腸管上皮のHDAC3とMHCクラスIIは微生物特異的な免疫を調整する

研究論文GastroenterologyImmunology Open Access|10.1172/JCI162190
腸管上皮のHDAC3とMHCクラスIIは微生物特異的な免疫を調整する
https://www.jci.org/articles/view/162190#.Y-71_BrMv8k.twitter
エミリー・エシュルマン、1,2 シャオ・ツウ、2,3,4 ヴィヴィアン・ウー、1,2 テイラー・ライス、1,2 ローラ・エングルマン、1,2 ベイリー J. ディドリクセン、1,2,4 ジョーダン・ウィット、1,2 デイビッド・ベイ・ハスラム、3 シンシンウェイ、2,3 テレーザ・アレンハット 1,2人
2023年1月5日発行 - 詳細はこちら
PDFを見る
要旨
常在細菌に対する異常な免疫応答は、炎症性腸疾患やその他の慢性炎症疾患を促進する。しかし、粘膜組織において微生物特異的な免疫反応がどのように制御されているかは、まだ十分に理解されていない。我々は、微生物特異的ヒストン脱アセチル化酵素3(HDAC3)を上皮に発現しないマウスが、腸内の常在菌特異的CD4+ T細胞の蓄積を増加させることを発見し、上皮のHDAC3が局所微生物特異的免疫に指示を与えるのではないかという仮説を引き起こした。これと一致して、微生物特異的CD4+T細胞と上皮HDAC3の発現は、早期の微生物叢のコロニー形成に伴って同時に誘導された。さらに、HDAC3の上皮内在性アブレーションは、腸内細菌特異的Tregを減少させ、腸内細菌特異的Th17細胞を増加させ、T細胞駆動性大腸炎を促進させることがわかった。HDAC3は、NF-κB依存的な上皮性MHCクラスII(MHCII)の制御に不可欠であることが明らかになった。上皮内在性のMHCIIは、成体マウスにおける宿主特異的Th17細胞の局所的蓄積を抑制し、微生物が誘発する炎症から保護した。さらに、HDAC3は、腸内細菌が上皮細胞にMHCIIを発現させ、腸内細菌特異的T細胞の数を制限することを可能にした。これらのデータを総合すると、常在菌を認識し炎症を制御する組織内在性のCD4+T細胞サブセットの動的バランスを指令する上で、上皮のヒストン脱アセチル化酵素が中心的な役割を担っていることが明らかになった。
グラフィカルアブストラクト

グラフィカルアブストラクト
はじめに
消化管には、微生物叢と総称される何兆もの微生物が生息しており、それらは哺乳類細胞と共生関係を形成し、健康や疾病の媒介に重要な役割を担っている。広域抗生物質や無菌マウスモデルを用いた広範な実験により、宿主の免疫系の発達と機能において微生物叢が必要であることが明らかにされている(1-5)。特に、生後数年の間に重要な免疫教育や較正が行われるため、微生物叢の相互作用は生後間もない時期に大きな影響を与える(6-9)。実際、この生後間もない時期に微生物叢の組成やコロニー形成が乱れると、喘息、アレルギー、炎症性腸疾患(IBD)などの慢性炎症性疾患を発症しやすくなる免疫の長期的な変化と関連している(1, 4, 8)。
宿主と微生物叢の関係は共生的であるにもかかわらず、粘膜表面に豊富に存在し、抗原的に外来微生物と密接に関連しているため、病的な炎症を刺激する潜在的な危険性を持っている。そのため、腸管免疫応答は、侵入してくる病原体に対する防御的な免疫を可能にする一方で、無害な常在菌に対する炎症反応を抑制するよう、厳密に制御されなければならない。常在菌は制御性T細胞の分化を促進し(10)、微生物反応性エフェクターおよびメモリーT細胞は、マウスおよびヒトの両方に存在する(11-17)。これらの常在菌特異的T細胞は、防御サイトカインを誘導し、病原体に対する交差反応性を提供することにより、バリア機能を促進する(14, 18, 19)。しかし、微生物叢に対する異常な免疫応答は、IBDのような炎症状態の引き金にもなる(1, 20-22)。大腸炎モデルマウスでは、腸内細菌は、部分的には、微生物叢反応性CD4+ T細胞を刺激することによって、炎症を駆動する(23-25)。さらに、IBD患者の微生物特異的CD4+ T細胞は、健常患者のT細胞と比較して、機能的に変化し、IL-17などの炎症性サイトカインをより多く産生することが示されている(14, 26-30)。しかし、腸内細菌特異的CD4+ T細胞の組織内制御を指示するメカニズムは、まだ十分に理解されていない。
腸管上皮細胞(IEC)は、微生物叢と免疫細胞の直接的な界面に存在するため、微生物抗原に応答して局所免疫を指示するユニークな態勢をとっている。IECは病原体認識受容体を発現し、微生物のシグナルを感知して、サイトカイン、ケモカイン、成長因子の分泌を介して腸管免疫応答を制御する(1, 31)。上皮に存在する特殊な小葉細胞や杯細胞関連抗原通路は、下層の抗原提示骨髄系細胞に抗原を送達するが(32-34)、IECは、免疫応答を制御しうる古典的抗原処理・提示経路も備えている(35、36)。しかしながら、健全な微生物-免疫関係を調整するIEC指向のメカニズムについては、まだあまり理解されていない。
標準的なセンサー以外にも、エピジェネティック修飾酵素であるヒストン脱アセチル化酵素3(HDAC3)の上皮発現が微生物叢に応答し、哺乳類の代謝、腸の恒常性、炎症を制御することが最近わかってきた(37-40)。ここで、我々は、最初の微生物叢のコロニー化後の微生物叢依存的な常在特異的CD4+ T細胞の拡大が、HDAC3の上皮の発現と同時に起こることを見いだした。この関係を考慮して、我々はIEC内在性のHDAC3が、微生物叢に刺激されたCD4+ T細胞の拡大および分化を制御しているかどうかを調べた。実際、IECにおけるHDAC3発現の消失は、腸の炎症と、上皮のMHCIIによって誘導される微生物特異的Th17細胞の蓄積を引き起こした。無菌状態で飼育したマウスの組織をさらに分析したところ、HDAC3は、微生物が上皮MHCII依存的に微生物特異的CD4+ T細胞の制御を誘導するのに必要であることが示された。これらのデータを総合すると、微生物叢は、組織内在性T細胞の上皮制御を指示することにより、常在耐性を誘導し、炎症を制限していることが明らかになった。
研究成果
上皮のHDAC3発現は、腸の通性CD4+ T細胞を制限する。微生物叢のコロニー形成は出生時に始まり、微生物叢の複雑さと密度は乳児期に最も顕著に増加する(41-43)。幼少期から始まる微生物叢への曝露は免疫学的教育を促進し、この時期に破壊されると慢性炎症性疾患が増加する可能性がある(42-46)。出生時の初期コロニー形成が、常在菌に特異性を持つ局所T細胞の蓄積を促すことが予測されているが、これは直接的に検証されたことはない。そこで、生後間もない微生物叢のコロニー形成が、腸の常在菌特異的T細胞応答を誘導しているかどうかを調べるために、無菌仔(GF)と通常飼育(CNV)の新生児の大腸からのCD4+ T細胞を、まずMHCII制限四量体を用いて常在フラゲcBir1ペプチドに対するT細胞受容体特異性を分析した。生後1週間以内に、GFとCNVの仔マウスは、大腸におけるcBir1+CD4+T細胞の存在量が同様に少なかった(図1、AおよびB)。比較として、微生物存在下で飼育された3週齢の仔は、年齢をマッチさせたGF対照と比較して、cBir1+CD4+T細胞の蓄積の増加を示し(図1、AおよびB)、これは離乳までに微生物叢コロニー化が徐々に増加することと並行している(44)。したがって、微生物叢由来のシグナルは、腸内微生物特異的CD4+T細胞の初期膨張のプライミングに不可欠である。
上皮性HDAC3発現が腸内常在菌特異的CD4+ T細胞を制限している図1
上皮性HDAC3発現は、腸内常在菌特異的CD4+ T細胞を制限する。(A と B) GFとCNVの新生仔の大腸から分離したcBir1+テトラマー特異的CD4+ T細胞の数。 (C) GFとCNV仔の大腸から分離したIECにおけるHDAC3のmRNA発現。 (D と E) 全腸CD3+の頻度。(FおよびG)全腸CD8a+(F)およびCD4+(G)T細胞の頻度。(H-K)HDAC3FFおよびHDAC3ΔIECマウスの大腸におけるcBir1+テトラマー特異的CD4+ T細胞の数(HおよびI)およびRORγt+(J)およびFoxP3+(K) cBir1+CD4+ T細胞の頻度。 cBir1+テトラマー細胞はライブ、CD45+、系統(CD11b-B220-Ly6G-、CD11c-CD8a-、CD4+でゲートされる。データは、少なくとも2回の実験、1グループあたり3-4匹のマウスの代表値である。*P < 0.05, **P < 0.01, ****P < 0.0001, 1-way ANOVA with Tukey's multiple-comparison test (B) or unpaired 2-tailed Student's t test (C-K)による.
