
食物繊維によるグルコース代謝の改善はプレボテラの存在量増加と関連する

clinical and translational report|第22巻 第6号 p971-982, 2015/12/01
食物繊維によるグルコース代謝の改善はプレボテラの存在量増加と関連する
ペティア・コヴァチェヴァ・ダッチャリー
アンネ・ニルソン
Rozita Akrami
Eric Martens
Inger Björck
フレドリック・ベックヘド
すべての著者を表示
オープンアーカイブ公開日:2015年11月06日DOI:https://doi.org/10.1016/j.cmet.2015.10.001
PlumX メトリクス
ハイライト
プレボテラ/バクテロイデスは、大麦の穀粒に対する有益な反応と関連している
プレボテラに富む微生物間相互作用は、大麦カーネル反応物質において高い。
プレボテラはバクテロイデスによるグルコース不耐性を防ぐ
プレボテラはマウスの肝グリコーゲン貯蔵量の増加を促進する
まとめ
腸内細菌叢は、宿主の食事と相互作用することでヒトの健康に重要な役割を果たすが、食事に対する反応にはかなりの個人差がある。ここでは、大麦穀粒入りパン(BKB)を3日間摂取した後に糖代謝の改善を示した健康な被験者と、この食事介入に最も反応しなかった被験者の腸内細菌叢組成を比較検討した。プレボテラ/バクテロイデス比は、BKB後の反応者で非反応者より高かった。メタゲノム解析の結果、BKB後、反応者の腸内細菌叢はプレボテラ・コプリに富み、複合多糖類を発酵する能力が高まっていることが示された。最後に、応答者であるヒトドナーの微生物叢を移植した無菌マウスは、非応答者の微生物叢を移植した無菌マウスと比較して、糖代謝の改善、プレボテラの存在量の増加、肝臓グリコーゲン量の増加を示した。このことから、プレボテラはBKBによる糖代謝の改善に関与しており、グリコーゲンの蓄積を促進することで特定の個体で観察される可能性が示唆された。

グラフの概要
図 サムネイルfx1
大きな画像を見るダウンロード 高解像度画像
はじめに
腸内細菌叢は食事と相互作用し、宿主の生理・代謝に影響を与える(Tremaroli and Bäckhed, 2012)。蓄積されたデータにより、肥満(Cotillard et al., 2013, Le Chatelier et al., 2013)、2型糖尿病(Karlsson et al., 2013, Qin et al., 2012)、脳卒中(Wang et al., 2011)の患者において腸内細菌叢が変化することが示され、代謝疾患の発症に関わる重要な環境因子である可能性が指摘されています。腸内細菌叢と代謝性疾患との直接的な関連は、脂肪率が低下し、食事誘発性肥満の発症から保護される無菌(GF)マウスを用いて確立されています(Bäckhedら、2004年、Bäckhedら、2007年)。
腸内微生物の生態は、ヒトでは食事によってかなりの程度調節される(Davidら、2014):脂肪およびタンパク質の高摂取はバクテロイデスのレベルの増加と関連し、一方、繊維の高摂取はプレボテラのレベルの増加と関連している(De Filippoら、2010、Koethら、2013、Wuら、2011)。したがって、食事介入は、腸内細菌叢を調節し、さらに宿主の健康に影響を与える潜在的なツールである(Dewulfら、2013、Kovatcheva-DatcharyおよびArora、2013)。我々は以前に、大麦穀粒ベースの夜食(非デンプン多糖類と耐性デンプンが豊富)が、正常な体格指数(BMI)の健康な被験者の耐糖能を改善することを示し(Johanssonら、2013、Nilssonら、2006、Nilssonら、2008)、結腸発酵とグルコース代謝の間のリンクの証拠を提供した(Johanssonら、2013)。しかし、食事に対する反応には大きな個人差があり(Lampe et al., 2013)、最近の研究では、腸内細菌組成を利用して、食事介入から恩恵を受ける被験者を特定できることが示されている(Korpela et al., 2014, Le Chatelier et al.)
ここでは、腸内細菌の違いが一部の個人におけるBKBの有益な効果を説明できるかどうかを調べるために、大麦核を用いたパン(BKB)後にグルコース代謝の改善を示した健康な被験者とこの食事介入に最も反応しなかった被験者の腸内細菌叢の組成を比較しました。
結果および考察
BKB介入は健康なヒトのサブセットでグルコース代謝を改善する
我々は以前、BKBの摂取が食後グルコース代謝の改善と関連することを示した(Nilsson et al.、2006)。本研究では、BKBに対する反応に応じて2つのグループを選択した。まず、39人の健常者を対象に、クロスオーバー試験でBKBまたは白小麦粉パン(WWB)を3日間連続して食事介入し、食後のグルコースおよびインスリン応答を測定した(これらの被験者の生体測定値を表S1に示す)。予想通り、標準化された朝食後の4日目に測定された平均血糖値および血清インスリン反応は、全グループでWWBと比較してBKB後に改善された(図S1AおよびS1B)。また、朝食時に測定した呼気中水素排泄量は、BKB摂取後、WWBと比較して高くなった(図S1C)。全群から、BKBに続く標準化朝食後に、グルコースおよびインスリン反応の改善が最も少ないか全く見られない被験者(非応答者と呼ぶ;図1Aおよび1B )および最も顕著な改善を示した被験者(応答者と呼ぶ;図1Cおよび1D)をWWBと比較して10名選択した。選択基準に関するさらなる詳細は、「実験手順」に含まれている。空腹時呼気水素は、BKBの消費後に両群で増加し(図1E)、難消化性炭水化物の存在が、反応者と非反応者の両方において結腸微生物叢の発酵活性を増加させることを示した。
図サムネイルgr1
図1標準化された朝食後の非応答者と応答者のグルコースおよびインスリンプロファイル(WWBまたはBKBの3日間の摂取後
キャプションを表示
大きな画像を表示するダウンロード 高解像度画像
腸内細菌の16S rRNA配列決定により、BKB摂取後の反応者にプレボテラが増加することが示された。
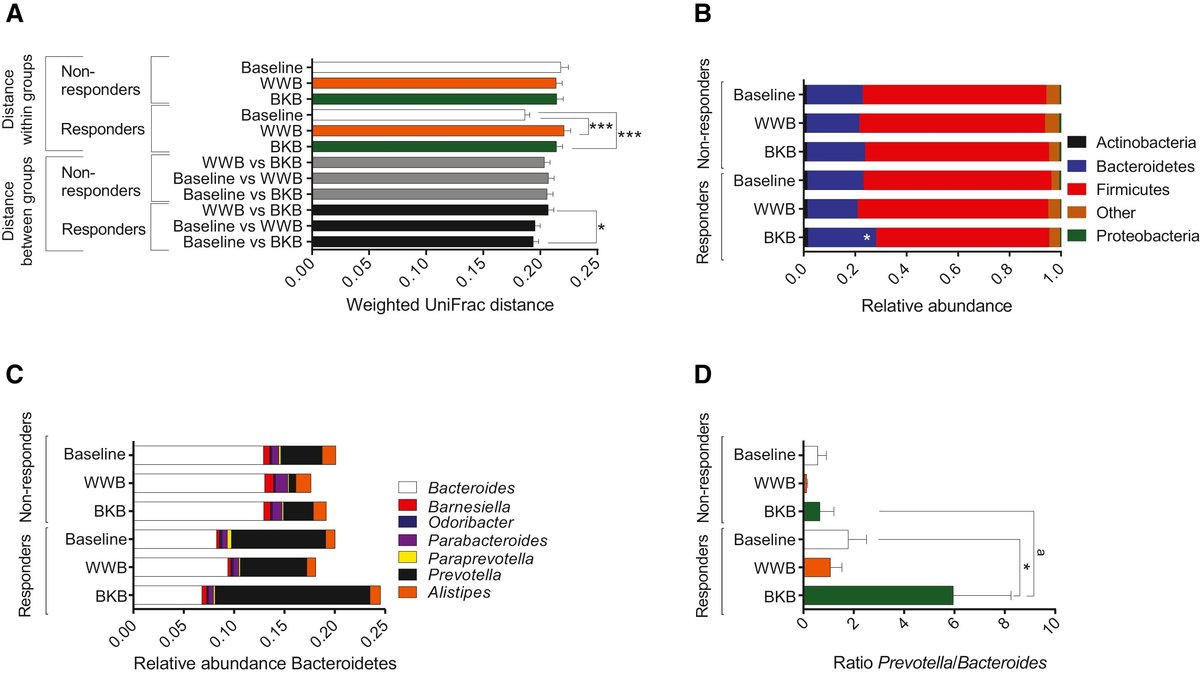
反応者と非反応者の腸内細菌叢組成に違いがあるかどうかを調べるため、これら2つのグループの糞便サンプルを16S rDNA遺伝子の454/Rocheパイロシークエンスによって解析した。個体の非重み付け(存在/不在情報)UniFrac距離(Lozupone et al., 2006)に基づく糞便マイクロバイオータ配列データの主座標分析(PCoA)は、主に被験者によってクラスタリングされ(図S2A)、以前の研究(Wu et al., 2011)と一致して、各個人の微生物群集が異なる微生物群の集合体からなることが示唆された。さらに、WWB、BKB、およびベースライン時の反応者と非反応者間の非重み付けおよび重み付けされたUniFrac距離に基づいて微生物群集を評価しました。我々は、食事にWWBまたはBKBを補充することで、反応者の微生物群に影響を与えるが、非反応者では影響を与えないことを示した(図2AおよびS2B)。グループ間のUniFrac距離の重み付けにより、WWB摂取と比較したBKB摂取は、反応者では糞便微生物叢の特定のメンバーの相対存在量に有意な変化をもたらしたが、非反応者ではそうではなかったことが明らかになった(図2A)。

図のサムネイル gr2
図2WWBまたはBKBの3日間摂取による非応答者と応答者の腸内細菌叢の変化
キャプションを表示
大きな画像を見るダウンロード 高解像度画像
この知見にさらに取り組むため、次に、ベースラインとBKBおよびWWB摂取後の反応者と非反応者の腸内細菌叢組成を分析した。固形細菌とバクテロイデーテスは、反応者と非反応者の両方で最も一般的な門であり、反応者と非反応者の間で、ベースラインとWWB摂取後の門レベルでの存在量の違いは観察されなかった(図2B)。しかし、BKB摂取後、Bacteroidetesの存在量は、反応者で増加し、非反応者では増加しなかった(図2B)。BacteroidetesはBacteroidesとPrevotellaの2つの属からなり、反応者のBKB介入後のBacteroidetesの増加はPrevotellaのブルームによるものであった(図2C)。非応答者では、BKB介入後もPrevotellaは増加しなかった(図2C)。プレボテラのレベルが高い被験者は、通常、バクテロイデスのレベルが低い(Arumugam et al.、2011、Koren et al.、2013、Wu et al.、2011)。また、反応者ではベースラインに対してBKB介入後にプレボテラ/バクテロイデス比が増加し、反応者では非反応者に対してBKB後にプレボテラ/バクテロイデス比が高くなることが観察された(図2D)。食物繊維を豊富に含む食事の長期摂取はプレボテラの存在量の増加と関連しているため、プレボテラ/バクテロイデス比の増加は、反応者が習慣的に食物繊維を多く摂取していることの結果であると考えられる(David et al.、2014、 De Filippo et al.、2010、Wu et al.、2011)。しかし、反応者と非反応者の両方が、WWB介入に対して同様のグルコース反応を示した(図1Aおよび図1C)。
