IgG4 のユニークな特性と健康と病気におけるその役割


元→The unique properties of IgG4 and its roles in health and disease | Nature Reviews Immunology
The unique properties of IgG4 and its roles in health and disease
IgG4 のユニークな特性と健康と病気におけるその役割
Abstract
IgG4 is the least abundant subclass of IgG in human serum and has unique functional features.
IgG4 は、ヒト血清中の IgG の中で最も少ないサブクラスであり、独自の機能的特徴を持っています。
IgG4 is largely unable to activate antibody-dependent immune effector responses and, furthermore, undergoes Fab (fragment antigen binding)-arm exchange, rendering it bispecific for antigen binding and functionally monovalent.
IgG4 は、抗体依存性免疫エフェクター応答を活性化することがほとんどできず、さらに Fab (フラグメント抗原結合) アーム交換を受けて、抗原結合に対して二重特異性になり、機能的に一価になります。
These properties of IgG4 have a blocking effect, either on the immune response or on the target protein of IgG4.
IgG4 のこれらの特性は、免疫応答または IgG4 の標的タンパク質のいずれかに遮断効果をもたらします。
In this Review, we discuss the unique structural characteristics of IgG4 and how these contribute to its roles in health and disease.
このレビューでは、IgG4 のユニークな構造特性と、これらが健康と疾患におけるその役割にどのように寄与しているかについて説明します。
We highlight how, depending on the setting, IgG4 responses can be beneficial (for example, in responses to allergens or parasites) or detrimental (for example, in autoimmune diseases, in antitumour responses and in anti-biologic responses).
設定に応じて、IgG4 応答がどのように有益 (たとえば、アレルゲンや寄生体への応答) または有害 (たとえば、自己免疫疾患、抗腫瘍応答、および抗生物学的応答) になるかを強調します。
The development of novel models for studying IgG4 (patho)physiology and understanding how IgG4 responses are regulated could offer insights into novel treatment strategies for these IgG4-associated disease settings.
IgG4 (病態) 生理学を研究し、IgG4 応答がどのように調節されているかを理解するための新しいモデルの開発は、これらの IgG4 関連疾患設定に対する新しい治療戦略への洞察を提供する可能性があります。
Introduction
Humoral (antibody-mediated) immune responses are important for protection against pathogen invasion but can also cause disease.
体液性 (抗体媒介性) 免疫応答は、病原体の侵入に対する防御に重要ですが、病気の原因にもなります。
Antibodies recognize and bind specific structures of pathogens through their Fab (fragment antigen binding) arms.
抗体は、Fab (フラグメント抗原結合) アームを介して病原体の特定の構造を認識して結合します。
In addition to mediating direct neutralization of the pathogen, this opsonization of pathogen structures can result in the activation of various immune effector pathways through the Fc (fragment crystallizable) region of antibodies.
病原体の直接的な中和を媒介することに加えて、病原体構造のこのオプソニン化は、抗体の Fc (フラグメント結晶化可能) 領域を介したさまざまな免疫エフェクター経路の活性化をもたらす可能性があります。
The antibody Fc region interacts with Fc receptors on immune cells such as macrophages, neutrophils and natural killer cells, resulting in antibody-dependent cell-mediated cytotoxicity (ADCC) or antibody-dependent cellular phagocytosis (ADCP).
抗体の Fc 領域は、マクロファージ、好中球、ナチュラル キラー細胞などの免疫細胞上の Fc 受容体と相互作用し、抗体依存性細胞媒介性細胞傷害 (ADCC) または抗体依存性細胞食作用 (ADCP) を引き起こします。
The antibody Fc region can also interact with the complement system, resulting in antibody-dependent complement deposition, which further primes pathogens for cellular uptake and destruction.
抗体の Fc 領域は補体系と相互作用することもでき、その結果、抗体依存性の補体沈着が起こり、病原体の細胞への取り込みと破壊をさらに促進します。
The ability of an antibody to elicit these immune responses depends on the type of Fc tail and modifications thereof (for example, glycosylation).
これらの免疫応答を誘発する抗体の能力は、Fcテールのタイプとその修飾(例えば、グリコシル化)に依存します。
In humans, five classes of antibody are recognized based on their Fc tail: IgM, IgD, IgE, IgA and IgG.
ヒトでは、Fc テイルに基づいて、IgM、IgD、IgE、IgA、および IgG の 5 つのクラスの抗体が認識されます。
IgM antibodies are produced in the early stages of a primary, adaptive immune response.
IgM 抗体は、一次適応免疫応答の初期段階で産生されます。
IgM and IgD form the B cell receptors (BCRs) on naive B cells.
IgM と IgD は、ナイーブ B 細胞の B 細胞受容体 (BCR) を形成します。
The primary IgM response can be followed by a second, long-lasting wave of IgG or IgA antibodies, the latter being involved particularly in mucosal immune responses.
一次 IgM 応答に続いて、長期にわたる IgG または IgA 抗体の第 2 波が発生する可能性があり、後者は特に粘膜免疫応答に関与しています。
IgE probably evolved as a defence against parasitic worms and is also involved in various allergic diseases.
IgE はおそらく寄生虫に対する防御として進化し、さまざまなアレルギー疾患にも関与しています。
Such mature antibody responses are produced by antigen-stimulated B cells that have undergone a process known as class-switch recombination, in which a B cell rearranges its DNA to produce another class of antibody with the same specificity.
このような成熟抗体応答は、クラススイッチ組換えとして知られるプロセスを経た抗原刺激 B 細胞によって生成されます。 B 細胞がその DNA を再構成して、同じ特異性を持つ別のクラスの抗体を生成します。
B cells can undergo repeated rounds of class-switch recombination until the DNA has been recombined using the most 3′ Fc tail gene segment (Fig. 1).
B細胞は、最も3'側のFcテール遺伝子セグメントを使用してDNAが組換えられるまで、クラススイッチ組換えのラウンドを繰り返すことができます(図1)。

Four subclasses of IgG exist, numbered according to the order of abundance.
IgG には 4 つのサブクラスがあり、豊富な順に番号が付けられています。
IgG1 has the largest relative contribution to total IgG, followed by IgG2 then IgG3 and IgG4.
IgG1 は総 IgG に対する相対的な寄与が最も大きく、次に IgG2、IgG3、IgG4 の順です。
Although IgG4 is the least abundant IgG subclass overall, specific responses can be dominated by IgG4, often associated with chronic or repeated antigen exposure.
IgG4 は全体的に最も少ない IgG サブクラスですが、特定の応答は IgG4 によって支配される可能性があり、多くの場合、慢性的または反復的な抗原曝露に関連しています。
IgG4 has a unique set of properties compared with the other IgG subclasses that has led to IgG4 being widely regarded as an anti-inflammatory, ‘benign’ antibody that may have beneficial functions in allergic disease (Table 1).
IgG4 は他の IgG サブクラスと比較して独自の一連の特性を持っているため、IgG4 はアレルギー疾患において有益な機能を持つ可能性のある抗炎症性の「良性」抗体として広く見なされています (表 1)。

However, evidence is accumulating that IgG4 also has a pathogenic role in a range of diseases.
しかし、IgG4 がさまざまな疾患において病原性の役割も果たしているという証拠が蓄積されています。
Research in the past decade has shown that IgG4 can have detrimental roles in IgG4 autoimmune diseases (IgG4-AIDs), in tumour immunology and in IgG4-related diseases (IgG4-RDs).
過去 10 年間の研究は、IgG4 が IgG4 自己免疫疾患 (IgG4-AID)、腫瘍免疫学、および IgG4 関連疾患 (IgG4-RD) において有害な役割を果たしている可能性があることを示しています。
The IgG4-AIDs and IgG4-RDs are chronic conditions and for most patients no cure currently exists.
IgG4-AID と IgG4-RD は慢性疾患であり、現在ほとんどの患者にとって治療法はありません。
Moreover, the increasing use of biological therapies warrants a better understanding of why certain drugs elicit IgG4 anti-drug responses that limit their efficacy.
さらに、生物学的療法の使用の増加は、特定の薬物がその有効性を制限する IgG4 抗薬物応答を誘発する理由をよりよく理解することを保証します。
In this Review, we highlight how the unique structural and functional characteristics of IgG4 contribute to disease onset and progression in these settings.
このレビューでは、IgG4 のユニークな構造的および機能的特性が、これらの設定で疾患の発症と進行にどのように寄与するかを強調します。
Furthermore, we provide an overview of our current understanding of how IgG4 responses are regulated.
さらに、IgG4 応答がどのように調節されているかについての現在の理解の概要を提供します。
By understanding these processes, future therapeutic strategies could be shaped to prevent pathogenic IgG4 responses or induce beneficial IgG4 responses.
これらのプロセスを理解することで、病原性 IgG4 応答を防止したり、有益な IgG4 応答を誘導したりするための将来の治療戦略を立てることができます。
Although we appreciate that most antibody responses involve a range of different (sub)classes, in this Review we focus on IgG4-associated diseases that have a predominant IgG4 (antigen-specific) response.
ほとんどの抗体応答にはさまざまな (サブ) クラスが関与していることを理解していますが、このレビューでは、IgG4 (抗原特異的) 応答が優勢な IgG4 関連疾患に焦点を当てています。
Development of an IgG4 response
IgG4 応答の発生
The production of IgG4 requires that B cells undergo class-switch recombination (Fig. 1).
IgG4 の産生には、B 細胞がクラススイッチ組換えを受ける必要があります (図 1)。

A direct switch from IgM to IgG4 can occur, but indirect switching through, for example, IgG1 is also — at least theoretically — possible.
IgM から IgG4 への直接的な切り替えが発生する可能性がありますが、たとえば IgG1 を介した間接的な切り替えも、少なくとも理論的には可能です。
Although direct evidence is lacking, in BCR repertoire analyses only limited clonal overlap between IgG1 and IgG4 responses has been observed, which suggests that indirect switching to IgG4 via IgG1 is a minor route in vivo.
直接的な証拠は不足していますが、BCR レパートリー分析では、IgG1 と IgG4 応答の間の限られたクローン重複のみが観察されており、これは、IgG1 を介した IgG4 への間接的な切り替えが in vivo でのマイナーな経路であることを示唆しています。
In keeping with this, in vitro, naive IgM+ B cells readily switch towards IgG4 production, whereas IgG1+ memory B cells do not.
これに合わせて、in vitro では、ナイーブ IgM+ B 細胞は IgG4 産生に容易に切り替わりますが、IgG1+ 記憶 B 細胞はそうではありません。
Interestingly, substantial clonal overlap between IgG4 and IgA2 was observed, which may reflect that common food antigens often induce both IgG4 and IgA2 responses.
興味深いことに、IgG4 と IgA2 の間にかなりのクローン重複が観察されました。これは、一般的な食品抗原が IgG4 と IgA2 の両方の応答を誘導することが多いことを反映している可能性があります。
This could indicate either that there are similar requirements for the development of IgA2 and IgG4 responses or that substantial sequential class-switching from IgG4 to IgA2 occurs.
これは、IgA2 および IgG4 応答の発生に同様の要件があること、または IgG4 から IgA2 への実質的な連続クラス切り替えが発生することを示している可能性があります。
Class-switching from IgE to IgG4 is not possible owing to the order of class-switch elements in the genome (the heavy-chain constant region segments for IgE being downstream (3′) of those for IgG4).
IgE から IgG4 へのクラス スイッチは、ゲノム内のクラス スイッチ エレメントの順序により不可能です (IgE の重鎖定常領域セグメントは IgG4 のセグメントの下流 (3') にあります)。
Therefore, the allergen-specific IgG4 that is induced by specific immunotherapy in patients with IgE-mediated allergic disease must be derived from either precursor B cells capable of switching to both IgE and IgG4 (for example, non-switched or IgG1+ memory B cells) or newly recruited (naive) B cells.