このことは、これらのT細胞サブセットを制御する経路の障害が、病的な炎症への感受性に影響を与える可能性があることを示唆している。興味深いことに、IBD患者のIECはHDAC3酵素のレベルを低下させており(37)、他の施設での知見と一致して、HDAC3のIEC固有発現を欠くマウス(HDAC3ΔIECマウス)は直腸脱を特徴とする慢性腸炎に対する感受性を増加させていた(補足図1A;この記事とともにオンラインで利用できる補足資料;https://doi. org/10.1172/JCI162190DS1) 、炎症性バイオマーカーのリポカリン-2のレベル上昇(補足図1B)、炎症細胞の浸潤(補足図1、C-E)、慢性炎症に一致する組織学的変化(補足図1F)(37)。驚くべきことに、HDAC3の上皮発現も1週齢の仔と比較して3週齢の仔で劇的に誘導された(図1C)。しかし、GF条件下で飼育した仔マウスでは、これらの発生段階において上皮のHDAC3発現は低いバックグラウンドレベルに留まっていた(図1C)。このことは、マイクロバイオータのコロニー形成が腸の初期段階の上皮HDAC3発現を増加させることを示唆している。
上皮内在性HDAC3発現と微生物叢コロニー化後の通性CD4+T細胞の拡大との間の時間的関連は、IEC内在性HDAC3が微生物叢特異的T細胞免疫を制御しているのではないかという仮説を引き起こした。腸管固有層では、HDAC3ΔIECマウスでは、Cre陰性同腹子HDAC3FF対照と比較して、総CD3+ T細胞は変化していなかった(図1、DおよびE)。しかしながら、上皮HDAC3の喪失は、CD8a+細胞ではなく、腸CD4+ T細胞の上昇をもたらし(図1、FおよびG)、それは大腸(図1、HおよびI)および小腸(補足図2、AおよびB)において増加したcBir1+通性特異性を呈した。メタゲノム解析では、HDAC3ΔIECマウスの大腸における常在菌組成の変化が確認され(補足図3A)、多様性の減少(補足図3B)およびビフィズス菌の減少(補足図3C)により特徴づけられ、先行する16S配列決定(37)と一致した。さらに、HDAC3ΔIECマウスの腸内細菌叢では、cBir1の発現自体が、同腹のコントロールと比較して減少していた(補足図3、DおよびE)。したがって、HDAC3ΔIECマウスにおけるcBir1-同胞特異的T細胞の上昇は、cBir1発現種のレベルと並行していない。HDAC3ΔIECマウスのcBir1+ CD4+T細胞は、炎症性RORγt+ Th17細胞への分化が増加し(図1J)、フロックス化コントロールと比較して、相互的にFoxP3+ T制御分化が減少している(図1K)ことが示された。コメンサル特異的なTh1およびT濾胞ヘルパー(Tfh)細胞は、上皮HDAC3の喪失によって変化しなかった(補足図4A)。これらのデータを総合すると、上皮HDAC3は、腸内の通性特異的TregとTh17細胞のバランスを調節するのに重要な役割を果たすことが示される。
上皮性HDAC3欠損マウスのCD4+ T細胞は、重篤な大腸炎を誘発する。HDAC3ΔIECマウスの制御不全CD4+ T細胞が腸の炎症を促進するかどうかを決定するために、慢性大腸炎のT細胞移植モデル(47-49)を用い、精製CD4+ T細胞をHDAC3FFおよびHDAC3ΔIECマウスから分離してRag1-/-受容体マウスに養子移入した(図2A)。コントロールマウスからCD4+ T細胞を受け取ったRag1-/-レシピエントとは対照的に、HDAC3ΔIECマウスからCD4+ T細胞を受け取ったRag1-/-レシピエントはより著しい体重減少(図2B)、結腸短縮(図2C)、および炎症細胞浸潤、クリプト過形成、および壁肥厚によって特徴づけられる重度の結腸炎病理を示した(図2、DおよびE)。さらに、HDAC3ΔIECマウスからCD4+T細胞を受け取ったRag1-/-マウスでは、炎症性バイオマーカーであるリポカリン-2の内腔濃度も有意に上昇した(図2F)。したがって、IEC内在性のHDAC3発現は、炎症性大腸菌CD4+ T細胞のプライミングを抑制するのに必須である。
上皮HDAC3欠損マウスのCD4+ T細胞は、重篤な大腸炎を誘発する.Figure 2
上皮性HDAC3欠損マウスのCD4+ T細胞は、重篤な大腸炎を誘発する。(A) HDAC3FFおよびHDAC3ΔIECマウスから分離したナイーブCD4+T細胞をRag1-/-宿主に移植したT細胞大腸炎モデルの実験概略図。(B-D)HDAC3FFまたはHDAC3ΔIECマウスからのT細胞を受け取ったRag1-/-ホストの体重(B)、結腸長(C)、およびH&E染色結腸切片(D)の変化。スケールバー:20μM。(E)Dの切片の組織学的スコアリング(F)リポカリン-2の糞便中濃度。(GおよびH)HDAC3FFまたはHDAC3ΔIECマウスからT細胞を受け取ったRag1-/-ホストの大腸におけるcBir1+四量体特異的CD4+ T細胞の数。生きている、CD45+、系統(CD11b-B220-Ly6G-、CD11c-CD8a-)、CD4+にゲーティングした。(I) cBir1+ CD4+T細胞のTh17 (RORγt+)の頻度。(J-L)HDAC3FFまたはHDAC3ΔIECマウスからのT細胞を受け取った水(CNV)または抗生物質(ABX)で処理したRag1-/-マウスの大腸における体重(J)、cBir1+四量体特異的CD4+ T細胞数(K)およびRORγt+ cBir1+ CD4+ T細胞(L)頻度の変化。データは少なくとも2回の独立した実験(1群3-4匹)の代表値である。*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, by unpaired 2-tailed Student's t test (B-I) or 1-way ANOVA with Tukey's multiple-comparison test (J-L).
HDAC3ΔIECマウスから分離した大腸菌CD4+ T細胞をさらに調べるために、常在細菌発現抗原に対する特異性を評価した。HDAC3ΔIECマウスからCD4+T細胞を受け取ったRag1-/-レシピエントは、HDAC3FF細胞を受け取ったホストと比較して、上昇した微生物叢特異的cBir1+CD4+T細胞を示した(図2、GおよびH)。これらの実験から、移植後のcBir1+細胞の大部分はRORγt+ Th17細胞であることが示された(図2Iおよび補足図4B)。HDAC3ΔIECマウスにおけるこれらの細胞の偏った分化と同様に、HDAC3ΔIECマウスに由来する場合、HDAC3FF同腹のコントロールと比較して、より高頻度に交感神経特異的T細胞が炎症性RORγt+ Th17細胞に分化した(図2I)。HDAC3ΔIECマウス由来のコメンサル特異的T細胞は、FoxP3+ Tregへの分化が減少し、Th1細胞には差がなかった(補足図4B)。次に、これらの細胞が微生物叢に応答しているかどうかを調べるために、広域スペクトル抗生物質を用いて常在菌を枯渇させることにより、T細胞移入試験を行った(50)。以前の結果と一致して、HDAC3ΔIECマウスからT細胞を受け取ったマイクロバイオータレプリエントマウスにおいて、体重減少の増加(図2J)およびcBir1+ Th17細胞の拡大が検出された(図2、KおよびL)。しかしながら、大腸炎による体重減少(図2J)およびcBir1+ Th17細胞は、マイクロバイオータ枯渇とともに失われ(図2、KおよびL)、このモデルにおいて、交雑物特異的Th17細胞が腸の炎症を駆動することが示された。集合的に、これらのデータは、上皮HDAC3の損失がマイクロバイオータ特異的なコリトジェニックCD4+ T細胞の増加をもたらしたので、上皮HDAC3の発現が腸におけるCD4+ T細胞の発達の制御に必要であることを示す。
HDAC3は、IEC上のMHCIIの表面発現を制御している。サイトカインIFN-γは、CD4+ T細胞のTh1エフェクターへの分化を促進する一方で、Th17細胞を含む他のTh系譜への分化を抑制する(51, 52)。以前の研究では、細菌感染時に、上皮内T細胞によるIFN-γ産生が、上皮HDAC3欠損マウスで減少することが示された(39)。しかし、定常状態ではIFN-γレベルは低く、有意差は検出されなかった(補足図4、CおよびD)(39)。また、1型自然免疫系リンパ球(ILC1s)はIFN-γを産生することができる。しかし、上皮性HDAC3の欠損は、ILC1sの頻度や、大腸で優勢なILC系譜であるILC3sには影響を与えなかった(補足図4E)。さらに、外因性IFN-γを投与されたHDAC3ΔIECマウスでは、通性特異的Th17細胞は上昇したままであった(補足図4F)。したがって、IFN-γは、HDAC3ΔIECマウスで観察された通性特異的Th17細胞の基礎的な違いの直接的または主要な原因を反映しているとは考えられない。
MHCIIを介した抗原提示は、抗原特異的CD4+T細胞応答を指示するのに重要である。しかし、全MHCII+腸造血細胞(図3A)およびMHCIIhi腸樹状細胞やマクロファージを含む古典的抗原提示細胞の頻度は、上皮HDAC3欠失によって影響を受けなかった(図3、BおよびC)。しかし驚くべきことに、CD45+造血細胞ではなく、EpCAM+IECが腸-微生物叢界面でMHCIIの大部分を発現していることがわかった(図3, DおよびE)。IECは微生物の合図に反応する。そこで次に、微生物叢がIEC内在性のMHCII発現を調節する役割を担っているかどうかを評価した。他の研究(53-57)と同様に、CNVマウスから分離したIECはGFマウスと比較してMHCIIの高い表面発現を示した(図3、FおよびG)。さらに、新生児GF仔は、MHCIIのβ鎖をコードする遺伝子であるH2-Ab1の上皮の発現を低く維持していた(図3H)。しかし、上皮のH2-Ab1発現は、HDAC3の制御(図1C)と同様に、最初の微生物叢のコロニー化後に強固に誘導された(図3H)。
HDAC3はIECs上のMHCIIの発現を制御する図3
HDAC3は、IECs上のMHCIIの発現を制御する。(A)結腸固有層における全MHCII+細胞の頻度。(BおよびC) HDAC3FFおよびHDAC3ΔIECマウスの大腸固有層における樹状細胞およびマクロファージの頻度。ライブ、CD45+、MHCII+でゲーティング。(D)大腸内腔表面のMHCII+細胞。(E)Dにおける全MHCII+細胞の頻度。(FおよびG)GFおよびCNVマウスの大腸におけるMHCII+ EpCAM+細胞の頻度。(H)GFおよびCNV仔マウスの大腸から分離したIECにおけるH2-Ab1のmRNA発現。 IおよびJ)HDAC3FFおよびHDAC3ΔIECマウスの大腸におけるMHCII+ EpCAM+細胞の頻度。(KおよびL)HDAC3FFおよびHDAC3ΔIECマウスの大腸から単離したIECにおけるH2-Ab1(K)およびCIITA(L)mRNA。(M)タモキシフェン(4-OHT)で処理したHDAC3FFおよびHDAC3ΔIEC-INDオルガノイドにおけるHDAC3 mRNA。 N)IKK-16で処理したHDAC3FFおよびHDAC3ΔIEC-INDオルガノイドにおけるH2-Ab1 mRNAの発現。データは、少なくとも3つの独立した実験、1群あたり3-5匹のマウスの代表値である。*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, by unpaired 2-tailed Student's t test (E-M) or 1-way ANOVA with Tukey's multiple-comparison test (N).
