PM2.5による神経毒性は、腸内細菌叢の乱れによるビタミンB12代謝のアンバランスを介して誘導される

メインコンテンツへスキップ
テイラー・アンド・フランシス・オンライン・ホームページ
ログイン|会員登録
カート
腸内微生物
第15巻 2023年-第2号
オープンアクセス
281
閲覧数
0
これまでのCrossRef引用
2
アルトメトリック
聞く
研究論文
PM2.5による神経毒性は、腸内細菌叢の乱れによるビタミンB12代謝のアンバランスを介して誘導される
https://www.tandfonline.com/doi/full/10.1080/19490976.2023.2267186
朱玄,夏燕亭,王煥煥,石麗華,殷宏平,顧明爾 & show all
論文 2267186|2023年4月25日受領、2023年10月02日受理、オンライン公開:2023年10月16日
引用
https://doi.org/10.1080/19490976.2023.2267186
クロスマークのロゴクロスマーク
この記事で
PDF EPUB
共有アイコン
要旨
大気中の微小粒子状物質(PM2.5)は、有毒で有害な物質を伴いやすく、認知機能障害など人の健康に深刻な害を及ぼす。ビタミンB12(VitB12)は細菌によって合成される必須微量栄養素であり、栄養およびシグナル伝達分子として神経伝達物質の合成に寄与する。しかし、PM2.5曝露におけるVitB12による認知機能障害の減弱と腸内細菌叢の調節との関係は解明されていない。本研究では、線虫(C. elegans)において、PM2.5が神経伝達物質受容体の著しい遺伝子発現変化やVitB12含量の低下とともに、行動障害や神経細胞障害を引き起こすことを明らかにした。VitB12アナログであるメチルコバラミン(MeCbl)は、線虫におけるPM2.5誘発神経毒性を緩和した。さらに、in vivoおよびin vitroモデルを用いて、PM2.5への長期曝露が腸内細菌叢の構造の変化を引き起こし、その結果、VitB12関連代謝経路のバランスが崩れ、認知障害が生じることを発見した。MeCblの補充は、細菌の多様性を増加させ、有害物質の含有量を減少させ、短鎖脂肪酸(SCFA)と神経伝達物質の濃度を対照群のレベルまである程度回復させることができた。ここで、PM2.5による害を軽減する新たな標的が発見され、腸内および細胞内の神経伝達障害を緩和するMeCblが供給された。また、この結果は、PM2.5に暴露された人々の代謝障害を改善するために、ヒトの腸内細菌叢の調整をターゲットとしたVitB12の使用の参考となる。
keywords: ビタミンB12メチルコバラミン大気中微粒子物質試験管内大腸シミュレーションシステム腸内細菌神経伝達物質
前の記事
次の記事
はじめに
大気汚染は、粒子状物質(PM)、一酸化炭素、鉛、二酸化窒素、オゾン、二酸化硫黄の複雑な混合物である。中でも、空気力学的直径が2.5μm未満の大気中の粒子状物質、すなわちPM2.5は、人間の健康に最も有害である。PM2.5の組成は複雑で、ベンゾ[a]ピレン、多環芳香族炭化水素、ニトロ、酸素系PAHsなどの有機物だけでなく、硫酸塩、硝酸塩、金属などの無機化学物質も豊富に含んでいる。 引用1,引用2 過去20年間の累積的な証拠から、大気汚染は喘息などの呼吸器疾患、心血管疾患、代謝障害、がんと密接に関連していることが示されている。 引用7 さまざまな大気汚染成分の中でも、PM2.5は認知障害と強く関連する主な要因の一つである。引用8,引用9 PM2.5は血液脳関門を通過して神経系に入り込み、過剰な活性酸素種(ROS)と炎症因子を産生し、神経細胞のアポトーシスを引き起こす、 引用12 PM2.5に暴露されたNeuro-2a細胞では、オートファジー-リソソーム機能障害も調査され、リソソーム膜の透過性に関連した細胞死が引き起こされた。引用13 とはいえ、PM2.5による神経毒性の具体的なメカニズムは依然として不明である。
引用14 脳腸軸は、消化管と中枢神経系の間の双方向コミュニケーションネットワークである。腸内微生物が産生する関連代謝産物が宿主と相互作用し、中枢神経系の機能と行動を制御している。PM2.5は、バクテロイデーテス属/ファーミキューテス属の比率の減少を伴う、成体マウスの体重の明らかな減少を引き起こす可能性がある。 引用17 PM2.5が腸内細菌に及ぼす影響は広く懸念されており、引用18 PM2.5と腸内細菌叢の具体的な関係を詳細に調べることは特に重要である。
コバラミンとしても知られるビタミンB12(VitB12)は、多くの必須生化学プロセスにおいて補酵素または補酵素として作用し、VitB12の生理的役割は、コバルトを介した化学反応の中心、特にコバルトイオンが直接コバルト-炭素結合を形成する能力を中心に展開する。その生合成プロセスには様々な酵素の関与が必要であり、その合成経路は極めて複雑である。VitB12の特定種の指向性生産と選択的吸収は、他の細菌との競合を減らすことができ、細菌間の相互依存的共生の確立を助長する。引用20,引用21 私たちの以前の研究では、メチルコバラミン(MeCbl)の補給が、C57BL/6マウスにおいてPM2.5によって誘発された認知機能障害を緩和することがわかった。PM2.5におけるMeCblによる認知機能障害の減弱と腸内細菌叢の調節との関係は解明されていない。
本研究では、物質の神経毒性を決定する優れたモデルである線虫を用いて、まずPM2.5による神経毒性を評価した。さらに、マウス実験とin vitro模擬大腸発酵モデル実験を組み合わせて、PM2.5による代謝異常の緩和におけるMeCblの基本的なメカニズムを調べた。まず、PM2.5、細菌群集組成および体内VitB12レベルの関係を明らかにすることを目的とした。さらに、PM2.5とVitB12の影響を受ける腸内細菌叢と神経伝達物質との相互作用を理解することを目指した。この結果は、MeCblの作用機序と腸内細菌叢の標的制御に関するさらなる研究の参考となる。
研究結果
PM2.5曝露が線虫に誘発する毒性はMeCblによって軽減される
線虫は、その明確な遺伝的背景、単純な構造、短いライフサイクル、培養の容易さから、老化研究の分野では古典的なモデル生物となっている。線虫の腸内構造はヒトの腸内構造と類似しており、豊富で多様なマイクロバイオームを保有しているためである。線虫の神経系は、その運動行動を制御・調節することができ、線虫の運動行動能力は通常、頭部屈曲頻度、胴体屈曲頻度、咽頭ポンピングによって評価・分析される。Citation25 対照群と比較して、PM2.5投与群では胴体屈曲頻度が低かったが、頭部屈曲頻度は高かった。さらに、PM2.5またはMeCblの両補給は、線虫の嚥下機能に明らかな影響を与えなかった(図1c)。さらに、PM2.5に暴露された線虫の生存率は明らかに低下し、9日目には半数が死亡したのに対し、対照群では14日目でも50%の生存率を維持した。MeCblによる介入は生存率を効果的に改善した;ミミズにMeCblを供給すると、9日目に約70%の生存率が観察された(図1d)。これらのデータを総合すると、PM2.5誘発毒性はMeCblによって緩和されることが示唆される。
図1. 線虫におけるPM2.5誘発毒性に対するMeCblの影響。(a-c)ミミズの体曲げ時間(1分)、頭振り時間(20秒)、咽頭ポンプ速度(20秒)に対するMeCbl介入の影響。*p < .05; **p < .01; ns: スチューデントのt検定で有意ではなかった。(d) 異なる処理を行ったミミズの寿命アッセイ。(e) 化学走性を介した連合学習モデル;(f) MeCblの有無に関わらず、72時間PM2.5曝露後の化学走性学習行動;(g) 線虫の化学走性を解析するための模式図。(h)MeCblの有無に関わらず72時間PM2.5に暴露した線虫の走化性行動;化学物質に対する線虫の感受性を調べるためにインドールとデカナール(エタノールに溶解)を用いた;(i)線虫の採餌行動を解析するための模式図;(j)MeCblの有無に関わらず72時間PM2.5に暴露した線虫の採餌行動。異なる文字のバーで示した値は有意に異なる(p < .05、一元配置分散分析)。CI、走化性指数。
