BA.2/5 亜種の上部および下部呼吸器系における複製と固有の病原性の異なる軌跡

Divergent trajectory of replication and intrinsic pathogenicity of SARS-CoV-2 Omicron post-BA.2/5 subvariants in the upper and lower respiratory tract
SARS-CoV-2 オミクロン BA.2/5 亜種の上部および下部呼吸器系における複製と固有の病原性の異なる軌跡
Summary
Background
Earlier Omicron subvariants including BA.1, BA.2, and BA.5 emerged in waves, with a subvariant replacing the previous one every few months. More recently, the post-BA.2/5 subvariants have acquired convergent substitutions in spike that facilitated their escape from humoral immunity and gained ACE2 binding capacity. However, the intrinsic pathogenicity and replication fitness of the evaluated post-BA.2/5 subvariants are not fully understood.
BA.1、BA.2、BA.5 などの以前のオミクロン サブバリアントは波状に出現し、数か月ごとにサブバリアントが以前のサブバリアントに取って代わりました。最近では、ポスト BA.2/5 サブバリアントはスパイクの収束的置換を獲得し、体液性免疫からの脱出を容易にし、ACE2 結合能力を獲得しました。ただし、評価されたポスト BA.2/5 サブバリアントの固有の病原性と複製適応性は完全には理解されていません。
Methods
We systemically investigated the replication fitness and intrinsic pathogenicity of representative post-BA.2/5 subvariants (BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5) in weanling (3–4 weeks), adult (8–10 weeks), and aged (10–12 months) mice. In addition, to better model Omicron replication in the human nasal epithelium, we further investigated the replication capacity of the post-BA.2/5 subvariants in human primary nasal epithelial cells.
我々は、離乳マウス(3~4週)、成体マウス(8~10週)、老齢マウス(10~12か月)における代表的なポストBA.2/5サブバリアント(BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5)の複製適応度と固有の病原性を体系的に調査しました。さらに、ヒト鼻腔上皮におけるオミクロン複製をより適切にモデル化するために、ポストBA.2/5サブバリアントのヒト一次鼻腔上皮細胞における複製能力をさらに調査しました。
Findings
We found that the evaluated post-BA.2/5 subvariants are consistently attenuated in mouse lungs but not in nasal turbinates when compared with their ancestral subvariants BA.2/5. Further investigations in primary human nasal epithelial cells revealed a gained replication fitness of XBB.1 and XBB.1.5 when compared to BA.2 and BA.5.2.
評価された BA.2/5 以降のサブバリアントは、その祖先サブバリアント BA.2/5 と比較した場合、マウスの肺では一貫して弱毒化されているが、鼻甲介では弱毒化されていないことがわかりました。一次ヒト鼻上皮細胞でのさらなる調査により、BA.2 および BA.5.2 と比較した場合、XBB.1 および XBB.1.5 の複製適応度が向上していることが明らかになりました。
Interpretation
Our study revealed that the post-BA.2/5 subvariants are attenuated in lungs while increased in replication fitness in the nasal epithelium, indicating rapid adaptation of the circulating Omicron subvariants in the human populations.
私たちの研究により、BA.2/5以降の亜変異体は肺では弱毒化される一方で、鼻上皮での複製適応度は増加していることが明らかになりました。これは、循環するオミクロン亜変異体がヒト集団内で急速に適応していることを示唆しています。
Funding
The full list of funding can be found at the Acknowledgements section.
Introduction
SARS-CoV-2 Omicron (B.1.1.529) was designated as the fifth variant of concern (VOC) by the WHO on November 26th, 2021, after Alpha, Beta, Gamma, and Delta. Omicron contains over 30 changes in spike when compared to ancestral SARS-CoV-2, leading to its robust capacity in evading humoral immunity and high transmissibility.
SARS-CoV-2 オミクロン(B.1.1.529)は、2021年11月26日にWHOによって、アルファ、ベータ、ガンマ、デルタに続く5番目の懸念される変異体(VOC)に指定されました。オミクロンは、祖先のSARS-CoV-2と比較してスパイクに30を超える変化があり、体液性免疫を回避する強力な能力と高い伝染性につながっています。
Omicron quickly replaced Delta as the dominant circulating SARS-CoV-2 variant and is currently the only circulating SARS-CoV-2 VOC as of July 2023.
オミクロンは急速にデルタに取って代わり、SARS-CoV-2の優勢な循環型変異体となり、2023年7月現在、唯一の循環型SARS-CoV-2 VOCとなっています。
Continuous surveillance of Omicron evolution revealed that the subsequently emerged SARS-CoV-2 Omicron subvariants frequently outcompeted previously dominant Omicron subvariants every few months, in the order of BA.1, BA.2, and BA.5.
オミクロンの進化を継続的に監視した結果、その後出現したSARS-CoV-2オミクロン亜種が、BA.1、BA.2、BA.5の順に、数か月ごとに以前に優勢だったオミクロン亜種を頻繁に打ち負かしていることが明らかになりました。
More recently, although additional Omicron subvariants have emerged after BA.5, these newly emerged subvariants were unable to successfully outcompete other subvariants to become the dominant Omicron lineage worldwide, leading to the cocirculation of multiple Omicron subvariants (Fig. 1a).
最近では、BA.5の後にさらにオミクロン亜種が出現しましたが、これらの新しく出現した亜種は他の亜種を打ち負かして世界中で優勢なオミクロン系統になることができず、複数のオミクロン亜種が共存する結果となりました(図1a)。

Interestingly, these different post-BA.2 and post-BA.5 subvariants have similarly acquired convergent substitutions at a number of specific locations on spikes, including R346, K444, L452, N460, and F486, that contribute to immune evasion or increase in ACE2 binding affinity.
興味深いことに、これらの異なるポスト BA.2 およびポスト BA.5 サブバリアントは、R346、K444、L452、N460、および F486 を含むスパイク上の特定の場所で、免疫回避または ACE2 結合親和性の増加に寄与する収束的置換を同様に獲得しています。
The pathogenicity of previous dominant Omicron variants, BA.1, BA.2, and BA.5 have been evaluated in vivo, and were in general demonstrated to be attenuated when compared with ancestral SARS-CoV-2 or previous VOCs.