興味深いことに、HDAC3発現の喪失は、大腸(図3、IおよびJ)および小腸(補足図5、AおよびB)において上皮表面MHCII発現を劇的に減少させた。HDAC3ΔIECマウスのIECは、H2-Ab1遺伝子発現の低下を示し(図3Kおよび補足図5C)、MHCIIコード遺伝子の転写調節が変化していることが示唆された。HDAC3ΔIECマウスのIECでは、H2-Ab1遺伝子の近傍でヒストンアセチル化の有意な変化は見られなかった(補足図5D)ことから、HDAC3はこの遺伝子を直接標的としていないことが示唆された。クラスIIコアクチベータCIITAの発現は、HDAC3ΔIECマウス由来のIECにおいて、フロックス化対照由来のIECと比較して減少していた(図3L)。CIITAはNF-κBによって制御され、HDAC3は複数の細胞系においてNF-κB活性化を促進することが示されている(58-61)。したがって、NF-κBがMHCIIのHDAC3依存性調節を媒介するかどうかを試験するために、タモキシフェンがHDAC3発現を著しく低下させるHDAC3FFおよび誘導性HDAC3ΔIEC-INDマウスモデルの結腸から腸オルガノイドが生成された(図3M)。NF-κB活性の阻害は、野生型オルガノイドにおけるH2-Ab1の発現を抑制した(図3N)。しかし、NF-κBの阻害は、HDAC3を欠くオルガノイドにおけるMHCIIの発現に影響を与えなかった(図3N)。まとめると、これらのデータは、HDAC3が、部分的に、NF-κBの活性化を介して、上皮のMHCII発現を調節することを支持する。予想通り、HDAC3FFオルガノイドはIFN-γ刺激後にH2-Ab1発現を上昇させた(補足図5E);しかしながら、HDAC3発現欠損オルガノイドにおいても同様の誘導が起こり(補足図5E)、HDAC3欠損上皮細胞においてIFN-γ依存性のMHCII調節がそのまま残っていることが支持された。IFN-γはin vivoでは相乗的な役割を果たすかもしれないが、これらのオルガノイドのデータは、HDAC3が上皮のMHCII発現を制御する上皮内在性のNF-κB依存性メカニズムを示唆している。
上皮性MHCIIは、常在菌特異的CD4+T細胞と腸の炎症を制限する。上皮性MHCIIは、保護的および有害な免疫反応の両方で機能することが示唆されているが(53, 57, 62, 63)、通性特異的T細胞反応および微生物が誘発する炎症の制御におけるその役割は不明なままであった。そこで、これを検証するために、IECにおけるMHCIIを条件付きで欠損させたマウス(MHCIIΔIEC)を、フロックスドH2-Ab1マウス(MHCIIFF)とビリンプロモーターの下流でCreリコンビナーゼを発現するマウスとの交配により作成した(64, 65)。MHCIIΔIECマウスのIECにおけるMHCII発現の有意な低下は、mRNA(補足図6A)および表面タンパク質分析(補足図6B)により確認された。MHCIIの発現低下はIECに限られ、CD45+細胞では同レベルであった(補足図6C)。興味深いことに、古典的抗原提示細胞におけるMHCIIの活性化作用とは対照的に、IEC内在性MHCIIの欠損は、結腸(図4、AおよびB)および小腸における通性特異的cBir1+CD4+T細胞の蓄積を上昇させることが分かった(補足図6D)。MHCIIΔIECマウスの腸は、腸内細菌特異的Tregの減少(図4C)と腸内細菌特異的Th17細胞の増加(図4D)を示したが、腸内細菌特異的Th1細胞とTfh細胞は同様であった(補足図6E)。重要なことは、腸内細菌叢の組成と多様性は、MHCIIΔIECマウスとフロックス化同胞対照で同様であり(補足図7、A-C)、cBir1遺伝子を保有する細菌の割合が含まれていた(補足図7、DおよびE)。したがって、MHCIIΔIECマウスの腸内でcBir1常在菌特異的T細胞が増加したことは、常在菌の組成の変化を反映していないことがわかった。
上皮性MHCIIは常在菌特異的T細胞を制御している(図4
上皮性MHCIIは常在菌特異的T細胞を制御している。(AおよびB)MHCIIFFおよびMHCIIΔIECマウスの大腸におけるcBir1+特異的CD4+T細胞数.(C と D) MHCIIFF と MHCIIΔIECマウスの大腸におけるFoxP3+ (C) と RORγt+ (D) cBir1+ CD4+T細胞の頻度. (E) 2W1S-Candida albicans常在菌のコロニー形成の図。(F) MHCIIFFおよびMHCIIΔIECマウスの大腸におけるC. albicans-2W1S+特異的CD4+細胞の数。(G)2W1Sペプチド摂食モデルの図。(H)MHCIIFFおよびMHCIIΔIECマウスの大腸における2W1S+特異的CD4+細胞の数。(I)MHCIIFFおよびMHCIIΔIECマウスの直腸脱の頻度。(J)MHCIIFFマウスおよび脱腸したMHCIIΔIECマウス(MHCIIΔIEC*)のH&E染色された大腸切片。スケールバー:20μM。(KおよびL)MHCIIFFおよびMHCIIΔIEC*の便中の骨髄系細胞浸潤の頻度(K)およびリポカリン-2レベル(L)。(M-O) コントロールおよび増殖したMHCIIΔIECおよびHDAC3ΔIECマウスの大腸におけるcBir1+特異的CD4+ T細胞数(M)、FoxP3+ (N) およびRORγt+ (O) cBir1特異的T細胞頻度。テトラマー細胞は、生細胞、CD45+、系統(CD11b-B220-Ly6G-、CD11c-CD8a-)、CD4+にゲーティングされている。データは、少なくとも2つの独立した実験(A-H)の代表であるか、または少なくとも2つの独立した実験(I-O)からプールしたもので、1グループあたり3-6匹のマウス。*P < 0.05, **P < 0.01, ***P < 0.001, by unpaired 2-tailed Student's t test (B-H and L-O) or Mantel-Cox test (I).
この常在菌特異的応答が別の微生物抗原で起こるかどうかを調べるために、マウスはI-Eαエピトープの2W1S55-68バリアントを発現するCandida albicansでもコロニー形成した(図4E) (16, 17)。cBir1+CD4+T細胞の制御と一致して、MHCIIΔIECマウスは、C.アルビカンスコロニー化が同程度のMHCIIFFマウス(図4F)と比較してC.アルビカンス-2W1S特異的CD4+T細胞の数が増加していることを表示した。さらに、上皮MHCIIは、2W1Sペプチドを経口投与した後の2W1S+CD4+T細胞の腸内蓄積を制限するのに必要だった(図4、GおよびH)(66)。上皮性MHCIIが組織内T細胞を制御する機構を調べるため、MHCIIFFおよびMHCIIΔIECマウスで微生物特異的T細胞の増殖、生存、アネルギーが比較検討された。上皮型MHCIIの欠損により、増殖マーカーKi67(補足図8A)、アネルギーマーカーFR4、CD73(補足図8、B、C)に最小限の差異しか生じなかった。驚くべきことに、MHCIIΔIECマウスのcBir1+CD4+T細胞は、アポトーシスマーカーBim(補足図8、DおよびE)およびアネキシンV(補足図8F)の低下を示し、上皮性MHCIIがアポトーシスを促進して、MHCIIΔIECマウスの局所蓄積につながる可能性を示している。
IEC内在性HDAC3欠損マウスは、慢性腸炎に対する感受性が上昇した(補足図1)。驚くべきことに、HDAC3ΔIECマウスと同様に、上皮性MHCIIを欠くマウスも、腸の炎症を示す直腸脱の発生率が年齢とともに増加することを示した(図4I)。さらに、脱出したMHCIIΔIECマウス(MHCIIΔIEC*)は、慢性大腸炎に一致する病理学の増加(図4J)、ミエロイド細胞の浸潤の増加(図4K)、および糞便リポカリン-2の上昇(図4L)を示した。興味深いことに、脱出傾向が増加したMHCIIΔIECマウス及びHDAC3ΔIECマウスは、微生物特異的FoxP3+トレグ(図4N)の減少及び微生物特異的RORγt+炎症性Th17細胞(図4O)の増加を反映する、通性特異的CD4+T細胞(図4M)のレベル上昇によって特徴づけられた。これらのデータを総合すると、上皮のHDAC3は、微生物が誘発するMHCII指向性の交雑物特異的免疫および炎症を制御する上で重要である可能性が示唆された。
HDAC3は、マイクロバイオータによる上皮依存的なコミュンサル特異的免疫の制御を可能にする。MHCII依存性腸管炎症および交感神経特異的免疫における微生物叢の必要性を検証するために、MHCIIΔIECおよび対照MHCIIFFマウスの飲料水に、交感神経を著しく枯渇させる広域抗生物質を補充した(50)。以前の結果(図4I)と一致して、MHCIIΔIECマウスは直腸脱の有病率の増加を示したが(図5A)、微生物相の枯渇はMHCIIΔIECマウスの自然腸炎を防いだ(図5B)。さらに、微生物叢を枯渇させたMHCIIΔIECマウスは、常在菌特異的Th17細胞の蓄積(図5B)およびIL-17レベルの上昇(図5C)を示した。しかし、抗生物質で処理したMHCIIΔIECマウスは、MHCIIFFマウスと比較して、通性特異的Th17細胞(図5B)またはIL-17発現(図5C)の増加を示さなかった。これらのデータを総合すると、上皮のMHCII依存性の腸の炎症および通性特異的免疫応答の引き金に微生物叢が必要であることが浮き彫りにされた。
HDAC3がマイクロバイオータによる上皮依存的な腸管特異的免疫反応の制御を可能にする(図5
HDAC3は、微生物叢が上皮依存的な腸内細菌特異的免疫反応を制御することを可能にする。(A)コントロール(CNV)および抗生物質投与(ABX)したMHCIIFFおよびMHCIIΔIECマウスにおける直腸脱の発生頻度。(B と C) コントロールおよび ABX 処理した MHCIIFF および MHCIIΔIEC マウスの大腸における RORγt+cBir1+特異的 CD4+T細胞 (B) と IL-17 mRNA の発現頻度 (C).(DおよびE)GF-およびCNV-HDAC3FFおよびHDAC3ΔIECマウスの大腸におけるMHCII+ EpCAM+細胞の頻度。(F-H)cBir1+テトラマー特異的CD4+ T細胞の数(F)、GF-およびCNV-HDAC3FFおよびHDAC3ΔIECマウスから分離したFoxP3+(G)およびRORγt+(H)cBir1+ T細胞の周波数。 cBir1+テトラマー細胞は生きた、CD45+、系統(CD11b-B220-Ly6G-、CD11c-CD8a-)、CD4+でゲートされたものである。データは、少なくとも2つの独立した実験、1グループあたり3-5マウスの代表値である。*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, Mantel-Cox (A) または Tukeyの多重比較検定付き1-way ANOVA (B-H) によるものであった。
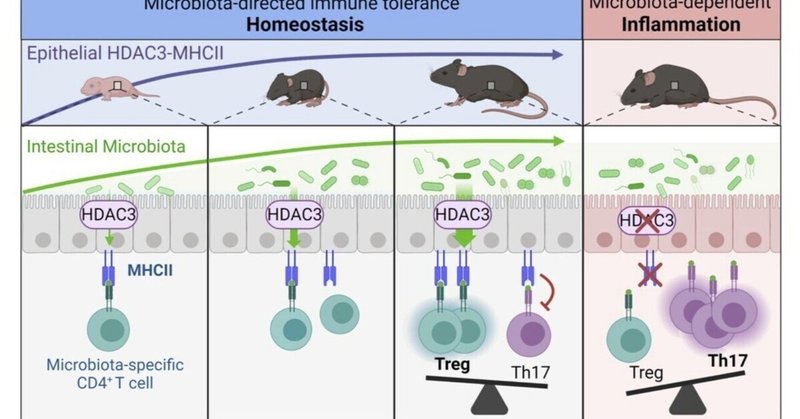
微生物叢による上皮細胞および免疫細胞のこの制御の仲介にHDAC3が必要かどうかを調べるために、GF由来のHDAC3FFおよびHDAC3ΔIECマウスをCNV-HDAC3FFおよび-HDAC3ΔIECマウスと比較検討した。野生型CNVマウスおよびGFマウスを用いた以前のデータ(図3、FおよびG)と一致して、IEC内在性MHCII発現は、GF-HDAC3FFマウスと比較してCNV-HDAC3FF対照で有意に誘導されたが(図5、DおよびE)、CNV-HDAC3ΔIECマウスはMHCIIを上昇させることができなかった(図5、DおよびE)。GFマウスでは、上皮HDAC3発現の喪失は、IEC上のMHCIIのタンパク質レベルに影響を与えず(図5、DおよびE)、上皮MHCIIの微生物叢依存性調節の仲介におけるHDAC3の特定の必要性が示された。さらに、CNV-HDAC3ΔIECマウスは、CNV-HDAC3FF同腹のコントロールと比較して、cBir1+通性特異的FoxP3+の減少および通性特異的Th17細胞の上昇を示した(図5、F-H)。しかし、GF-HDAC3ΔIECは、GF-HDAC3FFマウスのものと同様のレベルの交感神経特異的TregおよびTh17細胞を有した(図5、F-H)。これらのデータは、微生物叢が腸内細菌特異的な免疫を制御するためにHDAC3が必要であることを示している。これらの知見を総合すると、微生物叢のコロニー形成は、上皮のHDAC3/MHCII制御経路を通じて、局所組織環境における交配種特異的免疫反応を直接減弱させることにより、交配種の自己寛容を誘導できることが明らかになった(補足図9)。