したがって、IgE 媒介性アレルギー疾患患者の特異的免疫療法によって誘導されるアレルゲン特異的 IgG4 は、IgE と IgG4 の両方に切り替えることができる前駆 B 細胞(たとえば、非スイッチまたは IgG1+ メモリー B 細胞) または新しく動員された (ナイーブ) B 細胞のいずれかに由来する必要があります。
Class-switching towards IgG4 is mostly associated with T helper 2 (TH2) cell responses.
IgG4 へのクラス切り替えは、主に T ヘルパー 2 (TH2) 細胞応答に関連しています。
The type 2 cytokines IL-4 and/or IL-13 are important for the induction of IgG in general, but class-switching to IgG4 may more strictly depend on these cytokines than does class-switching to IgG1.
タイプ 2 サイトカイン IL-4 および/または IL-13 は、一般に IgG の誘導に重要ですが、IgG4 へのクラス切り替えは、IgG1 へのクラス切り替えよりもこれらのサイトカインに厳密に依存する可能性があります。
IL-10 and regulatory T (Treg) cells may also skew the antibody response towards IgG4 (relative to IgE and, possibly, also IgG1) (Fig. 1).
IL-10 および制御性 T (Treg) 細胞は、抗体応答を IgG4 (IgE および、おそらく IgG1 に対しても) に向けてゆがめる可能性もあります (図 1)。
However, the role of IL-10 is not fully clarified, and studies in vitro have yielded conflicting results depending on, amongst other factors, which cell types are present in addition to naive B cells (Box 1).
しかし、IL-10 の役割は完全には解明されておらず、in vitro での研究では、ナイーブ B 細胞に加えてどの細胞タイプが存在するかなどの要因に応じて、相反する結果が得られています (ボックス 1)。
IgG4-switched B cells have similar potential for terminal differentiation towards antibody-secreting cells to that of IgG1-switched B cells; hence, limitations in the development of an IgG4 antibody response are not owing to intrinsic limitations of IgG4+ B cells.
IgG4 スイッチ B 細胞は、IgG1 スイッチ B 細胞と同様の抗体分泌細胞への最終分化の可能性を持っています;したがって、IgG4 抗体応答の発生における制限は、IgG4+ B 細胞の固有の制限によるものではありません。
However, IgG4-switched B cells differ phenotypically from IgG1-switched B cells in several aspects.
ただし、IgG4 スイッチ B 細胞は、いくつかの側面で IgG1 スイッチ B 細胞とは表現型が異なります。
In particular, they have an altered chemokine receptor profile with lower levels of expression of CXCR3, CXCR4, CXCR5, CCR6 and CCR7 — chemokine receptors involved in germinal centre reactions and the generation of long-lived plasma cells.
特に、CXCR3、CXCR4、CXCR5、CCR6、およびCCR7の発現レベルが低く、ケモカイン受容体プロファイルが変化しています — 胚中心反応および長寿命形質細胞の生成に関与するケモカイン受容体。
In the circulation, numbers of IgG4+ B cells reflect serum IgG4 concentrations, and their levels follow similar patterns throughout life.
循環中の IgG4+ B 細胞の数は、血清 IgG4 濃度を反映しており、そのレベルは生涯を通じて同様のパターンに従います。
IgG4+ cell numbers in blood are low compared with IgG1+ cells and have a relatively low abundance in secondary lymphoid organs.
血液中の IgG4+ 細胞の数は IgG1+ 細胞に比べて少なく、二次リンパ器官の量は比較的少ないです。
Furthermore, IgG4 production by antibody-secreting cells can be markedly shorter lived than for other IgG subclasses, requiring continuous input from newly differentiating B cells.
さらに、抗体分泌細胞による IgG4 産生は、他の IgG サブクラスよりも著しく短命である可能性があり、新たに分化する B 細胞からの継続的なインプットが必要です。
Indeed, rituximab (anti-CD20) therapy for B cell depletion has been shown to be particularly beneficial in autoimmune diseases characterized by pathogenic IgG4 (auto)antibodies.
実際、B 細胞枯渇に対するリツキシマブ (抗 CD20) 療法は、病原性 IgG4 (自己) 抗体を特徴とする自己免疫疾患に特に有益であることが示されています。
During infancy, the proportion of IgG4 in circulation rises slowly with age, and IgG4 titres generally continue to increase throughout life until the fifth decade, after which a small gradual decline is observed.
乳児期に、循環中の IgG4 の割合は年齢とともにゆっくりと上昇します。IgG4力価は一般に、50代まで生涯を通じて増加し続けます。 その後、緩やかな減少が見られます。
Serum levels of IgG4 show great variation in the healthy population, although intra-individual levels are generally stable.
IgG4 の血清レベルは、健康な集団では大きな変動を示しますが、個人内のレベルは一般的に安定しています。
One of the hallmarks of most IgG4 responses is that they develop slowly over time for reasons that are not well understood.
ほとんどの IgG4 応答の顕著な特徴の 1 つは、十分に理解されていない理由で時間の経過とともにゆっくりと発生することです。
Prolonged or repeated exposure to antigen seems to be a necessary — but not sufficient — factor for the development of an IgG4-dominated response.
抗原への長期的または反復的な曝露は、IgG4 優位の応答の発生に必要な要素ですが、十分ではないようです。
For example, individuals hyperimmunized with tetanus toxoid have an IgG1-dominated response with little IgG4 despite repeated antigen exposure, whereas individuals repeatedly vaccinated with SARS-CoV-2 mRNA were shown, in some cases, to have increased proportions of IgG4 after a third vaccination, requiring at least 6 months to develop.
例えば、破傷風トキソイドで過剰免疫された個体は、繰り返し抗原に曝露されたにもかかわらず、IgG1 優位の応答を示し、IgG4 はほとんどありません。いくつかのケースでは、SARS-CoV-2 mRNAを繰り返しワクチン接種した個人が示されました。3 回目のワクチン接種後に IgG4 の割合が増加し、発生までに少なくとも 6 か月を要します。
IgG4 is not commonly part of the antibody response to bacterial or viral infection.
IgG4 は通常、細菌またはウイルス感染に対する抗体反応の一部ではありません。
The range of situations in which specific IgG4 is or can be a dominant factor is wide and includes responses to allergens, therapeutically administered proteins, autoantigens and helminth infections.
特定の IgG4 が支配的な因子である、または支配的な因子である可能性がある状況の範囲は広く、アレルゲン、治療的に投与されたタンパク質、自己抗原、および蠕虫感染への応答が含まれます。
With the exception of helminths, the absence of an infectious agent seems to be a common feature of IgG4 responses and it is tempting to speculate that the absence of certain danger signals such as pathogen-associated molecular patterns (PAMPs) is a prerequisite for B cells to differentiate towards IgG4-secreting cells in vivo.
蠕虫を除いて、感染病原体が存在しないことは IgG4 応答の一般的な特徴のようです。また、病原体関連分子パターン (PAMP) などの特定の危険信号が存在しないことが、in vivo で B 細胞が IgG4 分泌細胞に分化するための前提条件であると推測しがちです。
Indeed, the proportion of IgG4 antibodies was smaller in individuals receiving whole-cell pertussis vaccine than in individuals receiving acellular pertussis vaccine (although IgG4 was only a small fraction of the total IgG response even in the latter).
実際、IgG4 抗体の割合は、無細胞百日咳ワクチンを投与された個人よりも全細胞百日咳ワクチンを投与された個人の方が小さかった (ただし、IgG4 は後者の場合でも全 IgG 応答のごく一部に過ぎなかった)。
Structure and function of IgG4
IgG4の構造と機能
Despite the high levels of homology between human IgG subclasses, each subclass has a specific set of functional characteristics owing to particular structural features (Table 1).
ヒト IgG サブクラス間の高いレベルの相同性にもかかわらず、各サブクラスは、特定の構造上の特徴により、特定の一連の機能特性を持っています (表 1)。

IgG4 is unique in that it has a lesser affinity than other IgG subclasses for many effector molecules, such as Fc receptors and complement, and also because of structural features that affect interactions through the Fab region, such as Fab-arm exchange and a greater propensity for acquiring glycosylation in the variable domains (Fig. 2).
IgG4 は、Fc 受容体や補体などの多くのエフェクター分子に対して、他の IgG サブクラスよりも親和性が低いという点で独特です。また、Fabアーム交換や可変ドメインでグリコシル化を獲得する傾向が高いなど、Fab領域を介した相互作用に影響を与える構造的特徴のためでもあります(図2)。

Fc-dependent effector functions
Fc依存エフェクター機能
IgG4 differs in several key amino acid positions from the other IgG subclasses, resulting in a modified binding pattern to Fcγ receptors (Table 1 and Fig. 2).
IgG4 はいくつかの重要なアミノ酸位置が他の IgG サブクラスと異なり、その結果、Fcγ 受容体への結合パターンが変化します (表 1 および図 2)。
In particular, relative to IgG1, the amino acid changes at L234F, A327G and P331S in IgG4 are implicated in effects on Fcγ receptor binding.
特に、IgG1 と比較して、IgG4 の L234F、A327G、および P331S でのアミノ酸変化は、Fcγ 受容体結合への影響に関与しています。
Binding to most Fcγ receptors is reduced for IgG4 (although not completely abrogated), resulting in IgG4 having poor ADCC activity and, probably, also ADCP activity (although this has not been studied in detail).
IgG4 ではほとんどの Fcγ 受容体への結合が減少し (完全に無効化されたわけではありません)、その結果、IgG4 の ADCC 活性は低下し、おそらく ADCP 活性も低下します (ただし、これは詳細には研究されていません)。
Interestingly, binding of IgG4 to the inhibitory receptor FcγRIIb is not affected, which skews Fcγ receptor signalling induced by IgG4 away from cellular activation and towards inhibition.
興味深いことに、抑制性受容体 FcγRIIb への IgG4 の結合は影響を受けず、これは IgG4 によって誘導される Fcγ 受容体シグナル伝達を細胞の活性化から阻害へとゆがめます。
In the case of IgG1, interaction with the activating receptor FcγRIIIa is markedly increased if the conserved Fc glycan does not contain the core fucose moiety.
IgG1 の場合、保存された Fc グリカンがコア フコース部分を含まない場合、活性化受容体 FcγRIIIa との相互作用が著しく増加します。
This is also true for IgG4, and an afucosylated variant of IgG4 was found to induce ADCC, albeit still less efficiently than did IgG1.
これは IgG4 にも当てはまり、IgG4 のアフコシル化バリアントが ADCC を誘導することがわかりましたが、それでも IgG1 ほど効率的ではありませんでした。
However, naturally occurring afucosylated antibody responses seem to be restricted to antiviral responses or alloimmunity to blood cells and platelets, which normally do not have high levels of IgG4.
しかし、天然に存在するアフコシル化抗体応答は、抗ウイルス応答または血液細胞および血小板に対する同種免疫に限定されているようです。通常、高レベルの IgG4 はありません。
The limited signalling of IgG4 through activating Fc receptors might attenuate the impact of certain autoantibody responses; for example, it has recently been shown in a mouse model of thrombotic thrombocytopenic purpura that recombinant IgG1 antibodies to ADAMTS13 (a disintegrin and metalloproteinase with thrombospondin motifs 13) are more pathogenic than their IgG4 counterparts in an Fcγ-dependent manner.