以前の優勢なオミクロン変異体である BA.1、BA.2、および BA.5 の病原性はin vivoで評価されており、祖先の SARS-CoV-2 または以前の VOC と比較すると、一般的に弱毒化されていることが実証されています。
In the current study, we further investigated the intrinsic pathogenicity and replication fitness of a number of representative post-BA.2/5 subvariants carrying the convergent substitutions that confer immune evasion or increase in ACE2 binding affinity.
本研究では、免疫回避または ACE2 結合親和性の増加をもたらす収束的置換を持つ、代表的な BA.2/5 以降のサブ変異体の固有の病原性と複製適合性をさらに調査しました。
We demonstrated that the intrinsic pathogenicity and replication fitness of the evaluated post-BA.2/5 subvariants (BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, XBB.1.5) are attenuated in the lungs but not in the nasal turbinates of infected mice.
評価された BA.2/5 以降のサブ変異体 (BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5) の固有の病原性と複製適合性は、感染したマウスの肺では弱毒化されていますが、鼻甲介では弱毒化されていないことが実証されました。
Furthermore, in primary human nasal epithelial cells, XBB.1 and XBB.1.5 demonstrated higher replication fitness than that of BA.2 and BA.5.2.
さらに、初代ヒト鼻上皮細胞において、XBB.1 および XBB.1.5 は BA.2 および BA.5.2 よりも高い複製適合性を示しました。
Results
Post-BA.2/5 subvariants including BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5
BA.2/5以降の亜種はBL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5を含む
BA.2 contains eight spike substitutions that are not present in BA.1, including three substitutions (T19I, L24S, V213G) and one deletion (25–27 deletion) in the N-terminal domain (NTD), and four substitutions (S371F, T376A, D405N, R408S) in the receptor-binding domain (RBD).
BA.2 には、BA.1 には存在しない 8 つのスパイク置換が含まれています。これには、N 末端ドメイン (NTD) の 3 つの置換 (T19I、L24S、V213G) と 1 つの欠失 (25~27 欠失)、および受容体結合ドメイン (RBD) の 4 つの置換 (S371F、T376A、D405N、R408S) が含まれます。
BA.5 and BA.2.75 are two phylogenetically independent subvariants that are believed to diversify independently from BA.2.
BA.5 と BA.2.75 は、BA.2 とは独立して多様化すると考えられる、系統学的に独立した 2 つのサブバリアントです。
Compared with BA.2, BA.5 possesses three additional changes (69-70del, L452R, and F486V) and one reversion mutation (R493Q) in its spike (Fig. 1b).
BA.2 と比較すると、BA.5 はスパイクに 3 つの追加変更 (69-70del、L452R、F486V) と 1 つの復帰突然変異 (R493Q) を持っています (図 1b)。

BQ.1 and BQ.1.1 are descendants of BA.5, which possess four (K444T, L452R, N460K, and F486V) or all five (R346T, K444T, L452R, N460K, and F486V) convergent substitutions on spikes, respectively (Fig. 1b).
BQ.1 と BQ.1.1 は BA.5 の子孫で、スパイクにそれぞれ 4 つの収束置換 (K444T、L452R、N460K、F486V) または 5 つの収束置換 (R346T、K444T、L452R、N460K、F486V) を持っています (図 1b)。
BL.1 and CH.1.1 originated from BA.2.75, which is a progeny of Omicron BA.2.
BL.1 と CH.1.1 は、Omicron BA.2 の子孫である BA.2.75 に由来しています。
BL.1 and CH.1.1 have acquired two (R346T and N460K) or all five (R346T, K444T, L452R, N460K, and F486S) convergent substitutions on spikes when compared to BA.2, respectively (Fig. 1b).
BL.1 と CH.1.1 は、それぞれ BA.2 と比較すると、スパイクに 2 つの収束的置換 (R346T と N460K) または 5 つの収束的置換 (R346T、K444T、L452R、N460K、および F486S) を獲得しています (図 1b)。
XBB sublineages likely originated through the recombination of two BA.2 descendants, BJ.1 and BM.1.1.1 (progeny of BA.2.75).
XBB 亜系統は、2 つの BA.2 子孫、BJ.1 と BM.1.1.1 (BA.2.75 の子孫) の組み換えによって発生したと考えられます。
XBB.1 spike contains R346T, N460K, and F486S, while XBB.1.5 possesses R346T, N460K, and F486P on spike (Fig. 1b).
XBB.1 スパイクには R346T、N460K、および F486S が含まれ、XBB.1.5 にはスパイクに R346T、N460K、および F486P が含まれます (図 1b)。
Replication fitness of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 in the nasal turbinates of weanling, adult, and aged C57BL/6J mice
離乳児、成体、老齢の C57BL/6J マウスの鼻甲介における BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5 の複製適応度
We first explored the replication fitness of the post-BA.2/5 subvariants in the nasal turbinates of infected animals (Fig. 2a–f).
まず、感染動物の鼻甲介における BA.2/5 以降の亜変異体の複製適応度を調査しました (図 2a~f)。

To this end, we intranasally challenged weanling C57BL/6J mice (3–4 weeks), adult C57BL/6J mice (8–10 weeks), and aged C57BL/6J mice (10–12 months), with 1 × 10⁵ PFU of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1 or XBB.1.5.
この目的のため、離乳した C57BL/6J マウス (3~4 週)、成体 C57BL/6J マウス (8~10 週)、老齢 C57BL/6J マウス (10~12 か月) に、BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、または XBB.1.5 の 1 × 10⁵ PFU を鼻腔内投与しました。
In addition, we included BA.2 (ancestral of BL.1, XBB.1, CH.1.1, and XBB.1.5) and BA.5.2 (ancestral of BQ.1 and BQ.1.1) as comparison groups.
さらに、比較グループとして BA.2 (BL.1、XBB.1、CH.1.1、XBB.1.5 の祖先) と BA.5.2 (BQ.1 および BQ.1.1 の祖先) を含めました。
We sacrificed the infected mice at 2 days post infection (dpi) to collect nasal turbinate tissues for virological assessment.
感染後 2 日目に感染マウスを屠殺し、ウイルス学的評価のために鼻甲介組織を採取しました。
We found that BQ.1 and BQ.1.1, which are descendants of BA.5.2, replicated less efficiently than BA.5.2 in the nasal turbinates of infected adult mice, as evidenced by their significantly lower subgenomic envelope (sgE) gene expression and infectious titer when compared with BA.5.2 (Fig. 2b and e).
