘキサデカノアルグリセロール(TG(16:0/16:0/16:0))、1,2,3-トリオクタデカノイルグリセロール(TG(18: 0/18:0/18:0))および3β-ヒドロキシ-5-コレステン-3-ステアリン酸(ChoE(18:0))、3β-ヒドロキシ-5-コレステン-3-リノレイン酸(ChoE(18:2))をLarodanから入手し、各内部標準1250ng/mLを含む以下の濃度レベル:100、500、1000、1500、2000および2500ng/mL(CHCl3:MeOH、2:1、v/v中)に調製した。
国際脂質分類命名委員会(International Lipid Classification and Nomenclature Committee: ILCNC)は、脂質の包括的な分類フレームワークであるLIPID MAPS 133を導入した。この化学的な分類システムは、脂質を脂肪アシル、グリセロ脂質(GL)、グリセロリン脂質(GP)、スフィンゴ脂質(SP)、糖脂質(SL)、ポリケチド(PK)、プレノール脂質(PR)、ステロール脂質(ST)の8つのクラスに分類している。分類は脂質クラス、脂肪酸組成、炭素数、二重結合含有量に基づいている。PCs、Pes、Pis、SMs、セラミドなどの特定の脂質種は、そのヘッドグループに結合した2つの脂肪アシル基を持つが、lysoPCsとlysoPEsは1つの脂肪アシル基を持つという特徴がある。CE、DG、TGなどのクラスは、脂肪アシル基の数がそれぞれ1個、2個、3個と異なるのが特徴である。さらに脂質は、ホスファチジルコリン(PC)、リゾホスファチジルコリン(lysoPC)、ホスファチジルエタノールアミン(PE)、ジ-およびトリアシルグリセロール(DG、 TG)、スフィンゴミエリン(SM)、セラミド(Cer)、ホスファチジルイノシトール(PI)、ホスファチジルグリセロール(PG)、モノヘキソシルセラミド(HexCer)、ラクトシルセラミド(LacCer)。TGの分類は、脂肪酸組成に基づき、飽和、一価不飽和、多価不飽和に区別される。特に、PCとPEは、アルキルエーテルPCまたはPE(プラズマローゲン)を含むサブカテゴリーを示し、それぞれがわずかに異なる構造と生物学的機能を有する。詳細な構造が決定されていない場合、命名は炭素と二重結合の合計に基づいている。
データはMZmine 2.53.134を使用して処理した。同定はカスタムデータベースを使用し、同定レベル1と2を設定した。すなわち、本物の標準化合物(レベル1)に基づき、Metabolomics Standards Initiativeに基づくMS/MS同定(レベル2)に基づく。品質管理は、プールされた品質管理サンプル(各サンプルからプールされたアリコート)をサンプルと共に分析することによって行われた。さらに、品質管理手順の一環として、基準標準物質(NIST 1950 基準血漿)、抽出ブランクサンプル、および標準物質を分析しました。
極性および半極性代謝物の分析
血清サンプル40μlを、内部標準混合物(バリン-d8、グルタミン酸-d5、コハク酸-d4、ヘプタデカン酸、乳酸-d3、クエン酸-d4)を含む冷MeOH/H2O(1:1、v/v)90μlと混合した。3-ヒドロキシ酪酸-d4、アルギニン-d7、トリプトファン-d5、グルタミン-d5、1-D4-CA,1-D4-CDCA,1-D4-CDCA,1-D4-GCA,1-D4-GCDCA,1-D4-GLCA,1-D4-GUDCA,1-D4-LCA,1-D4-TCA,1-D4-UDCA)。チューブをボルテックスし、3分間超音波処理した後、遠心分離した(10000rpm、5分間)。遠心後、溶液の上層90μlをLCバイアルに移し、窒素ガス下で蒸発乾固した。乾燥後、サンプルを60μlのMeOH:H2O(70:30)に再溶解した。
分析は、トリプル四重極質量分析計(Waters Corporation, Milford, USA)と結合したAcquity UPLCシステムを用い、マイナスイオンモードで動作する大気圧エレクトロスプレーインターフェースを用いて行った。サンプル10μLのアリコートをAcquity UPLC BEH C18 2.1 mm × 100 mm、1.7μmカラム(Waters Corporation)に注入した。移動相は(A) 2 mM NH4Ac in H2O: MeOH (7:3)と(B) 2 mM NH4Ac in MeOHから成る。グラジエントは以下のようにプログラムした: 0-1分、1%溶媒B; 1-13分、100%溶媒B; 13-16分、100%溶媒B; 16-17分、1%溶媒B、流速0.3 mL/分。分析カラムの再調整を含む総運転時間は20分であった。
BAsとPFASの定量は7点内部検量線を用いて行った。同定は、Metabolomics Standards Initiativeに基づく同定レベル1と2のカスタムデータベースを用いて行った。品質管理は、プールされた品質管理サンプル(各サンプルからプールされたアリコート)をサンプルとともに分析することによって行われた。さらに、品質管理手順の一環として、基準標準物質(NIST 1950 基準血漿)、抽出ブランク試料および標準物質を分析した。
ヒト白血球抗原遺伝子型とNDサブタイプ間の分析
ヒト白血球抗原(HLA)クラスIIの遺伝子型は、3,783人の小児の血液スポットについて、ランタニド標識オリゴヌクレオチドプローブを用いた配列特異的ハイブリダイゼーションを用いて決定した。自己免疫疾患との遺伝的重複と併存症が文献で観察されたため、自己免疫で一般的に報告されているリスク対立遺伝子、特にDR4-DQ8とDR3-DQ2の有病率を、Python 3.11.4のオッズ比を用いてNDとコントロールで比較した。
便サンプルの収集と保存
1,748人の参加乳児から1年後に便サンプルが採取された。ウェルベビークリニックから提供された滅菌ヘラとチューブを用いて、おむつからサンプルを採取した。採取後、乳児の自宅またはクリニックで直ちに冷凍保存した。自宅で採取した検体については、凍結クランプを使用することで、ウェルベビークリニックへの凍結輸送が容易になり、その後の-80℃での乾燥保存が維持された。便サンプルは平均11.93±2.94ヵ月齢で採取され、採取時の対照群とND群の間に有意な年齢差はなかった。
微生物量の配列決定と定量化
抽出と塩基配列決定
便検体からのDNA抽出と、それに続くV3-V4領域を標的とした16S rRNA-PCR増幅が行われた。米国フロリダ州ゲインズビルにあるフロリダ大学の学際的生物工学研究センター(ICBR)で、イルミナMiSeq 2x300 bpを用いて、確立されたプロトコールに従い、10プールに分けた合計1,748サンプルの塩基配列を決定した127,129。標的V3-V4 16S rRNAシーケンス用のアンプリコンは、Standard Illumina Read 1シーケンス/インデキシングプライマー341F(NNNNCCTACGGGAGGCAGCAG)および806R(GGGGACTACVSGGGTATCTAAT)を用いて作製した。
フォワードプライマー:5'- P5 -- アダプター -- リンカー -- SBS3 -- 16S -3'
5' - AATGATACGCGACCGAGCIWTHTAYGGIAARGGIGGIATGIAA - 3'
リバースプライマー:5' - P7 アダプター -- リンカー -- バーコード -- SBS12 -- 16S - 3'
5'-caagcagaagacggcatacgagat- (barcode)- gtgactggagttcagacgtgtgcttccgstctgggactacvsgggtatctaat - 3'
プーリングには、11ヌクレオチド長のバーコードを使用した。各PCRサンプルはプールする前にスピンコラム精製し、Qubitで定量した。
ペアエンドジョイニングとデマルチプレックス
アンプリコンはまずQiime1,135で処理され、ペアエンドリードの結合(join_paired_ends.py)、デマルチプレックス(split_libraries_fastq.py)、別々のfastqファイルの生成(split_sequence_file_on_sample_ids.py)が行われた。最初のQIIME1処理では、フィルタリングが下流の品質スコアの意味に与える潜在的な影響に留意しました。そのため、split_libraries_fastq.pyステップのdemultiplexingオプションを意図的にできるだけ緩やかに設定し、エラー訂正を1つの一貫した方法で実行できるようにしました。具体的には、以下のパラメーターを選びました: -q (maximum unacceptable Phred quality score): q(許容できないPhred品質スコアの最大値):"0 "に設定し、下流処理のためにリードを最も甘く通過させる: 0」に設定。Phred品質スコアが「0」の場合、連続した低品質塩基コール は許容されないため、すべての品質スコアのリードを通過させる: 100 "に設定。これらのパラメータは、DADA2136 Rパッケージでフィルタリングと品質管理の大部分を行うことを意図して、緩やかな選択を行った。
フィルタリングとサンプル推論
DADA2を使用して、品質の低い配列を除去し、高分解能のアンプリコン配列バリアント(ASV)を導出した。DADA2にはエラーモデルが組み込まれており、配列の各位置におけるエラー率を推定することで、生物学的変異とシーケンシングエラーを区別することができます136。このパッケージを使用して、未加工のデマルチプレックスリードの最初の検査(plotQualityProfile)、フィルタリングとトリミング(filterAndTrim)、フィルタリングされたリードのその後の検査(plotQualityProfile)、エラー率の学習(learnErrors、plotErrors)、およびサンプル推論(dada)が実行されました。filterAndTrimステップでは、以下のパラメータを用いた品質管理プロセスを採用した: truncation length (truncLen=c(421))は421塩基より短いリードを廃棄するように設定し、trimming (trimLeft=21)はバーコードを除去するために最初の21塩基を除去するように設定し、その結果、400塩基長のリードが得られるように設定し、maxNは "0"(デフォルト)に設定し、すべてのあいまいな塩基または未確定の塩基(N "文字として表される)を除去するように設定した、 最大期待誤差(maxEE)を "2 "に設定し、この閾値を超える期待誤差(EE = sum(10ˆ(-Q/20)))を持つリードを廃棄し、ΦXゲノムと一致するリードを除去し、入力ファイルを並列にフィルターするためにマルチスレッドを有効にした。その後、リード数が非常に少ない5サンプルを除外した。サンプル推論はdada関数を用いてマルチスレッドで行い、コンセンサス法によるキメラ除去を適用する前に10プールの配列表をマージした。分類学的割り当てはSilva 138137データベースを用いて決定し、必要に応じてNational Center for Biotechnology Information (NCBI) Basic Local Alignment Search Tool (BLAST)138 を用いて検証した。
合計で102,972,740リード(サンプルあたりの平均リード数:61,994.4;中央値: 57,860、最大: 中央値:57,860、最大値:776,158、最小値:10,059): 10,059)、12,844のユニークなASVを含んでいた。このうち、4,938個のASVが少なくとも2人の子供で共有され、2,444個が5人以上の子供に存在した。表S7にASVの全塩基配列を示す。コア分類群の同定には、希薄化データセットを使用し、カウントを21,800リードのシーケンス深度に標準化した結果、27の対照サンプルと636の対応するASVが除去された。
NDの状態を調べるマイクロバイオーム解析では、リード数が少ないサンプルと、NDを併発していないにもかかわらず後に自己免疫疾患と診断された乳児のサンプルを除去した。このうち116人は後にNDと診断され(ABISND)、87人は2つ以上のNDを発症していた。ABISND116人のうち、14人が後に言語障害、7人が知的障害、85人がADHD、39人がASDと診断された。環境因子の解析には、ABISの全サンプルを考慮し、数が少ないサンプル(n=1,743)は除外した。
1年後の便のグローバルおよび標的メタボロミクス
非標的LC-MSメタボローム解析は、出生時の生物学的性別と市町村で傾向スコアマッチングにより選択された46人のサブセット(n=23 ABISASDおよびn=23 ABISC)の便サンプルで実施され、採便時の年齢に差はなかった(ABISControls:10.89±3.51ヵ月、ABISASD:11.19±2.37、p=0.726)。平均重量11.4mgの便サンプルは、細胞抽出を行い、サンプルタンパク質含量の前正規化を行った。