次に、反応者と非反応者の間で観察されたBacteroidesとPrevotellaの存在量の違いが、他の微生物叢のメンバーを含むより広範囲なシフトを伴うかどうかを調査した。ベースライン時、反応者と非反応者の腸内細菌叢は、Doreaの相対量が増加し(p < 0.05, Kruskal-Wallis test with a false discovery rate [FDR] correction)、Roseburiaが増加する傾向にあった(p < 0.1, Kruskal-Wallis test with a FDR correction; Figure S2C);DoreaとRoseburiaはどちらも、食事性多糖の代謝が知られている堅果類の属であった(Duncan et al, 2007). しかし、これらの属の存在量はBKB添加後に減少したことから(図S2C)、これらの属は応答者における難消化性糖質の分解に関与する主要な分類群ではないことが示された。メタン生成古細菌は、H2や他の発酵最終生成物の蓄積を防ぐことによって腸内の多糖類発酵の効率を向上させるため、腸内細菌叢の有益な効果に寄与する可能性もある(Pimentel et al.、2012)。そこで、メチルコエンザイムM還元酵素遺伝子を定量することにより、WWBおよびBKBの摂取前後の反応者と非反応者におけるメタン生成アーキアの存在量も評価しました。その結果、BKB摂取後のメタン生成菌の存在量は、非応答者に比べて応答者で高いことがわかった(図S2D)。これは、これらの個体における多糖類発酵の効率上昇に寄与していると考えられる(Samuel and Gordon、2006、Zhang et al.、2009)。
我々は、反応者と非反応者で優占する細菌系統が異なるため、微生物叢によって複合糖質が短鎖脂肪酸(SCFA)などの代謝物に発酵される方法が異なる可能性があると仮定している。しかし、反応者と非反応者の間で糞便SCFAプロファイルに有意差は観察されず(図S2E)、これは以前の食事介入研究(Wuら、2014)で示唆された腸内細菌叢組成の個人間変動と関連している可能性がある。
応答者と非応答者の表現型は1年間安定したままである
これらの被験者における微生物叢の経時的安定性を調べるために、最初の研究の12カ月後に、元の被験者20人のうち14人(反応者7人と非反応者7人)でBKBとWWBによるクロスオーバー研究を繰り返した。ここでも、当初反応者と定義された被験者ではBKB介入後に食後血糖値の改善が観察されたが、当初非反応者と定義された被験者では見られなかった(図S3AおよびS3B)。さらに、BKB後のこの時点では、反応者対非反応者でプレボテラ/バクテロイデス比(16S qPCRで測定)が増加する傾向があり(図S3C)、微生物叢が経時的に比較的安定していることが示された。
腸内細菌叢のショットガン・メタゲノム配列決定により、反応者ではプレボテラ・コプリが増加し、BKBに反応してメタゲノムが機能的に変化していることが示された。
非応答者と応答者の微生物叢の違いを検証するために、ショットガンメタゲノムシーケンスによる糞便微生物叢の解析も行いました。被験者1人あたり平均1040万(±0.89 SD)のペアエンドリードを生成した(表S2)。454シーケンサーのデータと一致するように、反応者のメタゲノムでは非反応者に比べてPrevotellaceaeが濃縮される傾向があり、反応者ではベースラインと比較してBKB後にBacteroidaceaeの豊度が減少したが非反応者では見られなかった(図3A; 表S2)。プレボテラ/バクテロイデス比は、反応者ではベースラインと比較してBKB後に高く、反応者対非反応者ではBKB後に高かった(図3B)。また、反応者ではPrevotella copriがPrevotellaceaeの中で最も多く存在することが示された(図3C;表S2)。

図 サムネイル gr3
図3ベースライン時とBKB摂取後の非応答者と応答者のメタゲノムにおける分類学的組成
キャプションを表示
大きな画像を見るダウンロード 高解像度画像
さらに、BKB摂取後に非応答者と応答者のメタゲノムに誘発される機能的変化をベースラインと比較して検討し、糖質消化の主要酵素であるグリコシドヒドロラーゼをコードする遺伝子の存在量を定量した(Dodd et al.、2011)。BKB投与後、キシラン1,4-β-キシロシダーゼ、グルカンエンド-1,3-β-D-グルコシダーゼ、グルカン1,6-α-グルコシダーゼ、リチェニナーゼおよびセルラーゼをコードする遺伝子の存在量が反応者で著しく増加したが非反応者では観察されなかった(Table S2)。これらの酵素は、BKBにおけるβグルカンやその他の複合多糖類の消化に不可欠である。したがって、これらの知見は、BKBに存在する食物繊維が、非応答者よりも応答者のメタゲノムによってより効率的に消化できることを示唆し、プレボテラの機能レパートリーを、食事からの複合多糖を発酵する腸内細菌叢の能力向上と関連付けた最近のメタゲノム解析と一致する(Rampelliら、2015年)。
BKB後の応答者におけるネットワークの複雑性の増大とプレボテラに富む微生物相互作用
特定の微生物の共起がBKBに対する反応の違いを説明できる可能性があるかどうかを調べるために、相関ネットワーク解析を用いて、ベースラインとBKB後の非応答者と応答者の微生物叢において、BacteroidesとPrevotella種が他の種とどのように相互作用するかを調査しました。その結果、ベースライン時とBKB後において、ノード(種)とエッジ(種間の相互作用[相関])の数は、非応答者の方が応答者より多かった(表S3)。しかし、種間の正の相互作用の数および相互作用の複雑さ(平均近傍数およびネットワーク密度[平均近傍数の正規化]で示される)は、BKB後に非応答者と比較して応答者で増加した(表S3)。
ネットワーク密度がどのように分布しているかを知ることができるネットワーク中心化パラメータを用いると、BKB後の反応者において相互作用密度が最も中心化されていることが観察された(表S3;図S4)。中心性は、非応答者のバクテロイデス属から応答者のプレボテラ属にシフトしており、ハブノード(最も相互作用の多いノード)は、非応答者ではバクテロイデス属、応答者ではプレボテラ属であった(図S4)。また、ネットワーク解析の結果、ベースライン時の応答者ではPrevotella属菌が非常に多く、非応答者ではBacteroides属菌が優占していた(表S3;図S4)。反応者のメタゲノムで高濃度であったP. copriは(図3C)、ベースラインではすべての個体に存在したが、反応者では2倍以上高く、BKB後には反応者のみで(存在量と他の種との相互作用の数において)増加した(表S3;図S4)。
食餌に応じた微生物種の存在量の変化は、その微生物種と群集の他のメンバーとの相互作用だけでなく、撹乱に応じたそれらの相互作用の安定性と、その後の集団の対処法にも依存する(Harmon et al.) ネットワーク解析の結果、プレボテラとバクテロイデスは、ベースライン時の応答者と非応答者の両方で、放線菌、ファーミキューテス、プロテオバクテリア門の種と競合(すなわち負の相関)していた(表 S3;図 S4)。ActinobacteriaとFirmicutesのメンバーには、複合糖質の主要な分解者である種が含まれることが示唆されており(Flint et al.2012、El Kaoutari et al.2013、Leitch et al.2007、Ze et al.2012)、プレボテラとバクテロイデスがこれらの種と同じ基質をめぐって競合する可能性が示唆された。しかし、回答者のBKB後では、プレボテラ属は他のプレボテラ属だけでなく、放線菌、ファーミキューテス、プロテオバクテリア、アーキアの多くの種と共起しており(表S3;図S4)、ニッチが拡大したことが示唆された。このことから、プレボテラ属細菌は他の生物群集と積極的に相互作用し、反応者の炭水化物発酵の促進を図っていると推測された。一方、非応答者のBKB後では、ネットワークはBacteroides属が優勢なままであり、微生物間の負の相互作用の数が増加した(表S3)。これは、非応答者ではより多くの種が同じ基質をめぐって競合していることを示唆している。