BA.5.2 の子孫である BQ.1 と BQ.1.1 は、感染した成体マウスの鼻甲介で BA.5.2 よりも複製効率が低いことがわかりました。これは、BA.5.2 と比較した場合、サブゲノムエンベロープ (sgE) 遺伝子発現と感染力価が著しく低いことから明らかです (図 2b および e)。
In the nasal turbinates of infected weanling and aged mice, BQ.1 and BQ.1.1 replicated to similar levels when compared with BA.5.2 (Fig. 2a, c, d and f).
感染した離乳マウスと老齢マウスの鼻甲介では、BQ.1 と BQ.1.1 は BA.5.2 と比較して同様のレベルで複製されました (図 2a、c、d、f)。
Descendants of BA.2 including BL.1, XBB.1, CH.1.1, and XBB.1.5, replicated to largely comparable levels with that of BA.2 in the nasal turbinates of infected mice of all three age groups (Fig. 2a–f).
BL.1、XBB.1、CH.1.1、XBB.1.5を含むBA.2の子孫は、3つの年齢群すべての感染マウスの鼻甲介において、BA.2とほぼ同等のレベルで複製されました(図2a~f)。
Antigen expression and intrinsic pathogenicity of post-BA.2/5 sublineages in the nasal turbinates of weanling, adult, and aged C57BL/6J mice
離乳児、成体、老齢の C57BL/6J マウスの鼻甲介における BA.2/5 以降の亜系統の抗原発現と固有の病原性
We next performed immunohistochemistry staining to detect the level of SARS-CoV-2 nucleocapsid (N) protein expression in the nasal turbinates of infected mice.
次に、感染したマウスの鼻甲介における SARS-CoV-2 ヌクレオカプシド (N) タンパク質の発現レベルを検出するために免疫組織化学染色を行いました。
In keeping with previous reports from us and others, N protein was readily detected in the nasal epithelium of BA.2- and BA.5.2-infected C57BL/6J mice (Fig. 3a–e).
私たちや他の研究者による以前の報告と一致して、BA.2 および BA.5.2 に感染した C57BL/6J マウスの鼻上皮で N タンパク質が容易に検出されました (図 3a~e)。


We found that N expression levels in the nasal turbinates of BQ.1- or BQ.1.1-infected adult mice were significantly lower than that of BA.5.2-infected adult mice (Fig. 3b and e).
BQ.1 または BQ.1.1 に感染した成体マウスの鼻甲介における N 発現レベルは、BA.5.2 に感染した成体マウスのそれよりも有意に低いことがわかりました (図 3b および e)。
Similarly, infection of BQ.1 or BQ.1.1 also resulted in lower N expression than that of BA.5.2 in nasal turbinates of infected weanling and aged mice but the difference did not reach statistical significance (Fig. 3a, c and e).
同様に、BQ.1 または BQ.1.1 に感染すると、感染した離乳児および老齢マウスの鼻甲介における N 発現も BA.5.2 よりも低くなりますが、その差は統計的に有意には達しません (図 3a、c、e)。
In parallel, the expression of N protein in the nasal epithelium of BL.1-, XBB.1-, CH.1.1-, and XBB.1.5-infected mice was detected at a similar level when compared with that of BA.2 in infected mice of all three age groups (Fig. 3a–c and e).
同時に、BL.1、XBB.1、CH.1.1、XBB.1.5 に感染したマウスの鼻上皮における N タンパク質の発現は、3 つの年齢群すべての感染マウスにおける BA.2 の発現と同程度であることが検出された (図 3a~c および e)。
We then evaluated the N expression rate in the nasal turbinate of infected mice against the peak count date of the different Omicron subvariants with linear regression.
次に、感染マウスの鼻甲介における N 発現率を、異なる Omicron サブバリアントのピークカウント日に対して線形回帰で評価した。
While no trend was observed for weanling and adult mice, a weak trend of reduced replication was observed in aged mice (−R = 0.42, p = 0.0072) (Fig. 3d).
離乳マウスと成体マウスでは傾向は見られなかったが、高齢マウスでは複製が減少する弱い傾向が見られた(−R = 0.42、p = 0.0072)(図3d)。

In parallel, we examined histopathological changes of nasal turbinate tissues collected from infected mice at 2 dpi with haematoxylin and eosin (H&E) staining.
並行して、感染マウスから2 dpi に採取した鼻甲介組織の組織病理学的変化をヘマトキシリンおよびエオシン(H&E)染色で調べた。
We found that while the nasal turbinates of BQ.1-, BQ.1.1-, XBB.1- and CH.1.1- infected mice demonstrated a milder level of epithelium destruction and submucosal immune cells infiltration when compared to BA.2-/BA.5.2-infected mice, no apparent damage in the nasal turbinate was observed after BL.1 and XBB.1.5 infection (Fig. 4).
BQ.1、BQ.1.1、XBB.1、CH.1.1 に感染したマウスの鼻甲介では、BA.2/BA.5.2 に感染したマウスと比較して上皮破壊と粘膜下免疫細胞浸潤のレベルが軽度であったものの、BL.1 および XBB.1.5 感染後には鼻甲介に明らかな損傷は見られなかったことがわかりました (図 4)。

Together, these results indicated that while subtle differences might be observed for individual subvariants, including the reduced replication of BQ.1/BQ.1.1 and the low tissue damage of BL.1/XBB.1.5, the overall replication fitness and pathogenicity of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 are largely comparable with that of BA.2 and BA.5.2 in the nasal turbinates of infected mice.
これらの結果を総合すると、BQ.1/BQ.1.1 の複製の減少や BL.1/XBB.1.5 の組織損傷の低さなど、個々のサブバリアントで微妙な違いが見られるかもしれないが、感染したマウスの鼻甲介における BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、および XBB.1.5 の全体的な複製適応度および病原性は、BA.2 および BA.5.2 とほぼ同等であることが示唆されました。
Replication fitness of post-BA.2/5 sublineages in the lungs of weanling, adult, and aged C57BL/6J mice
離乳児、成体、老齢の C57BL/6J マウスの肺における BA.2/5 以降の亜系統の複製適応度
Next, we investigated the replication fitness of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 in the lungs of infected animals harvested at 2 dpi.