考察
微生物叢は、宿主の免疫系の発達・教育に不可欠である。しかし、微生物叢に対する不適切な免疫反応がいくつかの慢性炎症疾患の根底にあり、微生物叢特異的免疫の制御方法を理解する必要性が浮き彫りになっている。本研究では、微生物叢の初期コロニー形成により、上皮のHDAC3発現が上昇し、常在菌特異的T細胞が拡大することを見出した。IECにおけるHDAC3の選択的欠失は、上皮性MHCII発現の低下と、上皮性MHCIIによって制御されている commensal-specific CD4+ T cellの制御障害を引き起こした。重要なことは、HDAC3またはMHCIIの上皮発現の喪失は、通性特異的Th17細胞の増加と同時に、通性特異的Tregの減少を招いたことである。これらの結果は、腸内細菌特異的T細胞が、健康な人の寛容細胞から、活動性のクローン病患者の炎症性IL-17分泌細胞へと切り替わることを示唆する最近のヒトのデータと一致している(14、67)。HDAC3ΔIECとMHCIIΔIECマウスモデルでは、常在菌の組成が異なるが、両モデルともcBir1+常在菌特異的CD4+ T細胞が同様に増加した。微生物特異的T細胞の宿主内在性制御と一致して、HDAC3は、微生物が微生物特異的T細胞の上皮MHCII依存性制御を誘導するのに必要であった。したがって、常在微生物は、常在特異的CD4+ T細胞サブセットの局所動態およびマイクロバイオータ感受性疾患への感受性を制御するHDAC3依存性上皮MHCII経路を誘導することによって、自己寛容を指示することができる(補足図9)。
マイクロバイオータ特異的CD4+ T細胞は、胸腺で中枢的に生成され(68)、末梢粘膜組織で生成される(15, 69-71)。自己反応性CD4+ T細胞は胸腺ネガティブセレクションによって制限されることがよく知られているが(72-74)、通性特異的細胞を制御するメカニズムはあまりよく理解されていないままである。以前の研究では、活性化した commensal特異的T細胞の負の選択は、MHCII発現のグループ3自然リンパ系細胞(ILC3s)によって媒介されうることが記述されている(75、76)。ILC3は腸管内腔表面では比較的まれな細胞であるが、我々はIECが腸管MHCIIの主要な供給源であることを示している。上皮細胞によるHDAC3依存的なMHCII発現は、腸内細菌特異的Th17の蓄積を制限しており、腸内細菌特異的CD4+T細胞の制御と微生物叢に対する病原性応答の抑制に対する内腔表面の主要なメカニズムである可能性がある。通性特異的T細胞蓄積の制御に加え、上皮HDAC3またはMHCIIの損失は、RORγt+抗原提示細胞での最近の観察(77-79)と同様に、通性特異的Tregの著しい減少をもたらした。しかしながら、微生物叢に対するCD4+ T細胞応答は、HDAC3に限定されないin vivoでの複数のシグナル伝達経路を反映していると思われる。このように、異なる非典型的な細胞における抗原提示経路の相補的かつ本質的な役割は、健康な腸内組織の恒常性を確立し維持するために極めて重要であると思われる。
腸の健康と疾病の制御における上皮性MHCIIに関する研究は、様々な、あるいは相反する結果をもたらし、この制御の文脈依存的な役割を浮き彫りにしている。以前の研究では、上皮性MHCIIの欠損が、T細胞による腸の炎症と移植片対宿主病のモデルマウスを保護することが示唆されていた(53, 62)。一方、他の報告では、上皮MHCIIは、CD4+ T細胞によるバルクTregの発達とIL-10の発現を促進し(57、80)、CD4+ T細胞の活性化を制限している(81)ことが示されている。さらに、IECは限られたコスティミュレーション分子を発現しており(82-84)、上皮性MHCIIの発現は、古典的な抗原提示細胞とは異なり、寛容なT細胞応答を促進する可能性が示唆される。実際、我々のデータは、上皮性MHCIIが腸内常在菌特異的T細胞応答を制御することにより腸の恒常性を促進するという、これらの後者の研究による予測と一致している。実際、上皮のHDAC3依存性MHCIIを欠損すると、腸内細菌特異的Tregが減少し、同時にTh17細胞が増加することが判明した。Tuganbaevらによる予測(57)と一致するように、我々の2W1Sのデータも、上皮性MHCIIが摂取した抗原に向けられた腸の免疫応答を制御している可能性を裏付けている。このように、上皮性MHCII発現細胞は、常在微生物および食物抗原の両方に対する免疫寛容をより広く制御し、腸の病理学的炎症を制御する可能性がある。
HDAC3依存性の上皮性MHCIIの欠損により、常在菌特異的なTh17活性が上昇し、微生物叢主導の腸の炎症への感受性が高まった(補足図9)。IFN-γはTh1反応を増幅する一方で、Th17細胞を含む他のThサブセットの分化と機能を阻害する(51, 52)。さらに、IL-17はTh1の分化を制限しながらIFN-γの産生を抑制することができる(85)。しかし、我々の系では、IFN-γは、HDAC3ΔIECマウスで観察されるTh17表現型を駆動する主要な因子ではない。さらに、上皮 MHCII を失ったマウスでは、常在菌特異的な Th17 細胞の上昇も見られた。驚くべきことに、活動性IBDの患者は、健常対照者に見られるものと比較して、機能的に異なる微生物反応性CD4+ T細胞を持ち、より高いレベルのIL-17を分泌する(14, 26, 28-30, 67)。IL-17は、IBDを含むいくつかの自己免疫疾患や炎症疾患の発症や増悪に関連しています(28, 30, 86)。さらに、IL-17のシグナル伝達が阻害されたマウスモデルは腸の炎症から保護されており(87, 88)、腸の炎症におけるIL-17の病原的な役割を裏付けている。さらに、微生物特異的なTh17細胞は、T細胞依存性大腸炎の駆動に必要であり(89)、RORγt-またはIL-17欠損T細胞の移入は、腸の炎症発生を防ぐ(90)。腸内細菌特異的T細胞は、特異的四量体陽性細胞の希少性から直接移植することはできないが、cBir1+ Th17細胞の増殖には微生物叢の存在が必要であり、腸内細菌の反応性Th17が腸炎誘導に重要であることが示唆される。しかし、IBD患者を対象としたIL-17を枯渇させる臨床試験は効果がなく、場合によっては病状を悪化させることもあった(91)。腸のホメオスタシスを制御するIL-17のこのような多面的な役割は、IL-17自体を広く中和するのではなく、特定のIL-17産生者をターゲットにすることが、より効果的な治療法となる可能性を示唆している。実際、他の細胞型からのIL-17産生を維持しつつ、腸内常在菌に特異的なTh17細胞を標的とする治療薬が、腸の炎症を抑制することが示されている(92)。我々のデータは、HDAC3活性の増強により上皮のMHCII発現を促進することで、炎症性腸内常在菌特異的T細胞をさらに制限し、健康な腸のホメオスタシスを促進する可能性を示唆している。
微生物叢と哺乳類免疫細胞のクロストークには、パターン認識受容体の関与と微生物叢由来の代謝物シグナルを介した情報伝達が含まれる。我々のデータは、微生物叢に感受性のある酵素HDAC3が腸の上皮のMHCII発現を制御するという、異なるレベルの上皮の制御を明らかにしている。実際、上皮のHDAC3の欠損は、腸における微生物叢依存的なMHCIIの発現を消失させるのに十分であった。これまでの研究で、微生物叢がIEC内在性のMHCII発現に必要であることが示されている(53-57)。これらの研究の多くは、免疫細胞がIFN-γシグナルを介してどのように上皮性MHCIIを誘導するかに着目している(53、93、94)。in vitroの研究に用いられたIFN-γの量は、腸の基礎的な恒常性濃度に比べて比較的高い。しかし、この制限にもかかわらず、我々の研究は、微生物叢がHDAC3を介して上皮MHCIIの発現を促進する、上皮細胞固有の追加のメカニズムを示唆するものであった。さらに、正規のパターン認識経路の破壊は、腸管バリアの完全性の喪失と炎症の増加をもたらし(95-97)、TLRシグナルアダプターであるMyD88とTRAFは、特に小腸において上皮性MHCIIの発現を誘導することが示されている(53)。MyD88/TRAF シグナルは NF-κB の活性化を促進し、CIITA と MHCII を駆動することができるが、HDAC3 は NF-κB の活性化を制御することが分かっている (58, 59, 61, 98, 99)。我々のデータは、NF-κBがHDAC3依存的に上皮性MHCIIの発現を誘導することを示すものである。HDAC3は、ヒストンおよび非ヒストン標的を脱アセチル化し、遺伝子発現や細胞機能を変化させることができる多面的な酵素である。したがって、HDAC3ΔIECマウスで観察される転写の違いは、ヒストン、および潜在的に非ヒストン標的や酵素に依存しない役割の制御が変化した結果である(59, 60, 100-103)。このIEC内在性のメカニズムにもかかわらず、上皮のMHCIIを促進するIFN-γの相乗的役割、またはin vivoでのMHCIIのHDAC3依存性の制御が、細胞内在性および外来性の経路の統合的ネットワークを反映していることを除外することはできない。
肺上皮細胞、皮膚ケラチノサイト、および線維芽細胞を含むいくつかの組織特異的、非造血性細胞が、MHCIIと組織内在性のCD4+T細胞応答を制御するのに必要な機構を発現することを示す証拠が増えている(104-106)。HDAC3のユビキタスな性質を考えると、HDAC3が他の非古典的な抗原提示細胞においてもMHCII依存性の経路を促進することは、したがってもっともなことである。興味深いことに、HDAC3の組織特異的な欠損は、糖尿病、心臓病、アルツハイマー病、IBDの慢性炎症性疾患モデルと関連している(37, 38, 107-109)。重要なことは、今回の我々の発見により、非造血幹細胞MHCIIのHDAC3誘導が微生物叢によって大きく誘導され、局所の自己指向性免疫応答を弱めるという、免疫制御に関する基本的な新しい考え方が明らかにされたことである。これらの知見は、微生物叢が常在菌指向性免疫の制御に利用する中心的な宿主メカニズムを明らかにし、この制御経路が慢性炎症疾患の治療のターゲットになることを示唆している。
研究方法
マウス C57BL/6Jマウスは、The Jackson Laboratoryから購入し、Cincinnati Children's Hospital Medical Center(CCHMC)の特定の病原体を含まない従来型(CNV)施設で維持した。無菌(GF)マウスは、CCHMC Gnotobiotic Mouse Facilityのフレキシブルなアイソレーターで維持し、オートクレーブした飼料と水を与え、微生物が存在しないことをモニターした。HDAC3FF、HDAC3ΔIEC、およびHDAC3ΔIEC-INDマウスは、以前に記載されたように作製した(37)。H2-Ab1(MHCII)FFおよびRag1-/-マウスは、The Jackson Laboratoryから購入し、CCHMCで維持した。MHCIIFFマウスをC57BL/6J-Villin-Creと交配して、MHCIIΔIECマウスを作製した。すべての研究において、性・年齢を一致させた同腹仔マウスを使用した。マウスは1ケージあたり4匹まで飼育し、12時間明期/12時間暗期のサイクルで、水と餌に自由にアクセスできる換気ケージシステムで飼育した。すべてのマウス研究は、CCHMCのInstitutional Animal Care and Use Committeeの承認を得て実施された。これらのプロトコルは、米国公衆衛生局および農務省が制定した基準に従っている。すべての実験は、Animal Researchが定める基準に従った。Reporting of In Vivo Experiments (ARRIVE)が定める基準に従った。