メタボロミクスプロファイリングは、フロリダ大学(フロリダ州ゲインズビル)のSoutheast Center for Integrated Metabolomics(SECIM)で実施した。Dionex UHPLCとオートサンプラーを備えたThermo Q-Exactive Oribtrap質量分析計では、m/z 200で35,000の質量分解能を持つ正および負の加熱エレクトロスプレーイオン化を別々の注入として使用しました。分離にはACE 18-pfp 100 x 2.1 mm, 2 μmカラムを用い、0.1%ギ酸水溶液(移動相A)とアセトニトリル(移動相B)で構成された。この極性固定相は包括的なカバレッジを提供する。非常に極性の高い化学種のカバレッジにおいて、この技術にはいくつかの限界があることに注意すべきである。流速は350μL/min、カラム温度は25℃、注入量はマイナスイオンで4μL、プラスイオンで2μLであった。
合計4,105のフィーチャーが検出され、ポジティブモードでは1,250、ネガティブモードでは2,855であった。MZmine134フリーウェアを使用して、フィーチャー同定、脱同位、アライメント、および最初のアライメントアルゴリズムで見落とされた可能性のあるフィーチャーのギャップ充填を行った。データは付加体および複合体を除去するために処理された。メタボロームフィーチャーは、既存のメタボロームデータベース(Metlin、ChemSpider、pubChem)に基づいて同定するために、1,414化合物からなるSECIMの内部保持時間代謝物ライブラリーを参照しました。
フロリダ大学(フロリダ州ゲインズビル)のSEICでは、選択反応モニタリング(SRM)を使用して、脂肪酸(22代謝物)とトリトファン(6代謝物)についてもターゲットメタボロミクスを実施しました。脂肪酸は LipidMatch140 in silico ライブラリを用いて定量・同定した。
定量化と統計解析
妊娠から幼児期までの環境リスク因子
ABISコホートは、4つの異なる神経発達状態に層別化された: ASD、ADHD、言語障害、知的障害である。複数の診断を受けた子どもは、重複の可能性を考慮し、それぞれの診断カテゴリーに含まれた。これらの各状態の中で、IBM SPSSバージョン29を用いて環境因子(家族病歴、感染症、暴露、生活環境を含む)の有病率を調べた。これらの環境因子は、出生時、1年時、3年時、5年時に実施されたABIS保護者アンケートから得られた。
偽発見率が膨れ上がる可能性を避けるため、的を絞ったアプローチを採用した。まず、条件間で有病率に15%以上の差がある因子を同定した。その後、Python 3.11.4を用いて、各選択因子と診断群の組み合わせについてオッズ比計算を行った。各比較群内の比較数は意図的に50以下に制限した。このような意図的な抑制は、所見の頑健性を維持し、比較の焦点を絞った性質から偽発見補正の必要性をなくすために行われた。同様に、家族の病歴、感染症、暴露、生活環境に関わる因子のみが考慮された。
3年後と5年後の気分と消化器症状のクラスター
ABIS研究に参加した親は、子どもの3歳時と5歳時の診察時に、成長、気分、胃腸の問題に関する11~12項目の二者択一式の質問を含む広範なアンケートに回答した。症状は、成長不良、体重増加不良、食欲不振、腹痛、腹部膨満感/ガス状胃、1日3回以上の下痢、1日3回以上の嘔吐、便秘、疲労、全般的な苛立ち、不機嫌/悲鳴、睡眠の質の低下などを評価するために、「その子供は、以下の症状に苦しんでいるか、または影響を受けていると思いますか」という質問を中心に構成された。
主成分分析(PCA)は、IBM SPSSバージョン29を用いて、小児の3歳時受診時に報告された12の症状および5歳時受診時に報告された11の症状に対する二値回答について実施された。成分はカイザー正規化を伴うバリマックス回転にかけられ、成分得点は回帰により計算された。このアプローチにより、KMO測定が満足のいくサンプリングの妥当性を示し(3歳時のKMO = 0.854、5歳時のKMO = 0.839)、有意なバートレットの球性の検定(いずれもp値 < 0.001)により、PCAに適したデータであることが確認され、成分の解釈が可能となった。
個々の症状および3年間および5年間の累積発生を評価するために、Python 3.11.4を用いてオッズ比を算出した。
耳炎と微生物の比較有病率
生後12ヵ月の耳炎感染に基づくマイクロバイオームの差異を、傾向一致群(n=576)内で求めた。出産様式、妊娠中の抗生物質と喫煙、母乳育児の総月数に基づいて、耳炎感染の有無別に1対1で小児を選択した。非対照群には、耳炎のデータが入手可能なABISの全サンプル(n=1307)が含まれた。FDR補正後のDESeq2の結果に基づいて、有病率検定用に属を選択した。これらの結果に基づいて、Python 3.11.4でCitrobacterとCoprococcusの有病率のオッズ比を算出し、早期耳炎および将来のND転帰との関連を評価した。
腸内細菌叢の交絡因子
腸内細菌叢の組成(β-多様性)に対する交絡因子の影響を調べるために、組成を変換した計数に対してBray-Curtis距離を用い、1000回の並べ替えを採用した並べ替え多変量分散分析(PERMANOVA;adonis関数、veganパッケージ141;R Foundation)を実施した。解析の目的は、生物学的性別、分娩様式、妊娠中の母親の喫煙、地理的地域(スウェーデンの郡)、乳児の抗生物質使用、早期の呼吸器感染症または胃腸感染症、子どもの脆弱性指数スコアなど、いくつかの因子の影響を評価することであった。
ABISControlsとABISNDの間の存在量の差
DESeq2142パッケージを用いて、対照群と将来のND間の腸内細菌叢変動の差次的発現解析を行った。ディスバイオシスも反映する可能性のある将来の自己免疫疾患を有する小児は解析から除外した。サイズ因子と分散の推定値が計算され、負の二項一般線形モデル(GLM)が、平均強度に対する分散の局所型フィッティングを伴うWald統計を使ってフィッティングされた。
GLM解析により、log2 fold change (FC)と標準誤差(lfcSE)の値が得られた。多重比較を考慮するため、Benjamini-Hochberg法により、各解析において偽発見率(FDR)調整p値を算出した。比較は、コントロールとNDを一括して(いずれかの適格NDの診断)、さらに診断の種類(すなわち、ASD、ADHD、言語障害)で層別化して行った。
症状クラスターとリスク因子、およびHLAによる存在量の違い
マイクロバイオーム全コホート(n=1743)を用いて、症状、リスク因子、HLAに関連する関連を、各比較においてFDR補正を行ったDESeq2142パッケージを用いて調査した。3年後および5年後の症状との関連における1歳時の微生物分類群のプロスペクティブ評価では、疲労、腹痛、全身苛立ち、下痢、睡眠の質などの症状の2値分類、および1つ以上の将来のNDの発症と最も強い関連を示した症状クラスターを用いた。1年目と出生時の調査から得られた危険因子と保護因子を二分して分析した。危険因子には、1年目の胃腸炎、抗生物質を必要とする感染症、耳炎、心理社会的脆弱性の増加、妊娠中の母親の喫煙、完全/完全母乳育児の月数の少なさ、1年目のチョコレート、フライドポテト、チップスの頻度の多さ、帝王切開による出産が含まれ、保護因子はその逆(すなわち、 感染症に感染していない;心理社会的脆弱性が減少している、またはない;喫煙していない;完全/排他的母乳育児の期間が長い;最初の1年間にチョコレート、フライドポテト、チップスを食べなかった、または少なかった;経膣分娩による出産)。乳児の食事の特徴は、頻度(毎日または週3~5回と、めったにまたは週1~2回)に基づいて二分した。母乳育児の月数(合計/排他的)は、1~4ヵ月と5ヵ月以上とで二分した。心理社会的脆弱性は、総指標に基づいて高リスク/低リスクに二分した。
1つ目はDR4-DQ8ホモ接合体とDR4-DQ8を持たない人との比較、2つ目はDR3-DQ2/DR4-DQ8を持つ人とこれらのリスク対立遺伝子のいずれかを持たない人との比較である。
出生時の性別で層別化したASD群における診断年齢による存在量の違い
早期のASD診断が表現型の重症度に及ぼす潜在的な影響に対処するため、ABISASDサブグループ内の診断年齢に基づく差異も検討した。これらの解析は、診断年齢の平均が男性ではるかに低い(男性13.4±4.0歳、女性17.6±2.6歳)ことを考慮し、男性と女性に分けて行った。
微生物叢交絡因子とNDリスク因子をコントロールした後の存在量の差(ABISND-Match)
神経発達リスクまたは腸内細菌叢組成に影響を及ぼす交絡因子を制御するために、matchIt48 Rパッケージを用いて傾向スコアマッチングを適用した。考慮した交絡因子は、生物学的性別、分娩様式、地理(スウェーデンの地域/郡)、有害物質曝露(妊娠中の喫煙など)、心理社会的脆弱性指数の合計、乳児の食事(母乳育児の総月数、生後12ヵ月間の牛肉、チョコレート、その他のキャンディー、チップス/チーズドーナッツの摂取頻度を含む)であった。これらの交絡因子のデータが欠落している症例は除外され、82例のND症例(ABISND-Match)が残った。対照群(n=163)は、これらの交絡因子の分布のバランスをとるために、最近傍傾向スコアマッチングを用いて選択された。HLA遺伝子型(p's > 0.162)および抗生物質の使用(p's > 0.203)を含む他の追加的交絡因子は、ABISND-Matchとこれらの選択された対照との間で本質的にバランスがとれていることがわかった。データセットには15,120,951リードが含まれ、サンプルあたり平均61,718.2リード(中央値57,208、最大328,839、最小11,440)で、3,847のユニークなASVにまたがった。1,573個のASVが少なくとも2人の子供に見られ、878個が4人の子供に見られた。ABISND-Match群では、26人の乳児が将来のASD、59人が将来のADHD、13人が将来の言語障害と診断された。ほとんどの乳児は後に1つのND診断しか受けず、ADHD43人、言語障害11人、ASD8人、知的障害1人であったが、19人は複数のND診断を受けた。
DESeq2解析は、前述したように、このマッチさせた乳児群で繰り返された。さらに、最適な分類を達成するために有病間隔に基づいて分類群をフィルタリングするPIME144 Rパッケージを用いて、全体的なマイクロバイオームコミュニティ組成の違いを評価した。これは、マッチさせたNDと対照の間の差異については属レベルで、NDのサブタイプ(すなわち、複数の診断、ASD、ADHD、および言語障害)を分離するためにASVレベルで実行された。
NDサブグループ間でのコア微生物叢の選択
マイクロバイオームの複雑さを考慮し、我々の解析は、NDの状態に基づいて、一般的で広く共有されている細菌種を評価するためにも拡張された。対照群と将来のND(ASD、ADHD、言語障害、知的障害)について決定されたコア分類群は、組成変換されたASVカウントについてマイクロバイオームRパッケージを用いて評価された。
我々のアプローチは2つのデータセットを包含した:完全データセットと希薄データセット(後者はサンプリング深度を均等にするために21,800カウントに制限した。このアプローチでは、有病率が最低10%(言語障害と知的障害については20%)を超えたことのない分類群は除外された。NDサブグループ全体に一貫して存在する主要な微生物分類群を特定するために、microbiome Rパッケージのcore_members関数を採用した。コア微生物は、特定のNDグループに属するサンプルの50%以上で相対存在量が0.01%を超える分類群と定義した。得られたコア分類群は、plot_core R関数を用いてさらに可視化した。
今回の分析結果の信頼性を高めるため、元の希薄化されていないデータセットと希薄化されたデータセットの両方を分析した。注目すべきことに、希薄化のプロセスはコア分類群の同定結果に明確な影響を及ぼさなかった。一貫性を保つため、希薄化データセットは21,800リードの均一なシーケンス深度に調整された。
便メタボロミクス解析と1年後の微生物量との相関
MetaboAnalyst145 5.0を用いて、便サンプル(n=23 ABISASDおよびn=23 ABISControls)における将来のASDとの関連について代謝物濃度を評価し、NDアウトカム、特にASDおよび併存ASD/ADHDとの関連に注目した。その後、選択した代謝物を細菌の相対量および絶対量(後者はqPCRを用いて算出)と比較した。データはサンプルごとの代謝物の合計に正規化され、代謝物濃度はlog-10変換された。便代謝物はポジティブイオンモードとネガティブイオンモードで別々に分析され、既知の571代謝物(ネガティブイオンモードで300代謝物、ポジティブイオンモードで271代謝物)の積算ピーク高強度も含まれました。後にASDと診断された乳児の臍帯血清または便サンプルにおいて濃縮または減少を示す代謝物は、Kruskal Wallis/Mann-Whitney U、部分最小二乗-判別分析(PLS-DA)、およびfold-change分析を用いて同定した。階層的クラスタリングと点双列相関も行った。
便サンプルの脂肪酸の違いは、RのMann-Whitney U検定を用いて将来のASD状態との関連を評価した。モル%が計算され、比較された。注目すべきは、将来ASDになる可能性のある小児は、しばしばパルミトレイン酸の欠乏を示すという観察がなされたことである。これをさらに調べるため、カイ二乗分析を用いてパルミトレイン酸の有病率の差を計算した。