次に、感染後 2 日目に採取した感染動物の肺における BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5 の複製適応度を調査しました。
We found that the virus gene copy and infectious titer of BA.5.2 progenies, BQ.1 and BQ.1.1, were both lower than that of BA.5.2 (Fig. 5a–f).
BA.5.2 の子孫である BQ.1 と BQ.1.1 のウイルス遺伝子コピー数と感染力価は、どちらも BA.5.2 よりも低いことがわかりました (図 5a~f)。

Similarly, the virus replication of BA.2 progenies, BL.1, XBB.1, CH.1.1, and XBB.1.5, were consistently less efficient when compared with BA.2 in the mouse lungs (Fig. 5a–f).
同様に、マウスの肺における BA.2 子孫である BL.1、XBB.1、CH.1.1、XBB.1.5 のウイルス複製は、BA.2 と比較して一貫して効率が低かった (図 5a~f)。
The lowered replication of BA.2/5 progenies was most appreciable in the lungs of adult mice.
BA.2/5 子孫の複製の低下は、成体マウスの肺で最も顕著であった。
In particular, the infectious titers of BQ.1 and BQ.1.1 were 55.0- (p = 0.0009) and 23.5- (p = 0.0011) folds lower than that of BA.5.2, respectively.
特に、BQ.1 と BQ.1.1 の感染力価は、BA.5.2 の感染力価よりそれぞれ 55.0 倍 (p = 0.0009) と 23.5 倍 (p = 0.0011) 低かった。
Meanwhile, the infectious titers of BL.1, XBB.1, CH.1.1, and XBB.1.5 were 19.6- (p = 0.013), 95.7- (p = 0.0092), 9.6- (p = 0.019), and 74.4-folds (p = 0.0094) lower when compared with BA.2, respectively (Fig. 5e).
一方、BL.1、XBB.1、CH.1.1、XBB.1.5の感染力価は、BA.2と比較するとそれぞれ19.6倍(p = 0.013)、95.7倍(p = 0.0092)、9.6倍(p = 0.019)、74.4倍(p = 0.0094)低かった(図5e)。
Antigen expression and intrinsic pathogenicity of post-BA.2/5 sublineages in the lungs of weanling, adult, and aged C57BL/6J mice
離乳児、成体、老齢の C57BL/6J マウスの肺における BA.2/5 以降の亜系統の抗原発現と固有の病原性
In parallel, we performed immunohistochemistry staining to detect the amount of SARS-CoV-2 nucleocapsid (N) protein in the lungs of virus-infected mice.
並行して、ウイルスに感染したマウスの肺における SARS-CoV-2 ヌクレオカプシド (N) タンパク質の量を検出するため、免疫組織化学染色を実施しました。
In keeping with the virus gene copy and infectious titer results, the amount of N detected in the lungs of BQ.1- and BQ.1.1-infected mice were significantly lower when compared to BA.5.2, while the amount of N detected in lungs of BL.1-, XBB.1-, CH.1.1-, and XBB.1.5-infected mice were significantly lower when compared to BA.2 (Fig. 6a–c and e).
ウイルス遺伝子コピーおよび感染力価の結果と一致して、BQ.1 および BQ.1.1 に感染したマウスの肺で検出された N の量は BA.5.2 と比較して有意に低く、BL.1、XBB.1、CH.1.1、および XBB.1.5 に感染したマウスの肺で検出された N の量は BA.2 と比較して有意に低かったです (図 6a~c および e)。


In the lungs of adult mice, N expression following BQ.1 and BQ.1.1 infection were 7.9- (p = 0.0059) and 2.5- (p = 0.041) folds lower than that of BA.5.2, respectively.
成体マウスの肺では、BQ.1 および BQ.1.1 感染後の N 発現は、BA.5.2 と比較してそれぞれ 7.9 倍 (p = 0.0059)、2.5 倍 (p = 0.041) 低かった。
Meanwhile, N expression upon BL.1, XBB.1, CH.1.1, and XBB.1.5 infection were 7.5- (p < 0.0001), 4.4- (p = 0.0003), 4.7- (p = 0.0002), and 74.5-folds (p < 0.0001) lower when compared with BA.2, respectively (Fig. 6b and e).
一方、BL.1、XBB.1、CH.1.1、XBB.1.5 感染時の N 発現は、BA.2 と比較してそれぞれ 7.5 倍 (p < 0.0001)、4.4 倍 (p = 0.0003)、4.7 倍 (p = 0.0002)、74.5 倍 (p < 0.0001) 低かった (図 6b および e)。
We then evaluated the N expression rate in the lungs of infected mice against the peak count date of the different Omicron subvariants with linear regression.
次に、感染マウスの肺における N 発現率を、さまざまな Omicron サブバリアントのピークカウント日に対して線形回帰で評価した。
Remarkably, we found a significant trend of reduced N expression from BA.2 to XBB.1.5 in mice of all three age groups, including weanling mice (−R = 0.79, p < 0.0001), adult mice (−R = 0.73, p < 0.0001), and aged mice (−R = 0.73, p < 0.0001) (Fig. 6d).
注目すべきことに、離乳マウス(−R = 0.79、p < 0.0001)、成体マウス(−R = 0.73、p < 0.0001)、老齢マウス(−R = 0.73、p < 0.0001)を含む3つの年齢群のマウスすべてにおいて、BA.2からXBB.1.5へのN発現の減少の顕著な傾向が見られました(図6d)。
We next examined the histopathological changes in mouse lungs upon virus infection with haematoxylin and eosin (H&E) staining and quantified lung pathology scores based on a semi-quantitative protocol (Fig. 7a–g).