プレボテラはマウスの糖代謝を改善する
プレボテラ属細菌による糖代謝改善の可能性を探るため、マウスにP. copriまたは対照菌を毎日7日間摂取させ、耐糖能について比較検討した。低脂肪・高タンパク・高繊維の標準的なチャウ食を与えたマウスでは、ヒト糞便由来の生きたP. copri(DSM 18205)を投与すると、加熱殺菌したP. copriと比較して耐糖能が改善した(2種類のマウスで実験、図4Aおよび4B)。一方、高脂肪・低食物繊維食を与えたマウスでは、P. copriは耐糖能に影響を与えなかった(図4C)。これらの結果は、プレボテラが腸内に定着し、糖代謝に有益な効果を促進するためには、食物繊維が重要であるという概念を支持するものである。
図サムネイルgr4
図4プレボテラのマウス糖代謝に対する有益な効果は食餌依存的であり、P. copriはB. thetaiotaomicron誘発性糖質不耐症から保護する。
キャプションを表示
大きな画像を見るダウンロード 高解像度画像
Bacteroides属とPrevotella属が糖代謝に及ぼす影響を直接的に調べるため、ヒト糞便由来のBacteroides thetaiotaomicron(ATCC 29148)とP. copri(DSM 18205)を14日間GFマウスに単コロン化および双コロン化した。すべてのマウスに標準的なチャウ食を与えた。B. thetaiotaomicronは、広範囲の動植物性グルカンを利用する一方で (Martens et al., 2008, Martens et al., 2011, Xu et al., 2003) 、他のBacteroides株と比較して、複合多糖類の分解に必須の遺伝子を欠いている (Dodd et al., 2010, Martens et al., 2011)。一方、P. copriは複雑な多糖の発酵と利用に不可欠な多くの酵素と遺伝子群を有している(Dodd et al.) 両菌はグノトビオティックマウスに同程度にコロニー形成し、バイコロニー形成では両菌のレベルが低下した(図4D)。B. thetaiotaomicronをコロニー形成したマウスでは、P. copriをコロニー形成したマウスと比較して耐糖能が損なわれていた(図4E)。重要なことは、B. thetaiotaomicronによる耐糖能の低下は、P. copriとの同時コロニー形成によって防止されたことである(図4E)。B. thetaiotaomicronのレベルは、モノコロニー化(R2 = 0.11; p = 0.22)またはバイコロニー化(R2 = 0.10; p = 0.42)のいずれのマウスでも、糖負荷試験の曲線下面積と相関がなく、バイコロニー化マウスにおけるB. thetaiotaomicronレベルの低下が、糖負荷改善の理由ではないことが示された。これらのデータは、P. copriとB. thetaiotaomicronがマウスの腸内で糖質発酵を行う能力の違いが、糖代謝の面で観察された違いに寄与していることを示唆し、耐糖能改善におけるP. copriの重要性を強調するものであった。
Bacteroidesはプロピオン酸を生産することが知られているが、Prevotellaはコハク酸を生産する(Macy et al.、1978)。予想通り、B. thetaiotaomicronを単独培養したマウスの盲腸ではプロピオン酸が主要代謝物であったが、P. copriを単独培養したマウスでは、盲腸のコハク酸レベルが高く、プロピオン酸の生産が見られなかった(図S5A)。プレボテラの有益な効果がコハク酸の高生産によって媒介されているかどうかを調べるために、コハク酸をプロピオン酸に変換できないB. thetaiotaomicron変異体、BT1686-89を作製した。この菌株をGFマウスにコロニー形成させると、糞便中のコハク酸レベルが高くなったが(図S5A)、野生型B. thetaiotaomicronをコロニー形成したマウスと比較して、グルコース応答は改善されなかった(図S5B)。したがって、P. copriを単独でコロニー形成したマウスで観察された高レベルのコハク酸は、この細菌種の有益な効果を説明するには十分でない。
耐糖能異常は、近位結腸および肝臓における遺伝子発現の変化と関連する
B. thetaiotaomicronとP.copriをコロニー形成したマウス間の耐糖能の違いを説明しうる遺伝子を同定するために、モノおよびバイコロニー形成マウスとGFマウスの近位結腸および肝臓生検のRNA配列決定(RNA-seq)を実施した。1サンプルあたり平均1400万リードを生成し、これらのリードの90%以上がマウスゲノムにマッピングされた(Table S4)。B. thetaiotaomicronのコロニー形成により有意に発現量が増加または減少し、P. copriのコロニー形成またはB. thetaiotaomicronとP. copriのバイコロニー形成(いずれもGFマウスと比較)により有意に変化しない遺伝子に着目し、近位結腸で601、肝臓で206の遺伝子を同定した(Table S4)。
近位結腸で最も有意に影響を受けた遺伝子は、グルコースおよびエネルギー恒常性に有益な効果を誘導する腸の糖新生の主要酵素であるグルコース-6-ホスファターゼをコードするG6pcであり(De Vadderら、2014);それはB. thetaiotaomicronのコロニー化によってダウンレギュレートされていた(表1)。肝臓のグルコース代謝に影響を与える遺伝子として、インスリンシグナル伝達の重要なタンパク質であるsorbin and SH3 domain containing 1をコードするSorbs1 (Lesniewski et al., 2007, Menzaghi et al.)と、肝臓のグルコース代謝に影響を与えるPyglの2つが最も有意に影響を受けた。Bollen et al., 1998, Izumida et al., 2013)、肝臓グリコーゲンホスホリラーゼ(グリコーゲンからグルコースへの分解を触媒する酵素)をコードするPyglが、B. thetaiotaomicronのコロニー化により上昇した(表1)。次に、Pyglの発現量の変化が肝臓のグリコーゲン貯蔵量の変化に反映されるかどうかを調べたところ、B. thetaiotaomicronを単コロニー化したマウスは、P. copriを単コロニー化したマウスと比較して肝臓のグリコーゲン量が著しく減少した(0.60 ± 0.14 対 2.25 ± 0.21 mg/g肝臓、p < 0.0001)ことが明らかになった。
ヒト腸内細菌叢の移植によるBKBの宿主糖代謝への有益な効果の伝達
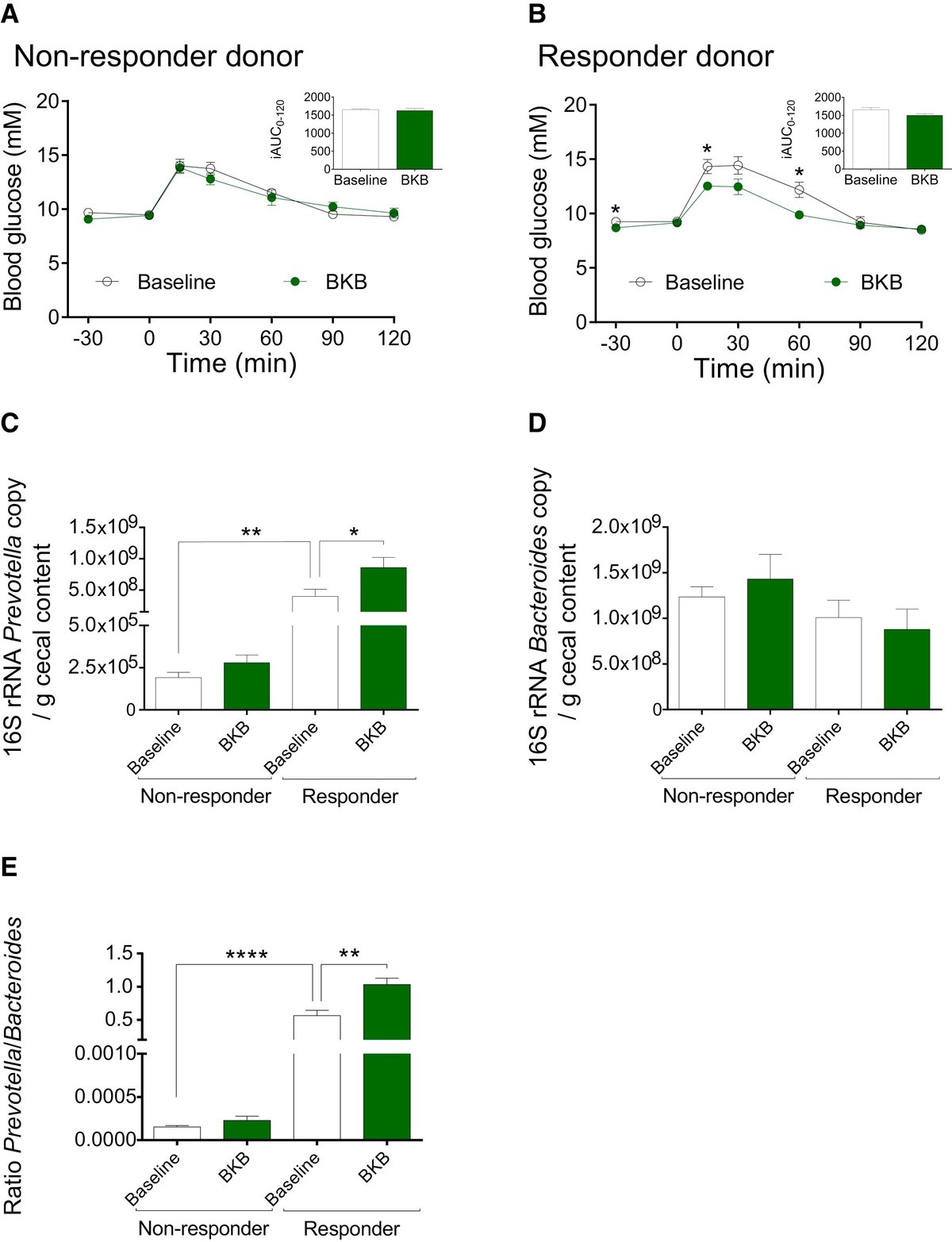
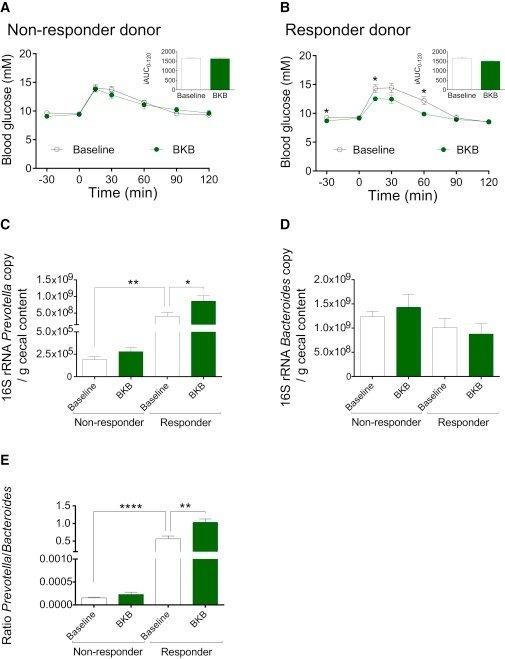
BKB後の宿主の糖代謝改善を促進する非応答者と応答者の関連微生物叢の可能性を調べるため、ベースラインとBKB後の非応答者2人と応答者2人のヒトドナーの微生物叢をGFマウスに移した。BKBおよびWWB後の食後血糖および血清インスリン反応、ならびに選択した非レスポンダーおよびレスポンダー・ドナーのベースライン時およびWWBおよびBKB後のプレボテラ/バクテロイデス比を図S6に示す。ベースラインまたはBKB後に採取した非レスポンダードナーのいずれかからの微生物叢で14日間コロニー化したマウスの食後血糖反応にいかなる差異も観察されなかった(図5AおよびS7A)。対照的に、ベースラインで採取した同じドナーからの微生物叢でコロニー形成したマウスと比較して、BKB後にレスポンダー・ドナーからの微生物叢で14日間コロニー形成したマウスにおいて改善されたグルコース応答が観察された(図5BおよびS7B)。
図サムネイルgr5
図5BKB後に採取したレスポンダーの腸内細菌叢をGFマウスに移植すると耐糖能が改善する。
キャプションを表示
大きな画像を見るダウンロード 高解像度画像