ビフィドバクテリウム(Bifidobacterium)、ロゼブリア(Roseburia)、フェーカリバクテリウム(Faecalibacterium)、アッカーマンシア(Akkermansia)、コプロコッカス(Coprococcus)の相対的な存在量を個別に最もよく予測する上位20の便のメタボローム特徴を、マイナスイオンモードで抽出するために機械学習を利用した。ランダムフォレストリグレッサーを各属で個別に実行し、存在量を予測する上位20代謝物を同定した。その後、スピアマン相関を用いて代謝物と細菌量を関連付けました。この解析はPython 3.11.4で行いました。
MetaboAnalyst 5.0146フレームワーク内でスピアマン相関を用いて、AkkermansiaおよびCoprococcusについて便代謝物濃度との関連性を追加評価した。この解析には、AkkermansiaおよびASV-88 Akkermansia muciniphilaの16Sコピーの総カウント数、Coprococcusの相対存在量、および正規化代謝物濃度が組み込まれた。
臍帯血清メタボロミクスと1年後の微生物量との相関
母親の感染症37、喫煙38、ストレス/深刻なライフイベント39、食事40,41、妊娠中のコーヒー摂取42が臍帯血清と胎児に及ぼす出生前のプログラミングの影響があるため、傾向スコアマッチング48を用いてこれらの要因をコントロールし、将来のASD症例(n=27)に対する対照(n=27)を選択した。このために、出生時の調査から得られた以下の変数について、1:1の最近傍法を採用した:感染症またはその他の感染症、重篤なライフイベント、喫煙、カフェイン摂取、ビタミン・ミネラル・鉄サプリメント、妊娠中の牛乳・乳製品・卵の摂取、さらに母親と父親の教育レベル、子どもの出生週。114例の臍帯血清検体から検出された既知の極性代謝物の濃度と、今回の調査で最も一貫してNDと関連したいくつかの分類群の相対存在量との関係を調べた。ピアソン相関はRで行った。将来のASDと対照の間の差は、RでKruskal Wallis/Mann Whitney検定によって決定した。
謝辞
ABISは、Barndiabetesfonden(スウェーデン小児糖尿病財団)、Swedish Council for Working Life and Social Research(スウェーデン労働生活社会研究評議会)助成金/賞番号FAS2004-1775およびFAS2004-1775、Swedish Research Council助成金/賞番号K2005-72X-11242-11A、K2008-69X-20826-01-4、K2008-69X-20826-01-4の支援を受けた; Östgöta Brandstodsbolag; Medical Research Council of Southeast Sweden (FORSS); JDRF Wallenberg Foundation grant/award number K 98-99D-12813-01A; Region Östergötland and Linköping University, SwedenからのALFおよびLFoU助成金; Joanna Cocozza Foundation。本研究は、JDRF助成金番号1-INO-2018-637-A-Nの助成を受け、助成金契約101094099の下、欧州連合(EU)のホライゾンヨーロッパプログラムの助成を受けた「Inflammation in human early life: targeting impacts on life-course health」(INITIALISE)コンソーシアムの支援を受けた。TriplettおよびLudvigsson研究室のメンバー、特にJordan Russell博士には、技術的支援と過去のシーケンス作業について、またIngela JohanssonおよびJorma Ilonen博士には、参加者サンプルの調達と管理、およびジェノタイピングについてご協力いただいた。UF ICBRおよびSECIMのスタッフの方々には、シーケンスおよびメタボローム解析をサポートしていただきました。原稿に貴重なコメントをいただいたBarry Ahrens博士とLina Colucci博士に感謝する。すべての漫画のスケマティックはBioRender.comのソフトウェアを用いて作成した。この仕事を共に始めた同僚であり友人でもあるPer Gustafssonの思い出に捧げる。
著者の貢献
構想、A.P.A.、E.W.T.、K.I.、T.H.、M.O.、J.L.、方法論、A.P.A.、T.H.、M.O.、E.W.T.、J.R.P.、K.I.、T.J.G.、J.L.、形式分析、A.P.A.、T.H.、J.R.P.、調査、A.P.A、 調査、A.P.A.、T.H.、J.R.P.、C.D.G.;データ管理、A.P.A.、T.H.、J.R.P.、T.J.G.、M.O.、E.W.T.、J.L.;執筆-原案、A.P.A.;執筆-校閲・編集、A.P.A、 E.W.T.、K.I.、T.H.、T.J.G.、J.R.P.、J.L.;可視化、A.P.A.、T.H.、J.R.P.、T.J.G.、E.W.T.;資金獲得、J.L.、M.O.、E.W.T.;リソース、E.W.T.、M.O.、T.J.G.、 プロジェクト管理、J.L.、監督、E.W.T.、M.O.、T.J.G.、J.L.。J.L.はデータおよび生物学的サンプルの収集を含むABIS研究を立ち上げ、現在も主導している。
利害関係の宣言
著者らは、競合する利害関係はないと宣言している。
補足情報
pdfダウンロード(.04 MB)
pdfファイルのヘルプ
表S1. 図4に関連する腸内細菌叢組成の交絡因子
腸内細菌組成に影響を及ぼす交絡因子のパーミュテーショナル多変量分散分析結果。PERMANOVAは、ASVレベルのBray-Curtis距離を用いて腸内細菌組成について実施した。腸内細菌組成と、性別、分娩様式、妊娠中の母親の喫煙、地理、抗生物質による感染、生後1年間の自己申告による疾病、および小児の脆弱性指数(>3)との関連を評価した。PERMANOVAは、adonis Rパッケージを用いて1,000回の並べ替えで計算した。欠損データのため、関連は2つの別々のモデルで解析した。最初のモデルは、欠損データのない1,557検体で観察された13,197分類群について、性別、分娩様式、地理との関連を同時に評価した。2番目のモデルは、欠測データのない1,087サンプルで観察された3,888分類群について、残りの変数の関連を評価した。
.xlsxのダウンロード (.04 MB)
xlsxファイルのヘルプ
表S2. ABISControls(N=1,456)およびABISND(N=116)における、図4に関連する分類学的ランク間の存在量の差。
負の二項分布による差分発現解析の結果。log2 fold change (FC)|が1以上、正規化塩基平均が10以上の有意な分類群を、FCが小さい順に並べた。q値はBenjamini-HochbergによるFDR補正を反映。数値で先行する分類群はASV IDを表す。ASVの全塩基配列は表S7に示す。Log2 fold changeの値が0を超えるものは、コントロールで高い分類群を表す。
.xlsxのダウンロード (.17 MB)
xlsxファイルのヘルプ
表S3. ABISControlsおよび将来のNDにおいてより豊富な腸内細菌叢、ならびにそれらの保護因子およびリスク因子との関連(図4および5に関連)。
DESeq2 Rパッケージの負の二項一般線形モデルを用いた腸内細菌叢の存在量の差分析で、各変数の偽発見率補正を考慮した。表には、変数の統計的結果と、様々な差分解析モデル(例えば、すべてのND、ND-Match、ADHD、ASD、または言語障害)におけるコントロールおよび将来の神経発達障害(ND)に対する保護因子およびリスク因子と関連する分類群間の比較が含まれている。ND-Matchは、説明したように、傾向スコアマッチングによって選択された対照と将来のNDとの比較から得られた差分豊度を表す。乳児の食事頻度(毎日/週3~5回 vs めったに/週1~2回)、母乳育児の月数(合計/排他的)、心理社会的脆弱性(高リスク/低リスク)など、出生時および1年間の質問票の変数は、解析のために二分された。危険因子は、胃腸炎、抗生物質が必要な感染症、1年目の耳炎、心理社会的脆弱性の増加、妊娠中の母親の喫煙、授乳期間の短縮、チョコレート、フライドポテト、チップスの頻繁な摂取、帝王切開出産を考慮し、防御因子はそれらの逆数を示した。分類群および同定された危険因子に関連する特定のND群は、「危険因子」タブに明記されている。
.xlsxのダウンロード (.02 MB)
xlsxファイルのヘルプ
表S4. 図4に関連する、将来の神経発達障害(ND)を有する乳児とマッチさせた対照を比較し、NDのサブタイプで分離した有病率モデルにおける最大の寄与者
アンプリコン配列変異体(ASV)の有病率に基づき、マッチドコントロールと将来のNDを比較したPIMEモデルおよびNDサブタイプ別に将来のNDを比較したPIMEモデルにおける最大の寄与者。最初のモデルでは、NDリスク因子とマイクロバイオームの交絡因子に関する最近傍傾向スコアマッチングを用いて、マッチさせた対照を2:1の割合で選択した。ABISND-Match(n=81)をマッチさせた対照(n=163)または将来のND(n=114;すなわち、多発性、ASD、ADHD、または言語障害)と区別するランダムフォレストモデルの統計。平均減少精度(MDA)に基づく上位分類群の寄与を示す。分類群は有病率に基づいてフィルタリングされ、OOB(out-of-bag)誤差が最も小さく、分類群の数が最も多い有病区間が選択された。
pdfをダウンロード(.04 MB)
pdfファイルのヘルプ
表S5. 図5に関連する、対照群と比較して将来の神経発達障害(ND)症例全体で有意に豊富な腸内細菌叢
DESeq2による将来のすべてのND(n = 116)、ASD(n = 39)、ADHD(n = 85)、言語障害(n = 14)における症例対対照の統計(log-fold変化)。正規化塩基平均数≧10で存在する細菌のみが示されている。
pdfをダウンロードする (.05 MB)
pdfファイルのヘルプ
表S6. 図6に関連する、将来の自閉症スペクトラム障害(ASD)症例と対照(n = 46)間で発現に差のあるその他の便中代謝産物
ポジティブイオンモードとネガティブイオンモードで発現に有意差を示した1歳時の便中代謝物。m/zと保持時間は、LC-MSで明確なm/z値を持つ複数のピークで表される代謝物を区別するために記載した。平均値と標準偏差 (SD) が描かれており、p 値は R で実行した Mann-Whitney U 検定に対応しています。
ダウンロード.xlsx (1.72 MB)
xlsxファイルのヘルプ
表S7. 図4および5に関連するABISアンプリコン配列バリアント(ASV)の完全配列
DADA2解析の結果得られた各ABIS ASVの400塩基16S rRNA全塩基配列。Silva138データベースを用いて分類した。
参考文献
Thomas R.
サンダース S.
Doust J.
ベラー E.
グラスジウP.
注意欠陥/多動性障害の有病率:系統的レビューとメタ分析。
小児科。2015; 135: e994-e1001
https://doi.org/10.1542/peds.2014-3482
論文で見る
スコープス (1268)
PubMed
クロス
グーグル奨学生
フェイヤド J.
デ・グラーフR.
ケスラーR.
アロンソJ.
アンガーマイヤー M.
デミテネール K.
デ・ジローラモ G.
ハロ J.M.
カラム E.G.
ララ C.
ら。
成人の注意欠陥多動性障害の国家を越えた有病率と相関関係。
Br. J. Psychiatry. 2007; 190: 402-409
https://doi.org/10.1192/bjp.bp.106.034389
論文で見る
スコープス (1039)
PubMed
クロス
グーグル奨学生
ポズナーJ.
ポランチクG.V.
ソヌガ-バークE.
注意欠陥多動性障害。
ランセット。2020; 395: 450-462
https://doi.org/10.1016/S0140-6736(19)33004-1
記事で見る
スコープス (381)
パブコメ
概要
全文
全文PDF
グーグル奨学生
ブラックL.I.
バハラチアンA.
ホフマンH.J.
3~17歳の子どものコミュニケーション障害と介入サービスの利用: 米国、2012年。
NCHS Data Brief。2015; 2012: 1-8
記事で見る
グーグル・スカラー
ノーバリー C.F.
グーチ D.
レイC.
ベアードG.
チャーマン T.
サイモノフ E.
ヴァンヴァカス G.
ピクルスA.
言語障害の有病率と臨床症状における非言語能力の影響:集団調査からの証拠。
J. 児童精神医学。Psychiatry. 2016; 57: 1247-1257
https://doi.org/10.1111/jcpp.12573
論文で見る
スコープス (507)
PubMed
クロス
グーグル奨学生
シャオ E.Y.
マクブライド S.W.
Hsien S.
シャロンG.
ハイド E.R.
マキュー T.
コデリ J.A.
チャウ J.
ライズマン S.E.
ペトロシーノ・J.F.
他
微生物叢は神経発達障害に伴う行動的・生理的異常を調節する。
Cell. 2013; 155: 1451-1463
https://doi.org/10.1016/j.cell.2013.11.024
記事で見る
日本学術振興会特別研究員
パブコメ
概要
全文
全文PDF
グーグル奨学生
ベルディング K.
ドノバン S.M.