マウス大腸炎モデルおよび2W1Sモデル。T細胞移植大腸炎モデルについては、HDAC3FFおよびHDAC3ΔIECマウスの脾臓および腸間膜リンパ節から、MojoSort Mouse CD4 Naive T Cell Isolation(BioLegend)を介して5×105個のナイーブCD4+ T細胞を単離した。細胞の純度をフローサイトメトリーで確認し、年齢と性をマッチさせたRag1-/-レシピエントにT細胞をi.p.インジェクションした。抗生物質処理として、MHCIIFFおよびMHCIIΔIECマウスの仔には、離乳時に1 mg/mL コリスチン(ミリポアシグマ)、1 mg/mL アンピシリン(ミリポアシグマ)および5 mg/mL ストレプトマイシン(ミリポアシグマ)を添加した水を与え、16週間抗生物質を投与して維持させた。Rag1-/-マウスは、T細胞移植の7-10日前に同じ抗生物質カクテルを与えられ、その後、実験期間中、抗生物質水で維持された。抗生物質水は7-10日ごとに更新された。PBSまたは10μgの組換えIFN-γ(PeproTech)i.p.を受けたマウスを、24時間後に分析した。C. albicans-2W1Sコロニー形成は、以前に記載されたように実施した(16、17、110)。簡単に言えば、マウスをコロニー形成の2日前にアンピシリン-水(1mg/mL)で前処理し、実験の期間中アンピシリン-水で維持した。マウスに、2W1S55-68ペプチドを発現する組換えC. albicansを口腔内に滴下した(111)。マウスは、コロニー形成の14日後に採取した糞便CFUをプレーティングすることにより、コロニー形成をモニターされた。2W1Sの摂食については、MHCIIFFおよびMHCIIΔIECマウスに、0日目、2日目、および4日目に100μgの2W1Sペプチドを経口投与し、そして6日目に2W1S特異的細胞を既報(66)通りに採取した。
細胞の単離。大腸を摘出し、開腹し、PBSで洗浄した。IECについては、組織を予め温めたストリップバッファー(PBS、5%FBS、1mM EDTA、1mM DTT)中に入れ、37℃で45°の角度で180rpmで振盪しながら15分間インキュベートした。固有層分離のために、組織をPBSで洗浄してEDTAとDTTを除去した後、予め温めておいた消化バッファ(RPMI with 1 mg/mL Collagenase/Dispase [MilliporeSigma] )中で37℃、180 rpmで振りながら30分インキュベートした。インキュベーション後、組織をボルテックスし、70μmのセルストレーナーを通過させた。
フローサイトメトリー。FACSバッファ(2%FBS、0.01%アジ化ナトリウム、PBS)で希釈した以下の抗体を用いて、細胞をフローサイトメトリー用に染色した。BV711-抗CD326(EpCAM)(クローンG8.8、BioLegend)、BUV395-抗CD45.2(クローン104、BD Biosciences)、APC-またはFITC-抗MHCII(クローンM5/114.15. 2、eBioscience)、APC-eFluor 780-抗CD4(クローンRM4-5、eBioscience)、PE-Cy7-抗CD44(クローンIM7、eBioscience)、Alexa Fluor 647-またはBV650-抗RORγt(クローンQ31-378、BD Biosciences)、PerCP-eFluor710-またはAPC-抗CD8a(クローン53-6. 7、eBioscience)、PerCP-Cy5.5-抗CD3(クローン17A2、eBioscience)、PerCP-eFluor 710-抗B220(クローン RA3-6B2、eBioscience)、PerCP-eFluor 710-抗Ly6G(クローン 1A8-Ly6g, eBioscience)、PerCP-Cy5. 5-、eFluor 450-、またはPE-抗CD11b(クローンM1/70、eBioscience)、PerCP-Cy5.5-抗CD11c(クローンN418、eBioscience)、BV650-抗CXCR5(クローン138D7、BioLegend)、アレクサフルオロ647-抗FR4(クローン12A5、BioLegend)、eFluor450-抗CD73(クローンeBioTY/11. 8、eBioscience)、Alexa Fluor 488-anti-Bim(C34C5, Cell Signaling Technology)、BV421-anti-Tbet(clone 4B10、BioLegend)、FITC-anti-CD90.2(clone 53-2. 1、eBioscience)、PE-Cy7-抗CD127(クローンA7R34、eBioscience)、eFluor 450-抗Ki67(クローンSolA15、eBioscience)、およびPE-抗IFN-γ(クローンXMG1.2、eBioscience)である。サイトカイン染色については、細胞を50ngのPMAと1μgのイオノマイシンで37℃、3〜4時間刺激した。細胞は、Annexin V染色バッファー(BD Pharmingen)で希釈したAlexa Fluor 488またはAPC標識annexin V(eBioscience) で染色された。死細胞はFixable Aqua Dead Cell Stain Kit (Invitrogen)で除外した。細胞内染色にはBD Fix/Permキットを使用した。クラスII制限四量体(cBir1:YSNANILSQ;および2W1S:EAWGALANWAVDSA)はPE結合で、NIH tetramer coreによって生成・提供されたものであった。テトラマー染色では、サンプルをテトラマー(1:100)およびFc Block(抗マウスCD16/CD32、eBioscience)と共に、室温で1時間インキュベートした。サンプルはBD LSRFortessaで取得し、FlowJo Software (Tree Star)で解析した。
RNAおよび定量PCR解析。RNAは、RNeasy Kit(Qiagen)を用いて単離した。全組織からの RNA の場合、サンプルは TRIzol でホモジナイズした。cDNAはVerso reverse transcriptase kit (Thermo Fisher Scientific)を用いて製造者のプロトコルにしたがって合成した。リアルタイムPCRは、SYBR Green(Applied Biosystems)を用いて行い、以下のマウスプライマー配列を使用して解析した。HPRTフォワード5′-GATTAGCGATGAACCAGGT-3′、HPRTリバース5′-CCTCCCATCTCCTTCATGACA-3′、H2-Ab1フォワード 5′-TGTGAGTCCTGTGACTGCCATTA-3′, H2-Ab1リバース5′-TCGCCCATGAACTGGTACGAAA-3′、IL-17フォワード5′-ACCGCAATGAAGACCCTGAT-3′, IL-17 reverse 5′-TCCCTCCGCATTGACACA-3′, HDAC3 forward 5′-TTGGTATCCTGGAGCTGCTT-3′, HDAC3 reverse 5′-GACCCGGTCAGTGAGGTAGA-3′, IL-17 reverse 5′-TCCCTCCGCATTGACA-3′, CIITA forward 5′-CCCTGCGTGATGATGTC-3′,CIITA reverse 5′-ATCTCAGACTGATCCTGGCAT-3′である。
リポカリン-2 ELISA。糞便ペレットを100mg/mLの濃度でPBS中でホモジナイズした後、高速で10分間遠心分離した。上清を回収し、マウスLipocalin-2/NGAL ELISAキット(R&D Systems)を用いて、製造者の説明書に従ってリポカリン-2レベルを決定した。
腸管オルガノイド。腸オルガノイドは、HDAC3FFおよびHDAC3ΔIEC-INDマウスの結腸から、以前に記載されたように生成した(38、112)。簡単に言えば、結腸を開き、小片に切断し、キレート緩衝液(PBS中2mM EDTA)中で4℃、30分間、回転させながらインキュベートした。次に組織を振盪バッファー(PBS、43.3 mMショ糖、54.9 mMソルビトール)に移し、2〜5分間手で振盪した。結腸陰窩を再懸濁し、オルガノイド培養液(10mM HEPES、2mM l-グルタミン酸を補充した60% Advanced DMEM/F12 medium [Gibco, Thermo Fisher Scientific], 40% L-WRN conditioned medium, 1× N2 supplement, 1× B27 supplement, 50 ng/mL murine EGF, 10 μM Y-27632 ROCK inhibitor)とともにマトリゲル(コーニング)内にプレーティングした。培養液は2-3日おきにリフレッシュした。HDAC3 欠損を誘発するために、オルガノイドを 1 μM ヒドロキシタモキシフェン(4-OHT、MilliporeSigma) で 24 時間処理した。オルガノイドを、ビヒクル(DMSO)または5μMのIKK-16(Selleckchem)で24時間処理した。IFN-γ刺激については、オルガノイドを100 U/mLのマウス組換えIFN-γ(BioLegend)で24時間処理した。インキュベーション後、オルガノイドをPBSで3回洗浄し、RNeasyキット(Qiagen)を用いて溶解した。
微生物叢の解析 ショットガンメタゲノムシーケンスのために、製造者の推奨に従って、PowerFecal DNA isolation kit (Qiagen Inc.) を用いて約0.1 gの便からDNAを抽出した。シーケンスライブラリーは、Nextera XTプロトコル(Illumina)を使用して微生物DNAから生成した。配列決定は、Illumina NovaSeq 6000マシンで、150bp DNAペアエンドリードを用いて、サンプルあたり約4G塩基対の深さで実施した。生シーケンスデータは、デマルチプレックスしてFASTA形式に変換し、下流解析に供した。各サンプルのペアエンドシーケンスリードを、ヒトゲノムと約40,054の細菌、真菌、ウイルス、寄生虫ゲノムからなるカスタムゲノムデータベースに対してKraken (113) でアライメントしました。このデータベースは、まず RefSeq ゲノムデータベース (https://www.ncbi.nlm.nih.gov/refseq/) に含まれるすべての細菌、真菌、ウイルスと、ヒトゲノム (GR38Ch; https://www.ncbi.nlm.nih.gov/projects/genome/guide/human/index.shtml) から導出されました。NCBI Assemblies や PATRIC のドラフトゲノムなど、手動によるキュレーションでゲノムを追加している。グループ間の全体的なマイクロバイオーム組成の比較は、RのVeganパッケージ(114)を用いて、多重応答並べ替え手順(MRPP)により実施した。同腹のHDAC3FFおよびHDAC3ΔIECならびにMHCIIFFおよびMHCIIΔIECマウスから得られたマイクロバイオームショットガン配列決定リードは、プログラムDiamond(116)のtblastコマンドおよびデフォルト設定を使用して、cBir1ペプチド配列(115)(AY551005.1)に対してアラインされた。この遺伝子の有病率は、マッピングされた100万個の細菌リードあたりのcBir1カウントとして表される。cBir1定量PCR(qPCR)分析のために、便DNAは、QIAamp Fast DNA Stool Extract Kit(Qiagen)を用いて、製造者の説明書に従って抽出された。細菌DNAは、以下のプライマーセット:cBir1 forward 5′-AAGTACTTTACGGCAGGCG-3′, cBir1 reverse 5′-TCTGTTCCGTCAGCACCTAC-3′ を用いたqPCR分析によってcBir1遺伝子発現を評価した。