また、16S qPCRを用いて、ベースラインとBKB後に非レスポンダーとレスポンダーの微生物叢をコロニー形成したマウスの盲腸におけるプレボテラとバクテロイデスのレベルを測定した。プレボテラのレベルは、ベースラインで採取したレスポンダー対非レスポンダーのドナー微生物叢でコロニー形成したマウスで高く、BKB後に採取したレスポンダー微生物叢でコロニー形成したマウスでさらに増加した(図5C及びS7C)。バクテロイデスのレベルは、非レスポンダー対レスポンダーのドナー微生物叢でコロニー形成されたマウスの盲腸でわずかに、しかし有意に高く、ベースラインまたはBKB後に採取した微生物叢でコロニー形成されたマウスで同様であった(図5DおよびS7D)。さらに、プレボテラ/バクテロイデス比は、ベースラインで採取したレスポンダー対非レスポンダーのドナー微生物叢でコロニー形成したマウスで有意に高く、BKB後に採取したレスポンダー微生物叢を受け取ったマウスでさらに増加した(図5EおよびS7E)。これらの観察は、BKBによって誘発された微生物叢の変化が、宿主における有益な応答を促進するのに十分であり、特に、BKB後のプレボテラの存在量の増加が耐糖能を改善するという我々の仮説をさらに支持するものである。
また、B. thetaiotaomicronとP.copriでコロニー形成されたマウスで制御が異なる遺伝子、およびグルコース代謝に関与する可能性のある遺伝子(表1)の相対発現を、レスポンダーと非レスポンダーからの微生物叢をベースラインとBKB後に受けたマウスの組織で測定した。BKB後に採取したレスポンダーと非レスポンダーの微生物叢でコロニー化したマウスの近位結腸生検において、G6Pcの発現が有意に増加していることを確認した(図6A)。また、BKB後に採取したレスポンダーおよびノンレスポンダーの微生物叢でコロニー形成されたマウスの肝臓では、ベースラインで採取したそれぞれの微生物叢と比較して、Sorbs1の有意な減少が観察された(図6B)。しかし、Pyglは、BKB後に採取したレスポンダーではなくノンレスポンダーの微生物叢でコロニー形成されたマウスの肝臓で、ベースラインで採取した微生物叢と比較して有意に減少した(図6C)。
表1B. thetaiotaomicronを単独コロニー形成したマウスの結腸近位部および肝臓の生検で有意に制御された上位10遺伝子
組織 遺伝子名 Log2 Fold change p値 p-adj 説明
大腸近位部 G6pc -2.75 1E-10 5.4E-08 グルコース-6-ホスファターゼ、触媒作用
Apoa4 -3.51 4E-08 8.1E-06 アポリポタンパク質A-IV
Sprr2a3 1.15 5E-08 1E-05 小型プロリンリッチタンパク質2A3
Saa2 -1.09 8E-08 1.6E-05 血清アミロイド A 2
Pepd -0.57 8E-08 1.6E-05 ペプチダーゼ D
Enpep -1.01 2E-07 2.9E-05 グルタミルアミノペプチダーゼ
Gsdmcl2 2.41 3E-07 5E-05 ガスデルミンC様2型
Nfkbia -0.39 4E-07 6.1E-05 B細胞におけるκライトポリペプチド遺伝子エンハンサー核因子阻害剤
Slc38a2 0.36 1E-06 0.00015 溶質キャリアーファミリー38のメンバー2
Mfsd8 -0.45 1E-06 0.00015 メジャーファシリテータースーパーファミリードメイン含有8番染色体
Edn1 -0.82 2E-06 0.00025 エンドセリン
肝臓 Sorbs1 0.60 5E-06 0.00033 ソルビン・SH3ドメイン含有1型
Bdh2 0.97 1E-05 0.00062 3-ヒドロキシ酪酸デヒドロゲナーゼ・2型
Chka -0.81 1E-05 0.00068 コリンキナーゼα
Camk2b 1.57 1E-05 0.00073 カルシウム/カルモジュリン依存性プロテインキナーゼII, β
Cd9 -0.69 2E-05 0.00102 CD9抗原
Lrig2 -0.60 3E-05 0.00138 ロイシンリッチリピートおよび免疫グロブリン様ドメイン2
Hist1h1e -0.85 5E-05 0.00188 ヒストンクラスター1, H1e
Ergic1 0.41 6E-05 0.00223 小胞体-ゴルジ体中間コンパートメント(ERGIC) 1
Pygl 0.41 7E-05 0.00244 肝臓グリコーゲンホスホリラーゼ
Atp6v0e2 1.10 8E-05 0.00268 ATPase, H+ transporting, lysosomal V0 subunit E2
表S4に関連する。
表を新規タブで開く

図のサムネイルgr6
図6BKB後に採取した応答者の腸内細菌叢をGFマウスに移植すると、肝グルコースのグリコーゲンとしての貯蔵が促進される。
キャプションを表示
大きな画像を見るダウンロード 高解像度画像
次に、Pyglの発現量の変化が、レスポンダーと非レスポンダーの微生物叢を持つマウスの肝臓のグリコーゲン蓄積量の変化に反映されているかどうかを調べた。その結果、BKB後に採取したレスポンダー微生物叢を保有するマウスの肝臓のグリコーゲン量は、ベースラインで採取したレスポンダー微生物叢を保有するマウス、BKB後またはベースラインで採取した非レスポンダー微生物叢を保有するマウスと比較して有意に高いことが示された(図6D)。これらの知見は、プレボテラが、グリコーゲン貯蔵量の増加を促進することによって、一部の個体(すべてではない)で観察されたBKBによる糖代謝の改善に寄与している可能性を示唆するものであった。
展望
要約すると、我々は、健康なヒトのサブグループにおいて、腸内細菌叢の変化が、難消化性糖質の短期間の食事補充によって誘導される有益な代謝上の利点に寄与していることを示した。また、本研究は、腸内細菌叢の個々のメンバー(存在量および他の微生物種との相互作用の両方)が宿主の代謝および食事の変化に対する反応に潜在的に重要であることを強調している。この結果は、宿主の代謝を改善するための個別化アプローチの重要性を支持し、代謝異常の治療に食事と微生物叢の両方を含むシンバイオティクス・アプローチを用いる可能性を強調するものである。しかし、この知見は、より大規模なコホートにおいて検証される必要があることに留意することが重要である。さらなるステップとして、食事介入の恩恵を受ける個人を特定する戦略として腸内細菌叢組成を使用する前向き追跡調査を実施する予定である。
実験手順
被験者と試験デザイン
合計39人の被験者(男性6人、女性33人)がこの研究に参加した(表S1)。参加基準は、年齢50〜70歳、BMI普通〜やや過体重(BMI18〜28kg/m2)、空腹時血糖値6.1mmol/l以下、非喫煙者、全身健康、代謝異常や食物アレルギーが知られていないことであった。降圧剤と、抗炎症作用のない処方箋不要の鎮痛剤は受け入れた。本研究の参加者には、パートナーや配偶者の参加者はいなかった。本研究は、スウェーデン・ルンド市の地域倫理審査委員会により承認された(文献2010/457)。
我々は、BKB対WWBのグルコース代謝への影響を検証するために、無作為クロスオーバーデザインを使用した。BKB及びWWBの構成及び実験手順は、先に記載した(Nilssonら、2015)。簡単に言えば、各被験者は、最低2週間の間隔を空けて、BKBまたはWWBのいずれかを無作為の順序で摂取する2回の3日間介入に参加した。介入期間3日目の夜食後、被験者は翌朝に実験部門で提供される標準化された朝食まで絶食した(試験日)。8時の朝食前に空腹時採血を行い、呼気水素を測定した。血糖値測定のため、指刺し毛細血管採血を行った(HemoCue B-glucose)。血清インスリンを測定するために静脈血を採取した。血清は遠心分離により分離し、分析まで直ちに冷凍庫(-40℃)で保存した。呼気中の呼気水素は、Gastro+(Bedfont EC60 Gastrolyzer)を用いて測定した。このうち20名の被験者におけるグルコースおよびインスリン反応と呼気水素排泄量に関する結果は、以前に報告されている(Nilsson et al.、2015)。
糞便サンプルは、食事介入前(ベースラインと定義)、およびBKBまたはWWB後4日目(試験日)に発生した最初の排便から採取された。サンプルは直ちに凍結され、24時間以内に実験部門に引き渡され、分析まで-80℃で継続保管された。
反応者、非反応者の選択