Fc受容体の活性化によるIgG4の制限されたシグナル伝達は、特定の自己抗体応答の影響を弱める可能性があります;例えば、最近、血栓性血小板減少性紫斑病のマウスモデルにおいて、ADAMTS13 (トロンボスポンジンモチーフを有するディスインテグリンおよびメタロプロテイナーゼ 13) に対する組換え IgG1 抗体が、Fcγ 依存的に対応する IgG4 抗体よりも病原性が高いことが示されました。
In addition to the ‘classical’ Fcγ receptors (FcγRI–FcγRIII), two ‘non-classical’ Fcγ receptors — Fc receptor-like protein 4 (FCRL4) and FCRL5 — have been reported to bind IgG4, albeit weakly.
「古典的な」Fcγ 受容体 (FcγRI–FcγRIII) に加えて、2 つの「非古典的な」Fcγ 受容体 — Fc 受容体様タンパク質 4 (FCRL4) および FCRL5 — は弱いながらも IgG4 に結合することが報告されています。
These receptors are mostly expressed on B cells and have been described to either inhibit or enhance BCR signalling, the latter only if CD21 is simultaneously engaged, thereby augmenting or counteracting the role of FcγRIIb.
これらの受容体は主に B 細胞で発現し、BCR シグナル伝達を阻害または増強することが報告されていますが、後者は CD21 が同時に関与している場合にのみ、FcγRIIb の役割を増強または対抗します。
A specific role of IgG4 in this signalling route is as yet unknown.
このシグナル伝達経路における IgG4 の特定の役割は、まだ不明です。
Furthermore, IgG4 is a poor activator of complement, resulting in a poor capacity for inducing antibody-dependent complement deposition and ADCP.
さらに、IgG4 は補体の活性化が不十分なため、抗体依存性の補体沈着および ADCP を誘導する能力が低くなります。
Complement has an important role in clearing pathogens and promoting inflammation, which consequently is limited when IgG4 dominates the antibody response.
補体は、病原体の除去と炎症の促進に重要な役割を果たします。したがって、IgG4 が抗体応答を支配する場合、炎症は制限されます。
This results mainly from reduced binding to C1q, caused mainly by residue S331 of IgG4, which is the counterpart of P331 in IgG1 (the homologous P436S mutation in IgM also markedly affects C1q binding).
これは主に、IgG1のP331の対応物であるIgG4の残基S331によって主に引き起こされるC1qへの結合の減少に起因します(IgMの相同P436S変異もC1q結合に著しく影響します)。
Nevertheless, some studies suggest that IgG4 can activate complement in specific contexts.
それにもかかわらず、いくつかの研究は、IgG4 が特定の状況で補体を活性化できることを示唆しています。
For example, artificially enforcing the hexamerization of IgG4 — a process that normally would take place ‘spontaneously’ as part of complex formation of antibody with C1 — results in complement activation by the classical route.
たとえば、IgG4 の 6 量体化を人為的に強制すると (これは通常、C1 との抗体の複合体形成の一部として「自然に」起こるプロセスです)、古典的な経路による補体の活性化をもたらします。
This shows that IgG4 has a reduced ability to activate complement but is not completely ‘silent’ in this respect.
これは、IgG4 が補体を活性化する能力が低下していることを示していますが、この点で完全に「沈黙」しているわけではありません。
In patients with membranous nephropathy, all of whom have complement deposition in the kidneys, IgG4 autoantibodies to phospholipase A2 receptor 1 (PLA2R1) are associated with disease.
膜性腎症の患者では、その全員が腎臓に補体沈着を起こしています。 ホスホリパーゼ A2 受容体 1 (PLA2R1) に対する IgG4 自己抗体は、疾患に関連しています。
Recent work indicates a possible role for these IgG4 autoantibodies in complement activation via the lectin pathway, whereby decreased galactosylation levels on the autoantibodies allow for mannose-binding lectin (MBL) binding and complement deposition.
最近の研究は、レクチン経路を介した補体活性化におけるこれらの IgG4 自己抗体の役割の可能性を示しています。 これにより、自己抗体のガラクトシル化レベルが低下すると、マンノース結合レクチン(MBL)の結合と補体の沈着が可能になります。
This is in contrast to increased levels of galactosylation promoting IgG1 hexamerization and complement activation by the classical route, and other studies suggest a pathogenic role of IgG4 autoantibodies to PLA2R1 independent of complement.
これは、従来の経路による IgG1 六量体化および補体活性化を促進するガラクトシル化レベルの増加とは対照的です。他の研究は、補体とは無関係にPLA2R1に対するIgG4自己抗体の病原的役割を示唆しています。
Complement activation could be shown in vitro for glyco-engineered recombinant IgG4 antibodies, but only at high antigen density and high antibody concentration, and no contribution of the lectin pathway was observed.
補体の活性化は、糖操作された組換え IgG4 抗体の in vitro で示されましたが、高抗原密度および高抗体濃度でのみ示され、レクチン経路の寄与は観察されませんでした。
Fab-arm exchange
Fabアームの交換
Uniquely, serum IgG4 typically does not cross-link antigen.
ユニークなことに、血清 IgG4 は通常、抗原を架橋しません。
In fact, IgG4 often behaves effectively as a monovalent antibody in the circulation.
実際、IgG4 は循環中において一価抗体として効果的に作用することがよくあります。
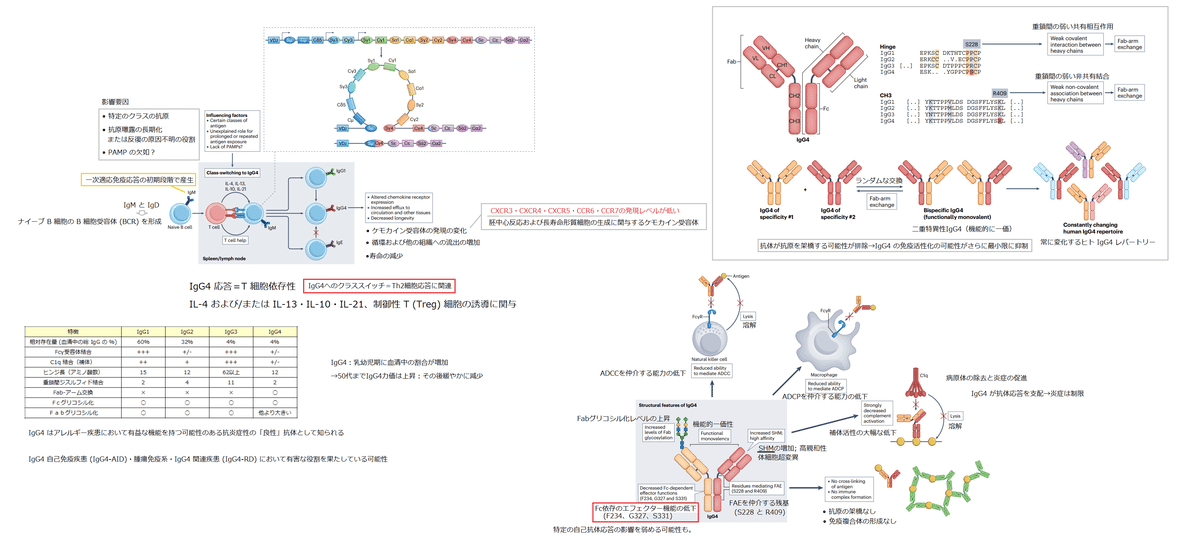
IgG4 molecules are produced as bivalent, monospecific antibodies but can, subsequently, engage in a process in which half-molecules of IgG4 (heavy chain and light chain) are randomly exchanged with other IgG4 half-molecules, through a process known as Fab-arm exchange (Box 2).
IgG4分子は、二価の単一特異性抗体として産生されます。しかし、続いて、Fab アーム交換として知られるプロセスを通じて、IgG4 の半分子 (重鎖と軽鎖) が他の IgG4 半分子とランダムに交換されるプロセスに関与することができます (ボックス 2)。
This makes most IgG4 molecules in the blood bispecific.
これにより、血液中のほとんどの IgG4 分子が二重特異性になります。
Early evidence of this process included the ability of serum IgG4 to cross-link two different allergens.
このプロセスの初期の証拠には、血清 IgG4 が 2 つの異なるアレルゲンを架橋する能力が含まれていました。
However, in many cases, the second antigen specificity of an IgG4 molecule will be irrelevant because the exchange is random, and the resulting antibody will behave as if monovalent.
しかし、多くの場合、IgG4 分子の 2 番目の抗原特異性は、交換がランダムであるため無関係であり、得られる抗体は一価のように振る舞います。
Thus, effective binding and downstream signalling of functionally monovalent IgG4 will require high affinity for antigen, as IgG4 cannot benefit from the accumulated binding strength (avidity) of multiple Fabs with the same specificity.
したがって、機能的に一価の IgG4 の効果的な結合と下流のシグナル伝達には、抗原に対する高い親和性が必要です。IgG4 は、同じ特異性を持つ複数の Fab の蓄積された結合強度 (アビディティ) の恩恵を受けることができないためです。
Interestingly, in patients with eosinophilic oesophagitis, very high titres of specific IgG4 to cow milk protein have been observed (together with deposits of IgG4 in the oesophageal wall), to such a degree that a substantial portion of IgG4 may still be bivalent in this context; this suggests that the effective monovalency of IgG4 is not absolute but depends on the relative levels of specific IgG4 and total IgG4.
興味深いことに、好酸球性食道炎の患者では、牛乳タンパク質に特異的な IgG4 の非常に高い力価が観察されています (食道壁への IgG4 の沈着とともに)。この状況では IgG4 のかなりの部分が依然として二価である可能性があります; これは、IgG4 の有効な一価性が絶対的なものではなく、特定の IgG4 と総 IgG4 の相対的なレベルに依存することを示唆しています。
The process of Fab-arm exchange is controlled by redox conditions and can be promoted in vitro by choosing an appropriate redox buffer.
Fab アーム交換のプロセスは、酸化還元条件によって制御され、適切な酸化還元バッファーを選択することによって in vitro で促進できます。
Comparing IgG4 with IgG1, two mutations in the latter are required to enable Fab-arm exchange: a P228S mutation in the hinge region allowing for the disulfide bonds that normally connect the heavy chains to be easily broken, and a K409R mutation in the carboxy-terminal domains that results in weaker non-covalent interactions between the heavy chains.
IgG4 と IgG1 を比較すると、Fab アーム交換を有効にするには後者は 2 つの変異が必要です:ヒンジ領域の P228S 変異により、通常は重鎖を接続するジスルフィド結合が容易に切断されます。また、カルボキシ末端ドメインの K409R 変異により、重鎖間の非共有相互作用が弱くなります。
Conversely, therapeutic IgG4 monoclonal antibodies often contain a S228P mutation to prevent Fab-arm exchange in vivo.
逆に、治療用 IgG4 モノクローナル抗体には、in vivo での Fab アーム交換を防ぐために S228P 変異が含まれていることがよくあります。
A major functional consequence of the effective monovalency of the majority of IgG4 in vivo is that it further reduces the ability for signalling, antigen cross-linking and immune activation (Fig. 2).
in vivo での大部分の IgG4 が有効な一価であることの主な機能的結果は、シグナル伝達、抗原架橋、および免疫活性化の能力をさらに低下させることです (図 2)。

Furthermore, Fab-arm exchange seems to further decrease the limited potential of IgG4 for complement activation.
さらに、Fab アーム交換は、補体活性化に対する IgG4 の限られた可能性をさらに低下させるようです。
Therefore, Fab-arm exchange together with the overall reduced ability of IgG4 to activate Fcγ receptors and complement means that IgG4 is often regarded as a natural type of ‘blocking’ antibody — a high-affinity monovalent binder with limited potential to induce inflammatory responses.