組織学的解析。大腸組織切片を4%パラホルムアルデヒドで4℃、一晩固定し、パラフィン包埋、切片作成、H&E染色を行った。組織学的スコアリングのため、スライドを以下のパラメーターに基づいて評価した:免疫細胞浸潤、1-5;粘膜肥厚/浮腫、1-5;陰窩長、1-5;および陰窩脱落/侵食、1-5。
統計。結果は、平均値±SEMで示した。2群間の検定は両側Studentのt検定で行った。多重比較にはTukey's test付き1-way ANOVAを使用した。脱出曲線はlog-rank Mantel-Cox検定で評価した。結果は、*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001で有意であるとみなされた。統計的有意性はPrism version 7.0 (GraphPad Software)を用いて算出した。
研究の承認 すべての動物実験は、Cincinnati Children's Hospital Medical CenterのInstitutional Animal Care and Use Committeeにより承認された。
著者による貢献
EME、SSW、TAが本研究の構想を練り、研究方法を考案した。EME、TYA、VW、TR、BJD、LE、JW、DBHが実験を実施した。EME、TYS、TR、DBHがデータを解析した。EME、TA、SSWは原稿の執筆、レビュー、編集を行った。
補足資料
補足データを見る
謝辞
NIH Tetramer Core Facility (NIH-NIAID contract 75N93020D00005) にMHCモノマーおよび4量体試薬を提供いただいた。Way、Qualls、Deshmukhの各研究室には有益な議論を、Allenghatの研究員には原稿の重要な読み合わせをいただいた。CCHMC Veterinary Services、Research Flow Cytometry Core、Confocal Imaging Core、Pathology Research Coreのサービスおよび技術支援に感謝する。本研究は、NIH(DK114123, DK116868 to TA, DP1AI131080 to SSW, and F32AI147591 to EME)、およびTAへのKenneth Rainin Foundation awardによる支援を受けている。SSWとTAはそれぞれBurroughs Wellcome FundからInvestigators in the Pathogenesis of Infectious Disease Awardを授与されている。このプロジェクトはPublic Health Serviceの助成金P30DK078392によって部分的に支援された。
宛先は以下の通り。Theresa Alenghat, Cincinnati Children's Hospital Medical Center, 3333 Burnet Avenue, MLC 7038, Cincinnati, Ohio 45229, USA(シンシナティ小児病院医療センター、米国)。電話番号 513.803.7498; Email: Theresa.Alenghat@cchmc.org。または、下記まで。Sing Sing Way, Cincinnati Children's Hospital Medical Center, 3333 Burnet Avenue, MLC 7017, Cincinnati, Ohio 45229, USA.まで。電話番号 513.636.7603; Email: SingSing.Way@cchmc.org。
脚注
利益相反。著者らは、利益相反が存在しないことを宣言している。
著作権:© 2023, Eshleman et al. これは、Creative Commons Attribution 4.0 International Licenseの条件の下で公開されたオープンアクセス論文である。
参考情報 J Clin Invest. 2023;133(4):e162190. https://doi.org/10.1172/JCI162190.
参考文献
Hill DA, Artis D. Intestinal bacteria and the regulation of immune cell homeostasis(腸内細菌と免疫細胞のホメオスタシス制御)。Annu Rev Immunol. 2010;28(1):623-667.
この記事を見るには CrossRef PubMed Google Scholar
Belkaid Y, Harrison OJ. ホメオスタシス免疫と微生物叢。Immunity. 2017;46(4):562-576.
この記事を経由して見る CrossRef PubMed Google Scholar
本田 圭、Littman DR. 適応免疫のホメオスタシスと疾患における微生物叢(The microbiota in adaptive immune homeostasis and disease)。Nature. 2016;535(7610):75–84.
この記事を経由して表示する。CrossRef PubMed Google Scholar
香山 浩、他:腸内における微生物叢、上皮、免疫細胞の相互作用. Annu Rev Immunol. 2020;38:23-48.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Shao TY, et al. Friendly fungi: commensal Candida albicansとの共生. Trends Immunol. 2022;43(9):706-717.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Koenig JE, et al. 発達期の乳児の腸内細菌叢における微生物コンソーシアムの継代。2011;108(suppl 1):4578-4585.
この記事を見るには PubMed Google Scholar
Yatsunenko T, et al. 年齢や地理的な違いによるヒト腸内細菌群の変化。ネイチャー。2012;486(7402):222–227.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Zheng D, et al. 健康や病気における微生物叢と免疫の相互作用。Cell Res. 2020;30(6):492-506.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Bäckhed F, et al.生後1年間のヒト腸内細菌叢の動態と安定化。セルホストマイクロビー。2015;17(5):690-703.
この記事を経由して表示する。CrossRef PubMed Google Scholar
本田 圭、Littman DR. 感染症や炎症におけるマイクロバイオーム。Annu Rev Immunol. 2012;30(1):759-795.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Duchmann R, et al. 腸内細菌、バクテロイデス、ビフィズス菌、およびヒトの常在腸内細菌叢からの抗原に対するT細胞特異性と交差反応性。Gut. 1999;44(6):812-818.
この記事を経由して見る CrossRef PubMed Google Scholar
Duchmann R, et al. 腸内細菌叢に対する寛容性は存在するが、活動性の炎症性腸疾患(IBD)では壊される。Clin Exp Immunol. 1995;102(3):448-455.
この記事を経由して表示します。PubMed Google Scholar
Ergin A, et al. クローン病および強直性脊椎炎患者における大腸菌タンパク質に対する末梢Th1 CD4 T細胞応答の障害。J Clin Immunol. 2011;31(6):998-1009.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Hegazy AN, et al. 腸内細菌叢に反応する循環および組織常在のCD4+ t細胞は、健常者に多く、炎症時に機能が変化する。Gastroenterology. 2017;153(153):1320–1337.
この記事を経由して見る PubMed Google Scholar
Hand TW, et al. 急性消化管感染症は、長寿命の微生物特異的T細胞応答を誘導する。Science. 2012;337(6101):1553–1556.
この記事を経由して表示する CrossRef PubMed Google Scholar
カンジダ・アルビカンス(Candida albicans)は、全身性のTh17免疫学的反応を正に調整する。Cell Host Microbe. 2019;25(3):404-417.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Jiang TT, et al. Commensal fungi recapitulate the protective benefits of intestinal bacteria.腸内細菌の保護効果を再現する。Cell Host Microbe. 2017;22(6):809-816.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Su LF, et al. Virus-specific CD4(+) memory-phenotype T cells are abundant in unexposed adults.(ウイルス特異的CD4(+)メモリー表現型T細胞は、非被ばく成人において豊富に存在する。Immunity. 2013;38(2):373-383.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Campion SL, et al. 未被曝献血者におけるHIV特異的ナイーブおよびメモリーCD4(+)T細胞のプロテオームワイド解析. J Exp Med. 2014;211(7):1273–1280.
この記事を経由して見る CrossRef PubMed Google Scholar
Kaser A, et al. 炎症性腸疾患. Annu Rev Immunol. 2010;28(12):573-621.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Tindemans I, et al. IBDにおけるT細胞媒介性炎症の不均質性を解明する。Cells. 2020;9(1):110.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Clemente JC, et al. 腸内細菌叢のヒトの健康への影響:統合的な見方. Cell. 2012;148(6):1258–1270.
この記事を経由して表示します。CrossRef PubMed Google Scholar
腸内細菌に対するエフェクターT細胞およびメモリーT細胞の反応。Trends Immunol. 2013;34(6):299-306.
この記事を経由して表示 CrossRef PubMed Google Scholar
炎症性腸疾患における腸のホメオスタシスとその破綻。ネイチャー。2011;474(7351):298–306.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Cong Y, et al. 自然発症した大腸炎C3H/HeJBirマウスにおける腸内細菌抗原に反応するCD4+ T細胞:Tヘルパー細胞1型反応の増加と疾患移行能。J Exp Med. 1998;187(6):855-864.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Annunziato F, et al.ヒトTh17細胞の表現型と機能的特徴。J Exp Med. 2007;204(8):1849–1861.