全群から、BKB後の標準化朝食後に、WWBと比較して、(1)血糖値の増分面積(IAUC、0-90分)が25%以上減少、(2)総AUC(0-90分)が減少、(3)インスリンIAUC(0-90分)が15%以上減少、の全ての反応が見られた10名の応答者(男2、女8)のグループを選択した。残りの29名から、BKB後の標準化朝食後のグルコースおよび/またはインスリン反応の改善がWWBと比較して最も少なかった10名の無反応者(すべて女性)を選出した。このうち、BKBによってグルコース反応もインスリン反応も変化しなかったのは4名、グルコース反応は変化したがインスリン反応はわずかに変化しなかったのは3名、インスリン反応は変化したがグルコース反応はわずかに変化しなかったのは3名であった。
ゲノムDNAの精製、増幅、パイロシークエンス、およびデータ解析
ビーズ叩きを繰り返して糞便からゲノムDNAを分離した後(Salonenら、2010)、16 rRNA遺伝子のV1-V2可変領域を、既報(De Vadderら、2014)のように27Fおよび338Rプライマーを使用して増幅した。プールした産物は、National Genomics Infrastructure (Stockholm)で454 GS FLX SystemとTitanium Chemistry (Roche)を用いて塩基配列を決定した。
生データは、200塩基より短い配列、1,000塩基より長い配列、プライマーのミスマッチ、あいまいな塩基、修正不可能なバーコード、6塩基を超えるホモポリマーランを含む配列を取り除くために品質フィルターにかけられた。品質フィルターされたリードは、454アダプターおよびバーコード配列からトリミングされ、ソフトウェアパッケージQuantitative Insights into Microbial Ecology (QIIME) (version 1.5.0) で解析された。合計で755,963リードがquality filterを通過した(平均12,599配列/サンプル)。配列データはQIIMEのdenoise_wrapper.pyでデノイズした。
配列は、ペアワイズ同一性の97%の閾値でUCLUSTを使用してオペレーション分類学的ユニット(OTU)に割り当てられた。最も豊富な配列を各OTUの代表として選び、Ribosomal Database Project Classifierを用いて分類学的に割り当てた。代表的なOTUはPynastを用いて整列させ、FastTreeで系統樹を作成し、これを用いてサンプルのβ多様性を推定した(加重および非加重UniFrac)。
ショットガンメタゲノム解析
メタゲノムDNAサンプルは、ヨーテボリ大学のコア施設でNextSeq500を用いてペアエンドモード(2∗151 bp)で配列決定されました。各シングルリード配列は、Metagenome rapid annotation (MG-RAST) サーバー (Meyer et al., 2008) を用いて、機能および分類学的プロファイルの両方について個別にアノテーションされました。解析の詳細については、Supplemental Informationに記載されています。
ネットワーク共起解析
ネットワーク解析のために、各グループのベースラインとBKB後の反応者と非反応者において、微生物叢におけるBacteroidesとPrevotella種の存在量と他の種の間のすべての可能なスピアマンの相関を別々に計算した(メタゲノム解析のデータを使用した)。ネットワークの複雑さを軽減するために、4個体以上(各群10個体中)で最低5つの配列を持つ種のみを使用した。すべての統計解析はR(R Core Team, 2015)で行い、p値はFDR法(Benjamini and Hochberg, 1995)を用いて補正した。許容可能な共起性相互作用は、絶対相関係数>0.6および補正p値<0.05を有するとみなされた。ネットワークは、Prefuse Force Directed Layoutを用いたCytoscape 3.0.2 (Kofia et al., 2015)を用いて可視化された。CytoscapeのNetworkAnalyzerツールは、ネットワークを個別に分析するために使用された。
16S rRNA定量PCR
16S rRNA定量PCRは、CFX96 Real-Time System (Bio-Rad) を用いて行った。すべての反応は、1回の実行で2重にPCRを行った。qPCRは、以前に報告されたように行った:Bacteroidesについて (Walker et al., 2011), methyl coenzyme-M reductase遺伝子について (Denman et al., 2007), Prevotellaについて (Matsuki et al., 2002), B. thetaiotaomicronについて (Samuel and Gordon, 2006)。解析の詳細については、Supplemental Informationに記載されている。
P. copriを用いたマウス実験
我々は、P. copriの効果を調べるために、マウスで3つの実験を行った。最初の実験では、チャウ食を与えたSwiss WebsterおよびC57BL/6雄成熟マウス(Taconic)に、生きたまたは加熱殺傷した対照のP. copriを毎日7日間連続して投与した(1群5匹ずつ)。第二の実験では、C57BL/6雄性マウスに高脂肪食(脂肪由来40.6% kcal; TD.09683 Harlan Teklad)を4週間与え、生きたP. copriまたは媒体対照を7日間連続してギャバリングさせた。第3の実験では、10〜12週齢のGF Swiss Webster雄マウスに、108CFUのP. copri株DSM18205および/またはB. thetaiotaomicron株ATCC 29148または同系のB. thetaiotaomicron変異株(BT1686-89)を単回ギャバージして接種した。モノおよびバイコロニー化したマウスをアイソケージシステムで14日間飼育した。これらすべての実験の最後に、経口ブドウ糖負荷試験(OGTT)を実施した。すべてのマウス実験は、University of Gothenburg Animal Studies Committeeによって承認されたプロトコルを用いて実施された。
ヒトの糞便のマウスへの移植
ベースラインおよびBKB摂取後の選択された反応者および非反応者からの糞便サンプル(図S6参照)を、還元溶液(Na2SおよびシステインをNaHCO3緩衝液に溶解)を添加したPBS緩衝液に加え、10〜12週のGF Swiss Webster雄および雌マウス(1群5〜7)に経口ガベージで移した。マウスは14日間コロニー形成された。実験終了時にOGTTを実施した。肝臓、盲腸、および結腸を採取し、直ちに液体窒素中でスナップ冷凍し、さらなる処理まで-80℃で保存した。
RNA 抽出および RNA 配列決定
肝臓および近位結腸のマウス組織をTissueLyzer(QIAGEN)を用いてRLTバッファー中でホモジナイズした。RNAはQiacube (QIAGEN)を用いて、製造元のプロトコルにしたがって単離した。RNAの量と質はNanoDrop ND-1000 spectrophotometer (NanoDrop Technologies)を用いて調べた。
RNA サンプルは、ヨーテボリ大学のゲノムコア施設において、Nextseq500 (2∗75) を用いてペアエンドモードで配列決定された。配列(リード)の質は、Trimmometricソフトウェア(Bolgerら、2014)のMAXINFOアルゴリズムを用いて評価した。より詳細な情報は、Supplemental Informationに含まれています。
RT qPCR
cDNAは、High-Capacity cDNA Reverse Transcription Kit(Applied Biosystems)を用いて合成した。 qRT-PCR反応は、1×SYBR Green Master Mix buffer(Thermo Scientific)および900nM遺伝子特異的プライマー(または300nM L32プライマー)を含む10μl容量でセットアップされた。反応は、CFX96 Real-Time System (Bio-Rad) で行った。遺伝子発現データは、ΔΔCT法を用いてリボソームタンパク質L32に正規化した。プライマー配列は、補足情報に記載した。
グリコーゲン測定
グリコーゲンの測定は、以前に記載したように、すり潰した冷凍肝臓ホモジネートで行った(Pfleiderer, 1974)。
計算と統計方法