図1. 線虫におけるPM2.5誘発毒性に対するMeCblの効果。(a-c)ミミズの体曲げ時間(1分)、頭振り時間(20秒)、咽頭ポンプ速度(20秒)に対するMeCbl介入の影響。*p < .05; **p < .01; ns: スチューデントのt検定で有意ではなかった。(d) 異なる処理を行ったミミズの寿命アッセイ。(e) 化学走性を介した連合学習モデル;(f) MeCblの有無に関わらず、72時間PM2.5曝露後の化学走性学習行動;(g) 線虫の化学走性を解析するための模式図。(h)MeCblの有無に関わらず72時間PM2.5に暴露した線虫の走化性行動;化学物質に対する線虫の感受性を調べるためにインドールとデカナール(エタノールに溶解)を用いた;(i)線虫の採餌行動を解析するための模式図;(j)MeCblの有無に関わらず72時間PM2.5に暴露した線虫の採餌行動。異なる文字のバーで示した値は有意に異なる(p < .05、一元配置分散分析)。CIは走化性指数。
フルサイズ表示
線虫の走化性、摂食、学習、記憶行動に対するPM2.5曝露の影響
線虫は液体や空気中の化学物質に反応し、マイクロモル濃度域の水溶性誘引物質や揮発性物質を感知し、その結果、傾向行動や回避行動を引き起こす。このモデルでは、線虫をNaClと餌のないNGM培地で一定期間培養し、NaClシグナルと飢餓シグナルを統合させ、線虫が飢餓処理中のNaCl濃度を回避するようにした。PM2.5への曝露はNaCl領域における線虫数の増加をもたらし、MeCblの介入はこの異常を逆転させた(図1e,f)。走化性アッセイにはインドールとデカナールを選択した。対照群では、線虫はインドールに対する誘引とデカナールに対する反発を示したが、線虫がPM2.5に曝露されるとCI指数の絶対値が低下し、これはMeCblの補充によって部分的に回復した(図1g,h)。摂食行動は咽頭運動ニューロンによって制御されており、咽頭筋の収縮と弛緩を直接調整して摂食行動を完了させている。PM2.5は、非曝露グループと比較して、採餌能力の明らかな低下を引き起こした。この場合も、MeCblによる予防後はミミズの採餌行動が回復した(図1i,j)。これらの結果から、MeCblはPM2.5によって損傷を受けた線虫の化学走性行動の感受性と学習・記憶行動の障害に対して保護効果を示した。
線虫の神経細胞と神経伝達物質受容体に対するPM2.5曝露の影響
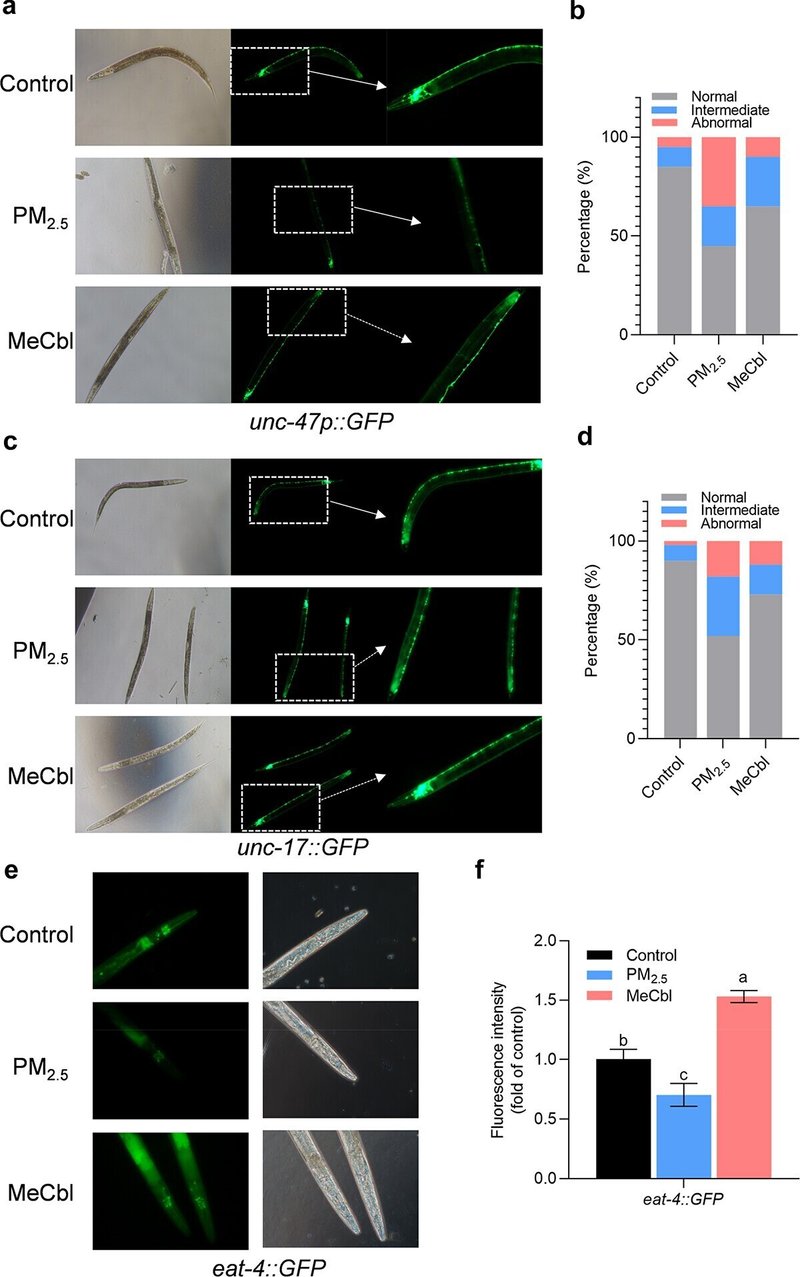
線虫の神経伝達物質は、合成、貯蔵、輸送、代謝、その他多くの生物学的プロセスを含め、哺乳類のものと高い類似性を持っている。PM2.5への曝露により、線虫の緑色蛍光タンパク質標識GABAニューロン(unc-47)、コリン作動性ニューロン(unc-17)、グルタミン酸作動性ニューロン(eat-4)の蛍光強度は程度の差こそあれ減少し、線虫の一連のニューロンがPM2.5曝露により損傷を受けたことが示唆された。MeCbl処理はPM2.5誘発神経毒性を部分的に防御した(図2)。
図2. PM2.5曝露72時間後のEG1285、LX929およびDA1240のGABAニューロン、コリン作動性ニューロンおよびグルタミン酸作動性ニューロンに対するMeCblの効果。(a)MeCbl処理の有無にかかわらず、PM2.5に曝露したワームのGABAニューロンの蛍光写真。(b) PM2.5およびMeCbl処理後のEG1285線虫の神経細胞傷害。(c)PM2.5とMeCbl処理の有無にかかわらず、線虫をPM2.5に暴露したときのコリン作動性神経細胞を示す蛍光写真。(d) PM2.5およびMeCbl処理後のLX929線虫の神経細胞傷害。神経細胞傷害の程度は、正常(神経細胞の完全性が良好)、中間(数個の神経細胞の劣化、5個未満)、異常(5個以上の神経細胞の劣化、不連続または誤配置)の3つのカテゴリーに分類した。(e)MeCbl処理の有無にかかわらず、PM2.5に曝露したときのミミズのグルタミン酸作動性ニューロンを示す蛍光写真。(f) 線虫におけるeat-4:GFPの相対蛍光強度。異なる文字のバーで示した値は有意に異なる(p < .05、一元配置分散分析)。
図2. PM2.5曝露72時間後のEG1285、LX929、DA1240のGABAニューロン、コリン作動性ニューロン、グルタミン酸作動性ニューロンに対するMeCblの影響。(a)MeCbl処理の有無にかかわらず、PM2.5に曝露したワームのGABAニューロンの蛍光写真。(b) PM2.5およびMeCbl処理後のEG1285線虫の神経細胞傷害。(c)PM2.5とMeCbl処理の有無にかかわらず、線虫をPM2.5に暴露したときのコリン作動性神経細胞を示す蛍光写真。(d) PM2.5およびMeCbl処理後のLX929線虫の神経細胞傷害。神経細胞傷害の程度は、正常(神経細胞の完全性が良好)、中間(数個の神経細胞の劣化、5個未満)、異常(5個以上の神経細胞の劣化、不連続または誤配置)の3つのカテゴリーに分類した。(e)MeCbl処理の有無にかかわらず、PM2.5に曝露したときのミミズのグルタミン酸作動性ニューロンを示す蛍光写真。(f) 線虫におけるeat-4:GFPの相対蛍光強度。異なる文字のバーで示した値は有意に異なる(p < .05、一元配置分散分析)。
フルサイズ表示
PM2.5曝露は線虫の神経細胞およびVitB12代謝に関与する遺伝子の発現を変化させた
メチオニン/アデノシルメチオニン(Met/SAM)サイクルは、葉酸代謝、プリンおよびチミン生合成を含む一炭素代謝の一部である。引用29,引用30 metr-1、mtrr-1、sams-1の遺伝子発現レベルの低下は、PM2.5に曝露した線虫で検出され、MeCblの補充によって回復した(図3a)。これら3つの遺伝子は、VitB12依存性のホモシステイン(Hcy)枯渇経路と関連していた(図3b)。Hcyは非タンパク質アミノ酸であり、ビタミンB群の助けを借りてメチオニンやシステインに再利用されるが、その欠乏は高ホモシステイン血症を引き起こす。VitB12が低いとHcyが蓄積し、神経変性疾患と関連性の高い深刻な酸化ストレスが引き起こされる。神経伝達物質が合成され放出された後、神経調節と神経信号の伝達の役割を果たすためには、対応する受容体に結合しなければならない。神経伝達物質の活性を調べるには、対応する受容体遺伝子の発現を検出することが重要である。神経伝達物質に対するPM2.5曝露とMeCbl介入の影響をさらに説明するために、9つの関連遺伝子のmRNAレベルを測定した。図3cに示すように、線虫をPM2.5に72時間曝露すると、eat-4、glt-3、unc-30、unc-46、unc-47遺伝子の発現が有意に上昇し、一方、MeCbl投与群では、非曝露群と比較してgab-1とacr-8遺伝子の発現が有意に低下した。興味深いことに、PM2.5は、メチル変換関連遺伝子よりも神経伝達物質受容体に関与する遺伝子発現に強い反応を引き起こし、これは、MeCblが一炭素サイクルへの参加に加えて、別の保護メカニズムを持つことを示唆している。線虫では、アシル-CoAデヒドロゲナーゼacdh-1はVitB12レベルのセンサー遺伝子として確立されており、VitB12レベルの低下はacdh-1の高発現を引き起こす。Pacdh-1:GFPワーム(VL749)は通常、内因性VitB12レベルを示すために用いられる。PM2.5に暴露された動物におけるVitB12レベルの低下を調べたところ、MeCblの補充によって回復したことから、VitB12の欠乏がPM2.5処理後のミミズの異常行動の原因のひとつである可能性が示唆された(図3d)。