この記事を経由して表示する CrossRef PubMed Google Scholar
Alexander M, et al. ヒト腸内細菌の代謝は、Th17の活性化と大腸炎を促進する。セルホストマイクロビー。2022;30(1):17-30.
経由してこの記事を表示します。CrossRef PubMed Google Scholar
Gálvez J. Role of Th17 cells in the pathogenesis of human IBD.ヒトIBDの病態におけるTh17細胞の役割。ISRN Inflamm. 2014;2014:1-14.
この記事を経由して表示します。PubMed Google Scholar
Fujino S, et al. 炎症性腸疾患におけるインターロイキン17の発現亢進. Gut. 2003;52(1):65-70.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Zhao J, et al. 炎症性腸疾患におけるTh17細胞:サイトカイン、可塑性、および治療法。J Immunol Res. 2021;2021:8816041.
この記事を経由して表示する。PubMed Google Scholar
Eshleman EM, Alenghat T. 上皮による微生物叢由来のシグナルの感知。Genes Immun. 2021;22(5–6):237–246.
この記事を表示する。PubMed Google Scholar
ローDD. 警戒か転覆か?粘膜組織における恒常性M細胞および誘導性M細胞。Trends Immunol. 2017;39(3):185-195.
この記事を経由して見る PubMed Google Scholar
Kulkarni DH, et al.ゴブレット細胞関連抗原通過は、口腔内耐性の誘導と維持をサポートする。Mucosal Immunol. 2020;13(2):271-282.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Kanaya T, et al. Intestinal M cells: tireless samplers of enteric microbiota(腸管M細胞:腸内細菌叢の疲れを知らないサンプラー). トラフィック. 2020;21(1):34-44.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Biton M, et al. Tヘルパー細胞サイトカインは、腸管幹細胞の再生と分化を調節する。Cell. 2018;175(5):1307–1320.
この記事を経由して表示する。CrossRef PubMed Google Scholar
神林 毅、Laufer TM. 非定型MHCクラスII発現抗原提示細胞:樹状細胞に代わるものはあるのか?Nat Rev Immunol. 2014;14(11):719-730.
この記事を経由して見る CrossRef PubMed Google Scholar
Alenghat T, et al. ヒストン脱アセチル化酵素3は、常在菌に依存した腸のホメオスタシスを調整する。Nature. 2013;504(7478):153–157.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Wu S, et al.腸内細菌叢由来の代謝産物がHDAC3活性を促進する。Nature. 2020;586(7827):108–112.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Navabi N, et al. 上皮型ヒストンデアセチラーゼ3は、局所リンパ球の活性化を調整することにより腸管免疫を指示する。Cell Rep. 2017;19(6):1165-1175.
この記事を経由して見る CrossRef PubMed Google Scholar
Kuang Z, et al. 腸内細菌叢はヒストンデアセチラーゼ3を通じて宿主の代謝の日内リズムをプログラムしている. Science. 2019;365(6460):1428–1434.
この記事を経由して見る CrossRef PubMed Google Scholar
Aagaard K, et al. Una destinatio, viae diversae: 膣マイクロバイオータへの曝露は乳児に健康利益を与え、曝露の欠如は疾患リスクを与えるか?EMBO Rep. 2016;17(12):1679-1684.
この記事を経由して見る CrossRef PubMed Google Scholar
Russell SL, et al. 幼少期の抗生物質による微生物叢の変化は、アレルギー性喘息への感受性を高める。EMBO Rep.2012;13(5):440-447。
この記事を経由して表示する。CrossRef PubMed Google Scholar
Gensollen T, et al. 幼少期の微生物相によるコロニー形成が免疫系を形成する仕組み。Science. 2016;352(6285):539–544.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Al Nabhani Z, et al. microbiotaに対する離乳反応は、成人における免疫病理学に対する抵抗性に必要である。Immunity. 2019;50(5):1276–1288.
この記事を経由して見る CrossRef PubMed Google Scholar
Wang T, et al.早世の抗生物質曝露と宿主の健康。微生物と免疫の相互作用の役割。Semin Perinatol. 2020;44(8):151323.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Gray J, et al. 腸内常在菌は肺粘膜免疫を媒介し、新生児マウスの感染に対する抵抗性を促進する。Sci Transl Med. 2017;9(376):1-14.
この記事を経由して見る PubMed Google Scholar
Ostanin DV, et al.慢性大腸炎のT細胞移植モデル:概念、考察、および取引のトリック。Am J Physiol Gastrointest Liver Physiol.2009;296(2):135-146。
この記事を経由して表示します。PubMedのGoogleスカラー
制御性T細胞の枯渇による免疫不全マウスの炎症性腸疾患の誘発。Curr Protoc Immunol. 1999;30(1):1-10.
この記事を経由して表示する。PubMed Google Scholar
Powrie BF, et al. CD45RB高およびCD45RB低CD4+ T細胞間の制御的相互作用は、防御的および病原性細胞媒介免疫間のバランスに重要である。J Exp Med. 1994;179(2):589-600.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Vétizou M, et al. CTLA-4遮断による抗がん免疫療法は、腸内細菌叢に依存する。Science. 2015;350(6264):1079–1084.
この記事を経由して見る CrossRef PubMed Google Scholar
Park H, et al. CD4T細胞の異なる系統は、インターロイキン17を産生することで組織の炎症を制御する。Nat Immunol. 2005;6(11):1133–1141.
この記事を経由して表示します。CrossRef PubMed Google Scholar
インターロイキン17を産生するCD4+エフェクターT細胞は、Tヘルパー1型および2型の系譜とは異なる系譜を経て発生する。Nat Immunol. 2005;6(11):1123–1132.
この記事を経由して見る CrossRef PubMed Google Scholar
腸管上皮によるMHCクラスII抗原提示は、移植片対宿主病を発症させ、微生物叢に影響される。Immunity. 2019;51(5):885-898.
この記事を経由して見る CrossRef PubMed Google Scholar
Matsumoto S, et al. γδTCRを持つ上皮内リンパ球は、マウス小腸上皮のクラスII主要組織適合性複合体分子の発現を制御する. 上皮細胞生物学, 1995;4(4):163-170.
この記事を経由して見る PubMed Google Scholar
分節型糸状菌は、上皮内リンパ球を活性化し、MHCクラスII分子とフコシルアシアロGM1糖脂質を無胚葉マウスの小腸上皮細胞上に誘導する腸内常在菌である。Microbiol Immunol. 1995;39(8):555-562.
この記事を経由して表示する。CrossRef PubMed Google Scholar
マウス腸管上皮細胞の細胞増殖および分化におけるガンマデルタT細胞受容体上皮内リンパ球の生理的役割(松本聡、他) Immunology. 1999;97(1):18-25.
この記事を経由して見る CrossRef PubMed Google Scholar
Tuganbaev T, et al. Diet diurnally regulates small intestinal microbiome-epithelial-immune homeostasis and enteritis(食事は小腸の微生物-上皮-免疫の恒常性と腸炎を調節する). Cell. 2020;182(6):1441–1459.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Ziesché E, et al. IL-1シグナルにおけるヒストンデアセチラーゼ3のコアクチベーターの役割には、p65 NF-κBの脱アセチル化が関与している。Nucleic Acids Res.2013;41(1):90-109に掲載されました。
この記事を経由して見る CrossRef PubMed Google Scholar
Chen X, et al. マクロファージにおける炎症性遺伝子発現プログラムに対するヒストン脱アセチル化酵素Hdac3の必要性. Proc Natl Acad Sci U S A. 2012;109(42):2865-E2874.
この記事を経由して見る PubMed Google Scholar
Nguyen HCB, et al. HDAC3活性の二律背反は、炎症反応を支配する。Nature. 2020;584(7820):286–290.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Leus NGJ, et al. HDAC 3選択的阻害剤RGFP966は、RAW264.7マクロファージおよびマウス精密切断肺スライスにおいて、NF-κB p65転写活性を減弱することにより抗炎症性を発揮する。Biochem Pharmacol. 2016;108:58-74.
この記事を経由して見る CrossRef PubMed Google Scholar
Jamwal DR, et al. 腸管上皮のMHCII発現は、マウスの化学物質、T細胞誘発性、および感染性大腸炎の重症度を決定する。Gastroenterology. 2020;159(4):1342–1356.
この記事を経由して見る CrossRef PubMed Google Scholar
Thelemann C, et al. インターフェロン-γは腸管上皮細胞のMHCクラスIIの発現を誘導し、マウスを大腸炎から保護する。PLoS One. 2014;9(1):1-10.
この記事を経由して見る PubMed Google Scholar
Hashimoto K, et al. 主要組織適合性IA-β鎖遺伝子の条件付きヌルアレル。Genesis. 2002;32(2):152-153.
この記事を経由して表示する CrossRef PubMed Google Scholar
El Marjou F, et al.腸上皮における組織特異的かつ誘導可能なCre媒介組換え。Genesis. 2004;39(3):186-193.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Hong SW, et al. 食物の免疫寛容は、CD4+ T細胞の機能不全の層によって媒介される。Nature. 2022;607(7920):762–768.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Pedersen TK, et al.クローン病において、常在菌由来エピトープに対するCD4+ T細胞応答が、寛容状態から炎症状態へと移行する。Immunity. 2022;55:1-15.
この記事を経由して見る CrossRef PubMed Google Scholar
Zegarra-Ruiz DF, et al.腸内細菌特異的T細胞の胸腺の発達。Nature. 2021;594(7863):413–417.
この記事を経由して表示する。CrossRef PubMed Google Scholar
ベルカイドY、ハンドTW. 免疫と炎症における微生物叢の役割。Cell. 2014;157(1):121-141.
この記事を経由して見る CrossRef PubMed Google Scholar
Xu M, et al. c-MAF依存性制御性T細胞は、腸内病原体に対する免疫寛容を媒介する。Nature. 2018;554(7692):373–377.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Yang Y, et al. 常在菌抗原に対する腸管TH17細胞の焦点化特異性. Nature. 2014;510(7503):152–156.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Blackman M, et al. 発育中のT細胞の正と負の選択におけるT細胞受容体の役割。科学。1990;248(15):1335–1341.
この記事を経由して表示します。PubMed Google Scholar
Mathis D, Benoist C. Aire. Annu Rev Immunol。2009;27:287-312.
この記事を経由して表示します。CrossRef PubMed Google Scholar
T細胞レパートリーのポジティブおよびネガティブ選択:胸腺細胞は何を見ているのか(そして見ていないのか)。Nat Rev Immunol. 2014;14(6):377-391.
この記事を経由して見る CrossRef PubMed Google Scholar
Hepworth MR, et al. 自然リンパ系細胞は腸内常在菌に対するCD4+ T細胞応答を制御する。Nature. 2013;498(7452):113–117.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Hepworth MR, et al.免疫寛容。グループ3自然リンパ系細胞は、常在菌特異的CD4+ T細胞の腸管選択を媒介する。Science. 2015;348(6238):1031–1035.