数値は平均値±SEMで表示した。グラフのプロットと統計解析には、特に指示のない限り、GraphPad Prism(version 6, GraphPad Software)を使用した。2群の統計的比較はStudent's t test, Wilcoxon matched-pairs signed rank test, Mann-Whitney U testで行った。3群以上の比較は一元配置分散分析(ANOVA)で行い、多重比較は Tukey post-tests で補正を行った。
著者による貢献
P.K.D.、A.N.、I.B.、F.B.はプロジェクトの構想および設計を行った。P.K.D.、A.N.、A.H.は試料を採取した。P.K.D., A.N., R.A., Y.S.L., F.D.V., T.A., A.H., and E.M.は実験を行った。P.K.D, A.N, R.A., Y.S.L, F.D.V., T.A., I.B., F.B. はデータの解析と解釈を行った。P.K.D.、A.N.、I.B.、F.B.は論文を執筆した。全著者が原稿にコメントした。
利益相反
A.N.、I.B.、F.B.はProPrev ABの創設者であり、株主である。
謝辞
原稿の編集はRosie Perkinsに、動物飼育はCarina Arvidsson, Sara Nordin-Larsson, Ulrica Enqvist, Caroline Wennberg, Zakarias Gulicに、454シーケンス解析はストックホルム国立ゲノム基盤に、ショットガンメタゲノム解析およびRNA-seq解析はSahlgrenska Academyのゲノム中核施設に、それぞれ感謝いたします。本研究は、Antidiabetic Food Center - a VINNOVA Centre of Excellence in Research and Innovation at Lund University、Swedish Research Council、Torsten and Ragnar Söderbergs財団、IngaBritt and Arne Lundberg財団、Swedish Foundation for Strategic Research、Knut and Alice Wallenberg財団、Västra Götaland地方とサルグランスカ大学病院間の医学研修と臨床研究に関する地域協定(ALF)から支援を受けて実施したものです。F.D.V.はEMBO Long-Term Fellowship ALTF 1305-2014の受給者です。F.B.はERC Consolidator Grant(欧州研究会議、コンソリデーターグラント615362 - METABASE)の受給者です。
Accession Numbers
本論文で報告された454パイロシーケンス配列のアクセッション番号はNCBI: SRP062889 です。本論文で報告したメタゲノム配列のアクセッション番号は、MG-RAST: 13068 (http://metagenomics.anl.gov/linkin.cgi?project=13068)です。この論文で報告されたRNA-seq配列のアクセッション番号は、NCBI: SRP064375 です。
補足情報
pdfをダウンロード (.79 MB)
pdfファイルに関するヘルプ
ドキュメントS1. 実験手順の補足、図S1〜S7、表S1〜S4
参考文献
アルムガム M.
レーズ J.
ペレティエ E.
ル・パズリエ D.
山田 毅
メンデD.R.
フェルナンデスG.R.
タップ J.
ブルルス T.
Batto J.-M.
他
MetaHITコンソーシアム
ヒト腸内細菌叢の腸型。
Nature. 2011; 473: 174-180
記事で見る
スコープス (4335)
PubMed
クロスリファレンス
グーグル奨学生
Bäckhed F.
Ding H.
Wang T.
フーパー L.V.
Koh G.Y.
ナギー A.
セメンコビッチ・C.F.
ゴードン J.I.
脂肪蓄積を制御する環境因子としての腸内細菌叢。
Proc. Natl. Acad. Sci. USA. 2004; 101: 15718-15723
記事で見る
スコープス (4237)
パブコメ
クロスリファレンス
グーグル奨学生
ベッケドF.
マンチェスターJ.K.
セメンコビッチC.F.
ゴードン J.I.
無菌マウスにおける食事誘発性肥満に対する抵抗性の基礎となるメカニズム。
Proc. Natl. Acad. Sci. USA. 2007; 104: 979-984
記事で見る
スコープス (1889)
PubMed
クロスリファレンス
グーグル奨学生
ベンジャミン・Y.
ホッホバーグ Y.
偽発見率をコントロールする。多重検定への実用的で強力なアプローチ。
王立統計学会誌。シリーズB(方法論)。1995; 57: 289-300
論文で見る
クロスリファレンス
Google Scholar
ボルガー A.M.
ローゼM.
ウサデル B.
Trimmomatic: イルミナ配列データのための柔軟なトリマー。
Bioinformatics. 2014; 30: 2114-2120
記事で見る
スコープス (26790)
PubMed
クロスリファレンス
グーグル奨学生
ボレンM.
ケペンスS.
Stalmans W.
肝臓におけるグリコーゲン代謝の特異的特徴。
Biochem. J. 1998; 336: 19-31
記事で見る
スコープス (321)
PubMed
クロスリファレンス
グーグル奨学生
コティヤールA.
ケネディS.P.
コングL.C.
プリフティ E.
ポンズ N.
ル・シャトリエE.
アルメイダ M.
クインキス B.
レヴェネス F.
ガレロン N.
et al.
ANR MicroObes コンソーシアム
腸内細菌遺伝子の豊かさにおける食事介入の影響。
Nature. 2013; 500: 585-588
記事で見る
スコープス (1194)
PubMed
クロスリファレンス
グーグル奨学生
デビッドL.A.
モーリスC.F.
カーモディR.N.
グーテンバーグD.B.
バトンJ.E.
ウルフ・B・E
リン A.V.
デブリン A.S.
ヴァルマ Y.
フィッシュバッハM.A.
他
食事がヒトの腸内細菌叢を迅速かつ再現性高く変化させる。
ネイチャー. 2014; 505: 559-563
記事で見る
スコープス (5404)
PubMed
クロスリファレンス
グーグル奨学生
デ・フィリッポC.
カヴァリエリD.
ディ・パオラM.
ラマゾッティM.
プーレ J.B.
マサール S.
コリーニ S.
ピエラッチーニ G.
リオネッティP.
ヨーロッパとアフリカ農村部の小児における比較研究により明らかになった腸内細菌叢の形成における食事の影響。
Proc. Natl. Acad. Sci. USA. 2010; 107: 14691-14696
記事で見る
スコープス (3629)
パブコメ
クロスリファレンス
グーグル奨学生
デバッダF.
コバチェバ-ダッチャリーP.
Goncalves D.
Vinera J.
ジトゥン C.
ドゥシャンプト A.
Bäckhed F.
Mithieux G.
微生物が生成する代謝産物は、腸-脳神経回路を介して代謝的利益を促進する。
Cell. 2014; 156: 84-96
記事で見る
スコープス (1228)
PubMed
概要
全文
全文PDF
Google Scholar
デンマン S.E.
トムキンスN.W.
マクスウィーニー C.S.
抗メタン生成化合物ブロモクロロメタンに反応したルーメンメタン生成菌集団の定量および多様性解析。
FEMS Microbiol. Ecol. 2007; 62: 313-322
記事で見る
スクープ (386)
PubMed
クロスリファレンス
グーグル奨学生
デュルフE.M.
カニP.D.
クラウス S.P.
フエンテス S.
ピュイラートP.G.
ネーリンクA.M.
ビンデルスL.B.
デ・ボスW.M.
ギブソンG.R.
ティッセン・J.-P.
デルゼンヌ N.M.
プレバイオティクス概念への洞察:肥満女性におけるイヌリンタイプフルクタンによる探索的二重盲検介入試験からの教訓。
Gut. 2013; 62: 1112-1121
記事で見る
スコープス (532)
PubMed
クロスリファレンス
グーグル奨学生
ドッド D.
ムーン Y.-H.
スワミナサン K.
マッキー R.I.
Cann I.K.O.
Prevotella bryantiiによるキシラン分解のトランスクリプトーム解析とキシラン分解性バクテロイデスのエネルギー獲得に関する洞察.