図3. PM2.5に曝露した線虫におけるコバラミン代謝および神経伝達物質に関連する遺伝子のmRNAレベルに対するMeCblの効果。(a) PM2.5およびMeCblの影響を受けたMet/SAMサイクル遺伝子のレベル。 (b) 線虫におけるVitB12関連代謝経路の図。(c) PM2.5に暴露された線虫の神経伝達関連遺伝子発現に対するMeCblの影響。相対的遺伝子発現はact-1遺伝子で正規化した。(d) PM2.5に曝露したPacdh1:GFPトランスジェニック動物(VL749株)の蛍光写真と相対蛍光強度(MeCbl処理の有無にかかわらず)。アシル-CoAデヒドロゲナーゼacdh-1はB12レベルのセンサー遺伝子として確立されており、B12レベルが低下するとacdh-1の発現が上昇する。異なる文字のバーで示した値は有意に異なる(p < 0.05、一元配置分散分析)。
図3. PM2.5に曝露した線虫におけるコバラミン代謝および神経伝達物質に関連する遺伝子のmRNAレベルに対するMeCblの効果。(a) PM2.5およびMeCblの影響を受けたMet/SAMサイクル遺伝子のレベル (b) 線虫におけるVitB12関連代謝経路の図。(c) PM2.5に暴露された線虫の神経伝達関連遺伝子発現に対するMeCblの影響。相対的遺伝子発現はact-1遺伝子で正規化した。(d) PM2.5に曝露したPacdh1:GFPトランスジェニック動物(VL749株)の蛍光写真と相対蛍光強度(MeCbl処理の有無にかかわらず)。アシル-CoAデヒドロゲナーゼacdh-1はB12レベルのセンサー遺伝子として確立されており、B12レベルが低下するとacdh-1の発現が上昇する。異なる文字のバーで示された値は有意に異なる(p < .05、一元配置分散分析)。
拡大表示
PM2.5曝露マウスにおける腸内細菌組成の変化
神経伝達物質は、化学シナプスでの情報伝達に関与する多様な化合物群である。引用32 以前の研究では、MeCblがPM2.5による学習・記憶機能の障害を回復させることが示された。PM2.5によって誘発された毒性を改善するための神経伝達物質調節におけるMeCblの役割は、私たちの関心を集めている。MeCblの補充により、PM2.5に暴露されたマウスの腸管長の短縮と体重減少が回復したが、体重増加には3群間で有意差はなかった(図S1A-C)。図4aに示すように、PM2.5曝露によりマウスの腸内細菌叢のα多様性レベルが低下したが、MeCblの補充により回復したことから、MeCblは腸内細菌叢をより安定かつバランスよく維持していることが示された。さらに、細菌叢の変化の全体的な構造には3群間で有意差はなかった(図S1D)。腸内細菌叢は、ファーミキューテス(Firmicutes)とバクテロイデーテス(Bacteroidetes)の2つの門の細菌が支配的であったCitation33。門レベル(図S1E)では、ファーミキューテスとバクテロイデーテスがマウスの腸内細菌叢の支配的な細菌で、全門の平均87.70%を占めていた。疣贅菌はPM2.5群では2ヶ月目に、MeCbl群では4ヶ月目と5ヶ月目に多かった。後期では、対照群とPM2.5群では放線菌の割合が多く、MeCbl群では少なかった。ヒートマップを用いて、グループ間の属レベルの比較を行った(図4b)。対照群と比較して、PM2.5群ではラクトバチルスとパラバクテロイデスの存在量が増加し、MeCbl群ではこれらの細菌の存在量が減少した。一方、属レベルでは、バクテロイデス、プレボテラ、ルミノコッカス、アロプレボテラがPM2.5群で減少し、MeCbl群ではその逆で、特にバクテロイデスが減少した。上記のLactobacillus、Bacteroides、Prevotella、Ruminococcusなどの細菌は、繊維基質を利用して短鎖脂肪酸(SCFAs)を産生することができる引用34、Lactobacillus、Bacteroides、Parabacteroides、Ruminococcusは、神経伝達物質の合成に関連することが報告されている引用35。
図4. PM2.5に曝露したマウスの腸内細菌構成と代謝産物に対するMeCblの影響。(a)糞便微生物叢のα-多様性-シャノン指数、(b)マウスの腸管内細菌属の相対的存在量のヒートマップ、(c)マウスの腸内代謝物の主成分分析(PCA1とPCA2の差は53. 43%と45.75%であった);(d)マウスの腸内短鎖脂肪酸濃度;(e)マウスの腸内細菌叢の代謝物ヒートマップ;(f)マウスの代謝物と腸内細菌叢の相関分析。
図4. PM2.5に曝露したマウスの腸内細菌構成および代謝産物に対するMeCblの効果。(a)糞便微生物叢のα-diversity-Shannon指数、(b)マウスの腸管内細菌属の相対的存在量のヒートマップ、(c)マウスの腸内代謝物の主成分分析(PCA1とPCA2の差は53. 43%と45.75%であった);(d)マウスの腸内短鎖脂肪酸濃度;(e)マウスの腸内細菌叢の代謝物ヒートマップ;(f)マウスの代謝物と腸内細菌叢の相関解析。
フルサイズ表示
PM2.5曝露によるマウスのメタボロームプロファイルの変化
2ヶ月目のマウスの糞便を選択し、非標的メタボローム解析を行った。同じグループのポイントはまとまっていたが、異なるグループのポイントは散らばっており、サンプルの平行性が良好で、コントロール、PM2.5、MeCblグループ間で明らかな違いがあることが示された(図4c)。PM2.5は、γ-アミノ酪酸(GABA)、グリシン、グルタミン酸、ノルエピネフリン、アスパラギン酸、ドーパミンのレベルを増加させ、ヒスタミンとフェニルエチルアミンのレベルを減少させた(図4e)。
マウスの糞便中のSCFA濃度を図4dに示す。4ヶ月目と5ヶ月目において、PM2.5曝露マウスの酢酸含量は対照群より有意に低く、酪酸はすべての月において対照群よりPM2.5群で低かった。それにもかかわらず、MebClの添加は、酪酸の濃度が5ヶ月目に増加し、対照群よりもさらに高くなったことを除いては、PM2.5群と比較して、上記の2つの物質の含量に明らかな変化を与えなかった。2ヶ月目と3ヶ月目では、MebCl群のバレレート含量はPM2.5群よりも有意に高かった。このことから、マウスをPM2.5に曝露すると、腸内のSCFAsの総濃度が低下し、特に酢酸と酪酸の濃度が低下すると結論できる。VitB12の摂取は腸内の酪酸とバレレート濃度を増加させたが、酢酸含量は増加させなかった。
細菌叢と代謝産物の相関分析により、マウスがPM2.5に曝露された場合、ラクトバチルス属の存在量の増加とバクテロイデス属の存在量の減少が、グルタミン酸、GABA、ドーパミン含量上昇の主な要因であると考えられ、これはMeCblによって逆転させることができた(図4f)。
PM2.5曝露とMeCbl補充によるin vitro結腸シミュレーション系での腸内細菌叢と代謝物の変化
マウスとヒトの腸内細菌叢はかなり異なることを考慮して、ヒト腸内細菌叢の増殖と代謝をシミュレートできるin vitro模擬大腸発酵モデルをさらなる研究に使用した。大腸腔のSCFA含量を図5aに示す。PM2.5と比較して、MeCbl処理群(MeCbl-MおよびMeCbl-H)では酪酸含量の増加が観察された。吉草酸はMeCbl-H群にのみ存在し、他の群には存在せず、この結果は粘膜のそれと一致した(図5aおよびS2A)。PM2.5処理後にFaecalibacterium、Ruminococcus_2、AnaeroglobusおよびBifidobacteriumの存在量が減少したことは、酪酸産生量が低下した一因かもしれない(図5b)。
図5. PM2.5に曝露した試験管内結腸シミュレーションシステムの腸管内腔における細菌組成と代謝産物の変化に対するMeCblの効果。(a)結腸の腸管内腔におけるSCFAの変化のヒストグラム、(b)PM2.5とMeCblの影響を受けた酪酸産生と腸内細菌叢の変化の関係の漫画、(c)細菌属の相対存在量のヒートマップ、(d)細菌の二次代謝強度のヒートマップ、(e)代謝産物と腸内細菌叢の相関分析、(f)生物種と神経伝達物質の間の正準相関分析(CCA)。まずデータに対してデトレンドコレスポンデンス分析(DCA)を実施し、本研究の関連データのDCA1は3.7928で3より大きいため、CCAモデルを選択した。