したがって、Fab アームの交換と、IgG4 の Fcγ 受容体および補体を活性化する能力の全体的な低下は、IgG4 がしばしば天然型の「ブロッキング」抗体と見なされることを意味します。 — 炎症反応を誘発する可能性が限られている高親和性一価結合剤。
Because of these weak effector functions and blocking ability, IgG4 is the second most widely used antibody format for therapeutic monoclonal antibodies, with examples including natalizumab, nivolumab and reslizumab.
これらのエフェクター機能と遮断能力が弱いため、IgG4 は、ナタリズマブ、ニボルマブ、レスリズマブなどの治療用モノクローナル抗体で 2 番目に広く使用されている抗体フォーマットです。
Glycosylation
グリコシル化
The glycan structure on IgG molecules can vary.
IgG 分子のグリカン構造はさまざまです。
Specific glycan profiles have been associated with (patho)physiological conditions, and the exact glycan structure can affect antibody functions such as Fcγ receptor activation and complement activation.
特定のグリカン プロファイルは (病理) 生理学的状態に関連付けられており、正確なグリカン構造は、Fcγ 受容体活性化や補体活性化などの抗体機能に影響を与える可能性があります。
This has been investigated in most detail for the conserved N-linked glycans in the Fc region.
これは、Fc領域の保存されたN結合型グリカンについて最も詳細に調査されています。
In general, Fc galactosylation of IgG4 seems to be decreased in pathological conditions.
一般に、病的状態では IgG4 の Fc ガラクトシル化が減少しているようです。
Furthermore, N-linked glycans are also present in the variable regions of immunoglobulins to different degrees.
さらに、N-結合型グリカンは、免疫グロブリンの可変領域にもさまざまな程度で存在します。
Variable domain glycosylation is largely dependent on acquiring mutations that introduce glycosylation motifs during somatic hypermutation, such that specific antibody responses may be highly enriched or depleted for variable domain glycans.
可変ドメインのグリコシル化は、体細胞超変異中にグリコシル化モチーフを導入する変異の獲得に大きく依存しているため、可変ドメインのグリカンに対して特異的な抗体応答が高度に濃縮または消耗する可能性があります。
In particular, certain autoantibodies, including those against muscle-specific tyrosine kinase, desmoglein 3 (DSG3) and proteinase 3, are found to be highly glycosylated in the variable domain.
特に、筋肉特異的チロシンキナーゼ、デスモグレイン 3 (DSG3) およびプロテイナーゼ 3 に対するものを含む特定の自己抗体は、可変ドメインで高度にグリコシル化されていることがわかっています。
Interestingly, IgG4 antibodies in general have increased levels of Fab glycans compared with other IgG subclasses.
興味深いことに、IgG4 抗体は一般に、他の IgG サブクラスと比較して Fab グリカンのレベルが増加しています。
This feature seems to be associated with the type 2 response-like characteristics of the IgG4 response, as BCR repertoire analysis of both IgG4 and IgE responses showed increased levels of N-glycosylation motifs.
IgG4 応答と IgE 応答の両方の BCR レパートリー分析が N-グリコシル化モチーフのレベルの増加を示したため、この特徴は IgG4 応答のタイプ 2 応答様特性に関連しているようです。
The functional consequences of these glycans are not well understood but may include attenuation of antibody-mediated signalling by engaging lectins such as CD22, elimination of autoreactivity or enhancement of BCR signalling.
これらのグリカンの機能的影響はよく理解されていませんが、CD22 などのレクチンとの結合による抗体媒介シグナル伝達の減衰、自己反応性の排除、または BCR シグナル伝達の増強が含まれる可能性があります。
The link between antibody glycosylation and pathogenicity of IgG4 autoantibodies warrants further investigation.
抗体のグリコシル化と IgG4 自己抗体の病原性との関連性については、さらなる調査が必要です。
Physiological roles of IgG4
IgG4 の生理的役割
In general, the ‘blocking’ nature of an IgG4 response may be beneficial when it prevents excessive immune activation.
一般に、過剰な免疫活性化を防ぐ場合、IgG4 応答の「ブロッキング」の性質が有益である可能性があります。
In particular, in both allergic responses and parasitic infections, IgG4 responses are beneficial for the host by inducing tolerance and limiting inflammation (Fig. 3).
特に、アレルギー応答と寄生虫感染の両方において、IgG4応答は寛容を誘導し、炎症を制限することにより、宿主にとって有益です(図3)。

Allergic responses
アレルギー反応
Allergy is characterized by hypersensitivity reactions that lead to symptoms including rash, swelling, itching, upper respiratory tract sensitivity and, in severe cases, shock.
アレルギーは、発疹、腫れ、かゆみ、上気道過敏症、重篤な場合にはショックなどの症状を引き起こす過敏反応によって特徴付けられます。
These reactions result from allergens that trigger IL-13 and IL-4 release by TH2 cells and subsequent class-switching of B cells to IgE production.
これらの反応は、TH2 細胞による IL-13 および IL-4 の放出とそれに続く B 細胞の IgE 産生へのクラス切り替えを引き起こすアレルゲンに起因します。
In response to FcεRI stimulation by IgE bound to allergen, mast cells and basophils release histamine, cytokines and chemokines, which have effects on the vasculature and tissues to cause the hypersensitivity symptoms.
アレルゲンに結合したIgEによるFcεRI刺激に応答して、マスト細胞と好塩基球はヒスタミン、サイトカイン、ケモカインを放出します。 血管系や組織に影響を与え、過敏症の症状を引き起こします。
Peripheral tolerance to allergens can be achieved by specific immunotherapy or by regular exposure to them, which may induce allergen-specific IgG4 that can contribute to reducing hypersensitivity reactions by competing with IgE for binding to allergen and by other mechanisms (Fig. 3a).
アレルゲンに対する末梢耐性は、特定の免疫療法またはそれらへの定期的な曝露によって達成できます。これは、アレルゲンへの結合について IgE と競合することによって、および他のメカニズムによって、過敏症反応の軽減に寄与できるアレルゲン特異的 IgG4 を誘導する可能性があります (図 3a)。

Allergen-specific IgG4 responses have been described for a range of allergies, including to grass and birch pollen, cats, bee venom, peanuts and milk.
アレルゲン特異的 IgG4 応答は、草や白樺の花粉、猫、ハチ毒、ピーナッツ、牛乳など、さまざまなアレルギーについて報告されています。
In individuals who are allergic, IgG4 can constitute more than 75% of allergen-specific IgG after continuous exposure to antigen.
アレルギーのある人では、抗原に継続的にさらされた後、IgG4 がアレルゲン特異的 IgG の 75% 以上を構成する可能性があります。
After specific immunotherapy, IgG1 and, in particular, IgG4 allergen-specific responses can increase in the range 10- to 100-fold, with levels starting to increase after 1 month of therapy.
特異的免疫療法の後、IgG1、特に IgG4 アレルゲン特異的応答は、10 倍から 100 倍の範囲で増加する可能性があり、治療の 1 か月後にレベルが増加し始めます。
An increase in the titre of allergen-specific IgG4 generally correlates with increased tolerance and reduced hypersensitivity symptoms.
アレルゲン特異的 IgG4 の力価の増加は、一般に耐性の増加および過敏症の症状の軽減と相関します。
The protection against symptoms of allergy mediated by IgG4 is thought to be the result of at least three modes of action: blocking the activity of IgE by competing for allergen binding and preventing mast cell and basophil degranulation; inhibiting antigen presentation to T cells by IgE on B cells and dendritic cells; and preventing immune complex formation through the functional monovalency of IgG4.
IgG4 によって媒介されるアレルギーの症状に対する保護は、少なくとも 3 つの作用機序の結果であると考えられています:アレルゲン結合をめぐって競合し、マスト細胞と好塩基球の脱顆粒を防ぐことにより、IgEの活性をブロック;B細胞および樹状細胞上のIgEによるT細胞への抗原提示の阻害;IgG4の機能的な一価性による免疫複合体形成の防止。
The induction of allergen-specific IgG4 responses is thought to result from prolonged exposure to the allergen and increased production of IL-10.
アレルゲン特異的な IgG4 応答の誘導は、アレルゲンへの長時間の曝露と IL-10 の産生増加に起因すると考えられています。
IL-10 not only induces T cell tolerance but also regulates antibody production, resulting in increased IgG production relative to IgE production.
IL-10 は T 細胞の寛容を誘導するだけでなく、抗体産生も調節し、IgE 産生に比べて IgG 産生を増加させます。
IgE-induced CD4+ T cell activation is a very potent route to maintain chronic inflammation.
IgE による CD4+ T 細胞の活性化は、慢性炎症を維持するための非常に強力な経路です。
Antigen presentation to T cells via B cells and dendritic cells may be facilitated by IgE.
B細胞および樹状細胞を介したT細胞への抗原提示は、IgEによって促進される可能性があります。
By blocking these effects, IgG4 halts this positive-feedback loop, which limits IgE production and puts a brake on the inflammatory response.
これらの効果をブロックすることにより、IgG4 はこの正のフィードバック ループを停止させ、IgE 産生を制限し、炎症反応にブレーキをかけます。
Parasitic infections
寄生虫感染症
IgG4 responses can also occur during parasitic infection (Fig. 3b).
IgG4応答は、寄生虫感染中にも発生する可能性があります(図3b)。

The host usually develops a broad B cell-mediated and T cell-mediated immune response against the parasite.
宿主は通常、寄生虫に対する幅広いB細胞媒介性およびT細胞媒介性免疫応答を発症します。
In an attempt to evade the host immune response, the parasite stimulates production of cytokines such as IL-10 and induction of Treg cells.
宿主の免疫応答を回避しようとして、寄生虫は IL-10 などのサイトカインの産生と Treg 細胞の誘導を刺激します。
As a consequence, in a subset of patients, the anti-parasite B cell response may undergo class-switching towards IgG4.
結果として、患者のサブセットでは、抗寄生虫 B 細胞応答が IgG4 へのクラス切り替えを受ける可能性があります。
IgG responses can consist of up to 90% IgG4 in these asymptomatic patients.
これらの無症候性患者では、IgG 応答は最大 90% の IgG4 で構成されます。
A dual role for IgG4 in these infections can be envisaged.
これらの感染症における IgG4 の二重の役割が考えられます。
On the one hand, the ratio of IgG4 to IgE may positively correlate with asymptomatic infection, potentially by preventing ongoing inflammation and damage to host tissues, for example in the case of Brugia malayi infection.
一方で、IgG4 と IgE の比率は、進行中の炎症や宿主組織への損傷を防ぐことにより、無症候性感染と正の相関がある可能性があります。たとえば、マラヤ ブルジア感染症の場合です。
On the other hand, the ratio of specific IgG4 to IgE may correlate with the intensity of infection and might represent escape of the parasite from host immunity.
一方、特定の IgG4 と IgE の比率は、感染の強度と相関している可能性があり、宿主免疫からの寄生虫の逃避を表している可能性があります。
Indeed, histamine release by IgE-opsonized basophils from patients with filariasis challenged with filarial antigen was blocked by patient-derived IgG4, and histamine release inversely correlated with IgG4 levels.
実際、フィラリア抗原で攻撃されたフィラリア症患者の IgE オプソニン化好塩基球によるヒスタミン放出は、患者由来の IgG4 によってブロックされ、ヒスタミン放出は IgG4 レベルと逆相関していました。
Interestingly, it has been suggested that a consequence of the IgG4-mediated attenuation of host immunity by the parasite may be protection of the host from autoimmune disease and allergies (see next section), although the role of parasitic infections in dampening allergy has not been unambiguously determined.