次に、ウイルス感染後のマウス肺の組織病理学的変化をヘマトキシリンおよびエオシン(H&E)染色で調べ、半定量的プロトコルに基づいて肺病理スコアを定量化しました(図7a~g)。

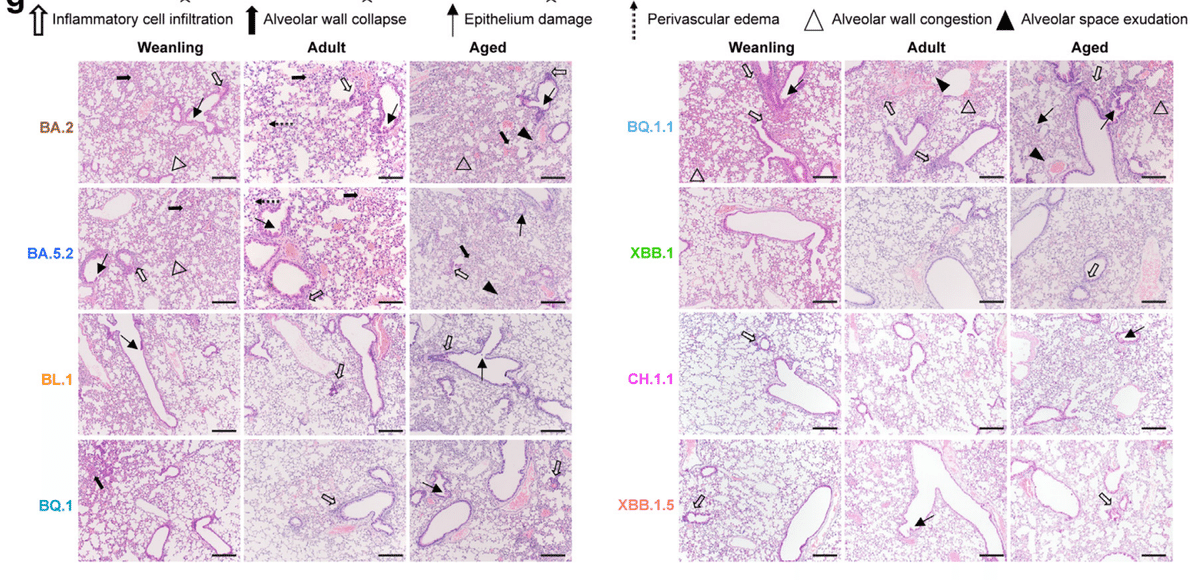
The pathological changes of lungs from BA.2- or BA.5.2-infected mice were featured by diffused alveolar infiltration, bronchiolar epithelial cell death, and alveolar wall collapse (Fig. 7a–c and g).
BA.2またはBA.5.2に感染したマウスの肺の病理学的変化は、拡散した肺胞浸潤、細気管支上皮細胞死、および肺胞壁の崩壊を特徴としていました(図7a~cおよびg)。
The lungs of BQ.1.1-infected mice demonstrated mild peribronchiolar infiltration and bronchiolar epithelial desquamation.
BQ.1.1 に感染したマウスの肺では、軽度の細気管支周囲浸潤と細気管支上皮の剥離がみられました。
BL.1-, BQ.1-, XBB.1-, CH.1.1-, and XBB.1.5-infected weanling and adult mice showed no apparent alveolar damage with mild perivascular and peribronchiolar immune cell infiltration and epithelium cell death (Fig. 7a, b and g), while lung damages in aged mice were slightly more severe, manifested as conspicuous perivascular infiltration and epithelial cell detachment (Fig. 7c and g).
BL.1、BQ.1、XBB.1、CH.1.1、XBB.1.5 に感染した離乳マウスと成体マウスでは、血管周囲と細気管支周囲に軽度の免疫細胞浸潤と上皮細胞死が見られましたが、明らかな肺胞損傷は見られませんでした (図 7a、b、g)。一方、老齢マウスの肺損傷はやや重度で、顕著な血管周囲浸潤と上皮細胞剥離が見られました (図 7c、g)。
Based on the lung pathology scores and the peak count date of BA.2, BA.5.2, BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5, we performed an ordinal logistic regression (OLR) and found that the evaluated Omicron subvariants are significantly declining in intrinsic lung pathology over time in weanling (p = 0.027) and adult (p = 0.023) mice (Fig. 7d and e).
BA.2、BA.5.2、BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5 の肺病理スコアとピークカウント日に基づいて、順序ロジスティック回帰 (OLR) を実行したところ、評価した Omicron サブバリアントは、離乳マウス (p = 0.027) と成体マウス (p = 0.023) で時間の経過とともに内因性肺病理が著しく減少していることがわかりました (図 7d および e)。
A modest trend of attenuation in lung pathology was observed in aged mice but the difference was not statistically significant (p = 0.075) (Fig. 7f).
老齢マウスでは肺病理の緩やかな減少傾向が見られましたが、その差は統計的に有意ではありませんでした (p = 0.075) (図 7f)。
Together, the histopathological findings are in line with the virological assessments, which collectively indicate that the replication and intrinsic pathogenicity of the post-BA.2/5 subvariants, BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5, are attenuated when compared with their ancestral BA.2 and BA.5.2 subvariants in lungs.
組織病理学的所見はウイルス学的評価と一致しており、全体として、肺におけるポスト BA.2/5 サブバリアント BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、および XBB.1.5 の複製および固有の病原性は、祖先の BA.2 および BA.5.2 サブバリアントと比較して弱まっていることを示しています。
In addition, we intranasally challenged 8–10 weeks old K18-hACE2 mice with 5 × 10³ PFU of BA.2, BA.5.2, BQ.1, BQ.1.1, XBB.1 or XBB.1.5.
さらに、8~10週齢のK18-hACE2マウスに、5×10³ PFUのBA.2、BA.5.2、BQ.1、BQ.1.1、XBB.1、またはXBB.1.5を鼻腔内投与しました。
We sacrificed the infected mice at 2 dpi to collect nasal turbinate and lung tissues for virological assessment.
感染マウスを2日後に屠殺し、ウイルス学的評価のために鼻甲介と肺組織を採取しました。
In the nasal turbinates of infected K18-hACE2 mice, while the replication of BA.5.2 descendants including BQ.1 and BQ.1.1 was largely similar when compared to their ancestral strain BA.5.2, the replication of XBB.1 (9.4-folds) and XBB.1.5 (26.4-folds) were more robust than that of their ancestral strain BA.2 (Supplementary Figure S1a).
感染したK18-hACE2マウスの鼻甲介では、BQ.1およびBQ.1.1を含むBA.5.2子孫の複製は、その祖先株BA.5.2とほぼ同様でしたが、XBB.1(9.4倍)およびXBB.1.5(26.4倍)の複製は、その祖先株BA.2よりも強力でした(補足図S1a)。
Next, we examined the replication capacity of SARS-CoV-2 Omicron post-BA.2/5 subvariants in the lungs of infected K18-hACE2 mice.