自閉症スペクトラム障害におけるマイクロバイオームと栄養:現在の知識と研究の必要性。
Nutr. Rev. 2016; 74: 723-736
https://doi.org/10.1093/nutrit/nuw048
論文で見る
スコープス (65)
PubMed
クロス
グーグル奨学生
マリンズC.
フィシェルG.
ツィエンR.W.
自閉症スペクトラムの統一見解: 自己調節フィードバックループの考察。
Neuron. 2016; 89: 1131-1156
https://doi.org/10.1016/j.neuron.2016.02.017
記事で見る
スコープス (127)
PubMed
概要
全文
全文PDF
グーグル奨学生
メルツァーA.
ヴァン・デ・ウォーターJ.
自閉症スペクトラム障害における免疫系の役割。
Neuropsychopharmacology. 2017; 42: 284-298
https://doi.org/10.1038/npp.2016.158
論文で見る
スコープス(308)
PubMed
クロス
グーグル奨学生
ウィルフォース C.
カールソンT.
アンデルリッドB.M.
ノルドグレンA.
Kostrzewa E.
ベルグレン S.
ロナルド A.
クジャ・ハルコラ R.
タミミエス K.
Bölte S。
不一致の双子の病歴と自閉症の環境病因。
翻訳。Psychiatry. 2017; 7e1014
https://doi.org/10.1038/tp.2016.269
論文で見る
スコープス (23)
クロスリファレンス
グーグル奨学生
メイヤー E.A.
パドヴァD.
ティリッシュK.
自閉症における脳腸軸の変化:併存疾患または原因メカニズム?
バイオエッセイ。2014; 36: 933-939
https://doi.org/10.1002/bies.201400075
記事で見る
スコープス (231)
パブコメ
クロス
グーグル奨学生
マケルハノンB.O.
マクラッケンC.
カーペンS.
シャープW.G.。
自閉症スペクトラム障害における胃腸症状:メタアナリシス。
小児科。2014; 133: 872-883
https://doi.org/10.1542/peds.2013-3995
論文で見る
スコープス (502)
PubMed
クロス
グーグル奨学生
ドシ-ベレスF.
葛 Y.
Kohane I.
自閉症スペクトラム障害における併存疾患クラスター:電子カルテ時系列分析。
Pediatrics. 2014; 133: e54-e63
https://doi.org/10.1542/peds.2013-0819
論文で見る
スコープス (264)
PubMed
クロス
グーグル奨学生
サンプソンT.R.
マズマニアン S.K.
マイクロバイオームによる脳の発達、機能、行動の制御。
Cell Host Microbe. 2015; 17: 565-576
https://doi.org/10.1016/j.chom.2015.04.011
論文で見る
スコープス (729)
PubMed
概要
全文
全文PDF
グーグル奨学生
マルケージJ.R.
アダムス D.H.
ファバ F.
エルメスG.D.A.
ハーシュフィールド G.M.
ホールド G.
クライシ M.N.
キンロス J.
スミット H.
トゥーヒー K.M.
ほか
腸内細菌叢と宿主の健康:新しい臨床フロンティア。
Gut. 2016; 65: 330-339
https://doi.org/10.1136/gutjnl-2015-309990
論文で見る
スコープス (1587)
PubMed
クロス
グーグル奨学生
クライアンJ.F.
ディナン T.G.
マイクロバイオーム革命を語る
Nat. Microbiol. 2019; 4: 552-553
https://doi.org/10.1038/s41564-019-0422-9
論文で見る
スコープス (17)
PubMed
クロス
グーグル奨学生
エドミストンE.
アッシュウッドP.
ヴァンデウォーターJ.
自己免疫、自己抗体、および自閉症スペクトラム障害(ASD)。
生物学。Psychiatry. 2017; 81: 383-390
https://doi.org/10.1016/j.biopsych.2016.08.031
論文で見る
スコープス (99)
パブコメ
概要
全文
全文PDF
グーグル奨学生
ベルカイド Y.
ハンドT.W.
免疫と炎症における微生物叢の役割。
Cell. 2014; 157: 121-141
https://doi.org/10.1016/j.cell.2014.03.011
記事で見る
スコープス(3131)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
本田 浩二
リットマンD.R.
適応免疫恒常性と疾患における微生物叢。
Nature. 2016; 535: 75-84
https://doi.org/10.1038/nature18848
論文で見る
スコープス (1182)
PubMed
クロス
グーグル奨学生
ウー・エイチ・ジェイ
ウー E.
免疫恒常性と自己免疫における腸内細菌叢の役割。
Gut Microbes. 2012; 3: 4-14
https://doi.org/10.4161/gmic.19320
記事で見る
スコパス (786)
PubMed
クロス
グーグル奨学生
鎌田直樹
Seo S.U.
チェン・G.Y.
ヌニェスG.
免疫と炎症性疾患における腸内細菌叢の役割。
Nat. Rev. Immunol. 2013; 13: 321-335
https://doi.org/10.1038/nri3430
論文で見る
スコープス (1603)
PubMed
クロス
グーグル奨学生
バーンC.S.
チェンバーズE.S.
モリソン D.J.
フロストG.
食欲調節とエネルギー恒常性における短鎖脂肪酸の役割。
イント。J.オブス。(ロンド)。2015; 39: 1331-1338
https://doi.org/10.1038/ijo.2015.84
論文で見る
スコープス (444)
PubMed
クロス
グーグル奨学生
Lyte M.
微生物-腸-脳軸における微生物内分泌学:神経化学物質の細菌産生と利用が行動に及ぼす影響。
PLoS Pathog. 2013; 9e1003726
https://doi.org/10.1371/journal.ppat.1003726
論文で見る
スコパス (285)
クロスリファレンス
グーグル奨学生
フォスター・J.A.
ライテ M.
マイヤー E.
Cryan J.F.
腸内細菌叢と脳機能: 神経科学の発展分野。
Int. J. Neuropsychopharmacol. 2016; 19pyv114
https://doi.org/10.1093/ijnp/pyv114
論文で見る
スコープス (138)
Crossref
グーグル奨学生
クライアンJ.F.
オリオダン K.J.
コーワン C.S.M.
サンドゥ K.V.
バスティアンスセン T.F.S.
ボーメ M.
コダニョーネ M.G.
クッソット S.
フリング C.
ゴルベバA.V.
他
微生物叢-腸-脳軸。
Physiol. Rev. 2019; 99: 1877-2013
https://doi.org/10.1152/physrev.00018.2018
論文で見る
スコープス (2137)
PubMed
Crossref
グーグル奨学生
ゲオルゲ C.E.
マーティンJ.A.
マンリケス F.V.
ディナン T.G.
クライアンJ.F.
クラークG.
本質に焦点を当てる:トリプトファン代謝と微生物-腸-脳軸。
Curr. 意見。Pharmacol. 2019; 48: 137-145
https://doi.org/10.1016/j.coph.2019.08.004
論文で見る
スコープス (115)
PubMed
クロス
グーグル奨学生
スグリッタ M.
ドーリング S.W.
バフィントンS.A.
モミンE.N.
フランシス M.B.
ブリトン R.A.
コスタ-マティオリM.
自閉症スペクトラム障害モデルマウスにおける社会的行動における微生物介在性変化の根底にあるメカニズム。
Neuron. 2019; 101: 246-259.e6
https://doi.org/10.1016/j.neuron.2018.11.018
論文で見る
スコープス (439)
PubMed
要旨
全文
全文PDF
グーグル・スカラー
バフィントン S.A.
ディ・プリスコG.V.D.
オーヒトゥング T.A.
アジャミ N.J.
ペトロシーノ・J.F.
コスタ-マティオリM.
微生物の再構成は、母親の食事が誘発する子孫の社会的およびシナプス欠損を逆転させる。
Cell. 2016; 165: 1762-1775
https://doi.org/10.1016/j.cell.2016.06.001
論文で見る
スコープス (763)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
カン D.W.
アダムス J.B.
グレゴリー A.C.
ボロディ T.
チティック L.
ファザーノ A.
コルッツ A.
ガイス E.
マルドナード J.
マクドノー=ミーンズS.
他。
微生物叢移植療法は腸内生態系を変化させ、胃腸症状と自閉症症状を改善する:非盲検試験。
Microbiome. 2017; 5: 10
https://doi.org/10.1186/s40168-016-0225-7
論文で見る
スコープス (869)
PubMed
クロス
グーグル奨学生
カン D.W.
アダムスJ.B.
コールマン D.M.
ポラード E.L.
マルドナード J.
マクドノー=ミーンズ S.
カポラソ J.G.
クラジュマルニク-ブラウンR.
自閉症症状と腸内細菌叢に対する微生物叢移植療法の長期的効果。
Sci. Rep. 2019; 9: 5821
https://doi.org/10.1038/s41598-019-42183-0
論文で見る
スコープス (404)
PubMed
クロス
グーグル奨学生
ヴオンH.E.
Hsiao E.Y.
自閉症スペクトラム障害における腸内細菌叢の新たな役割。
Biol. Psychiatry. 2017; 81: 411-423
https://doi.org/10.1016/j.biopsych.2016.08.024
論文で見る
スコープス (379)
パブコメ
概要
全文
全文PDF
グーグル奨学生
ブル=ラーセン S.
モハジェリ M.H.
ADHDの発症と進行における細菌マイクロバイオームの潜在的影響。
Nutrients. 2019; 11: E2805
https://doi.org/10.3390/nu11112805
論文で見る
スコープス (54)
PubMed
クロス
グーグル奨学生
ソルディロ J.E.
コリック S.
ラランホN.
キャリーV.
ワインストックG.M.
ゴールド D.R.
オコナー G.
サンデル M.
バカリエL.B.
ベイジェルマンA.
他
乳児腸内細菌叢と幼児期の神経発達アウトカムとの関連: VDAART無作為化臨床試験の付随研究。
JAMA Netw. Open. 2019; 2e190905
https://doi.org/10.1001/jamanetworkopen.2019.0905
論文で見る
スコープス (70)
PubMed
クロス
グーグル奨学生
カールソン A.L.
シャ・ケイ
アズカラートペリルM.A.
ゴールドマンB.D.
アーン M.
スタイナーM.A.
トンプソン A.L.
ゲン X.
ギルモア J.H.
ニックマイヤーR.C.
認知機能の発達に関連する乳児腸内細菌叢。
Biol. Psychiatry. 2018; 83: 148-159
https://doi.org/10.1016/j.biopsych.2017.06.021
論文で見る
スコープス (324)
パブコメ
概要
全文
全文PDF
グーグル奨学生
ラウエH.E.
コーリック S.A.
ベイカー E.R.
カラガス M.R.
マダン J.C.
3歳時点での自閉症に関連する社会的行動と乳児の腸内マイクロバイオームおよび微生物機能の前向き関連性。
Sci. Rep. 2020; 10: 15515
https://doi.org/10.1038/s41598-020-72386-9
論文で見る
スコープス (24)
パブコメ
クロス
グーグル奨学生
ラコルテE.
ジェルバシ G.
バチガルポI.
ヴァナコーレ N.
ラウッチ U.
パリシP。
神経発達障害を持つ子どもたちのマイクロバイオームのシステマティックレビュー。
フロント。Neurol. 2019; 10: 727
https://doi.org/10.3389/fneur.2019.00727
論文で見る
Crossref
グーグル・スカラー
マイヤー U.
イー・ビー・ケイ
フェルドンJ.
妊娠時期の違いによる出生前感染症の神経発達への影響:早ければ早いほど悪いのか?
神経科学者。2007; 13: 241-256
https://doi.org/10.1177/1073858406296401
記事で見る
スコープス (229)
PubMed
クロス
グーグル奨学生
フライシュ A.F.
リファス-シマンS.L.
ロコフL.B.
ハイヴァート M.F.
マンゾロス C.S.
オーケンE.
母親の出生前喫煙と臍帯血ホルモンとの関連: プロジェクトVivaコホート。
Metabolism. 2017; 72: 18-26
https://doi.org/10.1016/j.metabol.2017.04.001
論文で見る
スコープス (16)
PubMed
概要
全文
全文PDF
グーグル奨学生
グライニック・フィリップR.
ダヤンJ.
チョクロン S.
ジャケ A.Y.
トルジュマンS.
胎児と子供の発達に出生前ストレスの影響: 批判的な文献レビュー。
神経科学。Biobehav. Rev. 2014; 43: 137-162
https://doi.org/10.1016/j.neubiorev.2014.03.022
論文で見る
スコープス(262)
PubMed
クロス
グーグル奨学生
ハリス C.P.
ラムロチャンシンC.
ウールO.
デンメルメアH.