興味深いことに、寄生虫による宿主免疫の IgG4 媒介性減弱の結果は、自己免疫疾患およびアレルギーから宿主を保護する可能性があることが示唆されています (次のセクションを参照)。ただし、アレルギーの抑制における寄生虫感染の役割は明確に決定されていません。
Such a role would be consistent with the ‘hygiene hypothesis’, which suggests that lack of parasite exposure and the improved standards of hygiene in higher-income countries may cause increased prevalence of autoimmune and allergic diseases.
このような役割は、寄生虫への曝露の欠如と高所得国における衛生基準の改善により、自己免疫疾患やアレルギー疾患の有病率が増加する可能性があることを示唆する「衛生仮説」と一致します。
IgG4-dependent pathology
IgG4 依存性病理
Specific deficiency of IgG4 can occur either as an isolated phenomenon (in ~30% of cases) or in combination with a deficiency of other antibody (sub)classes, such as IgG2, IgA or IgG1.
IgG4 の特定の欠乏は、孤立した現象 (症例の約 30%) として、または IgG2、IgA または IgG1 などの他の抗体 (サブ) クラスの欠乏と組み合わせて発生する可能性があります。
A selective lack of IgG4 or severely reduced levels of IgG4 are extremely rare.
IgG4 の選択的欠如または IgG4 レベルの大幅な低下は極めてまれです。
In some individuals (mostly children), IgG4 deficiency is associated with recurrent respiratory tract infections, allergies, candidiasis, chronic diarrhoea and chronic fungal infections.
一部の個人 (主に子供) では、IgG4 欠乏症は再発性気道感染症、アレルギー、カンジダ症、慢性下痢、および慢性真菌感染症と関連しています。
These observations suggest that IgG4 may have an unexplored physiological role in mucosal immunity.
これらの観察結果は、IgG4 が粘膜免疫において未知の生理学的役割を果たしている可能性があることを示唆しています。
In line with this, a recent retrospective study observed IgG4 deficiency in ~20% of patients with inflammatory bowel disease, which was associated with worse disease outcome.
これに沿って、最近のレトロスペクティブ研究では、炎症性腸疾患患者の約 20% で IgG4 欠乏症が観察され、これは疾患転帰の悪化と関連していました。
Future studies should elucidate whether there is a causal relationship between these observations.
今後の研究では、これらの観察結果の間に因果関係があるかどうかを解明する必要があります。
IgG4 hypergammaglobulinaemia occurs in ~5% of the healthy population and seems without consequence.
IgG4 高ガンマグロブリン血症は、健康な人口の約 5% で発生し、結果はないようです。
Several diseases are associated with pathogenic IgG4 responses (Fig. 4).
いくつかの疾患は、病原性 IgG4 応答に関連しています (図 4)。

Here, we discuss examples of pathology that are directly dependent on IgG4, including autoimmune diseases, antitumour responses and anti-biologic responses.
ここでは、自己免疫疾患、抗腫瘍反応、抗生物質反応など、IgG4 に直接依存する病理学の例について説明します。
IgG4 autoimmune diseases
IgG4自己免疫疾患
IgG4-AIDs were first defined as a separate subgroup of antibody-mediated autoimmune disorders in 2015, and a first attempt at their classification based on the level of evidence for a pathogenic role of IgG4 was proposed soon thereafter.
IgG4-AID は、2015 年に抗体介在性自己免疫疾患の別のサブグループとして最初に定義され、その後まもなく、IgG4 の病原性役割の証拠レベルに基づく分類の最初の試みが提案されました。
IgG4-AIDs are characterized by autoantibody responses predominantly of the IgG4 subclass against a known antigen.
IgG4-AID は、主に既知の抗原に対する IgG4 サブクラスの自己抗体応答によって特徴付けられます。
These disorders can affect many organ systems, depending on the major site of action of the targeted antigen, including the kidneys, central and peripheral nervous systems, haematopoietic system and skin.
これらの障害は、標的抗原の主要な作用部位に応じて、腎臓、中枢および末梢神経系、造血系、皮膚など、多くの臓器系に影響を与える可能性があります。
So far, a direct pathogenic role of IgG4 autoantibodies has been established for six IgG4-AIDs through passive transfer of IgG4 in experimental animals (Table 2), but this group is likely to expand in the coming years with more evidence becoming available for 23 other candidate IgG4-AIDs.
これまでのところ、IgG4 自己抗体の直接的な病原性役割は、実験動物における IgG4 の受動伝達を通じて、6 つの IgG4-AID について確立されています (表 2)。 しかし、このグループは今後数年間で拡大する可能性が高く、他の 23 の候補 IgG4-AID についてより多くの証拠が利用可能になります。
Diagnosis of an IgG4-AID is based on clinical symptoms and the detection of serum IgG4 autoantibodies to the disease-specific antigen.
IgG4-AID の診断は、臨床症状と、疾患特異的抗原に対する血清 IgG4 自己抗体の検出に基づいて行われます。
Antigen-specific IgG4 levels correlate closely with disease severity.
抗原特異的 IgG4 レベルは、疾患の重症度と密接に相関しています。
As IgG4 is predominantly anti-inflammatory in nature and is not thought to induce pathology through Fc-dependent effector mechanisms, a key mode of action in all IgG4-AIDs is thought to be blocking essential protein–protein interactions of the target antigen.
IgG4 は本質的に主に抗炎症性であり、Fc 依存性エフェクターメカニズムを介して病状を誘発するとは考えられていないため、すべての IgG4-AID における重要な作用機序は、標的抗原の必須タンパク質間相互作用を遮断することであると考えられています。
For example, in muscle-specific kinase (MuSK) myasthenia gravis, which is a prototypical IgG4-AID, IgG4 autoantibodies to MuSK block its interaction with low-density lipoprotein receptor-related protein 4 (LRP4), thereby obstructing a key trophic signalling cascade at the neuromuscular junction and resulting in fatigable skeletal muscle weakness.
例えば、筋肉特異的キナーゼ (MuSK) 重症筋無力症は原型的な IgG4-AID であり、MuSK に対する IgG4 自己抗体が低密度リポタンパク質受容体関連タンパク質 4 (LRP4) との相互作用をブロックします。それにより、神経筋接合部での重要な栄養シグナル伝達カスケードを妨害し、疲労性の骨格筋の衰弱をもたらします。
Much overlap between the IgG4-AIDs can be found in terms of their inflammatory status and treatment response.
IgG4-AID 間の多くの重複は、炎症状態と治療反応の点で見られます。
For example, B cell depletion therapy with rituximab often results in long-term remission in all of these diseases.
たとえば、リツキシマブによる B 細胞除去療法は、多くの場合、これらすべての疾患で長期寛解をもたらします。
These observations suggest that although IgG4-AIDs can present with various symptoms depending on the target antigen, they share an underlying immunophenotype.
これらの観察結果は、IgG4-AID は標的抗原に応じてさまざまな症状を示す可能性がありますが、根本的な免疫表現型を共有していることを示唆しています。
Serum levels of IgG4 are, if at all, only slightly increased in patients with IgG4-AIDs and the numbers of IgG4+ plasma cells and IgG4+ B cells are normal in circulation (M.G.H., unpublished observations).
IgG4 の血清レベルは、たとえあったとしても、IgG4-AID の患者ではわずかにしか増加せず、循環中の IgG4+ 形質細胞と IgG4+ B 細胞の数は正常です (M.G.H.、未発表の観察結果)。
These observations do not support a hypothesis that these patients have an overall tendency to develop dominant IgG4 responses but, rather, support an antigen-driven aetiology.
これらの観察結果は、これらの患者が優勢な IgG4 応答を発現する全体的な傾向があるという仮説を支持するものではなく、むしろ抗原主導の病因を支持するものです。
A recent systematic review reports strong associations of IgG4-AIDs with HLA-DQB105 and HLA-DRB114, suggesting that these haplotypes predispose to the development of IgG4-AIDs.
最近のシステマティック レビューでは、IgG4-AID と HLA-DQB105 および HLA-DRB114 との強い関連性が報告されており、これらのハプロタイプが IgG4-AID の発症の素因となることを示唆しています。
This may occur through directing B cell development and cytokine production, or by facilitating antigen presentation.
これは、B 細胞の発生とサイトカインの産生を指示することによって、または抗原提示を促進することによって発生する可能性があります。
In the case of three archetypal IgG4-AIDs — MuSK myasthenia gravis, pemphigus vulgaris and thrombotic thrombocytopenic purpura — increased serum levels of the IgG4-promoting cytokine IL-10 have been reported.
3 つの典型的な IgG4-AID である MuSK 重症筋無力症、尋常性天疱瘡、および血栓性血小板減少性紫斑病の場合、IgG4 を促進するサイトカイン IL-10 の血清レベルの上昇が報告されています。
Although some autoimmune diseases mediated by IgG1, IgG2 or IgG3 are associated with tumour development, this has thus far not been reported for IgG4-AIDs, and the aetiology of IgG4-AIDs is expected to be different from that of such paraneoplastic syndromes.
IgG1、IgG2、または IgG3 によって媒介されるいくつかの自己免疫疾患は腫瘍の発生と関連していますが、これはこれまでのところ IgG4-AID について報告されておらず、IgG4-AID の病因はそのような腫瘍随伴症候群の病因とは異なると予想されます。
Interestingly, evidence for molecular mimicry resulting in an IgG4-dominated response to desmogleins in the skin epidermis is found in patients with endemic pemphigus foliaceus from Brazilian and Tunisian populations (Fig. 4a).
興味深いことに、皮膚表皮のデスモグレインに対する IgG4 優位の応答をもたらす分子模倣の証拠は、ブラジルおよびチュニジアの集団からの風土性葉状天疱瘡患者に見られます (図 4a)。

In these patients, antibodies develop against a salivary antigen from flies that are cross-reactive with desmogleins.
これらの患者では、デスモグレインと交差反応するハエの唾液抗原に対する抗体が発生します。
Moreover, monoclonal antibodies derived from patients with pemphigus vulgaris were shown to be cross-reactive with walnut antigen.
さらに、尋常性天疱瘡患者由来のモノクローナル抗体は、クルミ抗原と交差反応することが示された。
Both walnuts and flies carry allergens that are known to induce IgG4 responses.
クルミもハエも、IgG4 応答を誘発することが知られているアレルゲンを持っています。
These observations suggest that exposure to certain IgG4-inducing antigens in combination with a permissive HLA haplotype and an IgG4-promoting immune environment (such as increased IL-10 levels) might have a role in the development of certain IgG4-AIDs.
これらの観察結果は、寛容な HLA ハプロタイプおよび IgG4 を促進する免疫環境 (IL-10 レベルの増加など) と組み合わせた特定の IgG4 誘導抗原への曝露が、特定の IgG4-AID の発症に関与している可能性があることを示唆しています。
However, it is unclear which factors ultimately cause the IgG4 skewing of these responses.
しかし、どの要因が最終的にこれらの応答の IgG4 歪曲を引き起こすかは不明です。
Autoantibodies of other (sub)classes can also be found in patients with IgG4-AIDs, although usually of much lower titres.
他の (サブ) クラスの自己抗体も IgG4-AID の患者に見られますが、通常ははるかに低い力価です。
For several IgG4-AIDs, the pathogenicity of IgG1, IgG3 and IgM autoantibodies has also been confirmed.
いくつかの IgG4-AID については、IgG1、IgG3、および IgM 自己抗体の病原性も確認されています。
The mechanisms by which these autoantibodies induce pathology may differ between antibody (sub)classes and target antigens, and could include complement-mediated tissue damage and antigenic cross-linking and internalization causing surface depletion.