この記事を経由して見る CrossRef PubMed Google Scholar
Lyu M, et al. ILC3s select microbiota-specific regulatory T cells to establish tolerance in the gut.腸内細菌特異的制御性T細胞は、腸内細菌を選択する。Nature. 2022;610(7933):744–751.
この記事を経由して表示します。CrossRef PubMed Google Scholar
赤祖父B、他。 新規抗原提示細胞は、腸内細菌叢に対するTreg依存的な寛容を与える。Nature. 2022;610(7933):752–760.
この記事を経由して表示する。CrossRef PubMed Google Scholar
RORγt+細胞は、腸内細菌に特異的なTreg細胞の分化を誘導する。Nature. 2022;610(7933):737–743.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Westendorf AM, et al. CD4+Foxp3+制御性T細胞の拡張は、局所樹状細胞とは無関係に、腸管上皮細胞との抗原駆動型相互作用によって誘導される。Gut. 2009;58(2):211-219.
この記事を経由して表示する CrossRef PubMed Google Scholar
大腸上皮細胞を介したCD4 T細胞活性化の抑制。Gut. 2004;53(5):678-684.
この記事を経由して表示します。CrossRef PubMed Google Scholar
ヒト十二指腸上皮細胞は、抗原提示の分子コンポーネントを構成的に発現しているが、コスティミュレーション分子は発現していない。Hum Immunol. 2002;63(11):977-986.
この記事を経由して見る CrossRef PubMed Google Scholar
Heuberger C, et al.なぜ腸管上皮細胞はMHCクラスIIを発現するのか?Immunology. 2021;162(4):357-367.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Sanderson IR, et al. 腸管細胞およびリンパ系細胞におけるB7 mRNAの制御の違い。Immunology. 1993;79(3):434-438.
この記事を表示する。PubMed Google Scholar
T細胞が介在する腸の炎症におけるインターロイキン17Aの保護機能。Nat Immunol. 2009;10(6):603-609.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Ouyang W, et al. 炎症におけるTヘルパー17細胞エフェクターサイトカインの生物学的機能. Immunity. 2008;28(4):454-467.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Schmidt EGW, et al. 実験的大腸炎におけるTH17細胞の誘導とIL-17AおよびIL-17F遮断の効果. Inflamm Bowel Dis. 2013;19(8):1567–1576.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Tang C, et al. IL-17Aの抑制ではなく、IL-17Fの抑制は、腸内細菌叢の改変によるTreg細胞の誘導によって大腸炎に対する保護を提供します。Nat Immunol. 2018;19(7):755-765.
この記事を経由して見る CrossRef PubMed Google Scholar
Feng T, et al. Th17細胞は大腸炎を誘発し、IL-17による自然免疫IL-12およびIL-23産生の誘導を通じてTh1細胞応答を促進します。J Immunol. 2011;186(11):6313–6318.
この記事を経由して見る CrossRef PubMed Google Scholar
RORγを発現するTh17細胞は、IL-17AとIL-17Fの重複効果によりマウス慢性腸炎を誘発する。Gastroenterology. 2009;136(1):257-267.
この記事を見るには CrossRef PubMed Google Scholar
Hueber W, et al. ヒト抗IL-17Aモノクローナル抗体Secukinumabによる中等症から重症のクローン病の治療:無作為化二重盲検プラセボ対照試験の予想外の結果です。Gut. 2012;61(12):1693–1700.
この記事を経由して表示する。CrossRef PubMed Google Scholar
ROR-γtの一過性の阻害は、TH17細胞を減少させ、グループ3自然リンパ系細胞を維持することにより、治療的に腸の炎症を制限する。Nat Med. 2016;22(3):319-323.
この記事を経由して見る CrossRef PubMed Google Scholar
Blumberg RS, et al.腸管上皮細胞による抗原提示。Immunol Lett. 1999;69(1):7-11.
この記事を経由して表示します。CrossRef PubMed Google Scholar
Van Der Kraak LA, et al. IL-18の遺伝的および通性誘導は、IFNγを介して腸管上皮MHCIIを駆動する。Mucosal Immunol. 2021;14(5):1100–1112.
この記事を経由して見る CrossRef PubMed Google Scholar
Rakoff-Nahoum S, et al.腸のホメオスタシスに必要なtoll様受容体による常在菌の微生物叢の認識. Cell. 2004;118:229-241.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Rhee SH, et al. 大腸の炎症における細菌フラジェリンによるToll様受容体5の関与の病態生理学的役割。Proc Natl Acad Sci U S A. 2005;102(38):13610-13615.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Rose WAI, et al. TLR9は腸管損傷からの保護および腸管修復に重要である。サイ・レップ 2012;2:1-9.
この記事を経由して表示します。PubMed Google Scholar
Leus NGJ, et al. NF-κBを介した炎症における新たな創薬ターゲットとしてのヒストン脱アセチル化酵素3(hdac 3)。Curr Opin Chem Biol. 2016;33:160-168.
この記事を経由して見る PubMed Google Scholar
Zhu H, et al. ヒストンデアセチラーゼ3の活性化は、リポポリサッカライド刺激時の心筋細胞における腫瘍壊死因子-α(TNF-α)発現を促進する。J Biol Chem. 2010;285(13):9429–9436.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Greer CB, et al. ヒストン脱アセチル化酵素は、伸長機構を介して転写を正に制御している。Cell Rep. 2015;13(7):1444-1455.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Lewandowski SL, et al. ヒストンデアセチラーゼ3は、トランスフォーミング成長因子-β1(TGF-β1)のデアセチラーゼ非依存性エピジェネティックサイレンシングを調整して、第二心野形成をオーケストレーションする。J Biol Chem. 2015;290(45):27067–27089.
この記事を経由して見る CrossRef PubMed Google Scholar
Emmett MJ, et al. ヒストン脱アセチル化酵素3は、褐色脂肪組織を急性熱発生チャレンジに準備させる。Nature. 2017;546(7659):544–548.
この記事を経由して見る CrossRef PubMed Google Scholar
Wang Y, et al.腸内細菌叢はNFIL3と概日時計を介して体組成を調節している。Science. 2017;916(357):912–916.
この記事を経由して表示する。PubMed Google Scholar
Tamoutounour S, et al. ケラチノサイト内在性MHCII発現は、マイクロバイオータ誘導Th1細胞応答を制御する。Proc Natl Acad Sci U S A. 2019;116(47):23643-23652.
この記事を経由して見る CrossRef PubMed Google Scholar
Kerdidani D, et al. 肺腫瘍MHCII免疫は、線維芽細胞によるin situ抗原提示に依存する。J Exp Med. 2022;219(2):e20210815.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Shenoy AT, et al. 肺上皮細胞による抗原提示は、CD4+ TRM細胞の機能を指示し、バリア免疫を制御する。Nat Commun. 2021;12(1):5834.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Feng D, et al. ヒストン脱アセチル化酵素3によって編成される概日リズムは、肝脂質代謝を制御する。Science. 2011;331(6022):1315–1319.
この記事を経由して表示する。CrossRef PubMed Google Scholar
ヒストン脱アセチル化酵素3による心臓のエネルギー代謝の維持(Montgomery RL, et al. J Clin Invest. 2008;118(11):3588–3597.
この記事を見るには JCI CrossRef PubMed Google Scholar
Ning L, et al. 固形臓器損傷の病態におけるヒストン脱アセチル化酵素3の重要な役割. Cell Death Dis. 2021;12(8):1-13.
この記事を経由して見る PubMed Google Scholar
Shao TY, et al. Candida albicans oscillating UME6 expression during intestinal colonization primes systemic Th17 protective immunity.腸管コロニー形成時のカンジダ・アルビカンスのUME6発現は、全身性のTh17防御免疫を促進する。Cell Rep. 2022;39(7):110837.
この記事を経由して表示する。CrossRef PubMed Google Scholar
Igyártó BZ, et al.皮膚常在マウス樹状細胞サブセットは、別個かつ相反する抗原特異的Tヘルパー細胞応答を促進する。Immunity. 2011;35(2):260-272.
この記事を見るには CrossRef PubMed Google Scholar
Woo V, et al. 宿主の分節化した糸状菌由来のレチノイン酸は、腸管感染に対する宿主の防御を促進させる。Cell Host Microbe. 2021;29(12):1744–1756.
この記事を経由して見る CrossRef PubMed Google Scholar
ウッドDE、サルツバーグSL. Kraken: 正確なアラインメントを用いた超高速メタゲノム配列分類。Genome Biol. 2014;15(3):10.
この記事を経由して表示します。PubMed Google Scholar
vegan: Community Ecology Package(コミュニティエコロジーパッケージ)。Rパッケージバージョン2。Oksanen AJ, et al. 2022; https://cran.r-project.org/web/packages/vegan/index.html.
Lodes MJ, et al. Bacterial flagellin is a dominant antigen in Crohn disease(細菌性フラジェリンはクローン病における支配的な抗原である)。J Clin Invest. 2004;113(9):1296–1306.
この記事を経由して表示する JCI CrossRef PubMed Google Scholar
Buchfink B, Reuter K, Drost HG. DIAMONDを用いた生命の木スケールでの高感度タンパク質アラインメント。Nat Methods. 2021;18(4):366-368.
この記事を経由して表示する。CrossRef PubMed Google Scholar
バージョン履歴
バージョン1(2023年1月5日)。インプレスプレビュー
バージョン2(2023年2月15日)。電子出版
私たちは次のことを推奨します。
細菌性フラジェリンはクローン病における支配的な抗原である
マイケル・J・ロデスら、J Clin Invest誌、2004年
口腔内細菌叢が骨免疫調節作用を発揮する。
Jessica D. Hathaway-Schraderら、JCI Insight、2022年
微生物叢に敏感なエピジェネティックシグネチャーはクローン病の炎症を予測する
ダニエル・ケリーら、JCIインサイト、2018年
コレステロールの流入と流出のバランスが崩れたときに何が起こるかを学ぶ
出典はこちら DiscoverCholesterolEfflux.com|CMD-MY-0033 I 2022年11月号
HDAC3を介したLncRNA-LETの抑制は、MiR-548kを介して胃がん細胞の成長促進、侵入、移動、およびアポトーシスを制御する
Jie Zhangら、Journal of Environmental Pathology, Toxicology and Oncology(環境病理学、毒性学、腫瘍学)、2021年
健康と病気における非従来型腸管上皮内リンパ球
アリ・ナズミら、Crit Rev Immunol、2021年
を搭載
ツイッター フェイスブック RSS
著作権 © 2023 米国臨床研究学会
ISSN: 0021-9738(プリント), 1558-8238(オンライン)
メールアラートに登録する
あなたのEメールアドレス
この記事が気に入ったらサポートをしてみませんか?