図5. PM2.5に曝露したin vitro大腸シミュレーションシステムの腸管内腔における細菌組成および代謝産物の変化に対するMeCblの影響。(a)結腸の腸管内腔におけるSCFAの変化のヒストグラム、(b)PM2.5とMeCblの影響を受けた酪酸産生と腸内細菌叢の変化の関係の漫画、(c)細菌属の相対存在量のヒートマップ、(d)細菌の二次代謝強度のヒートマップ、(e)代謝産物と腸内細菌叢の相関分析、(f)生物種と神経伝達物質の間の正準相関分析(CCA)。まず、データに対してデトレンドコレスポンデンス分析(DCA)を実施し、本研究の関連データのDCA1は3.7928であり、3より大きいため、CCAモデルを選択した。
フルサイズ表示
VitB12の添加は、門の構造に明らかな変化をもたらした。PM2.5グループと比較して、Actinobacteria、Cyanobacteria、Acidobacteria、Armatimonadetes、Planctomycetes、Fibrobacters、Verrucomicrobia、Errucomicrobia、Patescibacteria、Chloroflexi、Gemmatimonadetes、BRC1の相対量は、3つのMeCbl処理グループすべてで減少した(図S3)。まとめると、PM2.5投与群とVitB12添加群では、菌叢構造が有意に異なり、VitB12の添加時間と濃度が増加するにつれて、菌叢構造は同じ傾向で変化し、中でもVerrucomicrobiaは明確な勾配変化を示した。
属のヒートマップ(図5c)では、PM2.5-0グループと比較して、PM2.5曝露によりBacillus属、Streptomyces属、Glutamicibacter属、Ureibacillus属、Parapedobacter属などの細菌属の存在量が増加した。しかし、VitB12を添加すると、これらの属の存在量は減少した。これらの属の相対的な存在量は、VitB12の補充時間が長くなり、VitB12濃度が高くなるにつれて減少した。中濃度および低濃度のVitB12は、これらの属の存在量をPM2.5-0グループと同程度まで減少させたが、高濃度のVitB12を長期間補充すると、これらの属の存在量は減少した。一方、PM2.5に暴露されると、Subdoligranulum属、Faecalibacterium属、Bacteroides属、Anaeroglobus属、Parasutterella属、Ruminococcus_2属、Lachnoclostridium属、その他の属の存在量が減少し、VitB12の添加によって増加した。これらの属の相対的な存在量は、時間およびVitB12濃度とともに増加した。さらに、MeCblの摂取は、細菌のリボソームスイッチを介したVitB12制御の結果として、Escherichia-Shigellaを増加させ、Lactobacillusを減少させた。
大腸内腔における二次代謝経路のうち(図5d)、PM2.5は、神経変性疾患、シグナル伝達分子と相互作用、環境適応、神経系、アミノ酸代謝、膜輸送、他のアミノ酸の代謝、遺伝情報処理、シグナル伝達、細胞プロセス、シグナル伝達の代謝強度、エネルギー代謝を含むすべての二次代謝経路を減弱させ、これは粘膜でも観察された(図S2B)。VitB12は代謝経路の一部を強化したが、代謝強度を部分的に回復させただけで、PM2.5-0のレベルまで完全に回復させることはできなかった。4日目に、さらに細菌叢と管腔内の代謝産物との相関を解析した。図5eに示すように、コール酸は最も明らかに濃縮された物質であり、乳酸桿菌およびバチルス菌と最も正の相関があり、ビフィドバクテリウム、バクテロイデス、ルミノコッカス_2およびアナエログロバスと負の相関があった。正準相関分析(CCA)は、2つの変数間の関係を記述する統計分析である。ここでは、神経伝達物質濃度と腸内細菌叢量の相関をCCAに用いた。ほとんどの微生物叢は第3象限と第4象限にあったのに対し、神経伝達物質は第1象限、第2象限、第3象限に分布していた(図5f)。
PM2.5に曝された発酵における毒素濃度、酵素活性、VitB12含量の変化
リポ多糖(LPS)の蓄積はPM2.5によって加速され、MeCblの介入はこの変化を遅らせ、発酵1週間後には対照群よりもさらに低くなった(図6a)。志賀毒素含量の変動傾向は対照群と同様であったが、MeCbl群ではより低い値を示した(図6b)。これらの結果は、MeCbl摂取が腸内毒素含量を減少させ、疾病のリスクを低減できることを示唆している。プロテアーゼ活性は、PM2.5処理群では2日目に有意に低下し、MeCblの介入により、発酵2日目のプロテアーゼ活性は対照群と同レベルを維持した。発酵を継続すると、PM2.5処理群の酵素活性は徐々に正常値に戻った。このことは、MeCblが、環境刺激によって誘発された腸障害時のプロテアーゼ活性の代謝レベルの安定化に有益であることを示唆している(図6c)。PM2.5は発酵2日目にアミラーゼ活性を低下させたが、発酵の進行に伴い、MeCbl介入群のアミラーゼ活性は4~6日目に上昇した(図6d)。体内には、ミトコンドリア内のアデノコバラミンと細胞質内のMeCblの2つの主な活性型があり、これらはメチルマロン酸(MMA)とホモシステイン(Hcy)の恒常性維持に必要である。一方、シアノコバラミンは結晶化しやすく、空気で酸化されにくいため、VitB12の食品添加物として使用されることが多い。ヒドロキソコバラミンはシアノコバラミンよりも活性型に変換されやすく、シアン化物イオンとの親和性が強いため、シアン化物中毒の解毒剤としても利用されている。その結果、模擬腸内発酵ブロス中のVitB12の主要形態はアデノコバラミンであり、PM2.5の添加により発酵過程で減少した。一方、MeCblを添加すると、すべてのタイプのVitB12の含有量が大幅に増加した(図6e-h)。
図6. 異なる発酵期間におけるin vitro大腸シミュレーションシステムの毒素、酵素活性およびVitB12含量。(a)LPS含量、(b)志賀毒素含量、(c)プロテアーゼ活性、(d)アミラーゼ活性、(e-h)4種類のVitB12(シアノコバラミン、メチルコバラミン、アデノシルコバラミン、ヒドロキソコバラミン)含量の変化の折れ線グラフ。
図6. 異なる発酵期間におけるin vitro結腸シミュレーションシステムの毒素、酵素活性およびVitB12含量。(a)LPS含量、(b)志賀毒素含量、(c)プロテアーゼ活性、(d)アミラーゼ活性、(e-h)4種類のVitB12(シアノコバラミン、メチルコバラミン、アデノシルコバラミン、ヒドロキソコバラミン)含量の変化の折れ線グラフ。
フルサイズ表示
MeCblはPM2.5によるVitB12合成代謝輸送障害を回復させた
対照群とMeCbl群とは対照的に、PM2.5群のマイクロバイオームでは、ウロポルフィリノーゲンIIIからのコビリン酸合成の好気性経路に関与するCobC、CobG、CobS、CobT、およびCobNの発現が亢進した(図7a,c)。一方、対照群とMeCbl群のマイクロバイオームでは、コビリン酸の嫌気性経路とアデノシル-コバラミン合成の遺伝子発現が増加した(図7b,c)。さらに、微生物叢の変化と相まって、PM2.5曝露は、デノボ生合成やサルベージ経路に関係なくVitB12合成に関与するFaecalibacterium、Bacteroides、Ruminococcus_2、Lachnoclostridium、Citrobacter、Collinsella、Escherichia-Shigellaの明らかなダウンレギュレーションをもたらしたCitation38(図7d)。バチルス・メガテリウム(Bacillus megaterium)はVitB12産生菌であり、工業的応用の候補と考えられていることは注目に値する。引用39 ヒトにおけるこの細菌の存在量は非常に低く、VitB12産生に関するバチルス菌に関する他の報告はないことから、我々はバチルス菌をVitB12を合成できる菌として分類しなかった。以上の結果から、PM2.5の補給はコバラミンの嫌気性合成能力を著しく破壊し、アミノ酸や一部の神経伝達物質の代謝に関与するアデノシルコバラミン濃度を阻害することが示された。MeCblの添加は、上記の細菌の存在量を増加させるか、VitB12濃度を直接増加させることによって、このような障害を回復させる可能性がある。BtuBは大腸菌の外膜に存在する輸送タンパク質で、VitB12の吸収を担っていると考えられている。メタトランスクリプトームデータ解析によると、BtuB、TonB、ExbD、ExbBの相対量はPM2.5処理群で明らかに減少しており、微生物のVitB12輸送システムが損なわれていることを示している。ExbBレベルは、エネルギー供給に重要なMeCbl処理によって回復した(図7e,f)。