これらの自己抗体が病理を誘発するメカニズムは、抗体 (サブ) クラスと標的抗原の間で異なる可能性があり、補体媒介組織損傷、抗原架橋、および表面枯渇を引き起こす内在化が含まれる可能性があります。
The functional consequences of IgG4 autoantibodies may also differ between diseases.
IgG4自己抗体の機能的結果も、疾患によって異なる場合があります。
The relative contribution of IgG4 to pathology compared with the contribution of other antibody (sub)classes has not been carefully delineated, but the effects of different antibody (sub)classes may function in parallel to increase disease severity.
他の抗体 (サブ) クラスの寄与と比較した病理学への IgG4 の相対的な寄与は、慎重に描写されていませんが、異なる抗体 (サブ) クラスの影響が並行して機能し、疾患の重症度を増加させる可能性があります。
Low titres of IgG1 autoantibodies are also found in non-symptomatic relatives of patients with pemphigus vulgaris, which suggests that having (low levels of) such antibodies alone is not sufficient to precipitate disease symptoms.
低力価の IgG1 自己抗体は、尋常性天疱瘡患者の無症候性近親者にも見られます。これは、このような抗体 (低レベル) だけを持っているだけでは疾患の症状を引き起こすのに十分ではないことを示唆しています。
It furthermore suggests that there is a subclinical stage in these autoimmune diseases that, upon early detection, could allow for IgG4-AID onset to be prevented.
さらに、これらの自己免疫疾患には無症状の段階があり、早期発見により IgG4-AID の発症を防ぐことができる可能性があることを示唆しています。
The role of autoantibodies of other (sub)classes in the pathophysiology of IgG4-AIDs requires further investigation.
IgG4-AID の病態生理における他の (サブ) クラスの自己抗体の役割については、さらなる調査が必要です。
Interestingly, in autoimmune diseases mediated by IgG1, IgG2 or IgG3 autoantibodies, a switch to an IgG4-dominant response may be therapeutic.
興味深いことに、IgG1、IgG2、または IgG3 自己抗体によって媒介される自己免疫疾患では、IgG4 優性応答への切り替えが治療に役立つ可能性があります。
Passive transfer of an IgG4 monoclonal antibody targeting acetylcholine receptor (AChR) inhibited subsequent complement-mediated damage and cytotoxicity induced by IgG1 binding to AChR, thus preventing the onset of AChR myasthenia gravis in rhesus macaques.
アセチルコリン受容体 (AChR) を標的とする IgG4 モノクローナル抗体の受動伝達は、その後の補体媒介性損傷および IgG1 が AChR に結合することによって誘発される細胞毒性を抑制し、アカゲザルにおける AChR 重症筋無力症の発症を予防しました。
Antitumour responses
抗腫瘍反応
Antitumour antibody responses can contain or even eliminate malignancies by binding to tumour cells and stimulating ADCC, antibody-dependent complement deposition and/or ADCP.
抗腫瘍抗体応答は、腫瘍細胞に結合し、ADCC、抗体依存性補体沈着および/またはADCPを刺激することにより、悪性腫瘍を抑制または排除することさえできます。
However, in 1977 a prospective study in patients with melanoma identified high levels of IgG4 as having negative effects on survival.
しかし、1977 年にメラノーマ患者を対象とした前向き研究により、高レベルの IgG4 が生存に悪影響を及ぼすことが確認されました。
It has since become evident that some malignancies evade host immune defences by inducing class-switching of the antitumour antibody response to IgG4.
それ以来、一部の悪性腫瘍は、IgG4 に対する抗腫瘍抗体応答のクラス切り替えを誘導することにより、宿主の免疫防御を回避することが明らかになりました。
IgG4 competes with other antibody (sub)classes for binding to tumour antigens and owing to its anti-inflammatory properties blocks the induction of antitumour immune responses (Fig. 4b).
IgG4は、腫瘍抗原への結合について他の抗体(サブ)クラスと競合し、その抗炎症特性により、抗腫瘍免疫応答の誘導をブロックします(図4b)。

In the absence of an immune response, tumour cells have increased ability to proliferate and metastasize, resulting in disease progression and decreased survival.
免疫応答がない場合、腫瘍細胞は増殖および転移する能力が高まり、その結果、疾患が進行し、生存率が低下します。
Immune evasion through class-switching to IgG4 has been observed in patients with melanoma, cholangiocarcinoma, colon cancer, pancreatic cancer and glioblastoma.
IgG4 へのクラス切り替えによる免疫回避は、メラノーマ、胆管がん、結腸がん、膵臓がん、膠芽腫の患者で観察されています。
Both the total IgG4 level and the number of IgG4+ B cells can be increased in the serum of patients with malignancies and are a negative prognostic indicaton.
総 IgG4 レベルと IgG4+ B 細胞数の両方が悪性腫瘍患者の血清で増加する可能性があり、これは予後不良の指標です。
These factors are also increased in the tumour microenvironment.
これらの要因は、腫瘍の微小環境でも増加します。
The development of antitumour IgG4 responses results from IL-4 and IL-10 production by tumour cells, which directs a modified type 2 response that stimulates class-switching to IgG4 (Fig. 4b).
抗腫瘍IgG4応答の発生は、腫瘍細胞によるIL-4およびIL-10産生に起因し、IgG4へのクラス切り替えを刺激する改変された2型応答を指示します(図4b)。
Furthermore, some tumour microenvironments contain tertiary lymphoid structures with functional germinal centres and Treg cells.
さらに、一部の腫瘍微小環境には、機能的な胚中心と Treg 細胞を含む三次リンパ構造が含まれています。
Crosstalk between these chronic inflammatory structures and the tumour may induce increased expression of IL-10 by Treg cells.
これらの慢性炎症構造と腫瘍の間のクロストークは、Treg 細胞による IL-10 の発現増加を誘発する可能性があります。
Importantly, it is not yet understood why certain tumours are capable of inducing IgG4 responses whereas others are not.
重要なことに、他の腫瘍はIgG4誘導をしていないのに、特定の腫瘍は IgG4 応答を誘導できる理由は未だ不明です。
In addition, although the reactivity of serum IgG4 antibodies from patients with cancer to tumour cells, for example to melanoma cells, has been confirmed, the precise antigen specificity of these antibodies has not yet been delineated.
また、がん患者の血清IgG4抗体がメラノーマ細胞などの腫瘍細胞に反応することは確認されていますが、これらの抗体の正確な抗原特異性はまだ明らかにされていません。
Broader study is needed to evaluate whether IgG4 has a pathogenic role in other cancer types.
IgG4 が他の種類のがんにおいて病原性を持っているかどうかを評価するには、より広範な研究が必要です。
Interestingly, in addition to the proposed role of IgG4 in blocking inflammatory responses, a pro-angiogenic IgG4+ B cell subset (CD49b+CD73+IL-10–) was recently identified.
興味深いことに、炎症反応のブロックにおける IgG4 の提案された役割に加えて、血管新生促進性 IgG4+ B 細胞サブセット (CD49b+CD73+IL-10–) が最近同定されました。
These B cells were increased in the serum of patients with melanoma and might facilitate tumour angiogenesis.
これらの B 細胞は黒色腫患者の血清で増加しており、腫瘍の血管新生を促進する可能性があります。
Anti-biologic responses
抗生物質への反応
IgG4-skewed responses can also occur as a result of chronic exposure to biological therapies.
IgG4 歪んだ応答は、生物学的療法への慢性的な暴露の結果として発生することもあります。
Such responses have been described for clotting factors FVIII and FIX used for the treatment of congenital haemophilia A or haemophilia B, for interferon-β used for the treatment of multiple sclerosis and for the tumour necrosis factor (TNF) inhibitors adalimumab and infliximab used for the treatment of inflammatory disorders such as rheumatoid arthritis and Crohn’s disease.
このような応答は、先天性血友病Aまたは血友病Bの治療に使用される凝固因子FVIIIおよびFIX、多発性硬化症の治療に使用されるインターフェロン-β、および腫瘍壊死因子(TNF)阻害剤アダリムマブ、関節リウマチやクローン病などの炎症性疾患の治療に使用されるインフリキシマブについて記載されています。
For each of these anti-biologic responses, the primary result is that the therapeutic effect of the biologic is impaired (Fig. 4c).
これらの各抗生物質応答の主な結果は、生物学的製剤の治療効果が損なわれることです (図 4c)。

Not all biologics trigger an IgG4-skewed anti-drug response.
すべての生物製剤が IgG4 に偏った抗薬物反応を引き起こすわけではありません。
For example, interferon-β induces prominent IgG4 skewing only in some of the patients who develop an antibody response, which argues against chronic exposure to the biologic being the sole determinant of IgG4 skewing.
たとえば、インターフェロン-β は、抗体反応を発症する一部の患者でのみ顕著な IgG4 の偏りを誘発します。これは、生物製剤への慢性的な曝露が IgG4 のゆがみの唯一の決定要因であることに反対するものです。
It is unclear why certain biologics cause these responses and others do not.
特定の生物製剤がこれらの反応を引き起こし、他の生物製剤がそうでない理由は不明です。
Haemophilia A and haemophilia B are severe clotting disorders caused by an inherited deficiency of FVIII or FIX, respectively.
血友病 A および血友病 B は、それぞれ FVIII または FIX の遺伝的欠乏によって引き起こされる重度の凝固障害です。
First-line treatment in these patients is chronic replacement of these clotting factors using either plasma-derived or recombinant proteins.
これらの患者の第一選択治療は、血漿由来タンパク質または組換えタンパク質のいずれかを使用して、これらの凝固因子を慢性的に置換することです。
Approximately 30% of patients treated with FVIII replacement therapy develop inhibitory antibodies predominantly of the IgG4 subclass.
FVIII 補充療法を受けた患者の約 30% は、主に IgG4 サブクラスの阻害抗体を発症します。
Low-affinity IgG antibodies to FVIII of all subclasses can be found in both healthy individuals and patients with haemophilia, but high-affinity, high-titre IgG4 blocking antibodies to FVIII are unique to patients.
すべてのサブクラスの FVIII に対する低親和性 IgG 抗体は、健康な個人と血友病患者の両方に見られますが、FVIII に対する高親和性で力価の高い IgG4 遮断抗体は患者に固有のものです。
Furthermore, high levels of anti-FVIII IgG4 correlate with decreased efficacy of FVIII replacement therapy.
さらに、高レベルの抗FVIII IgG4は、FVIII補充療法の有効性の低下と相関する。
Although these patients may also have antibodies to FVIII of other subclasses, IgG4 antibodies seem to be particularly detrimental.
これらの患者は他のサブクラスの FVIII に対する抗体も持っている可能性がありますが、IgG4 抗体は特に有害であるようです。
Interestingly, acquired autoimmune haemophilia A is also associated with a dominant IgG4 response to endogenous FVIII, which would thus classify this form of haemophilia as an IgG4-AID.
興味深いことに、後天性自己免疫性血友病 A は、内因性 FVIII に対する主要な IgG4 応答とも関連しているため、この型の血友病は IgG4-AID として分類されます。
For unknown reasons, FVIII — both endogenous and exogenous — has the propensity to induce an IgG4 response.
理由は不明ですが、FVIII (内因性と外因性の両方) は IgG4 応答を誘発する傾向があります。
Many inflammatory disorders, including rheumatoid arthritis and Crohn’s disease, can be successfully managed with TNF inhibitor therapy.
関節リウマチやクローン病を含む多くの炎症性疾患は、TNF阻害剤療法でうまく管理できます。
Monoclonal antibodies are immunogenic to varying degrees, depending, amongst other factors, on their extent of humanization.