J. Biol. Chem. 2010; 285: 30261-30273
記事で見る
スコープス (85)
PubMed
クロスリファレンス
グーグル奨学生
ドッドD.
マッキーR.I.
Cann I.K.O.
キシラン分解、ルーメンとヒト大腸菌に共通する代謝特性。
Mol. Microbiol. 2011; 79: 292-304
記事で見る
スコープス (159)
PubMed
クロスリファレンス
グーグル奨学生
ダンカン S.H.
ベレンゲールA.
ホルトロップG.
ジョンストン A.M.
フリント H.J.
Lobley G.E.
肥満者の炭水化物摂取量の減少により、糞便中の酪酸および酪酸産生菌の濃度が減少した。
Appl. Environ. Microbiol. 2007; 73: 1073-1078
論文で見る
スコープス (650)
PubMed
クロスリファレンス
グーグル奨学生
エル・カオタリA.
アルムーガムF.
ゴードンJ.I.
ラウール D.
Henrissat B.
ヒト腸内細菌叢における炭水化物活性酵素の存在量と多様性。
Nat. Rev. Microbiol. 2013; 11: 497-504
記事で見る
スコープス (837)
PubMed
クロスリファレンス
グーグル奨学生
フリント H.J.
スコットK.P.
ダンカンS.H.
ルイス P.
フォラノE.
腸内における複雑な炭水化物の微生物分解。
腸内細菌。2012; 3: 289-306
記事で見る
スコープス (1110)
PubMed
クロスリファレンス
グーグル奨学生
ハーモンJ.P.
モランN.A.
アイブス A.R.
環境変化に対する種の反応:食物網の相互作用と進化の影響。
サイエンス. 2009; 323: 1347-1350
記事で見る
スコープス (177)
PubMed
クロスリファレンス
グーグル奨学生
泉田裕子
矢作直樹
竹内洋子
西美和子
志鎌 淳
宝田明彦
増田洋平
久保田真一
松坂桃李
中川由紀夫.
他。
空腹時のグリコーゲン不足が肝臓-脳-脂肪の神経回路を刺激し、脂肪の利用を促進させる。
Nat. Commun. 2013; 4: 2316-2320
記事で見る
スコープス (60)
PubMed
クロスリファレンス
グーグル奨学生
ヨハンソンE.V.
ニルソンA.C.
エストマンE.M.
ビョルク I.M.
健康な成人における16時間の糖代謝、食欲および自発的な食物摂取に及ぼす大麦の難消化性炭水化物の効果。
Nutr. J. 2013; 12: 46
記事で見る
スコープス (74)
PubMed
クロスリファレンス
グーグル奨学生
カールソンF.H.
トレマローリV.
Nookaew I.
ベルグストロム G.
ベアC.J.
フェーガーバーグ B.
ニールセン J.
Bäckhed F.
欧州女性の正常、糖尿病性グルコースコントロール障害、および糖尿病性グルコースコントロールにおける腸内メタゲノム。
Nature. 2013; 498: 99-103
記事で見る
スコープス (1795)
PubMed
クロスリファレンス
グーグル奨学生
コエス R.A.
ワン・Z.
レビソンB.S.
バファ J.A.
オーグ E.
シーイーB.T.
ブレットE.B.
フー X.
ウー・Y.
Li L.
他。
赤身肉の栄養素であるL-カルニチンの腸内細菌叢による代謝は動脈硬化を促進する。
Nat. Med. 2013; 19: 576-585
記事で見る
スコープス (2646)
パブコメ
クロスリファレンス
グーグル奨学生
コフィアV.
イッセルリンR.
ブーチャンA.
バーダー G.
Social Network: a Cytoscape app for visualizing co-publication networks(ソーシャルネットワーク:共同出版ネットワークを可視化するためのCytoscapeアプリ)。
F1000Res. 2015; 4: 481
記事で見る
スコープス (11)
PubMed
クロスリファレンス
グーグル奨学生
コレンO.
ナイツD.
ゴンザレスA.
ウォルドロンL.
セガタ・N.
ナイト R.
ハッテンハワー C.
レイ・R.E.
人体全体のエンテロタイプのガイド:ヒトマイクロバイオームデータセットにおける微生物コミュニティ構造のメタ分析。
PLoS Comput. Biol. 2013; 9: e1002863
記事で見る
スコープス (355)
PubMed
クロスリファレンス
グーグル奨学生
コルペラ K.
フリントH.J.
ジョンストン A.M.
ラッピ J.
ポウタネン K.
デウルフ E.
デルゼンヌ N.
デ・ヴォスW.M.
サロネンA.
腸内細菌叢のシグネチャーは、肥満者における食事介入に対する宿主および微生物叢の反応を予測する。
PLoS ONE. 2014; 9: e90702
記事で見る
スコープス(139)
PubMed
クロスリファレンス
グーグル奨学生
コバチェバ-ダッチャリーP.
アローラT.
栄養、腸内細菌叢とメタボリックシンドローム。
ベストプラクティス。研究。Gastroenterol. 2013; 27: 59-72
記事で見る
スコープス (85)
パブコメ
概要
全文
全文PDF
Google Scholar
ランペ J.W.
ナバーロ S.L.
ハラーM.A.J.
Shojaie A.
食事介入に対する反応の個人間差:個人化された食事勧告に向けたオミックスプラットフォームの統合。
Proc. ニュートリションソサイエティー 2013; 72: 207-218
記事で見る
スコープス(61)
PubMed
クロスリファレンス
グーグル奨学生
ル・シャトリエE.
ニールセンT.
秦 J.
プリフティE.
ヒルデブランド F.
ファロニー G.
アルメイダ M.
アルムガムM.
バトー・J.-M.
ケネディ S.
他
MetaHITコンソーシアム
ヒト腸内細菌群の豊富さは、代謝マーカーと相関している。
Nature. 2013; 500: 541-546
記事で見る
スコープス (2687)
PubMed
クロスリファレンス
グーグル奨学生
ライチE.C.M.
ウォーカーA.W.
ダンカン S.H.
ホルトロップG.
フリント H.J.
ヒト糞便由来細菌による不溶性基質の選択的コロニー形成
Environ. Microbiol. 2007; 9: 667-679
記事で見る
スコープス (200)
パブコメ
クロスリファレンス
グーグル奨学生
レスニエフスキーL.A.
ホッシュ S.E.
ニールスJ.G.
デ・ルカC.
パシュムフォルーシュ M.
ルメンC.N.
チェン S.-H.
スカデングM.
サルティエル A.R.
オレフスキー J.M.
骨髄特異的Cap遺伝子欠失は高脂肪食誘発性インスリン抵抗性から保護される。
Nat. Med. 2007; 13: 455-462
記事で見る
スコープス (92)
PubMed
クロスリファレンス
グーグル奨学生
ロズポーンC.
ハマディM.
Knight R.
UniFrac--微生物コミュニティの多様性を系統的に比較するためのオンラインツール。
BMC Bioinformatics. 2006; 7: 371
記事で見る
スコープス (1155)
PubMed
クロスリファレンス
グーグル奨学生
メイシーJ.M.
リュングダールL.G.
Gottschalk G.
Bacteroides fragilisにおけるコハク酸およびプロピオン酸の生成経路。
J. Bacteriol. 1978; 134: 84-91
記事で見る
PubMed
クロスリファレンス
Google Scholar
マーテンス E.C.
チェンH.C.
ゴードン J.I.
粘膜糖鎖採食は、糖質分解ヒト腸内細菌共生体のフィットネスとトランスミッションを強化する。
Cell Host Microbe. 2008; 4: 447-457
記事で見る
スコープス (542)
PubMed
要旨
全文
全文PDF
Google Scholar
マーテンス E.C.
ロウE.C.
チェン H.
プードロN.A.
ウー・エム
マクナルティ N.P.
アボット D.W.
ヘンリサットB.
ギルバート・H.J.
ボラムD.N.
ゴードン J.I.
ヒト腸内共生細菌による植物細胞壁多糖の認識と分解。
PLoS Biol.
記事で見る
スコープス (498)
PubMed
クロスリファレンス
グーグル奨学生
松木利夫
渡辺和男
藤本純一
宮本佳明
高田 孝
松本和彦
小柳津浩二
田中 亮
ヒト糞便中の優勢菌の検出と同定のための16S rRNA遺伝子標的グループ特異的プライマーの開発.
Appl. Environ. Microbiol. 2002; 68: 5445-5451
記事で見る
スコープス (500)
パブコメ
クロスリファレンス
グーグル奨学生
メンザーギC.
パローニG.
デ・ボニスC.
ココ A.
ヴィーニャ C.
ミスキオG.
ランナ P.
タッシ V.
バッチ S.
Trischitta V.
プロテインチロシンホスファターゼ受容体タイプf(PTPRF)遺伝子座は、2型糖尿病における冠動脈疾患と関連している。
J. Intern. Med. 2008; 263: 653-654
記事で見る
スクープ (7)
PubMed
クロスリファレンス
グーグル奨学生
マイヤーF.
パールマンD.
D'Souza M.
オルソンR.
グラスE.M.
クバル M.
パックジアン・T
ロドリゲス A.