図7. KEGGパスウェイ解析によるin vitro結腸シミュレーション系(内腔)におけるコバラミン代謝および輸送の変化(4日目)。(a)PM2.5曝露により発現が増加したコバラミン代謝酵素の遺伝子、(b)PM2.5曝露により発現が減少したコバラミン代謝酵素の遺伝子、MeCbl溶液は最終濃度0.0125 mg/mL(MeCbl-L)および1.25 mg/mL(MeCbl-H)で発酵槽に添加した、(c)コバラミン代謝に関連する主要なパスウェイ。PM2.5処理後、発現が増加した遺伝子を赤で、発現が減少した遺伝子を青で示したが、MeCblの補充により、これらすべての遺伝子レベルが有意に逆転した;(d)細菌によるB12合成の主要経路のフローチャート。(e)大腸菌のVitB12輸送機構の図。(f)4日目の発酵におけるVitB12輸送に関与するBtuB、TonB、ExbD、ExbBの相対転写レベル。異なる文字のバーで示した値は有意に異なる(p < .05、一元配置分散分析)。
図7. KEGGパスウェイ解析による試験管内結腸シミュレーション系(内腔)におけるコバラミン代謝・輸送の変化(4日目)。(a)PM2.5暴露により発現が増加したコバラミン代謝酵素の遺伝子、(b)PM2.5暴露により発現が減少したコバラミン代謝酵素の遺伝子、MeCbl溶液は最終濃度0.0125 mg/mL(MeCbl-L)および1.25 mg/mL(MeCbl-H)で発酵槽に添加した、(c)コバラミン代謝に関連する主要なパスウェイ。PM2.5処理後、発現が増加した遺伝子を赤で、発現が減少した遺伝子を青で示したが、MeCblの補充により、これらすべての遺伝子レベルが有意に逆転した;(d)細菌によるB12合成の主要経路のフローチャート。(e)大腸菌のVitB12輸送機構の図。(f)4日目の発酵におけるVitB12輸送に関与するBtuB、TonB、ExbD、ExbBの相対転写レベル。異なる文字のバーで示した値は有意に異なる(p < .05、一元配置分散分析)。
フルサイズ表示
考察
我々の結果は、PM2.5への暴露が、腸内微生物のアンバランスと関連するVitB12代謝障害を引き起こすことを示している。重要なことは、本研究においてVitB12がPM2.5介入による新たな潜在的標的として同定されたことである。これまでの研究で、PM2.5への慢性的な曝露は、動物モデルおよび臨床研究の両方で認知機能の低下をもたらすことが示されているが、神経細胞実験では完全な答えは得られない。Citation42 PM2.5がヒトおよび動物モデルの両方で微生物叢の組成と存在量に影響を与える可能性があることは、一連の研究で証明されているが、微生物叢によって産生される神経伝達物質の交代に関する研究はない。本研究では、PM2.5が腸内細菌叢、代謝産物、有害マーカーに影響を及ぼすメカニズムをin vivoおよびin vitroで評価した。その結果、PM2.5が「腸脳軸」に基づく神経毒性に有効であることを示す確かな証拠が得られた。
線虫はPM2.5の毒性評価にうまく利用されており、重金属は潜在的にPM2.5の毒性に寄与している。引用44,引用45 この研究では、線虫の走化性行動の感受性と学習・記憶行動の障害はPM2.5によって著しく損傷され、MeCblの補充によって回復した。本研究では、GABAニューロン、コリン作動性ニューロン、グルタミン酸作動性ニューロンを緑色蛍光タンパク質で標識したトランスジェニック線虫を導入した。26個のGABA作動性ニューロンを持つトランスジェニック線虫EG1285では、GFPの発現が相対的に低かった。このことは、線虫においてPM2.5が抑制性および興奮性の神経伝達物質であるGABAの濃度を低下させ、線虫の体を曲げる活動につながっていることを意味している。このような同様の結果は他の研究者からも報告されており、Pb、Hg、Cu、Cr、Cd、Mnなどの重金属への曝露は、線虫のGABA作動性ニューロンの軸索変性や神経細胞喪失を引き起こす可能性がある。グルタミン酸は中枢神経系における重要な興奮性神経伝達物質である。脳内の過剰なグルタミン酸は、長期的な興奮性の亢進として現れ、脳細胞の損傷、不安、うつ病、その他の症状を引き起こす。同様に、アセチルコリン(Ach)は体壁や咽頭筋の刺激を制御するために不可欠であり、それゆえ運動や摂食に望まれる。
VitB12は、メチルマロニル・コエンザイムAミューターゼとメチオニン合成酵素の主要な補酵素であり、これらは一炭素サイクルと神経の正常な機能に不可欠である。興味深いことに、我々の研究では、これらの経路に関与する遺伝子は、線虫モデルにおけるPM2.5とMeCblの介入によって有意な変化は見られなかった。したがって、腸内微生物の制御がPM2.5誘発神経毒性に寄与していると考える理由がある。マウスモデルとin vitro微生物培養系を用いて、PM2.5、MeCbl、微生物ホメオスタシスの関係を調べた。Citation48。VitB12合成に関与するFaecalibacterium、Bacteroides、Ruminococcus_2、Lachnoclostridium、Citrobacter、Collinsella、Escherichia-Shigellaのレベルは、PM2.5による損傷後に劇的に低下した。 Citation49,Citation50その結果、PM2.5曝露はVitB12代謝経路の障害を引き起こし、神経伝達物質の異常分泌をもたらし、線虫の認知障害を引き起こすことがわかった。これらの異常な表現型は、MeCblの補充によって部分的に補うことができる。
SCFAは主に炭水化物(食物繊維)の細菌発酵によって産生され、様々なメカニズムを通して中枢プロセスに影響を与える。引用51,引用52 この研究では、MeCblは酪酸濃度を増加させ、その結果、Faecalibacterium、Ruminococcus_2、Anaeroglobusなどの酪酸産生細菌が増加した。ビフィズス菌は、直接酪酸を産生することはできないが、酪酸産生菌の増殖に適した環境を提供することができる。
我々のこれまでのマイクロバイオームおよびメタボローム解析データによると、PM2.5による神経代謝物質の異常分泌に対するMeCblの保護効果から、神経変性疾患に関連するシグナル伝達経路が大きな関心を集めている。この研究から、腸内細菌叢が宿主の神経伝達物質やその関連経路にも影響を及ぼし、その結果、宿主の生理や行動に影響を及ぼすことが明らかになった。腸内細菌によって分泌される神経伝達物質は、腸管神経系のシグナルをさらに制御する分子を放出するよう上皮細胞を誘導し、それによって脳の認知機能を制御することができる。Citation35 神経毒性の根本的な原因のひとつと考えられているグルタミン酸レベルの上昇が、マウスや試験管内微生物培養系にかかわらず、本研究のPM2.5曝露群で認められた。このことは、PM2.5曝露群におけるGlutamicibacterの増加がグルタミン酸蓄積の一因であることを示唆している。