次に、感染したK18-hACE2マウスの肺におけるSARS-CoV-2 OmicronのBA.2/5以降の亜種の複製能力を調べました。
We found that the sgE copies of BA.5.2 progenies, BQ.1 and BQ.1.1, were modestly lower than that of BA.5.2 by 2.1-folds and 1.4-folds, respectively.
BA.5.2の子孫であるBQ.1とBQ.1.1のsgEコピーは、BA.5.2のそれよりもそれぞれ2.1倍と1.4倍わずかに低いことがわかりました。
Similarly, the sgE copies of BA.2 progenies, XBB.1 and XBB.1.5, were 18.4-folds and 64.1-folds lower when compared to that of BA.2 in the infected mouse lungs, respectively (Supplementary Figure S1b).
同様に、感染したマウスの肺におけるBA.2の子孫であるXBB.1とXBB.1.5のsgEコピーは、BA.2と比較してそれぞれ18.4倍と64.1倍低かったです(補足図S1b)。
In parallel, we assessed the pattern of viral antigen expression in the lungs of the infected K18-hACE2 mice.
並行して、感染した K18-hACE2 マウスの肺におけるウイルス抗原発現のパターンを評価しました。
In keeping with the virus gene copy results, the expression of N protein in the lungs of XBB.1- and XBB.1.5-infected mice was lower when compared to BA.2, while the expression of N protein in lungs of BQ.1- and BQ.1.1-infected mice was moderately lower when compared to BA.5.2 (Supplementary Figure S1c).
ウイルス遺伝子コピーの結果と一致して、XBB.1 および XBB.1.5 に感染したマウスの肺における N タンパク質の発現は BA.2 と比較して低く、BQ.1 および BQ.1.1 に感染したマウスの肺における N タンパク質の発現は BA.5.2 と比較して中程度に低かったです (補足図 S1c)。
Overall, the pattern of Omicron subvariant replication in the lungs of K18-hACE2 transgenic mice was largely in agreement with results obtained from the wild-type C57BL/6J mouse models.
全体として、K18-hACE2 トランスジェニック マウスの肺における Omicron サブバリアントの複製パターンは、野生型 C57BL/6J マウス モデルから得られた結果とほぼ一致しました。
Replication fitness of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 in human lung epithelial cell Calu3 and primary human nasal epithelial cells
ヒト肺上皮細胞Calu3および初代ヒト鼻上皮細胞におけるBL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、およびXBB.1.5の複製適応度
To assess the replication kinetics of different Omicron post-BA.2/5 subvariants in vitro, we infected VeroE6-TMPRSS2 cells with BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, or XBB.1.5 and harvested supernatant samples at different time points post infection.
さまざまな Omicron の BA.2/5 後サブバリアントの複製速度を in vitro で評価するために、VeroE6-TMPRSS2 細胞に BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、または XBB.1.5 を感染させ、感染後のさまざまな時点で上清サンプルを採取しました。
By quantification of RNA-dependent-RNA-polymerase gene (RdRp) expression, we found that the replication fitness of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 were highly comparable with their ancestral subvariants BA.2/BA.5.2 in the VeroE6-TMPRSS2 cells, as all evaluated Omicron subvariants replicated to similar peak values with similar replication kinetics (Fig. 8a).
RNA 依存性 RNA ポリメラーゼ遺伝子 (RdRp) 発現の定量化により、BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、および XBB.1.5 の複製適合度は、VeroE6-TMPRSS2 細胞における祖先サブバリアント BA.2/BA.5.2 と非常によく似ていることがわかりました。評価したすべての Omicron サブバリアントは、同様の複製速度で同様のピーク値に複製されました (図 8a)。

In parallel, we assessed spike cleavage in the infected VeroE6-TMPRSS2 cells, and found similar cleavage efficiency of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 spike when compared with their ancestral subvariants BA.2/BA.5 (Supplementary Figure S2).
並行して、感染した VeroE6-TMPRSS2 細胞におけるスパイク切断を評価し、祖先サブバリアント BA.2/BA.5 と比較した場合、BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、および XBB.1.5 スパイクの切断効率は同様であることがわかりました (補足図 S2)。
Next, we evaluated the replication kinetics of Omicron post-BA.2/5 subvariants in human lung epithelial cells, Calu3.
次に、ヒト肺上皮細胞 Calu3 における Omicron ポスト BA.2/5 サブバリアントの複製速度を評価しました。
Our data demonstrated that the replication of BA.2 descendants including BL.1, XBB.1, CH.1.1, and XBB.1.5 was significantly less efficient when compared to BA.2 in Calu3 cells (Fig. 8b).
データから、BL.1、XBB.1、CH.1.1、および XBB.1.5 を含む BA.2 子孫の複製は、Calu3 細胞における BA.2 と比較して大幅に効率が低いことがわかりました (図 8b)。

Similarly, BQ.1 and BQ.1.1 did not replicate as efficiently as BA.5.2 (Fig. 8b).
同様に、BQ.1 および BQ.1.1 は BA.5.2 ほど効率的に複製されませんでした (図 8b)。
Overall, these results indicated that the replication of the post-BA.2/5 subvariants are attenuated in human lung epithelial cells when compared to their ancestral BA.2/5, which is in keeping with our in vivo findings.
全体的に、これらの結果は、BA.2/5 以降のサブバリアントの複製が、その祖先の BA.2/5 と比較してヒト肺上皮細胞で弱まっていることを示しており、これは私たちの生体内研究の結果と一致しています。
Our in vivo evaluation demonstrated that the overall replication fitness of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 are largely comparable with that of BA.2 and BA.5.2 in the nasal turbinates of infected mice.
我々の生体内評価では、感染マウスの鼻甲介における BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5 の全体的な複製適応度は、BA.2 および BA.5.2 の複製適応度とほぼ同等であることが実証されました。
To more accurately model the replication fitness of the post-BA.2/5 subvariants in the human nasal epithelium, which may be a key determinant of their transmissibility, we investigated the replication capacity of BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 in differentiated primary human nasal epithelial cells (hNECs).