ハインリッヒJ.
コレツコ B.
スタンドル M.
ティエリングE.
妊娠中の母親の食事と臍帯血中の代謝物プロファイルの関連性。
Biomolecules. 2022; 12: 1333
https://doi.org/10.3390/biom12101333
論文で見る
スコープス (0)
クロス
グーグル奨学生
プフェッファーレ P.I.
ビュシュレG.
Blümer N.
ロポネン M.
エゲ M.J.
クラウス・エッチマン S.
Genuneit J.
ヒュヴァリネン A.
ヒルボネン M.R.
ラウエナーR.
他。
臍帯血サイトカインは、妊娠中の母親の農業活動および農場乳製品の摂取によって調節される: PASTURE研究。
J. アレルギー臨床。Immunol. 2010; 125: 108-115.e1-3
https://doi.org/10.1016/j.jaci.2009.09.019
論文で見る
スコープス (153)
パブコメ
概要
全文
全文PDF
グーグル奨学生
セングピールV.
エリンド E.
バセリス J.
ニルソン S.
グローブ J.
マイレ R.
ハウゲン M.
メルツァー H.M.
アレクサンダー J.
ヤコブソンB.
他
妊娠中の母親のカフェイン摂取量は出生体重と関連するが、妊娠期間とは関連しない:大規模な前向き観察コホート研究の結果。
BMCメド。2013; 11: 42
https://doi.org/10.1186/1741-7015-11-42
記事で見る
スコープス (134)
パブコメ
クロス
グーグル奨学生
ケイル A.
ダニエルズJ.L.
フォルセンU.
ハルトマン C.
Cnattingius S.
セーデルベリ K.C.
フェイヒティング M.
スパレンP.
子孫の自閉症スペクトラム障害と関連する親の自己免疫疾患。
疫学。2010; 21: 805-808
https://doi.org/10.1097/EDE.0b013e3181f26e3f
論文で見る
スコープス (132)
パブコメ
クロス
グーグル奨学生
モロイ C.A.
モローA.L.
マインゼンデールJ.
ドーソンG.
ベルニエ R.
ダン M.
ハイマン S.L.
マクマホン W.M.
グーディ・ナイス J.
ヘップバーン S.
他
自閉症スペクトラム障害児における退行の危険因子としての家族性自己免疫性甲状腺疾患:CPEA研究。
J. Autism Dev. Disord. 2006; 36: 317-324
https://doi.org/10.1007/s10803-005-0071-0
論文で見る
スコープス (87)
PubMed
クロス
グーグル奨学生
ヒューズ H.K.
ミルズ・コE.
ローズ D.
アッシュウッドP.
自閉症スペクトラム障害の病理学的メカニズムとしての免疫機能障害と自己免疫。
フロント。細胞。Neurosci. 2018; 12: 405
https://doi.org/10.3389/fncel.2018.00405
論文で見る
スコープス (146)
PubMed
クロス
グーグル奨学生
アリー T.A.
井出 A.
ジャハロミM.M.
バーカー J.M.
フェルナンド M.S.
バブ S.R.
Yu L.
ミャオ D.
エルリッヒ・H.A.
フェイン P.R.
他
1A型糖尿病の極端な遺伝的リスク。
Proc. Natl. Sci. USA. 2006; 103: 14074-14079
https://doi.org/10.1073/pnas.0606349103
論文で見る
スコープス (183)
PubMed
クロス
グーグル奨学生
シェティ S.A.
ヒューゲンホルツF.
Lahti L.
Smidt H.
デ・ヴォス W.M.
腸内細菌叢の景観化:群集形成の洞察と微生物調節戦略への示唆。
FEMS Microbiol. Rev. 2017; 41: 182-199
https://doi.org/10.1093/femsre/fuw045
論文で見る
スコープス (139)
PubMed
クロス
グーグル奨学生
ホー D.E.
今井和彦
キングG.
スチュアート E.A.
MatchIt: パラメトリック因果推論のためのノンパラメトリック前処理。
J. Stat. Softw. 2011; 42: 1-28
https://doi.org/10.18637/jss.v042.i08
記事で見る
スコープス (2579)
クロスリファレンス
グーグル奨学生
バロン・コーエン S.
ロンバルド M.V.
Auyeung B.
アシュウィン E.
チャクラバルティB.
クニックマイヤーR.
自閉症スペクトラムはなぜ男性に多いのか?
PLoS Biol.
https://doi.org/10.1371/journal.pbio.1001081
記事で見る
スコープス (510)
クロスフィルム
グーグル奨学生
Orr S.K.
パルンボ S.
ボセッティ F.
マウント H.T.
カン J.X.
グリーンウッド C.E.
マ D.W.L.
セルハン C.N.
バジネットR.P.
未エステル化ドコサヘキサエン酸は神経炎症において保護的である。
J. Neurochem. 2013; 127: 378-393
https://doi.org/10.1111/jnc.12392
論文で見る
スコープス (133)
パブコメ
クロス
グーグル奨学生
オアー S.K.
トレパニエM.O.
バジネットR.P.
神経炎症を伴う動物モデルにおけるn-3多価不飽和脂肪酸。
プロスタグランジンLeukot。Essent。脂肪酸。2013; 88: 97-103
https://doi.org/10.1016/j.plefa.2012.05.008
記事で見る
スコープス (86)
パブコメ
概要
全文
全文PDF
グーグル奨学生
バジネR.P.
レイエ S.
脳機能と疾患における多価不飽和脂肪酸とその代謝産物。
Nat. Rev. Neurosci. 2014; 15: 771-785
https://doi.org/10.1038/nrn3820
論文で見る
スコープス (954)
PubMed
クロス
グーグル奨学生
カスティーヨ P.E.
ヨウンツ T.J.
チャベス A.E.
ハシモトダニ Y.
エンドカンナビノイドシグナル伝達とシナプス機能。
Neuron. 2012; 76: 70-81
https://doi.org/10.1016/j.neuron.2012.09.020
論文で見る
筑波大学
パブコメ
要旨
全文
全文PDF
グーグル奨学生
カイパースR.S.
ラックスウォルダ M.F.
オフリンガ P.J.
ボースマ E.R.
ダイク・ブルーワー D.A.J.
ムスキートF.A.J.
胎児の子宮内全身リノール酸、アラキドン酸、ドコサヘキサエン酸含量と付加率。
プロスタグランジンLeukot。Essent。脂肪酸。2012; 86: 13-20
https://doi.org/10.1016/j.plefa.2011.10.012
論文で見る
スコープス (110)
パブコメ
概要
全文
全文PDF
グーグル奨学生
Cunnane S.C.
プルード M.
ピフェリ F.
ベギン M.
フェアール C.
バーバーガー-ガトーP。
魚、ドコサヘキサエン酸とアルツハイマー病。
プログ。脂質研究2009; 48: 239-256
https://doi.org/10.1016/j.plipres.2009.04.001
記事で見る
スコパス(269)
PubMed
クロス
グーグル奨学生
マクナマラ R.K.
ハーンC.G.
ジャンダチェックR.
ライダーT.
ツォ P.
スタンフォード K.E.
リッチタンN.M.
大うつ病性障害患者の死後眼窩前頭皮質におけるオメガ3脂肪酸ドコサヘキサエン酸の選択的欠損。
Biol. Psychiatry. 2007; 62: 17-24
https://doi.org/10.1016/j.biopsych.2006.08.026
論文で見る
スコープス (219)
パブコメ
概要
全文
全文PDF
グーグル奨学生
キーリー・S.J.
ステア C.J.
ラジザック-マッギンレーN.K.
ウルソデオキシコール酸:炎症性腸疾患の有望な治療標的か?
Am. J.フィジオール。Gastrointest. Liver Physiol: G872-G881
https://doi.org/10.1152/ajpgi.00163.2019
論文で見る
スコープス (23)
Crossref
グーグル奨学生
アブデルカデル N.F.
サファル M.M.
サレムH.A.
ウルソデオキシコール酸はパーキンソン病のロテノンモデルにおけるアポトーシスカスケードを改善する: ミトコンドリア障害の調節。
Mol. Neurobiol. 2016; 53: 810-817
https://doi.org/10.1007/s12035-014-9043-8
論文で見る
スコープス (90)
クロスリファレンス
グーグル奨学生
キーン C.D.
ロドリゲス C.M.P.
アイヒ T.
チャブラ M.S.
ステア C.J.
ロー W.C.
胆汁酸の一種であるタウロウルソデオキシコール酸は、ハンチントン病のトランスジェニック動物モデルにおいて神経保護作用を示す。
Proc. Natl. Sci. USA. 2002; 99: 10671-10676
https://doi.org/10.1073/pnas.162362299
論文で見る
スコープス (264)
PubMed
クロス
グーグル奨学生
ロドリゲス C.M.P.
ソラ S.
ナン Z.
カストロ R.E.
リベイロ P.S.
ロー W.C.
ステア C.J.
タウロソデオキシコール酸は、ラットの急性出血性脳卒中後のアポトーシスを減少させ、神経傷害から保護する。
Proc. Natl. Sci. USA. 2003; 100: 6087-6092
https://doi.org/10.1073/pnas.1031632100
論文で見る
スコープス (184)
PubMed
クロス
グーグル奨学生
パリー G.J.
ロドリゲス C.M.P.
アラニャ M.M.
ヒルベルト S.J.
デイビー C.
ケルカー P.
ロー W.C.
ステア C.J.
筋萎縮性側索硬化症患者におけるウルソデオキシコール酸の安全性、忍容性および脳脊髄液透過性。
Clin. Neuropharmacol. 2010; 33: 17-21
https://doi.org/10.1097/WNF.0b013e3181c47569
論文で見る
スコープス (82)
PubMed
クロス
グーグル奨学生
クリスチャン N.P.
過フッ素化および多フッ素化アルキル物質(PFAS)の化学毒性。
in: Wexler P. Encyclopedia of Toxicology. 第4版。Academic Press, 2024: 747-756
https://doi.org/10.1016/B978-0-12-824315-2.01052-6
記事で見る
スコープス (0)
クロスリファレンス
グーグル奨学生
オモイケ O.E.
パック R.P.
マムドゥ H.M.
リュー Y.
ストラッサー S.
鄭 S.
オコロ J.
Wang L.
パーおよびポリフルオロアルキル物質と炎症および酸化ストレスマーカーとの関連。
Environ. 2021; 196: 110361
https://doi.org/10.1016/j.envres.2020.110361
論文で見る
スコパス (55)
クロスリファレンス
グーグル奨学生
フェントン S.E.
デュカットマンA.
ブービスA.
デウィット J.C.
ラウ C.
ン C.
スミス J.S.
ロバーツ S.M.
ペルフルオロアルキルおよびポリフルオロアルキル物質の毒性とヒトの健康に関するレビュー: 知見の現状と今後の研究に生かすための戦略。
Environ. Toxicol. Chem. 2021; 40: 606-630
https://doi.org/10.1002/etc.4890
論文で見る
スコープス (641)
パブコメ
クロス
グーグル奨学生
デ・マジストリスL.
ファミリアーリV.
パスコットA.
サポーネA.
フロリ A.
イアルディーノ P.
カルテニ M.
デ・ローザ M.
フランカヴィッラ R.
リーガーG.
他
自閉症スペクトラム障害患者とその一親等の近親者における腸管バリアの変化。
J. 小児科。Gastroenterol. 2010; 51: 418-424
https://doi.org/10.1097/MPG.0b013e3181dcc4a5
論文で見る
スコープス(402)
PubMed
クロス
グーグル奨学生
D'Eufemia P.
チェッリM.
フィノッキアーロR.
パシフィコL.
ヴィオッツィ L.
ザッカニーニ M.
カルディ E.
ジャルディーニO.
自閉症児における腸管透過性の異常。
1996; 85: 1076-1079
https://doi.org/10.1111/j.1651-2227.1996.tb14220.x
記事で見る
スコープス (310)
PubMed
クロス
グーグル奨学生
ルオ Y.
ラン C.
Li H.
Ouyang Q.
Kong F.
Wu A.
Ren Z.
Tian G.
Cai J.
Yu B.
他。
腸の健康をターゲットとしたAkkermansia muciniphilaの合理的考察:利点と課題。
NPJ Biofilms Microbiomes. 2022; 8: 81
https://doi.org/10.1038/s41522-022-00338-4
論文で見る
スコープス (19)
クロスフィルム
グーグル奨学生
エンゲヴィク M.A.
モーラ C.N.
ロートD.
エンゲヴィク K.
スピンラーJ.K.
デバラジ S.
クロフォード S.E.
エステス M.K.
カルカム M.
バーサロビッチJ.