代謝産物の数は交差摂食に依存しており、微生物の協力が腸内微生物の生態において主要な役割を果たしていると考えられる。PM2.5への長期曝露は、腸内微小環境のアンバランスに続き、VitB12、酪酸、神経伝達物質の異常産生を引き起こし、最終的に認知障害と神経毒性を引き起こした。MeCblを補給することで細菌叢のバランスが回復し、それによって関連代謝産物の濃度が正常に保たれ、結果として認知障害が改善される。微量栄養素を人間の健康、特に汚染物質が蓄積する環境に応用するためには、MeCblがPM2.5による障害を防ぐ有益な因子としてどのように働くかを理解することが重要である。わが国では環境の変化が激しいため、母集団データの収集が完了しておらず、本研究の結論をさらに検証するためには、PM2.5に長期暴露した母集団実験の関連データが不足している。
結論
まとめると、PM2.5曝露は、MeCblの補充によって回復しうる神経行動異常をもたらした。さらなる研究は、PM2.5が、バクテロイデス属、ビフィドバクテリウム属、ルミノコッカス属、およびフェカリバクテリウム属の変化率に関連する、VitB12の合成、代謝、および輸送の障害を引き起こしたという証拠を提供した。 MeCblの摂取は、関連する代謝の恒常性を維持することができる。さらに、MeCblは神経伝達物質の濃度を対照群のレベルまである程度回復させたが、これは神経伝達物質の合成に関与するだけでなく、乳酸桿菌やバクテロイデスなどの腸内細菌叢のバランスを維持することにも関係している。続いて、神経伝達物質の障害が緩和され、認知障害が寛解する。脂肪酸、ヒスチジン、フェニルアラニン代謝の経路もMeCblの改善効果に関与していた。今回の知見は、VitB12による腸内細菌叢の標的制御のための新たな手がかりを提供する可能性があり、長期にわたるPM2.5曝露による有害な影響が、食事構造を調整することによって腸内細菌叢を制御することで緩和されることを示唆している。
材料と方法
線虫の維持、同調および処理
線虫は、大腸菌OP50を含む標準線虫増殖培地(NGM)(3 g/L NaCl、2.5 g/L ペプトン、17 g/L 寒天、25 mM リン酸カリウム、1 mM CaCl2 .2 H2O、1 mM MgSO4 .7 H2O、12.9 mM コレステロール)で20℃で培養した。ほとんどのワムシがL4ステージに達した時点で、M9バッファーで回収し、同調によって卵を得るために溶解した。処理については、対照群を除き、同調したワムシをMeCbl(0.5μg/mL)の有無にかかわらずPM2.5(0.1mg/mL)に曝露した。72時間暴露後、運動性、動物の行動、蛍光強度などの指標を測定した。PM2.5サンプルはHuanhuan Wang(杭州師範大学、中国)の好意により提供された。無機成分では硫黄、カリウム、塩化物が多く、有機成分ではベンゾ(b)フルオランテン、インデノ(1,2,3-cd)ピレン、クリセンが多かった。
寿命アッセイ
同調させた若成虫を、NGM中で5-フルオロ-2′-デオキシ-β-ウリジン(5-FudR、50μg/mL)および異なる薬剤を含むOP50とともに寿命まで成長させた。各NGMで合計30~40匹の若成虫を培養し、1日おきにワムシを数えた。白金線で軽く触れても動かない虫は死んだとみなした。実験はすべて3回繰り返した。
運動アッセイ
同期の線虫(L4 期)を異なる処理下で NGM プレートに移した。3日目に実体顕微鏡で線虫を観察した。線虫の体の長軸に対する1波長の動きを体の曲がりとして記録した。線虫の頭部は左右に振れるので、回復した場合は頭部の振れとみなした。体曲げの回数は60秒以内に、頭部スイングの回数は20秒以内に、各群30匹以上の線虫でカウントした。
鍛造行動
大腸菌OP50を9cm寒天培地で中心から半径0.5cmの範囲で培養し、麻酔のために1M NaN3を1μL細菌コロニーに塗布した。次に、曝露したワムシをOP50の中心から4 cm離れた場所に置いた。各試験は試験培地に60匹のワムシを入れ、3回繰り返した。20℃で6時間培養後、OP50コロニーに到達した線虫の数を各プレートで数えた。線虫の到達レベルは、細菌コロニーに到達した線虫の数をプレート上の線虫の総数で割って算出した。
走化性(CTX)アッセイ
通常のCTXアッセイでは、5μLのにおい物質をA地点に滴下し、同じ溶媒をB地点に滴下した。記憶連想学習アッセイでは、中心から3cm離れた位置にNaCl寒天片(100mM)を置き、反対側の対応する位置をコントロールとした。培地はNaClの濃度勾配を得るために4℃で14時間保存した。アッセイの15分前にNaCl寒天を取り除き、プレートの中央に個体を配置した。培養4時間後、いずれかのスポットから半径2cm以内の動物を数え、対照スポットの動物数を差し引き、プレート上の動物の総数を除算して走化性指数(CI)を求めた。CIが陽性であれば塩への誘引を、陰性であれば嫌悪を示すことは文献に記載されている通りである。各濃度を3回反復した。
イメージングと蛍光定量
EG1285株[unc-47p:GFP+lin-15(+)]、DA1240株[eat-4:sGFP+lin-15(+)]、LX929株[unc-17:GFP]を別々に用いて、GABAニューロン、グルタミン酸作動性ニューロン、コリン作動性ニューロンを評価した。VL749[acdh-1p:GFP+unc-119(+)]ワームは内因性VitB12レベルを示すために用いた。同調させたミミズがL1期に達した時点で、対照群を除き、MeCbl(0.5μg/mL)添加/無添加のPM2.5(0.1mg/mL)に曝露した。72時間曝露後、無作為に選択した異なる処理群のワムシを、0.2%レバミソールを含むM9緩衝液を含むスライドに移し、固定曝露パラメータで蛍光顕微鏡を用いて捕獲した。蛍光強度の測定には1群につき少なくとも20匹のミミズを用い、各実験は3連で行った。蛍光はImage-Pro Plus 6.0を用いて定量化し、結果は対照動物の平均蛍光強度に対して正規化した相対蛍光単位で示した。
リアルタイム定量PCR(RT-qPCR)
同調させた野生型 N2 線虫を、若齢成虫期に MeCbl を含む PM2.5 または含まない PM2.5 に 72 時間曝露した後、すべての線虫を回収し、RNA 抽出後、市販キットの説明書に従って cDNA 合成を行った。qPCR は SYBR-Green PCR キット(Yeasen, Shanghai, China)を用いて行い、結果は 2-ΔΔCt 法で解析した。目的遺伝子の相対的mRNA発現は、act-1発現との関連で測定した。
マウスと処置
雄性C57BL/6野生型マウス(8週齢)48匹をShanghai Slack Laboratory Animal Co. 実験前に、マウスを25±1℃、湿度55±5%の環境下で7日間馴化させ(12時間明暗サイクル)、標準的な実験食と飲料水を与えた。マウスを無作為に3群に分け、順応期間後5ヶ月間飼育した。対照群の実験室はろ過空気(PM2.5含有量はゼロに等しい)で満たされ、PM2.5群とMeCbl群は環境PM2.5慢性リアルタイム暴露室で環境PM2.5に暴露された。MeCbl群には滅菌した0.125 mg/L MeCbl栄養液を毎日与え、他の群には滅菌した超純水を与えた。マウスは0日目と毎月体重を測定し、糞便を採取した。5ヵ月後、マウスを犠牲にし、結腸の長さを測定した。糞便はDNA抽出と短鎖脂肪酸測定のために-80℃で保存した。動物実験はThe Guide for Care and Use of Laboratory Animalsに従って行い、杭州師範大学医学部動物飼育使用委員会の承認を得た(承認番号:HSD20170301)。
試験管内大腸発酵シミュレーション