モノクローナル抗体は、他の要因の中でも特にヒト化の程度に応じて、さまざまな程度の免疫原性を示します。
However, even so-called fully human antibodies, of which the TNF inhibitor adalimumab is an early example, contain parts that are unique and foreign to recipients, namely the complementarity-determining regions responsible for target binding.
しかし、TNF阻害剤アダリムマブがその初期の例である、いわゆる完全ヒト抗体でさえ、レシピエントに固有で異質な部分、すなわち標的結合に関与する相補性決定領域を含んでいます。
B cell and T cell epitopes will be present in the biologic that can drive the development of high-affinity, class-switched (IgG4) antibodies.
B 細胞および T 細胞のエピトープは、高親和性のクラス スイッチ (IgG4) 抗体の開発を促進できる生物学的製剤に存在します。
Similarly, although FVIII is a human protein, it will be seen by the immune system as a partially foreign protein in patients with congenital haemophilia owing to genetic defects in endogenous FVIII.
同様に、FVIII はヒトタンパク質ですが、内因性 FVIII の遺伝的欠陥により、先天性血友病患者の免疫系によって部分的に外来タンパク質として認識されます。
This explains the potential for developing high-affinity, class-switched antibodies, although it is unclear why there is a tendency for these responses to favour class-switching to IgG4.
これは、高親和性のクラス スイッチ抗体を開発する可能性を説明していますが、なぜこれらの応答が IgG4 へのクラス スイッチを支持する傾向があるのかは不明です。
IgG4-related diseases
IgG4関連疾患
IgG4-RDs are a heterogeneous group of inflammatory disorders characterized by massive influx of IgG4+ B cells in affected organs and increased serum levels of IgG4.
IgG4-RD は、影響を受けた臓器への IgG4+ B 細胞の大量の流入と IgG4 の血清レベルの上昇を特徴とする、不均一な炎症性疾患のグループです。
Similar to IgG4-AIDs, a wide variety of organs can be affected in IgG4-RDs, including the thyroid, pituitary gland, pancreas, lungs, kidneys, gastrointestinal system and vasculature, with symptoms varying according to the organ affected.
IgG4-AID と同様に、甲状腺、脳下垂体、膵臓、肺、腎臓、胃腸系、血管系など、さまざまな臓器が IgG4-RD で影響を受ける可能性があり、症状は影響を受ける臓器によって異なります。
Diagnosis of an IgG4-RD is based on the histopathological finding of an IgG4+ B cell infiltrate, resulting in swelling of the organ, storiform fibrosis and obliterative phlebitis in a tissue biopsy.
IgG4-RD の診断は、IgG4+ B 細胞浸潤の組織病理学的所見に基づいており、組織生検で臓器の腫脹、ストリア状線維症、閉塞性静脈炎を引き起こします。
In addition, increased serum levels of IgG4 and of IgG4+ plasmablasts are often a good biomarker for both diagnosis and monitoring disease progression; 70–80% of patients have increased serum levels of IgG4.
さらに、IgG4 および IgG4+ 形質芽細胞の血清レベルの上昇は、多くの場合、診断と疾患進行のモニタリングの両方に適したバイオマーカーです;患者の 70 ~ 80% で IgG4 の血清レベルが上昇しています。
IgG4-AIDs and IgG4-RDs are currently considered to be separate disease entities as there is no evidence for large-scale influx of IgG4+ B cells in the affected organs in IgG4-AIDs or for significantly increased serum levels of IgG4.
IgG4-AID と IgG4-RD は、現在、IgG4-AID の影響を受ける臓器に IgG4+ B 細胞が大量に流入したり、IgG4 の血清レベルが大幅に上昇したりする証拠がないため、別個の疾患であると考えられています。
The typical histology observed in IgG4-RDs involving fibrosis and tissue damage does not seem to have a major role in IgG4-AIDs, although biopsy data in the latter are limited.
後者の生検データは限られていますが、線維症および組織損傷を伴う IgG4-RD で観察される典型的な組織学は、IgG4-AID で大きな役割を果たしているようには見えません。
Furthermore, the pathogenic role of specific IgG4 and IgG4+ B cells in IgG4-RDs remains enigmatic.
さらに、IgG4-RD における特定の IgG4 および IgG4+ B 細胞の病原性の役割は謎のままです。
BCR repertoire sequencing confirmed the clonal expansion of IgG4+ B cells in patients with IgG4-RDs, suggesting that some clones may contribute specifically to disease onset and progression, although their antigen specificity was not determined.
BCR レパートリー配列決定により、IgG4-RD 患者における IgG4+ B 細胞のクローン増殖が確認され、いくつかのクローンが疾患の発症と進行に特異的に寄与している可能性があることが示唆されましたが、それらの抗原特異性は決定されていませんでした。
Further research is needed to determine the similarities and differences between IgG4-RDs and IgG4-AIDs in terms of their pathology, aetiology, histology and clinical features.
IgG4-RD と IgG4-AID の病理学、病因学、組織学、および臨床的特徴の点での類似点と相違点を判断するには、さらなる研究が必要です。
There are three hypotheses to explain the role of IgG4 in the pathophysiology of IgG4-RDs.
IgG4-RD の病態生理における IgG4 の役割を説明するには、3 つの仮説があります。
First, IgG4-RDs, similar to IgG4-AIDs, are caused by IgG4 antibodies targeting autoantigens in the specific organ that is affected.
まず、IgG4-AID と同様に IgG4-RD は、影響を受ける特定の臓器の自己抗原を標的とする IgG4 抗体によって引き起こされます。
Second, patients with IgG4-RDs have a type 2-skewed inflammatory environment, for as yet unknown reasons, which triggers pleiotropic IgG4 responses and impaired homing of IgG4+ B cells.
第二に、IgG4-RD を有する患者は、まだ未知の理由で、タイプ 2 に偏った炎症環境を持っています。これは、多面的な IgG4 応答と IgG4+ B 細胞のホーミング障害を引き起こします。
Third, IgG4 is present in IgG4-RDs simply to dampen an ongoing immune response and, as such, does not contribute to the pathology.
第三に、IgG4 は単に進行中の免疫応答を弱めるために IgG4-RD に存在するため、病状には寄与しません。
Clinical clues to support an autoimmune hypothesis in these patients are the responsiveness to immunosuppressants, chronic disease course, presence of autoantibodies and HLA type II associations.
これらの患者における自己免疫仮説を支持する臨床的手がかりは、免疫抑制剤に対する反応性、慢性疾患の経過、自己抗体の存在、および HLA II 型関連性です。
Passive transfer of IgG1 and IgG4 from patients with IgG4-RDs can induce similar pathology in experimental animals.
IgG4-RD を有する患者からの IgG1 および IgG4 の受動的移入は、実験動物で同様の病理を誘発する可能性があります。
Although IgG antibodies to autoantigens (for example, nuclear antigens, lactoferrin, carbonic anhydrases II and IV, pancreatic secretory inhibitor, trypsinogens and annexin A11) have been found, none of these consistently correlates with an IgG4-RD.
自己抗原に対する IgG 抗体 (例えば、核抗原、ラクトフェリン、炭酸脱水酵素 II および IV、膵臓分泌阻害剤、トリプシノーゲン、アネキシン A11) が見つかっていますが、これらのいずれも一貫して IgG4-RD と相関していません。
Furthermore, these autoantibodies are mainly of the IgG1 subclass and target intracellular proteins that are unlikely to be the initial autoimmune trigger owing to lack of accessibility.
さらに、これらの自己抗体は主に IgG1 サブクラスのものであり、接近しにくいために最初の自己免疫の引き金になる可能性が低い細胞内タンパク質を標的としています。
By contrast, and in keeping with the second hypothesis, patients with IgG4-RDs generally have increased IgG4 reactivity against environmental antigens, suggesting that increased levels of IgG4 may be the result of a pleiotropic activation of IgG4+ B cells independent of their antigen specificity.
対照的に、2 番目の仮説に沿って、IgG4-RD の患者は一般に環境抗原に対する IgG4 反応性が増加しており、IgG4 レベルの増加は、抗原特異性とは無関係に IgG4+ B 細胞の多面的な活性化の結果である可能性があることを示唆しています。
Type 2 cytokines are increased in the serum of patients with IgG4-RDs, which could fit with both the first and second hypotheses.
タイプ 2 サイトカインは、IgG4-RD を持つ患者の血清で増加しており、これは最初の仮説と 2 番目の仮説の両方に適合する可能性があります。
Given the diverse nature of immune cells infiltrating affected organs in patients with IgG4-RDs, some argue that the IgG4 is induced in response to chronic immune stimulation and does not contribute to the pathology.
IgG4-RD患者の影響を受けた臓器に浸潤する免疫細胞の多様な性質を考えると、IgG4 は慢性的な免疫刺激に応答して誘導され、病状には寄与しないと主張する人もいます。
Indeed, passive transfer of IgG1 purified from a patient with pancreatitis reproduced disease in mice, whereas symptoms were markedly reduced upon co-transfer of IgG1 and IgG4 isolated from the same patient.
実際、膵炎患者から精製した IgG1 を受動的に移入すると、マウスで疾患が再現されましたが、同じ患者から分離された IgG1 と IgG4 を同時に移入すると、症状が著しく軽減されました。
Furthermore, in addition to IgG4, levels of IgE are often increased in IgG4-RDs.
さらに、IgG4 に加えて、IgG4-RD では IgE のレベルが上昇することがよくあります。
Upon treatment with rituximab, both IgG4 and IgE levels have a tendency to decrease.
リツキシマブで治療すると、IgG4 と IgE の両方のレベルが低下する傾向があります。
Interestingly, this was also observed upon treatment with abatacept, which interferes with T cell activation, albeit in a limited number of patients.
興味深いことに、これは、限られた数の患者ではあるものの、T細胞の活性化を妨げるアバタセプトによる治療でも観察されました。
Furthermore, dupilumab, which blocks the receptors for IL-4 and IL-13, has recently been considered for the treatment of IgG4-RDs.
さらに、IL-4 および IL-13 の受容体を遮断するデュピルマブは、IgG4-RD の治療に最近検討されています。
These studies point to a role of T cells (possibly TH2 cells) and IL-4 and/or IL-13 in the pathogenesis of IgG4-RDs, as well as indicating an apparent lack of persistence of IgG4 (and IgE) responses in this disease setting, which, at least for IgG4, seems to be a more general phenomenon.
これらの研究は、この疾患設定では IgG4 (および IgE) 応答の持続性が明らかに欠如していることを示していることと同様に、IgG4-RD の病因における T 細胞 (おそらく TH2 細胞) と IL-4 および/または IL-13 の役割を示しています。これは、少なくとも IgG4 については、より一般的な現象のようです。
Future research should focus on determining the sequence of events that lead to the production of IgG4 and confirming or acquitting a pathogenic role for IgG4 in IgG4-RDs.
今後の研究は、IgG4 の産生につながる事象の順序を決定し、IgG4-RD における IgG4 の病原性役割を確認または無罪にすることに焦点を当てる必要があります。
Some reports suggest an increased incidence of malignancies in patients with IgG4-RDs, through the effects of IgG4 on suppressing antitumour immune responses, although this is still a matter of debate.
いくつかの報告は、抗腫瘍免疫応答の抑制に対する IgG4 の影響を通じて、IgG4-RD を持つ患者における悪性腫瘍の発生率の増加を示唆していますが、これはまだ議論の問題です。
Having a tumour may also predispose to developing an IgG4-RD through cytokines secreted by the tumour that induce class-switching to IgG4.
腫瘍があると、IgG4 へのクラススイッチを誘導する腫瘍から分泌されるサイトカインを介して IgG4-RD が発生しやすくなる可能性もあります。
Whether having one type of IgG4-associated disease (IgG4-AID or IgG4-RD) can lead to a second type of IgG4-associated disease should be further investigated.