スティーブンス R.
ウィルケA.
他
メタゲノムRASTサーバー - メタゲノムの自動系統樹・機能解析のための公開リソース。
BMC Bioinformatics. 2008; 9: 386
記事で見る
スコープス (2542)
パブコメ
クロスリファレンス
グーグル奨学生
ニルソンA.
グランフェルトY.
オストマンE.
プレストン T.
ビョルクI.
その後の標準化された朝食で耐糖能に穀物ベースの夕方の食事の難消化性炭水化物のGIとコンテンツの効果。
Eur. J. Clin. 2006; 60: 1092-1099
記事で見る
スコープス (83)
PubMed
クロスリファレンス
グーグル奨学生
ニルソンA.
オストマンE.
プレストンT.
Björck私。
その後の標準化された朝食で耐糖能に夕方の食事の穀物繊維のGI対コンテンツの影響。
ユーロ。J.臨床。2008; 62: 712-720
記事で見る
スコープス (62)
PubMed
クロスリファレンス
グーグル奨学生
ニルソンA.C.
ヨハンソン-ボルE.V.
Björck I.M.
大麦の穀粒をベースとした製品の3日間介入後の腸内ホルモンおよびインスリン感受性指数の増加:健康な中年被験者における無作為化クロスオーバー試験。
Br. J. Nutr. 2015; Aug 11: 1-9
記事で見る
Google Scholar
プフライダーG.
グリコーゲン:アミログルコシダーゼによる定量。
バーグマイヤーHU, ディアフィールドビーチ, フロリダ州, US1974
論文で見る
Google Scholar
ピメンテル M.
Gunsalus R.P.
ラオ・S.S.C.
Zhang H.
ヒトの健康および疾病におけるメタン菌。
Am. J. Gastroenterol. 2012; 1: 28-33
記事で見る
クロスリファレンス
Google Scholar
Qin J.
Li Y.
Cai Z.
Li S.
Zhu J.
Zhang F.
Liang S.
Zhang W.
グァン Y.
シェンD.
他。
2型糖尿病における腸内細菌叢のメタゲノム・ワイド関連研究。
Nature. 2012; 490: 55-60
記事で見る
スコープス (3881)
PubMed
クロスリファレンス
グーグル奨学生
Rコアチーム
R: A language and enviorment for statistical computing.
R Foundation for Statistical Computing, Vienna, Austria2015
http://www.R-project.org/
記事で見る
Google Scholar
ランペリ S.
シュノアS.L.
コンソランディC.
トゥローニ S.
セヴェルニーニ M.
ペアーノC.
ブリギディP.
クリッテンデンA.N.
ヘンリーA.G.
カンデラ M.
ハドザ族狩猟採集民の腸内細菌叢のメタゲノム配列決定。
Curr. Biol. 2015; 25: 1682-1693
記事で見る
スコープス(211)
PubMed
アブストラクト
全文
全文PDF
Google Scholar
サロネン A.
ニッキラJ.
ヤランカ-トゥオビネンJ.
インモネン O.
ラジリッチ-ストヤノビッチM.
ケッコネンR.A.
パルヴァ A.
デ・ヴォス W.M.
糞便DNA抽出法と系統樹マイクロアレイの比較解析:機械的細胞溶解を用いた細菌および古細菌DNAの効果的な回収。
J. Microbiol. Methods. 2010; 81: 127-134
記事で見る
スコープス (377)
パブコメ
クロスリファレンス
グーグル奨学生
サロネンA.
ラハティL.
サロヤルヴィJ.
ホルトロップG.
コルペラ K.
ダンカン S.H.
デイト P.
ファルカソン F.
ジョンストン A.M.
ロブリーG.E.
他
肥満男性における腸内細菌叢の組成と発酵産物に対する食事と個人差の影響。
ISME J. 2014; 8: 2218-2230
記事で見る
スコープス (382)
PubMed
クロスリファレンス
グーグル奨学生
サミュエル・B.S.
ゴードン J.I.
宿主-古細菌-細菌相互作用のヒト化グノトビオティックマウスモデル。
Proc. Natl. Acad. Sci. USA. 2006; 103: 10011-10016
論文で見る
スコープス (471)
PubMed
クロスリファレンス
グーグル奨学生
トレマローリV.
Bäckhed F.
腸内細菌叢と宿主の代謝の間の機能的相互作用。
Nature. 2012; 489: 242-249
記事で見る
スコープス (2872)
PubMed
クロスリファレンス
グーグル奨学生
ウォーカー A.W.
インスJ.
ダンカンS.H.
ウェブスター L.M.
ホルトロップG.
ゼー X.
ブラウン D.
スターズM.D.
スコット P.
ベルジェラットA.
他。
ヒト大腸内細菌叢における優勢菌群および食事応答性菌群。
ISME J. 2011; 5: 220-230
記事で見る
スコープス (1094)
PubMed
クロスリファレンス
グーグル奨学生
ワン・Z.
クリプフェルE.
ベネットB.J.
コエス R.
レビソンB.S.
ドゥガーB.
フェルドスタインA.E.
ブレットE.B.
フー X.
チョンY.-M.
ら。
腸内細菌叢によるホスファチジルコリンの代謝は心血管系疾患を促進する。
Nature. 2011; 472: 57-63
記事で見る
スコープス (3387)
PubMed
クロスリファレンス
グーグル奨学生
ウー・ジー・ディー
チェン J.
ホフマンC.
ビッティンガー K.
チェン Y.Y.
キールボー S.A.
ビュートラ M.
ナイツ D.
ウォルターズ W.A.
ナイトR.
他
長期的な食事パターンと腸内細菌の腸型との関連性。
Science. 2011; 334: 105-108
記事で見る
スコープス (4000)
PubMed
クロスリファレンス
グーグル奨学生
ウーG.D.
コンファーC.
チェンE.Z.
スミス S.A.
シャー・R.D.
ビッティンガー K.
チェフードC.
アルベンバーグL.G.
ネッセル L.
ギルロイE.
他。
菜食主義者と雑食主義者の比較メタボロミクスにより、食事に依存した腸内細菌叢の代謝物産生に制約があることが明らかになりました。
Gut. 2014; (gutjnl-2014-308209)
https://doi.org/10.1136/gutjnl-2014-308209
記事で見る
スコープス(305)
クロスリファレンス
Google Scholar
Xu J.
ビュルセル M.K.
ヒムロッドJ.
Deng S.
カーマイケル・L.K.
チェン H.C.
フーパー L.V.
ゴードン J.I.
ヒト-バクテロイデス・テタイオオミクロン共生のゲノム的見解。
Science. 2003; 299: 2074-2076
記事で見る
スコープス (1014)
PubMed
クロスリファレンス
グーグル奨学生
ゼー X.
ダンカンS.H.
ルイスP.
フリント H.J.
Ruminococcus bromiiは、ヒト大腸におけるレジスタントスターチの分解に重要な役割を果たす種である。
ISME J. 2012; 6: 1535-1543
記事で見る
スコープス (552)
PubMed
クロスリファレンス
グーグル奨学生
張 浩
ディベイズJ.K.
ズッコロ A.
クドルナ D.
ブレイドッティ M.
ユウ Y.
パラメスワラン P.
クロウエル M.D.
ウィング R.
リトマンB.E.
Krajmalnik-Brown R.
肥満および胃ろう造設後のヒト腸内細菌叢。
プロック。Natl.Acad。Sci.USA。2009; 106: 2365-2370
記事で見る
スコープス (1350)
パブコメ
クロスリファレンス
Google Scholar
記事情報
出版年譜
掲載されました。2015年11月5日(木
受理されました。2015年10月7日(木
改訂版で受理。2015年7月3日(木
受理されました。2014年9月12日
識別情報
DOI: https://doi.org/10.1016/j.cmet.2015.10.001
著作権について
© 2015 Elsevier Inc. 発行:エルゼビア株式会社
ユーザーライセンス
エルゼビアユーザーライセンス|インフォメーションアイコンを再利用する方法
サイエンスダイレクト
ScienceDirectでこの論文にアクセスする
図
図サムネイルfx1
グラフィカルアブストラクト
図のサムネイル gr1
図1標準化された朝食後の非応答者と応答者のグルコースおよびインスリンのプロファイル。
図2:gr2
図2WWBまたはBKBを3日間摂取した非応答者および応答者における腸内細菌叢の変化
図2WWB/BKB摂取による非応答者と応答者の腸内細菌叢の変化
図3ベースラインとBKB摂取後の非応答者と応答者のメタゲノムの分類学的組成
図3.BKB摂取後の非応答者と応答者のメタゲノム構成
図4プレボテラ菌のマウス糖代謝への有益な効果は食事依存的であり、P. copriはB. thetaiotaomicron誘発の耐糖能異常から保護する。
図5:gr5
図5BKB後に採取した応答者の腸内細菌叢をGFマウスに移植すると耐糖能が改善される
図1.gr6
図6BKB後に摂取した応答者の腸内細菌叢をGFマウスに移植すると、肝グルコースのグリコーゲンへの貯蔵が促進される。
表
表1B. thetaiotaomicronを単独培養したマウスの大腸近位部および肝臓の生検で有意に制御された遺伝子トップ10