腸内葉酸産生の微生物代謝能と宿主葉酸受容体の調節。
フロント。Microbiol. 2019; 10: 2305
https://doi.org/10.3389/fmicb.2019.02305
論文で見る
スコープス (89)
PubMed
クロス
グーグル奨学生
ルイス P.
フリント H.J.
ヒト大腸内細菌叢によるプロピオン酸および酪酸の生成。
Environ. Microbiol. 2017; 19: 29-41
https://doi.org/10.1111/1462-2920.13589
論文で見る
スコープス (1420)
PubMed
クロス
グーグル奨学生
デリエン M.
ヴォーン E.E.
プラグC.M.
デ・ヴォス W.M.
ヒト腸管ムチン分解菌Akkermansia muciniphila gen.
Int. J. Syst. Evol. Microbiol. 2004; 54: 1469-1476
https://doi.org/10.1099/ijs.0.02873-0
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
ロイナネンJ.
カイヌライネンV.
Huuskonen L.
オットマン N.
ベルザー C.
フーティネン H.
デ・ヴォス W.M.
里仮理子
アッカーマンシア(Akkermansia muciniphila)は腸細胞に付着し、上皮細胞層の完全性を強化する。
Appl. Environ. Microbiol. 2015; 81: 3655-3662
https://doi.org/10.1128/AEM.04050-14
論文で見る
スコープス (390)
PubMed
クロス
グーグル奨学生
ロドリゲス V.F.
エリアス-オリベイラJ.
ペレイラ
ペレイラ J.A.
バルボーザ S.C.
マチャド M.S.G.
カルロスD.
Akkermansia muciniphilaと腸管免疫系: 炎症性腸疾患、肥満、糖尿病を抑制する仲良し。
フロント。Immunol. 2022; 13: 934695
https://doi.org/10.3389/fimmu.2022.934695
論文で見る
スコープス (60)
クロスリファレンス
グーグル奨学生
Wang J.
ブレイズ・J.
ハギギF.
キム・シュルツ S.
ラバル U.
トラゲザー K.J.
パシネッティ G.M.
ヒト単球における免疫炎症反応の減弱における新規微生物由来エピジェネティック修飾因子としての3(3,4-ジヒドロキシ-フェニル)プロピオン酸の特性。
Mol. Immunol. 2020; 125: 172-177
https://doi.org/10.1016/j.molimm.2020.07.003
論文で見る
スコープス (6)
クロスリファレンス
グーグル奨学生
ブルーメンダール M.
フロボースM.I.
ウェグマンJ.
ザンドベルトB.B.
ファンデレストO.
クールス R.
アーツ E.
健康な高齢者における反応的および積極的反応抑制に対するチロシン急性投与の神経認知効果。
eNeuro. 2018; 5eneuro.0035-17.2018
https://doi.org/10.1523/ENEURO.0035-17.2018
論文で見る
スコープス (19)
Crossref
グーグル奨学生
ヤングS.N.
ストレスの影響を緩和するL-チロシン?
J. 精神医学神経科学。2007; 32: 224
記事で見る
グーグル学者
ダンカン S.H.
バルセニーラ A.
スチュワート C.S.
プライドS.E.
フリント H.J.
ヒト大腸内酪酸産生菌における酢酸利用およびブチリル補酵素A(CoA):酢酸CoA転移酵素。
Appl. Environ. Microbiol. 2002; 68: 5186-5190
https://doi.org/10.1128/AEM.68.10.5186-5190.2002
論文で見る
スコープス (490)
PubMed
クロス
グーグル奨学生
玉内シャコオリZ.
スミダ I.
ブサールギンL.
ロレールO.
ムーリック V.
フォン S.B.
ボヌール・マレ M.
ジョリヴェ-グジョンA.
Roseburia属:健康のマーカー?
Future Microbiol. 2017; 12: 157-170
https://doi.org/10.2217/fmb-2016-0130
記事で見る
スコープス (419)
PubMed
クロス
グーグル奨学生
サルカー A.
ハーティ S.
ジョンソンK.V.-A.
モーラー A.H.
カーモディ R.N.
レト S.M.
エドマン S.E.
ダンバー R.I.M.
バーネット P.W.J.
社会的行動の神経生物学におけるマイクロバイオームの役割。
Biol. Rev. Camb. Philos. 2020; 95: 1131-1166
https://doi.org/10.1111/brv.12603
論文で見る
スコープス (61)
パブコメ
クロス
グーグル奨学生
バルセニーリャA.
プライドS.E.
マーティンJ.C.
ダンカン S.H.
スチュワート C.S.
ヘンダーソン C.
フリント H.J.
ヒト腸内酪酸産生菌の系統関係。
Appl. Microbiol. 2000; 66: 1654-1661
https://doi.org/10.1128/AEM.66.4.1654-1661.2000
論文で見る
スコープス (740)
PubMed
クロス
グーグル奨学生
ルイス P.
フリント H.J.
ヒト大腸内酪酸産生菌の多様性、代謝および微生物生態学的研究
FEMS Microbiol. Lett. 2009; 294: 1-8
https://doi.org/10.1111/j.1574-6968.2009.01514.x
論文で見る
酪酸菌
PubMed
クロス
グーグル奨学生
能登A.
ファノスV.
バルベリーニ L.
グラポフD.
ファットゥオーニ C.
ザファネッロ M.
カサノバ A.
フェヌ G.
デ・ジャコモ A.
デ・アンジェリスM.
他。
イタリアの自閉症児集団とその非罹患兄弟姉妹の尿中メタボロミクスプロファイル。
J. Matern. Fetal Neonatal Med. 2014; 27: 46-52
https://doi.org/10.3109/14767058.2014.954784
記事で見る
スコープス (89)
PubMed
クロス
グーグル奨学生
Liu Z.
Li W.
Geng L.
Sun L.
Wang Q.
Yu Y.
Yan P.
Liang C.
Ren J.
Song M.
他。
種を超えたメタボローム解析により、ウリジンは強力な再生促進因子であることが明らかになった。
Cell Discov. 2022; 8: 6
https://doi.org/10.1038/s41421-021-00361-3
論文で見る
スコープス (47)
PubMed
クロス
グーグル奨学生
Jiang N.
Zhao Z.
腸の老化は、in vivoおよびin vitroで炎症と酸化ストレスを制御するウリジンによって緩和される。
Cell Cycle. 2022; 21: 1519-1531
https://doi.org/10.1080/15384101.2022.2055252
論文で見る
スコパス (7)
PubMed
クロス
グーグル奨学生
Ma B.
Liang J.
Dai M.
Wang J.
Luo J.
Zhang Z.
Jing J.
中国の自閉症スペクトラム児における腸内細菌叢の変化。
Front. Cell. Infect. Microbiol. 2019; 9: 40
https://doi.org/10.3389/fcimb.2019.00040
論文で見る
スコープス (113)
PubMed
クロス
グーグル奨学生
マルティネス I.
ミュラーC.E.
ウォルターJ.
ヒト糞便微生物叢の長期的時間分析により、優勢な細菌種の安定したコアが明らかになった。
PLoS One. 2013; 8e69621
https://doi.org/10.1371/journal.pone.0069621
記事で見る
スコープス (131)
クロスリファレンス
グーグル奨学生
クラウス S.P.
エレーロ S.L.
ベルガーB.
クラウスL.
ブルッティンA.
モリーナ J.
パリス A.
ワント E.J.
ド・ワジエI.
クロアレックO.
他。
Colonization-Induced Host-Gut Microbial Metabolic Interaction.
mBio. 2011; 2e00271-10
https://doi.org/10.1128/mBio.00271-10
論文で見る
スコープス (327)
クロスリファレンス
グーグル奨学生
ジョンソン S.L.
カーク R.D.
ダシルバ N.A.
マー・H.
シーラム N.P.
バーティン M.J.
ポリフェノール微生物代謝産物は、腸および血液脳関門の透過性を示し、LPS誘導性炎症からマウスのミクログリアを保護する。
Metabolites. 2019; 9: 78
https://doi.org/10.3390/metabo9040078
論文で見る
スコープス (59)
PubMed
クロス
グーグル奨学生
ジョンソン S.L.
パーク H.Y.
ヴァテムD.A.
グランマスP.
マ H.
シーラム N.P.
食餌性イソフラボンダイゼインの血液脳関門透過性腸内微生物代謝産物であるエクオールは、ヒト神経芽細胞腫SH-SY5Y細胞および線虫において神経毒誘発毒性に対する神経保護作用を示す。
Plant Foods Hum. Nutr. 2020; 75: 512-517
https://doi.org/10.1007/s11130-020-00840-0
論文で見る
スコープス (20)
クロスリファレンス
グーグル奨学生
ヤップ C.X.
ヘンダース A.K.
アルバレス G.A.
ウッドD.L.A.
クラウス L.
タイソンG.W.
レストゥアディ R.
ウォレス L.
マクラーレンT.
ハンセル N.K.
他。
自閉症に関連した食事の嗜好が自閉症と腸内細菌叢の関連を媒介する。
Cell. 2021; 184: 5916-5931.e17
https://doi.org/10.1016/j.cell.2021.10.015
論文で見る
スコープス (151)
パブコメ
概要
全文
全文PDF
グーグル奨学生
ヤン K.
クルース R.L.
リン W.V.
Musher D.M.
肺感染症の原因としてのコリネバクテリア:症例シリーズと文献レビュー。
肺炎(ネイサン)。2018; 10: 10
https://doi.org/10.1186/s41479-018-0054-5
論文で見る
PubMed
クロスレフ
グーグル奨学生
諸石陽子
グイ J.
ホーエン A.G.
モリソン H.G.
ベイカー E.R.
ナドー K.C.
Li H.
リー Z.
マダン J.C.
カラガス M.R.
腸内細菌叢と新生児の呼吸器感染症リスクとの関係。
Commun. Med. 2022; 2: 1-8
https://doi.org/10.1038/s43856-022-00152-1
論文で見る
クロスリファレンス
グーグル奨学生
ライ D.
ゴールディング J.
マグヌッソンC.
ステア C.
ルイス G.
ダルマンC.
ストレスの多いライフイベントへの出生前および早期の曝露と自閉症スペクトラム障害のリスク: スウェーデンとイングランドの人口ベースの研究。
PLoS One。2012; 7e38893
https://doi.org/10.1371/journal.pone.0038893
記事で見る
スコープス (48)
クロスリファレンス
グーグル奨学生
カニナ A.
ラーション H.
シェランダー A.
ブトウィッカA.
テイラー M.J.
マルティーニ M.I.
リヒテンシュタイン P.
ルンドバーグ F.E.
オノフリオB.M.D.
ローゼンクヴィストM.A.
家族における累積的心理社会的逆境とADHDおよび自閉症との関連:家族ベースのコホート研究。
翻訳。精神医学。2023; 13: 1-8
https://doi.org/10.1038/s41398-023-02571-7
論文で見る
スコープス (0)
クロスリファレンス
グーグル奨学生
グムソグル S.B.
ファイン R.S.
マレー S.J.
バイトルJ.L.
スティーブンス H.E.
出生前ストレス後の神経発達におけるIL-6の役割。
脳行動。Immun. 2017; 65: 274-283
https://doi.org/10.1016/j.bbi.2017.05.015
論文で見る
スコープス (80)
PubMed
クロス
グーグル奨学生
コーカー M.O.
ホーエン A.G.
デイドE.
Lundgren S.
リーZ.
ウォン A.D.
ゼンス M.S.
パリス T.J.
モリソン H.G.
ソギン M.L.
他
分娩内抗生物質の特定のクラスと乳児の腸内細菌叢の成熟との関係:前向きコホート研究。
BJOG. 2020; 127: 217-227
https://doi.org/10.1111/1471-0528.15799
論文で見る
スコープス (76)
PubMed
クロス
グーグル奨学生
鄭D.
リウィンスキーT.
エリナブE.
健康と病気における微生物叢と免疫の相互作用。
Cell Res. 2020; 30: 492-506
https://doi.org/10.1038/s41422-020-0332-7
論文で見る
スコープス (1535)
PubMed
クロス
グーグル奨学生
オリバレス M.
ネフ A.
カスティーリョG.
パルマG.D.
バレア V.
カピージャ A.
パラウ F.
ノバ E.
マルコス A.
ポランコ I.
他。
HLA-DQ2遺伝子型は、セリアック病発症リスクの高い乳児の早期腸内細菌叢組成を選択する。
Gut. 2015; 64: 406-417
https://doi.org/10.1136/gutjnl-2014-306931
論文で見る
スコープス (227)
PubMed
クロス
グーグル奨学生
トーレス A.R.
ウェストオーバーJ.B.