ヒト鼻上皮における BA.2/5 以降の亜変異体の複製適応度 (その伝染性の重要な決定要因である可能性があります) をより正確にモデル化するために、分化した一次ヒト鼻上皮細胞 (hNEC) における BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5 の複製能力を調査しました。
We found that while BL.1, BQ.1, BQ.1.1, and CH.1.1 did not replicate as efficiently as their ancestral subvariants BA.2/BA.5.2 in the hNECs, the replication of XBB.1 and XBB.1.5 was more robust than that of BA.2 and BA.5.2 (Fig. 8c).
BL.1、BQ.1、BQ.1.1、およびCH.1.1はhNEC内でその祖先亜種BA.2/BA.5.2ほど効率的に複製されなかったが、XBB.1およびXBB.1.5の複製はBA.2およびBA.5.2よりも強力であった(図8c)。

At 72 h post infection (hpi), the virus gene copy in XBB.1- and XBB.1.5-infected hNECs were 5.3- (p = 0.012) and 4.3-folds (p = 0.0044) higher when compared with BA.2, respectively.
感染後 72 時間 (hpi) で、XBB.1 および XBB.1.5 に感染した hNEC のウイルス遺伝子コピーは、BA.2 と比較してそれぞれ 5.3 倍 (p = 0.012) および 4.3 倍 (p = 0.0044) 増加した。
Similarly, the virus gene copy in XBB.1- and XBB.1.5-infected hNECs were 3.7- (p = 0.034) and 3.0-folds (p = 0.023) higher when compared with BA.5.2 at 72 hpi (Fig. 8c).
同様に、XBB.1およびXBB.1.5に感染したhNECのウイルス遺伝子コピーは、72hpiでBA.5.2と比較した場合、それぞれ3.7倍(p = 0.034)および3.0倍(p = 0.023)高かった(図8c)。
Next, we performed immunofluorescence staining on BA.2-, BA.5.2-, BL.1-, BQ.1-, BQ.1.1-, XBB.1-, CH.1.1- and XBB.1.5-infected hNECs at 24 hpi to evaluate antigen expression.
次に、抗原発現を評価するために、感染後24時間でBA.2、BA.5.2、BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5に感染したhNECに免疫蛍光染色を行いました。
In keeping with the virus replication results, while the evaluated Omicron subvariants continue to target ACCTUB + ciliated cells in the hNECs, SARS-CoV-2 N expression was most prominent in XBB.1- and XBB.1.5-infected samples (Fig. 8d).
ウイルス複製の結果と一致して、評価されたOmicronサブバリアントはhNECのACCTUB +繊毛細胞を標的とし続けましたが、SARS-CoV-2 N発現はXBB.1およびXBB.1.5に感染したサンプルで最も顕著でした(図8d)。

Overall, these results revealed that XBB.1 and XBB.1.5 can more robustly replicate in the human nasal epithelium when compared to their ancestral subvariant, which provided biological insights on why XBB is becoming the dominating Omicron sublineage (Fig. 1a).
全体として、これらの結果から、XBB.1およびXBB.1.5は、祖先のサブバリアントと比較して、ヒトの鼻上皮でより堅牢に複製できることがわかり、XBBが支配的なOmicronサブ系統になりつつある理由に関する生物学的洞察が得られました(図1a)。
Discussion
Omicron BA.1 was first detected in South Africa in November 2021, which demonstrated a robust humoral evasion capacity and high transmissibility when compared with the previous SARS-CoV-2 VOCs.
オミクロン BA.1 は、2021 年 11 月に南アフリカで初めて検出され、以前の SARS-CoV-2 VOC と比較して、強力な体液回避能力と高い伝染性を示しました。
Subsequent Omicron subvariants emerged in waves, with a subvariant replacing the previous one every few months.
その後のオミクロンの亜種は波のように出現し、数か月ごとに亜種が以前の亜種に取って代わりました。
We and others have evaluated the virological features, replication, and pathogenicity of these dominant Omicron subvariants, including BA.1, BA.2, and BA.5.
私たちと他の研究者は、BA.1、BA.2、BA.5 など、これらの優勢なオミクロンの亜種のウイルス学的特徴、複製、病原性を評価しました。
More recently, the emerged post-BA.2/5 subvariants were unable to successfully outcompete other subvariants to become the dominant Omicron lineage worldwide, leading to the cocirculation of multiple Omicron subvariants.
最近では、出現したポスト BA.2/5 サブバリアントは、他のサブバリアントに打ち勝って世界中で優勢な Omicron 系統になることができず、複数の Omicron サブバリアントが共存するようになりました。
In this study, we investigated the replication fitness and intrinsic pathogenicity of a number of representative post-BA.2/5 subvariants, including BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5 in mouse models.
この研究では、マウス モデルで、BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、XBB.1.5 など、いくつかの代表的なポスト BA.2/5 サブバリアントの複製適応度と固有の病原性を調べました。
We found that the evaluated post-BA.2/5 subvariants are consistently attenuated in mouse lungs but not in nasal turbinates when compared with their ancestral subvariants BA.2/BA.5.2.
評価された BA.2/5 以降の亜変異体は、その祖先亜変異体 BA.2/BA.5.2 と比較した場合、マウスの肺では一貫して弱毒化されているが、鼻甲介では弱毒化されていないことがわかった。
Further investigations in primary human nasal epithelial cells revealed a gained replication fitness of XBB.1 and XBB.1.5 when compared to BA.2 and BA.5.2.
ヒトの一次鼻上皮細胞をさらに調査したところ、BA.2 および BA.5.2 と比較した場合、XBB.1 および XBB.1.5 の複製適合性が向上していることが明らかになった。
Together, our results indicate a divergent direction of replication fitness and intrinsic pathogenicity of emerging Omicron subvariants in the upper and lower respiratory tract.
これらの結果を合わせると、上気道および下気道における出現する Omicron 亜変異体の複製適合性と固有の病原性の方向性が異なることが示唆される。
The replication and pathogenicity of Omicron subvariants in human studies are potentially biased on vaccination status and previous history of infection/re-infection.
ヒト研究におけるオミクロン亜種の複製と病原性は、ワクチン接種状況や過去の感染/再感染歴によって偏っている可能性があります。
Here, we investigated the replication fitness and intrinsic pathogenicity of post-BA.2/5 subvariants carrying N501Y substitution in spikes in C57BL/6J mice, which allows objective evaluation of these subvariants without the impact of vaccines or previous infections.