1 種類の IgG4 関連疾患 (IgG4-AID または IgG4-RD) が 2 番目のタイプの IgG4 関連疾患につながる可能性があるかどうかをさらに調査する必要があります。
Case reports of co-occurrence of IgG4-AIDs with IgG4-RDs do exist but are generally rare.
IgG4-AID と IgG4-RD の同時発生の症例報告は存在しますが、一般的にはまれです。
Future directions
The role of IgG4 antibody responses in physiological and pathological settings is gaining increasing attention.
生理学的および病理学的設定における IgG4 抗体応答の役割は、ますます注目を集めています。
Although, historically, the anti-inflammatory nature of IgG4 was associated with dampening ongoing immune responses, it is increasingly recognized that these antibodies can also cause pathology.
歴史的に、IgG4 の抗炎症性は進行中の免疫応答の抑制に関連していましたが、これらの抗体が病状を引き起こす可能性があることもますます認識されています。
The first steps towards understanding the pathological mechanisms underlying these IgG4-associated diseases have been taken, but little is still known regarding what triggers and maintains these IgG4 responses.
これらの IgG4 関連疾患の根底にある病理学的メカニズムを理解するための第一歩が踏み出されましたが、これらの IgG4 応答を引き起こし、維持するものについてはまだほとんどわかっていません。
There is a clear need to better understand how IgG4 responses are regulated.
IgG4 応答がどのように制御されているかをよりよく理解する必要があることは明らかです。
This knowledge could then form the basis for novel therapeutic strategies targeting these responses.
この知識は、これらの反応を標的とする新しい治療戦略の基礎を形成する可能性があります。
Specifically, the aim would be to stimulate beneficial IgG4 responses, for example in allergy, or to inhibit IgG4 responses in autoimmune diseases and antitumour and anti-biologic responses.
具体的には、例えばアレルギーにおける有益な IgG4 応答を刺激すること、または自己免疫疾患および抗腫瘍および抗生物質応答における IgG4 応答を阻害することです。
Supporting these (pre)clinical ambitions will require better models to study the development of ‘natural’ human IgG4 responses (Box 1).
これらの(前)臨床的野心をサポートするには、「自然な」ヒト IgG4 応答の発生を研究するためのより優れたモデルが必要になります(ボックス 1)。
Box 1
In vitro and in vivo models to study IgG4 responses
IgG4 応答を研究するための in vitro および in vivo モデル
In vitro models
Model systems to study IgG4 in vitro generally involve either cell cultures or, more recently, organoids.
IgG4 を in vitro で研究するためのモデル システムには、通常、細胞培養または最近ではオルガノイドが含まれます。
IgG4+ memory B cells and IgG4-producing plasma cells can survive for several days to weeks in vitro when supplied with suficient trophic factors and cytokines including IL-21 and in the presence of other cell types such as stimulated T cells or CD40L-expressing cell lines.
十分な栄養因子および IL-21 を含むサイトカインが供給され、刺激された T 細胞または CD40L 発現細胞株などの他の細胞型の存在下で、IgG4+記憶B細胞およびIgG4産生形質細胞はin vitroで数日から数週間生存できます。
These models allow for, for example, isolation of antibody sequences, manipulation of antibody glycosylation or study of B cell receptor (BCR) signalling and recall responses.
これらのモデルは、例えば、抗体配列の分離、抗体グリコシル化の操作、または B 細胞受容体 (BCR) シグナル伝達およびリコール応答の研究を可能にします。
IgG4+ B cells readily diferentiate into antibody-secreting cells in vitro, similarly to IgG1+B cells.
IgG4+ B 細胞は、IgG1+ B 細胞と同様に、in vitro で抗体分泌細胞に容易に分化します。
Furthermore, in vitro class-switching of naive human B cells towards IgG4+ B cells has been shown by many groups to proceed readily, in an IL-4-dependent manner.
さらに、ナイーブ ヒト B 細胞の IgG4+ B 細胞への in vitro クラス切り替えは、IL-4 依存的に容易に進行することが多くのグループによって示されています。
Human tonsil organoids are another model system in which to study (de novo) IgG(4) responses.
ヒト扁桃腺オルガノイドは、(de novo) IgG(4) 応答を研究する別のモデル システムです。
These organoids can live for several weeks and produce varying amounts of antibodies, including IgG4.
これらのオルガノイドは数週間生存し、IgG4 を含むさまざまな量の抗体を産生します。
In vivo models
Studies of IgG4 in commonly used experimental animal models are challenging owing to the lack of a proper homologue of human IgG4.
一般的に使用される実験動物モデルでの IgG4 の研究は、ヒト IgG4 の適切な相同体がないため困難です。
Mice and rats each have an IgG subclass that, similar to human IgG4, has limited ability to activate complement, but the properties of these antibodies differ significantly from the anti-inflammatory properties of human IgG4.
マウスとラットはそれぞれ、ヒト IgG4 と同様に補体を活性化する能力が限られている IgG サブクラスを持っていますが、これらの抗体の特性は、ヒト IgG4 の抗炎症特性とは大きく異なります。
In addition, the IgG4-equivalent IgG subclasses do not undergo Fab-arm exchange under physiological conditions.
さらに、IgG4 と同等の IgG サブクラスは、生理学的条件下で Fab アーム交換を受けません。
Non-human primates (such as cynomolgus monkeys and rhesus macaques) have an IgG4 subclass that has many similarities to human IgG4, and these primates are therefore the favoured model for IgG4-related research.
非ヒト霊長類 (カニクイザルやアカゲザルなど) は、ヒト IgG4 と多くの類似点を持つ IgG4 サブクラスを持っているため、これらの霊長類は IgG4 関連の研究に好まれるモデルです。
Humanized animal models provide another method for studying IgG4 responses.
ヒト化動物モデルは、IgG4 応答を研究するための別の方法を提供します。
Recently, human IgG4 responses were successfully induced upon IL-10 exposure of immunodeficient NOD-scid IL2rγnull mice engrafted with human lymphocytes.
最近、ヒト IgG4 応答は、ヒトリンパ球を移植した免疫不全 NOD-scid IL2rγnull マウスの IL-10 暴露により首尾よく誘導されました。
A human IgG4 knock-in mouse model has been developed, but awaits formal peer-reviewed publication.
ヒト IgG4 ノックイン マウス モデルが開発されましたが、正式な査読済みの公開が待たれます。
Introducing human IgG4 into animal models is an interesting strategy but the extrapolation of results may be limited as Fcγ receptors have not been adjusted in these models, and the responses may therefore difer from the human situation.
ヒト IgG4 を動物モデルに導入することは興味深い戦略ですが、これらのモデルでは Fcγ 受容体が調整されていないため、結果の外挿は限られている可能性があります。したがって、反応は人間の状況とは異なる場合があります。
Although IgG(4) anti-drug responses are highly unwanted in patients, they form an important model to study de novo IgG4 responses in a human context.
IgG(4) 抗薬物応答は患者にとって非常に望ましくありませんが、人間のコンテキストで de novo IgG4 応答を研究するための重要なモデルを形成します。
Box 2
Fab-arm exchange
IgG antibodies are typically symmetrical molecules, with the exception of IgG4.
IgG抗体は通常、IgG4を除いて対称分子です。
IgG antibodies are covalent dimers of two half-molecules, each half consisting of a light chain and a heavy chain (see the figure).
IgG 抗体は、2 つの半分子の共有結合二量体であり、それぞれの半分は軽鎖と重鎖で構成されています (図を参照)。
The antibodies produced by plasma cells are bivalent and monospecific (having two antigen-binding sites of the same specificity).
形質細胞によって産生される抗体は、二価で単一特異性です (同じ特異性の 2 つの抗原結合部位を持っています)。
However, human IgG4 is an unusually dynamic antibody, and these half-molecules can dissociate and recombine with other IgG4 half-molecules in a process termed Fab (fragment antigen binding)-arm exchange.
しかし、ヒト IgG4 は異常に動的な抗体であり、これらの半分子は、Fab (フラグメント抗原結合) アーム交換と呼ばれるプロセスで解離し、他の IgG4 半分子と再結合することができます。
The resulting IgG4 antibodies are asymmetrical, bispecific structures with two diferent antigen-binding sites.
得られた IgG4 抗体は、2 つの異なる抗原結合部位を持つ非対称の二重特異性構造です。
Owing to the heterogeneity of the IgG4 pool, up to 99% of circulating IgG4 is considered to be bispecific.
IgG4 プールの不均一性により、循環 IgG4 の最大 99% が二重特異性であると見なされます。
Two features of IgG4 enable Fab-arm exchange to occur: the serine at position 228 (S228) in the core hinge and the arginine at position 409 (R409) in the CH3 domain.
IgG4 の 2 つの機能により、Fab アームの交換が可能になります。コア ヒンジの 228 位 (S228) のセリンと、CH3 ドメインの 409 位 (R409) のアルギニンです。
The former results in unusually labile covalent interactions between the two heavy chains through disulfide bonds.
前者は、ジスルフィド結合を介した 2 つの重鎖間の異常に不安定な共有相互作用をもたらします。
The latter weakens the non-covalent association between the two heavy chains through their CH3 domains.
後者は、CH3 ドメインを介して 2 つの重鎖間の非共有結合を弱めます。
Both features are required for Fab-arm exchange, as introducing only one of these residues in IgG1 does not result in Fab-arm exchange in vivo.
これらの残基の 1 つだけを IgG1 に導入しても、in vivo での Fab アーム交換は発生しないため、両方の機能が Fab アーム交換に必要です。
However, in vitro, where redox conditions can be varied more widely, a K409R mutant of IgG1 can be forced to participate in Fab-arm exchange in the presence of stronger reducing conditions than are typically required for IgG4 Fab-arm exchange.
しかし、レドックス条件をより広く変化させることができる in vitro では、IgG1 の K409R 変異体は、IgG4 Fab アーム交換に通常必要とされる条件よりも強い還元条件の存在下で、強制的に Fab アーム交換に参加させることができます。
This forms the basis of the Duobody platform, which enables the production of bispecific therapeutic antibodies through in vitro Fab-arm exchange that are stable in vivo.
これは、in vivo で安定な in vitro Fab アーム交換による二重特異性治療抗体の産生を可能にする Duobody プラットフォームの基礎を形成します。
For the other human IgG subclasses, particularly IgG3, the interactions between CH3 domains are also a lot weaker than for IgG1, but this is compensated by multiple disulfide bonds between heavy chains that are not present in IgG4.
他のヒト IgG サブクラス、特に IgG3 の場合、CH3 ドメイン間の相互作用も IgG1 よりもはるかに弱いですが、これは IgG4 には存在しない重鎖間の複数のジスルフィド結合によって補償されます。
A functional consequence of Fab-arm exchange is the effective monovalency of IgG4.
Fab アーム交換の機能的な結果は、IgG4 の効果的な一価性です。
The random nature of the exchange process results in the antigen specificity of one Fab arm recombining with any number of other, usually unrelated, antigen specificities in the other Fab arm.
交換プロセスのランダムな性質により、1 つの Fab アームの抗原特異性が、他の Fab アームの任意の数の他の (通常は無関係な) 抗原特異性と組み換えられます。
This eliminates the potential of the antibody to cross-link antigen, further minimizing the immune-activating potential of IgG4.
これにより、抗体が抗原を架橋する可能性が排除され、IgG4 の免疫活性化の可能性がさらに最小限に抑えられます。
Complement activation also seems to be further impaired by Fab-arm exchange.
補体の活性化は、Fab-arm交換によってさらに損なわれるようです。
以下省略。
この記事が気に入ったらサポートをしてみませんか?