ローゼンスパイアA.J.
自閉症におけるHLA免疫機能遺伝子。
Autism Res. 2012; 2012: 959073
https://doi.org/10.1155/2012/959073
論文で見る
パブコメ
クロス
グーグル奨学生
ウォーレン R.P.
オデルJ.D.
ウォーレンW.L.
バーガー・R.A.
マチューリスA.
ダニエルズ W.W.
トーレス A.R.
HLA-DRβ1の第3超可変領域と自閉症との強い関連。
J. Neuroimmunol. 1996; 67: 97-102
https://doi.org/10.1016/0165-5728(96)00052-5
論文で見る
スコープス (118)
パブコメ
概要
全文PDF
グーグル奨学生
ホール A.
Maw R.
イレズ=カヴェン Y.
グレゴリー S.
ライ D.
ゴールディングJ.
自閉的特徴と早期耳および上気道徴候との関連:Avon Longitudinal Study of Parents and Children (ALSPAC)の地理的に定義された小児集団の前向き観察研究。
BMJ Open. 2023; 13e067682
https://doi.org/10.1136/bmjopen-2022-067682
記事で見る
スコープス (2)
クロスリファレンス
グーグル奨学生
ペペレル C.
クスJ.V.
ガーダム M.A.
ヒューマーA.
バローズL.L.
複数の抗菌薬に対する耐性をコードする低病原性シトロバクター種。
Antimicrob. Agents Chemother. 2002; 46: 3555-3560
https://doi.org/10.1128/AAC.46.11.3555-3560.2002
論文で見る
スコープス (80)
PubMed
クロス
グーグル奨学生
リウ X.
Lu R.
Wu S.
Sun J.
Wnt/β-カテニン経路を介したサルモネラ菌による腸管幹細胞の制御。
FEBS Lett.
https://doi.org/10.1016/j.febslet.2010.01.024
論文で見る
スコープス (97)
PubMed
クロス
グーグル奨学生
リウ X.
Lu R.
ウー S.
Zhang Y.G.
Xia Y.
Sartor R.B.
Sun J.
Wnt2は腸上皮細胞における腸内細菌誘発性炎症を抑制する。
Inflamm. Bowel Dis. 2012; 18: 418-429
https://doi.org/10.1002/ibd.21788
論文で見る
スコープス (39)
PubMed
クロス
グーグル奨学生
カン E.
Zhou G.
Yousefi M.
Cayrol R.
Xia J.
Gruenheid S.
Citrobacter rodentium感染による疾患耐性の喪失は、上皮分化の障害およびT細胞応答の過活性化と関連している。
Sci. Rep. 2018; 8: 847
https://doi.org/10.1038/s41598-017-17386-y
論文で見る
スコープス (14)
クロスリファレンス
グーグル奨学生
ウマル S.
シトロバクター感染とWntシグナル。
Curr. Colorectal Cancer Rep.
https://doi.org/10.1007/s11888-012-0143-4
論文で見る
スコープス (16)
クロスリファレンス
グーグル奨学生
チェン・W.X.
チェン Y.R.
Peng M.Z.
Liu X.
Cai Y.N.
Huang Z.F.
Yang S.Y.
Huang J.Y.
Wang R.H.
Yi P.
他。
中国南部における自閉症スペクトラム障害児の血漿アミノ酸プロファイル: 110例の分析。
J. Autism Dev. Disord. 2023; 888
https://doi.org/10.1007/s10803-022-05829-z
論文で見る
スコープス (5)
クロスリファレンス
グーグル奨学生
Liu A.
Zhou W.
Qu L.
He F.
Wang H.
Wang Y.
Cai C.
Li X.
Zhou W.
Wang M.
自閉症スペクトラム障害児における尿中アミノ酸の変化。
Front. Cell. Neurosci. 2019; 13: 7
https://doi.org/10.3389/fncel.2019.00007
論文で見る
スコープス (36)
PubMed
クロス
グーグル奨学生
Zhang B.
Lv Z.
Li Z.
Wang W.
Li G.
Guo Y.
腸内細菌叢の変化と腸管傷害を軽減するl-アルギニンサプリメント。
Front. Microbiol. 2018; 9: 1716
https://doi.org/10.3389/fmicb.2018.01716
論文で見る
スコープス (63)
クロスリファレンス
グーグル奨学生
ファン・デル・ショール S.R.D.
リーズ P.J.
ステラード F.
ワティメナ J.D.L.
ザウアー P.J.J.
ビュラー H.A.
ヴァン・グードーヴァーJ.B.
早産児のリジンキネティクス:経腸栄養の重要性。
腸。2004; 53: 38-43
https://doi.org/10.1136/gut.53.1.38
記事で見る
スコパス (31)
PubMed
クロス
グーグル奨学生
ハレンA.
ジェイミーJ.F.
クーパーA.J.L.
哺乳類の脳におけるリジン代謝:最近の発見の重要性に関する最新情報。
アミノ酸。2013; 45: 1249-1272
https://doi.org/10.1007/s00726-013-1590-1
記事で見る
スコパス (101)
パブコメ
クロス
グーグル奨学生
ウー・G.
イェーガーL.A.
ベイザーF.W.
ロードスJ.M.
早産児のアルギニン欠乏:生化学的メカニズムと栄養の意味。
J. 栄養生化学。2004; 15: 442-451
https://doi.org/10.1016/j.jnutbio.2003.11.010
記事で見る
スコープス (197)
PubMed
クロス
グーグル奨学生
パク J.
Lee K.
キム K.
Yi S.J.
ヒストン修飾の役割:神経発達から神経疾患まで。
シグナル伝達。ターゲット。Ther. 2022; 7: 217
https://doi.org/10.1038/s41392-022-01078-9
論文で見る
スコープス (59)
パブコメ
クロス
グーグル奨学生
バドゥルディーン S.
ムロンゴ M.
バークリーJ.A.
アルギニン欠乏は早産新生児における重篤な感染症への感受性を高める。
Pediatr. Res. 2015; 77: 290-297
https://doi.org/10.1038/pr.2014.177
論文で見る
スコープス (47)
PubMed
クロス
グーグル奨学生
ポリカルプー E.
ザチャキ S.
ツォリア M.
パパエヴァンゲロウ V.
ポリカルプーN.
ブリアナ・D.
ガブリリ S.
コスタロス C.
Kafetzis D.
超低出生体重新生児における壊死性腸炎予防のためのL-アルギニン経腸的補充。
JPEN J. Parenter. Enteral Nutr.
https://doi.org/10.1177/0148607112471561
論文で見る
スコパス (48)
PubMed
クロス
グーグル奨学生
アミンH.J.
ザモラ S.A.
マクミランD.D.
フィック G.H.
ブッツナー J.D.
パーソンズ H.G.
スコット R.B.
アルギニン補充は未熟児における壊死性腸炎を予防する。
J. Pediatr: 425-431
https://doi.org/10.1067/mpd.2002.123289
論文で見る
スコープス (225)
パブコメ
概要
全文
全文PDF
グーグル奨学生
ニイニスト S.
タッキネン H.M.
エルルンド I.
アホネン S.
トッパリ J.
イロネン J.
ヴェイヨラ R.
クニップ M.
ヴァアララ O.
ヴィルタネン S.M.
乳児期の脂肪酸の状態は、1型糖尿病関連自己免疫のリスクと関連している。
Diabetologia. 2017; 60: 1223-1233
https://doi.org/10.1007/s00125-017-4280-9
論文で見る
スコープス (51)
PubMed
クロス
グーグル奨学生
ボーリューJ.
コスタG.
ルノーJ.
モワティエ A.
グレメ H.
セルジ D.
マルティノリ M.-G.
ミクログリア細胞およびミクログリア-ニューロン共培養において、パルミチン酸の神経炎症性および神経毒性はオレイン酸によって軽減される。
Mol. Neurobiol. 2021; 58: 3000-3014
https://doi.org/10.1007/s12035-021-02328-7
論文で見る
スコープス (17)
クロスリファレンス
グーグル奨学生
ラスケ C.
ミュラーS.
プライシェO.
ルシル V.
ムンク M.H.J.
オノルド I.
ペーター S.
ショップマイヤー U.
ウィルマン M.
腸内マイクロバイオームにおけるアルツハイマー病のシグネチャー: AlzBiom Studyの結果。
Front. Neurosci. 2022; 16: 792996
https://doi.org/10.3389/fnins.2022.792996
論文で見る
スコープス (9)
クロスリファレンス
グーグル奨学生
デ・フィリッピス F.
パゾッリE.
テット A.
タラッロ S.
ナッカラーティ A.
デ・アンジェリス M.
ネヴィアーニ E.
ココリン L.
ゴベッティ M.
セガタN.
他
ヒト腸内プレボテラ・コプリ株の異なる遺伝的および機能的形質は、異なる習慣的食事に関連している。
Cell Host Microbe. 2019; 25: 444-453.e3
https://doi.org/10.1016/j.chom.2019.01.004
記事で見る
スコープス (197)
PubMed
要旨
全文
全文PDF
グーグル奨学生
ベネット M.J.
ラグニ M.C.
フード I.
ヘイル D.E.
アゼライン酸とピメリン酸:代謝中間体または人工物?
J. 遺伝。Metab。Dis. 1992; 15: 220-223
https://doi.org/10.1007/BF01799635
論文で見る
スコープス (9)
クロス
グーグル奨学生
カシオ・L.
チェン・C.F.
ポーリー R.
スリカンス S.
ジョーンズ K.
スキナー C.D.
スティーブンソン R.E.
シュワルツ C.E.
Boccuto L.
大アミノ酸トランスポーターをコードする遺伝子の異常は自閉症スペクトラム障害のリスクを増加させる。
Mol. Genet. Genomic Med. 2020; 8e1036
https://doi.org/10.1002/mgg3.1036
論文で見る
スコープス (38)
クロスリファレンス
グーグル奨学生
アダムスJ.B.
オーディア・T.
マクドノー=ミーンズ S.
ルービン R.A.
クィグ D.
ガイス E.
ゲーン E.
ロレスト M.
ミッチェル J.
アトウッドS.
ら。
自閉症対神経質児の栄養と代謝の状態、および自閉症の重症度との関連。
栄養。Metab。2011; 8: 34
https://doi.org/10.1186/1743-7075-8-34
記事で見る
スコープス (287)
PubMed
クロス
グーグル奨学生
シャンバーガーR.J.
栄養とWICプログラムに関連する自閉症率。
J. J. Am. Coll。30: 348-353
https://doi.org/10.1080/07315724.2011.10719978
論文で見る
スコープス (20)
PubMed
クロス
グーグル奨学生
パワーズH.J.
リボフラビン(ビタミンB-2)と健康。
Am. J. Clin. Nutr. 2003; 77: 1352-1360
https://doi.org/10.1093/ajcn/77.6.1352
記事で見る
スコープス (721)
PubMed
要旨
全文
全文PDF
グーグル奨学生
マラシュリーE.T.
ボーレガ S.A.
リボフラビンには神経保護作用がある: パーキンソン病と片頭痛を中心に。
フロント。Neurol. 2017; 8: 333
https://doi.org/10.3389/fneur.2017.00333
論文で見る
スコープス (67)
PubMed
クロス
グーグル奨学生
ジャレット・H.
マクナルティ H.
ヒューズ C.F.
ペンティエヴァ K.
ストレイン J.J.
マッキャン A.
マカネナ L.
カニンガム C.
モロイ A.M.
フリンA.
ら。
18-102歳の成人におけるビタミンB-6とリボフラビン、それらの代謝的相互作用、およびMTHFR遺伝子型との関係。
Am. J. Clin. 2022; 116: 1767-1778
https://doi.org/10.1093/ajcn/nqac240
論文で見る
スコープス (5)
パブコメ
要旨
全文
全文PDF
グーグル奨学生
ラッセル J.T.
ロッシュ L.F.W.
エルドベリ M.
イロネン J.
アトキンソン M.A.
シャッツ D.A.
トリプレット E.W.
ルドヴィグソンJ.
自己免疫の遺伝的リスクは、ヒト腸内細菌叢の明確な変化と関連している。
Nat. Commun. 2019; 10: 3621
https://doi.org/10.1038/s41467-019-11460-x
論文で見る
スコープス (118)
パブコメ
クロス
グーグル奨学生
ルドヴィグソンJ.
ルドヴィグソンM.
セパA.
一般小児集団における糖尿病前症のスクリーニング:参加する母親の態度。
小児科。糖尿病。2002; 2: 170-174
https://doi.org/10.1034/j.1399-5448.2001.20405.x
記事で見る
スコープス (44)
クロス
グーグル奨学生
キンドグレンE.