Citation50 チューブ接続後、各発酵槽に 300 mL の腸管培養液と 15 個の粘膜ビーズを添加した。糞便微生物は4つの発酵槽に10%ずつ接種した(個人差をなくすために3人のドナーが糞便を提供した)。腸内発酵システムのパラメーターを設定することにより、0.5M NaOH溶液と0.5M HClが自動的に添加されてpHが5.5に調整され、温度はヒトの上行結腸の発酵環境を模擬した37℃で一定に保たれた。嫌気的環境を維持するため、発酵タンクに窒素を送り込んだ。接種24時間後、毎日100mLの新しい腸内培養液を供給し、ヒトの摂取と消化をシミュレートするために、同容量のタンクから発酵を排出した。粘膜の再生をシミュレートするため、毎日5個の新しい粘膜ビーズを使用し、タンク内の古い粘膜ビーズと交換した。
糞便微生物を接種し、微生物構造を安定させるために約1週間連続発酵させた後、必要なサンプルを添加した。滅菌シリンジを用いて、100 mg/mL PM2.5溶液28.7 μLを、2 Lのフィーディングタンク内の培地に添加した。MeCbl溶液を最終濃度0.0125 mg/mL、0.125 mg/mL、1.25 mg/mLで発酵槽に添加し、7日後に発酵を停止した。各発酵槽の発酵ブロスと粘液球は、第2ステージの0~7日目に採取し、試料は-20℃で保存した。
消化酵素と毒素の検出
超音波細胞破砕機を用いて菌体を破砕し、機械の作動電力を450W、作動時間を1分、超音波時間とインターバル時間を5秒に設定した。破砕した菌液を8000rpm、4℃で10分間遠心分離し、上清を関連酵素と毒素の検出に用いた。
アミラーゼ活性はヨウ素-デンプン比色法を用いて検出し、プロテアーゼ活性はFolin-Lowry protein assayを用いて測定した。2種類の消化酵素の活性は、標準曲線の希釈比に基づいて算出した。
志賀毒素およびリポ多糖の検出には志賀毒素ELISA試験キットおよびLPS ELISA試験キットを用いた。標準曲線はキットに添付されている標準液を用いて作成した。標準曲線に従い、希釈率を用いて2つの毒素濃度を算出した。
コバラミン含量の測定
ヒドロキソコバラミン、シアノコバラミン、アデノコバラミン、メチルコバラミンの4種類のVitB12標準試料溶液を精密に測定・調製した。高速液体クロマトグラフィー(HPLC)を用いて試料の保持時間を測定し、標準曲線を作成した。
菌液8mLを計量し、8000rpm、4℃で10分間遠心分離した後、培地沈殿を除去し、上清を回収した。VitB12の分離精製は固相抽出で行った。固相抽出カラム(Sep-Pak C18、Waters)を活性化し、回収した上清と結合させた。純水でカラム5容量をすすいだ後、溶離液を3mLのメタノールで2回溶出し、回収した。メタノールは37℃で窒素ブロワーにより完全に蒸発させ、得られた固形物を1mLの超純水に入れ、試験した。
クロマトグラフィー分離はすべて室温で行った。1.0mL/minの流速で、超音波水浴を用いて脱気した0.1%ギ酸水溶液(A)と純アセトニトリル(B)を移動相として用い、グラジエント溶出は以下のようにプログラムした: 注入量は20μLで、カラム溶出液は361nmでモニターした。クロマトグラフィーカラムはVenusil XBP C18(4.5×250、5μm、Agela Technologies)を使用した。
メタトランスクリプトーム解析
模擬発酵6日目の大腸サンプル(5 mL)を採取し、800 rpm、4℃で5分間遠心した。底部の固形培地を捨て、上清を新しい遠心チューブに移した。この液体を6000rpm、4℃で10分間遠心し、上清を取り除き、沈殿物を除去した。サンプルは2mLのRNA固定液とともに-80℃で保存され、マクロトランスクリプトームシーケンスのために上海明治生物医学技術有限公司に送られた。
統計解析
すべてのデータは平均値±標準偏差(SD)で示した。SPSS 22.0を用い、一元配置分散分析(One-way ANOVA)とポストホック多重範囲有意差(Duncan)検定およびt検定を行い、統計的有意性を決定した。有意差は*p < .05および**p < .01で示した。OTU表に従って、植物相のα-多様性を推定するためにシャノン指数とシンプソン指数を計算し、シーケンスデータに基づいて主成分分析(PCA)を行った。相関ヒートマップは、MeV. 4-9-0.とOrigin8.0を用いて相関ヒストグラムを作成した。さらに、PICRUStを用いて16S rRNAシーケンスから得られた配列データを解析し、遺伝子機能と代謝経路を予測した。
データおよび材料の入手可能性
本研究で作成または解析されたすべてのデータは、この発表論文[および補足情報ファイル]に含まれているか、NCBI SRAリポジトリ、PRJNA889242で入手可能である。
補足資料
補足資料
MS Wordをダウンロード (1.9 MB)
情報開示
著者による潜在的な利益相反は報告されていない。
補足資料
本論文の補足データは、https://doi.org/10.1080/19490976.2023.2267186 からオンラインでアクセスできる。
追加情報
資金提供
本研究は、中国国家自然科学基金会(助成金番号32072230)、浙江省自然科学基金会(助成金番号LY21C200007)および浙江公館大学エリート基金(3090JYN9921001G)の助成を受けた。
参考文献
Craig L, Brook JR, Chiotti Q, Croes B, Gower S, Hedley A, Krewski D, Krupnick A, Krzyzanowski M, Moran MD, et al. 大気汚染と公衆衛生:リスク管理者のためのガイダンス文書。J Toxicol Env Heal A. 2008;71(9-10):588-21. doi:10.1080/15287390801997732. [テーラー&フランシスオンライン] [PubMed] [Web of Science ®], [Google Scholar].
中国北部の都市周辺部から飛来する冬季の多環芳香族炭化水素結合粒子状物質が肺がん細胞の転移を促進する。Environ Sci Technol. 2015;49(24):14484–14493. doi:10.1021/es506280c. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Eze IC, Hemkens LG, Bucher HC, Hoffmann B, Schindler C, Künzli N, Schikowski T, Probst-Hensch NM. 欧州と北米における環境大気汚染と糖尿病の関連:系統的レビューとメタ分析。Environ Health Perspect. 2015;123(5):381–389. doi:10.1289/ehp.1307823. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
屋外での粒子状物質曝露と肺がん:系統的レビューとメタアナリシス。Environ Health Perspect. 2014;122(9):906–911. doi:10.1289/ehp/1408092. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Brook RD, Rajagopalan S, Pope CA, Brook JR, Bhatnagar A, Diez-Roux AV, Holguin F, Hong Y, Luepker RV, Mittleman MA, et al. 粒子状物質による大気汚染と心血管疾患。Circulation. 2010;121(21):2331–2378. doi:10.1161/CIR.0b013e3181dbece1. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
屋外大気汚染と小児および成人の喘息による救急外来受診:カナダ、アルバータ州北部におけるケースクロスオーバー研究。Environ Health. 2007;6(1):40. doi:10.1186/1476-069X-6-40. [Crossref] [PubMed], [Google Scholar].