ここでは、C57BL/6JマウスのスパイクにN501Y置換を持つポストBA.2/5亜種の複製適応度と固有の病原性を調査しました。これにより、ワクチンや過去の感染の影響なしにこれらの亜種の客観的な評価が可能になります。
The adult C57BL/6J wildtype mice develop a mild to moderate level of disease severity upon Omicron challenge, resembling Omicron pathogenesis in the human population.
成体のC57BL/6J野生型マウスは、オミクロン感染により軽度から中程度の重症度を発症し、ヒト集団におけるオミクロンの病原性に似ています。
In addition, we comprehensively evaluated the infection of these post-BA.2/5 subvariants in weanling, adult, and aged mice to account for the impact of age on the infection outcome, which is a key determinant of SARS-CoV-2 pathogenicity.
さらに、私たちは、SARS-CoV-2 の病原性の重要な決定要因である感染結果に対する年齢の影響を考慮するために、離乳マウス、成体マウス、および老齢マウスにおけるこれらの BA.2/5 以降の亜種について包括的に評価しました。
Overall, our results concluded that the post-BA.2/5 subvariants, BL.1, BQ.1, BQ.1.1, XBB.1, CH.1.1, and XBB.1.5, are attenuated in lungs of infected mice when compared with their ancestral BA.2/BA.5.2 subvariants based on their reduced sgE expression, infectious titers, antigen expression, and lung damage.
全体として、我々の結果は、BA.2/5 以降の亜変異体である BL.1、BQ.1、BQ.1.1、XBB.1、CH.1.1、および XBB.1.5 は、sgE 発現、感染力価、抗原発現、および肺損傷の減少に基づき、その祖先の BA.2/BA.5.2 亜変異体と比較した場合、感染マウスの肺で弱毒化されていると結論付けました。
Our results are in keeping with recent reports showing lower pathogenicity of BQ.1.1 than BA.5 and lower pathogenicity of XBB.1 than BA.2.75 in hamsters.
我々の結果は、ハムスターにおいて BQ.1.1 の病原性が BA.5 より低く、XBB.1 の病原性が BA.2.75 より低いことを示す最近の報告と一致しています。
More importantly, our study revealed a consistent pattern of attenuation of the BA.2 progenies when compared to BA.2, and that of the BA.5 progenies when compared to BA.5.
さらに重要なことは、私たちの研究により、BA.2 と比較した場合の BA.2 子孫の減衰パターンと、BA.5 と比較した場合の BA.5 子孫の減衰パターンが一貫していることが明らかになったことです。
The exact mechanism of the observed attenuation is currently not clear but may in part due to the convergent substitutions (R346, K444, L452, N460, and F486) that modulate immune evasion or ACE2 binding affinity.
観察された減衰の正確なメカニズムは現時点では明らかではありませんが、免疫回避または ACE2 結合親和性を調節する収束置換 (R346、K444、L452、N460、および F486) が一部原因である可能性があります。
In addition, recent studies have also demonstrated the role of non-spike substitutions in the pathogenicity of Omicron.
さらに、最近の研究では、オミクロンの病原性における非スパイク置換の役割も実証されています。
These viral determinants of attenuation in the lungs should be investigated in future studies.
肺での弱毒化のこれらのウイルス決定因子は、今後の研究で調査される必要があります。
While the evaluated post-BA.2/5 subvariants are consistently attenuated in the lungs of infected mice, their replication remained largely comparable to that of BA.2/BA.5.2 in the nasal turbinates of infected mice.
評価された BA.2/5 以降の亜変異体は感染マウスの肺で一貫して弱毒化されているが、その複製は感染マウスの鼻甲介における BA.2/BA.5.2 の複製とほぼ同等のままであった。
In primary human nasal epithelial cells, which is a more physiological-relevant model of the human nasal epithelium, XBB.1 and XBB.1.5 replicated more robustly than BA.2 and BA.5.2.
ヒト鼻上皮のより生理学的に関連のあるモデルである一次ヒト鼻上皮細胞では、XBB.1 と XBB.1.5 は BA.2 と BA.5.2 よりも強力に複製された。
Recently studies reported that Omicron subvariants have a replication advantage over ancestral SARS-CoV-2 or previous variants in human nasal epithelial cells or human nasal organoids.
最近の研究では、オミクロン亜変異体はヒト鼻上皮細胞またはヒト鼻オルガノイドにおいて祖先 SARS-CoV-2 または以前の変異体よりも複製上の利点があることが報告されている。
The increased replication fitness of the XBB subvariants in the human nasal epithelium may play a key role in their transmission among the human populations and in part explain why XBB and its related subvariants are becoming the dominant Omicron subvariants.
ヒトの鼻上皮における XBB 亜種の複製適応度の増加は、ヒト集団間での伝播において重要な役割を果たしている可能性があり、XBB とその関連亜種が優勢な Omicron 亜種になりつつある理由を部分的に説明するものと考えられます。
The divergent replication fitness of Omicron subvariants, including that of XBB.1 and XBB.1.5, in the upper and lower respiratory tract is highly intriguing.
XBB.1 および XBB.1.5 を含む Omicron 亜種の、上気道および下気道における異なる複製適応度は非常に興味深いものです。
The host and viral determinants for such discrepancies warrant further investigations.
このような相違の原因となる宿主およびウイルスの決定要因については、さらに調査する必要があります。
In summary, by evaluating the post-BA.2/5 subvariants in vivo and in human nasal epithelial cells, we found an interesting pattern of divergent trajectory of replication fitness and intrinsic pathogenicity of the post-BA.2/5 subvariants in the upper and lower respiratory tract.
要約すると、生体内およびヒトの鼻上皮細胞でポスト BA.2/5 サブバリアントを評価することで、上部および下部呼吸器におけるポスト BA.2/5 サブバリアントの複製適応度と固有の病原性の異なる軌跡という興味深いパターンが見つかりました。
The attenuated pathogenicity in lungs and increased replication fitness in the nasal epithelium indicated rapid adaptation of the Omicron subvariants in the human populations.
肺における病原性の弱まりと鼻上皮における複製適応度の増加は、オミクロン サブバリアントがヒト集団に急速に適応したことを示しています。
Continued surveillance should be carried out to further monitor the emerging Omicron subvariants.
出現するオミクロン サブバリアントをさらに監視するために、継続的な監視を実施する必要があります。
以下省略。
この記事が気に入ったらサポートをしてみませんか?