アーレンスA.P.
トリプレットE.W.
Ludvigsson J.
乳幼児の腸内細菌叢と環境は、若年性特発性関節炎発症の何年も前から、特に遺伝的に脆弱な小児において関連する。
EBioMedicine. 2023; 93: 104654
https://doi.org/10.1016/j.ebiom.2023.104654
論文で見る
スコープ (3)
要旨
全文
全文PDF
グーグル奨学生
ルドヴィグソン J.F.
アンデルソン E.
エクボム A.
フェイヒティング M.
キム J.L.
ロイターウォール C.
Heurgren M.
オラウソンP.O.
スウェーデン全国入院患者登録の外部レビューと検証。
BMC Public Health. 2011; 11: 450
https://doi.org/10.1186/1471-2458-11-450
論文で見る
スウェーデン
PubMed
クロス
グーグル奨学生
ベル C.C.
DSM-IV:精神障害の診断と統計マニュアル。
JAMA。1994; 272: 828-829
https://doi.org/10.1001/jama.1994.03520100096046
記事で見る
クロスリファレンス
グーグル
アメリカ精神医学会
精神障害の診断と統計マニュアル。
第5版。米国精神医学会、2013年
https://doi.org/10.1176/appi.books.9780890425596
記事で見る
クロスリファレンス
グーグル・スカラー
リービッシュ G.
フェイヒー E.
青木 J.
デニス E.A.
デュラン T.
エシング C.S.
フェドロワ M.
フォイスナー I.
グリフィス W.J.
ケフェラーH.
他。
LIPID MAPSの分類、命名法、MS由来の脂質構造の略記法に関する最新情報。
J. Lipid Res. 2020; 61: 1539-1555
https://doi.org/10.1194/jlr.S120001025
論文で見る
スコープス (333)
パブコメ
要旨
全文
全文PDF
グーグル・スカラー
プラスカルT.
カスティーヨ S.
ビジャール・ブリオネスA.
Oresic M.
MZmine 2: 質量分析ベースの分子プロファイルデータの処理、可視化、解析のためのモジュラーフレームワーク。
BMC Bioinformatics. 2010; 11: 395
https://doi.org/10.1186/1471-2105-11-395
論文で見る
筑波大学
PubMed
クロス
グーグル奨学生
カポラソJ.G.
クチンスキーJ.
ストンボーJ.
ビッティンガー K.
ブッシュマン F.D.
コステロ E.K.
フィーラー N.
ペーニャ A.G.
グッドリッチ J.K.
ゴードン J.I.
et al.
高スループットコミュニティシーケンスデータの解析を可能にするQIIME。
Nat. Methods. 2010; 7: 335-336
https://doi.org/10.1038/nmeth.f.303
論文で見る
論文リスト(27156)
パブコメ
クロス
グーグル奨学生
キャラハン B.J.
マクマーディ P.J.
ローゼン M.J.
ハン A.W.
ジョンソン A.J.A.
ホームズ S.P.
DADA2:イルミナアンプリコンデータからの高分解能サンプル推定。
Nat. Methods. 2016; 13: 581-583
https://doi.org/10.1038/nmeth.3869
論文で見る
スコープス (14940)
PubMed
クロス
グーグル奨学生
クアスト C.
プルエッセE.
Yilmaz P.
ゲルケンJ.
シュヴェールT.
ヤルザ P.
ペプリーズ J.
Glöckner F.O.
SILVAリボソームRNA遺伝子データベースプロジェクト:改良されたデータ処理とウェブベースのツール。
Nucleic Acids Res.
https://doi.org/10.1093/nar/gks1219
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
アルトシュール S.F.
ギッシュW.
ミラーW.
マイヤーズ E.W.
リップマン D.J.
基本的なローカルアライメント検索ツール。
J. Mol. Biol. 1990; 215: 403-410
https://doi.org/10.1016/S0022-2836(05)80360-2
論文で見る
日本学術振興会特別研究員
PubMed
クロス
グーグル奨学生
チェンバレン C.A.
ハッチM.
ギャレット T.J.
UHPLC-HRMSによるオキサロバクター・フォルミゲネス(Oxalobacter formigenes)HC1株およびOxWR株のメタボロームおよびリピドミクス特性解析。
Anal. Bioanal. Chem. 2019; 411: 4807-4818
https://doi.org/10.1007/s00216-019-01639-y
論文で見る
スコープス(17)
クロス
グーグル奨学生
ケルメルJ.P.
クルーガーN.M.
ウルマー C.Z.
ボーデン J.A.
パターソン R.E.
コクラン J.A.
ビーチャー C.W.W.
ギャレット T.J.
Yost R.A.
LipidMatch:非標的高分解能タンデム質量分析データを用いたルールベースの脂質同定のための自動ワークフロー。
BMC Bioinformatics. 2017; 18: 331
https://doi.org/10.1186/s12859-017-1744-3
論文で見る
スコープス (221)
PubMed
クロス
グーグル奨学生
オクサネン J.
シンプソンG.L.
ブランシェF.G.
キント R.
レジャンドル P.
ミンチン P.R.
オハラ R.B.
ソリモスP.
スティーブンス M.H.H.
Szoecs E.
他
ビーガン: 群集生態学パッケージ。
(バージョン2.6-4)
https://cran.r-project.org/web/packages/vegan/vegan.pdf
日付:2022年
記事で見る
グーグル・スカラー
ラブ M.I.
フーバー W.
Anders S.
DESeq2によるRNA-seqデータのフォルドチェンジと分散のモデレート推定。
Genome Biol.
https://doi.org/10.1186/s13059-014-0550-8
論文で見る
論文概要
PubMed
クロス
グーグル奨学生
ベンジャミニ Y.
ホッホバーグ Y.
誤発見率のコントロール: 多重検定への実践的で強力なアプローチ。
J. R. Stat. Soc. B Methodol. 1995; 57: 289-300
https://doi.org/10.1111/j.2517-6161.1995.tb02031.x
論文で見る
クロス
グーグル・スカラー
ロッシュ L.F.W.
ドブラー P.T.
パイロ V.S.
コラツコフスキーB.
ドリュー J.C.
トリプレット E.W.
pime: 微生物群集間の新規差異発見のためのパッケージ。
Mol. Ecol. Resour. 2020; 20: 415-428
https://doi.org/10.1111/1755-0998.13116
論文で見る
スコープス (29)
パブコメ
クロス
グーグル奨学生
パン Z.
Chong J.
Zhou G.
デ・リマ・モライス D.A.
Chang L.
バレット M.
ゴーティエ C.
ジャック P.É.
Li S.
Xia J.
MetaboAnalyst 5.0:生のスペクトルと機能的洞察のギャップを縮める。
Nucleic Acids Res. 2021; 49: W388-W396
https://doi.org/10.1093/nar/gkab382
論文で見る
スコープス (1908)
PubMed
クロス
グーグル奨学生
Pang Z.
Zhou G.
Ewald J.
チャン L.
ハカリス O.
Basu N.
Xia J.
グローバルメタボロミクスデータのLC-HRMSスペクトル処理、マルチオミクス統合、共変量調整にMetaboAnalyst 5.0を使用。
Nat. Protoc. 2022; 17: 1735-1761
https://doi.org/10.1038/s41596-022-00710-w
論文で見る
スコープス (460)
PubMed
クロス
グーグル奨学生
論文情報
出版履歴
出版 2024年4月3日
受理 受理:2024年2月28日
改訂版受理 2023年11月22日
受理:2023年11月22日 受理日:2023年4月7日
出版段階
インプレス、ジャーナル予稿集
識別
DOI: https://doi.org/10.1016/j.cell.2024.02.035
著作権
エルゼビア社発行
ユーザーライセンス
クリエイティブ・コモンズ 表示 (CC BY 4.0)|情報アイコンの再利用方法
サイエンスダイレクト
ScienceDirectでこの論文にアクセスする
図
図サムネイルfx1
グラフィカルアブストラクト
図のサムネイルgr1
図1神経発達障害(NDs)に関与する早期生活環境および医学的危険因子
図のサムネイルgr2
図22.5歳および5歳時の症状クラスターと1歳時の微生物種存在量との関連
図サムネイルgr3
図3将来のASDにおけるコード血清メタボロームと便脂肪酸の違い(ABISASD)
図サムネイルgr4
図4微生物交絡因子とNDリスク因子のマッチング(ABISND-Match)の前後における、全ABISControls(n = 1,456)と将来のNDであるABISND(n = 116)の分類学的およびコアマイクロバイオームの違い
サムネイル図1
図S1図1に関連する、ADHD、言語障害、知的障害に関与する早期の生活環境および医学的危険因子
図サムネイル図2
図S2図2に関連する、2.5歳時の症状と1歳時の微生物量との関連
図サムネイルgr5
図5将来の神経発達障害(ND)サブグループ間およびHLAリスクハプロタイプ間の腸内細菌叢の違い
サムネイル図3
図S3図4および図5に関連する、対照群と神経発達障害(ND)にまたがるコア微生物叢
図サムネイルfigs4
図S4ABIS対照群ではすべてのNDカテゴリーと比較して腸内細菌叢が増加している(図5に関連)。
図サムネイルgr6
図6便代謝物は将来のASD、および1年後のAkkermansiaとCoprococcusの存在量と強い相関がある
サムネイル図5
図S5将来の自閉症スペクトラム障害(ASD)における診断年齢による腸内細菌叢の違い(N=39)、図5関連
図サムネイルgr7
図7シトロバクター(Citrobacter)およびコプロコッカス(Coprococcus)の有病率、およびファスコラクトバクテリウム(Phascolarctobacterium)の存在量と、早期の耳炎感染および将来の神経発達障害(ND)の転帰との関連性
関連記事
広告
ライフ&メディカルサイエンスジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノム学
細胞宿主と微生物
細胞代謝
細胞レポート
セルレポーツ医学
細胞幹細胞
細胞システム
カレントバイオロジー
発生細胞
免疫
医学
分子細胞
ニューロン
構造
アメリカン・ジャーナル・オブ・ヒューマン・ジェネティクス (パートナー)
生物物理学雑誌 (パートナー)
生物物理レポート (パートナー)
Human Genetics and Genomics Advances (パートナー)
分子植物 (パートナー)
分子療法 (パートナー)
分子治療法と臨床開発 (パートナー)
分子療法核酸 (パートナー)
分子療法腫瘍学 (パートナー)
Plant Communications (パートナー)
幹細胞レポート (パートナー)
生化学の動向
癌の動向
細胞生物学の動向
生態学と進化の動向
内分泌学・代謝学の動向
遺伝学の動向
免疫学の動向
微生物学の動向
分子医学の動向
神経科学の動向
寄生虫学の動向
薬理学の動向
植物科学の動向
物理科学・工学ジャーナル
セルレポート 物理科学
化学
化学触媒
デバイス
ジュール
物質
化学の動向
学際ジャーナル
セルリポーツ・メソッド
セルリポーツ・サステイナビリティ
Heliyon
アイサイエンス
ワン・アース
パターン
STARプロトコル
ネクサス(パートナー)
イノベーション(パートナー)
バイオテクノロジーの動向
認知科学の動向
著者
論文投稿
複数ジャーナル投稿
STARメソッド
プレビュー - プレプリント
査読者
査読者向け情報
ニュース&イベント
ニュースルーム
細胞シンポジウム
コンソーシアムハブ
ウェビナー
ラボリンク
マルチメディア
セルプレスポッドキャスト
セルプレスビデオ
カラーリングとコミック
フィギュア360
セル画ショー
研究篇
セルプレスについて
セルプレスについて
オープンアクセス
COVIDハブ
持続可能性
インクルージョンと多様性
コンタクト
お問い合わせ
ヘルプ&サポート
採用情報
セルプレス採用情報
サイエンティフィックジョブボード
アクセス
登録する
請求
今すぐ読む
司書に推薦する
出版アラート
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
Nucleusコレクション
スナップショット・アーカイブ
インフォメーション
広告主の皆様へ
リクルーターの方へ
図書館員の方へ
プライバシーポリシー
ご利用条件
アクセシビリティ
本サイトのコンテンツは、あらゆる分野の医療従事者および研究者を対象としています。
当サイトでは、サービスの提供・向上およびコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を更新するには、このサイトのクッキー設定をご覧ください。
このサイトのすべてのコンテンツ Copyright © 2024 Elsevier Inc.、そのライセンサー、および寄稿者。
テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビア社に帰属します。
すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条件が適用されます。
プライバシーポリシー 利用規約 アクセシビリティ ヘルプ&サポート お問い合わせ
RELX
この記事が気に入ったらサポートをしてみませんか?