アラウージョJA。超微粒子は心血管疾患の危険因子ですか?Rev Esp Cardiol。2011;64(8):642–645. doi:10.1016/j.recesp.2011.05.002. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Ailshire JA, Clarke P. Fine particulate matter air pollution and cognitive function among U.S. older adults. J Gerontol B. 2015;70(2):322-328. doi:10.1093/geronb/gbu064. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Gatto NM, Henderson VW, Hodis HN, St. John JA, Lurmann F, Chen J, Mack WJ. ロサンゼルスの中高年における大気汚染の成分と認知機能。Neurotoxicology. 2014;40:1–7. doi:10.1016/j.neuro.2013.09.004. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Thiankhaw K, Chattipakorn N, Chattipakorn SC. PM2.5曝露とAD関連神経病理および認知アウトカムとの関連。Environ Pollut. 2022年;292(Pt A):118320. doi:10.1016/j.envpol.2021.118320. [Crossref] [PubMed], [Google Scholar].
空気中の微粒子の神経毒性:分子事象からヒト疾患まで。J Hazard Mater. 2023;457:131827. doi:10.1016/j.jhazmat.2023.131827. [クロスリフ] [PubMed] [Web of Science ®], [Google Scholar].
PM2.5中の有機抽出物はSH-SY5Y細胞のフェロプターシスを誘導する主要なトリガーである。Ecotoxicol Environ Saf. 2023;249:114350. doi:10.1016/j.ecoenv.2022.114350. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
PM2.5曝露は、神経細胞におけるオートファジー機能障害/p62-KEAP1-NRF2の活性化を介して、リソソーム膜透過による細胞死を誘発し、フェロプシス不感受性をもたらす。Ecotoxicol Environ Saf. 2022;248:114333. doi:10.1016/j.ecoenv.2022.114333. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
基本的なM、Dardevet D、Abuja PM、Bolsega S、Bornes S、Caesar R、Calabrese FM、Collino M、De Angelis M、Gérard P、ら。代謝および免疫疾患におけるマイクロバイオームの関連性が因果関係を反映しているかどうかを識別するためのアプローチ。Gut Microbes. 2022;14(1):2107386. doi:10.1080/19490976.2022.2107386. [Taylor & Francis Online] [PubMed] [Web of Science ®], [Google Scholar].
Dai、Wang Z、Yang Y、Du P、Li X. PM2.5によるマウスの体重減少は腸内微小環境の変化による:粘液バリア、腸内細菌叢、代謝プロファイリング。J Hazard Mater. 2022;431:128653. doi:10.1016/j.jhazmat.2022.128653. [クロスリフ] [PubMed] [Web of Science ®], [Google Scholar].
Mutlu EA, Comba IY, Cho T, Engen PA, Yazıcı C, Soberanes S, Hamanaka RB, Niğdelioğlu R, Meliton AY, Ghio AJ, et al. Inhalational exposure to particulate matter air pollution alters the composition of the gut microbiome. Environ Pollut. 2018;240:817–830. doi:10.1016/j.envpol.2018.04.130. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
LiS、Guo B、Dong K、Huang S、Wu J、Zhou H、Wu K、Han X、Liang X、Pei X、et al.環境PM2.5およびその成分への長期曝露と腸内細菌叢との関連:中国コホートからの証拠。Sci Total Environ. 2023;884:163577. doi:10.1016/j.scitotenv.2023.163577. [Crossref][PubMed][ウェブ・オブ・サイエンス®], [Google Scholar].
秦J、王J.粒子状物質曝露による肺障害に対する腸内細菌叢の影響に関する研究の進展。環境研究2023;233:116162.doi:10.1016/j.envres.2023.116162. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
スミスAD、ウォーレンMJ、レフサムH.第6章ビタミンB12。で: エスキンNAM、編集者。Advances in food and nutrition research. Vol.2018。Academic Press; 2018. p. 215-279. doi:10.1016/bs.afnr.2017.11.005. [Crossref], [Google Scholar].
Mccarter SJ、Coon EA、Savica R、St Louis EK、Bower JH、Benarroch EE、Sandroni P、Low P、Singer W.多系統萎縮症診断時のビタミンB12値の低下は、生存期間の短縮と関連する。Mov Disord. 2020;35(8):1462–1466. doi:10.1002/mds.28070. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Mccarter SJ, Teigen LM, Mccarter AR, Benarroch EE, St LE, Savica R. Low Vitamin B12 and Parkinson disease: potential link to reduced cholinergic transmission and severity of disease. Mayo Clin Proc. 2019;94(5):757-762. doi:10.1016/j.mayocp.2019.01.039. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
メチルコバラミンがPM2.5曝露による神経細胞アポトーシスと認知機能低下を緩和することを明らかにした。J Alzheimer Dis. 2022;86(4):1783–1796. doi:10.3233/JAD-215384. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
を用いた。C.エレガンスとその細菌食は、宿主と微生物叢の相互作用をシステムレベルで理解するためのモデルである。Curr Opin Biotechnol. 2017;46:74–80. doi:10.1016/j.copbio.2017.01.008. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
C. elegans: a biosensor for host-microbe interactions. Lab Anim (NY). 2021;50(5):127–135. doi:10.1038/s41684-021-00724-z. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
また、「哺乳類が哺乳類に感染すると、哺乳類が哺乳類に感染する可能性がある」ことを明らかにした。Chemosphere. 2017;181:55–62. doi:10.1016/j.chemosphere.2017.04.068. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
LDL受容体モチーフを持つ分泌タンパク質HEN-1は線虫の感覚統合と学習を制御する。Cell. 2002;109(5):639–649. doi:10.1016/S0092-8674(02)00748-1. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
ソアレスMV、チャランMF、ジャックMT、ドスサントスALA、ルシェセC、ピントンS、アビラDS。空気中のトルエン暴露は、線虫の神経行動学的変化を促進する生殖細胞系列のアポトーシスと神経細胞損傷を引き起こす。Environ Pollut. 2020;256:113406. doi:10.1016/j.envpol.2019.113406. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
MPAでキャップしたCdTe量子ドットの曝露は、グルタミン酸、セロトニン、ドーパミンのトランスポーターと受容体に遺伝子レベルで影響を与えるか、活性酸素を増加させるか、あるいはその両方によって、線虫Caenorhabditis elegansの神経毒性作用を引き起こす。Nanoscale. 2015;7(48):20460–20473. doi:10.1039/C5NR05914C. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Lam AB, Kervin K, Tanis JE. ビタミンB12は、メチオニン/S-アデノシルメチオニンサイクルを制御することで、アミロイドβ誘導性タンパク質毒性に影響を与える。この論文では、ビタミンB12がアミロイドβに及ぼす影響について、メチオニンとアデノシルメチオニンのサイクルを制御することで明らかにした。 [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
(注1)本論文は、本論文の一部である。線虫のメチオニン/S-アデノシルメチオニンサイクルの活性は、核ホルモン受容体によって感知され調整される。Elife. 2020;9:e60259. doi:10.7554/eLife.60259. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
秦 S、王 Y、李 L、劉 J、蕭 C、段 D、郝 W、秦 C、陳 J、姚 L、他。線虫における生殖成功を確実にするために、早生期のビタミンB12がSBP-1/SREBP1を介して脂質過酸化を制御する。このような遺伝子発現は、生物学的に重要な役割を担っている。 [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
Strandwitz P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018;1693:128-133. doi:10.1016/j.brainres.2018.03.015. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
神経性食欲不振症における微生物叢-治療の可能性。2022;1-51. doi:10.1017/S0954422422000130. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
腸内細菌叢と脳とのコミュニケーションにおける短鎖脂肪酸の役割。Nat Rev Gastroenterol Hepatol. 2019;16(8):461–478. doi:10.1038/s41575-019-0157-3. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Chen Y, Xu J, Chen Y. 腸内細菌叢による神経伝達物質の制御と神経疾患における認知への影響。Nutrients. 2021;13(6):2099. doi:10.3390/nu13062099. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
コバラミンはAcetobacter pasteurianus DSM 3509によって生産される。Appl Microbiol Biotechnol. 2019;103(9):3875–3885. doi:10.1007/s00253-019-09704-3. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
シアノコバラミンとメチルコバラミンが炎症性腸疾患と腸内細菌叢組成に及ぼす影響。J Agric Food Chem. 2019;67(3):916–926. doi:10.1021/acs.jafc.8b05730. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Guetterman HM, Huey SL, Knight R, Fox AM, Mehta S, Finkelstein JL. ビタミンB-12と消化管マイクロバイオーム:システマティックレビュー。2022;13(2):530から558まで。 [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Bacillus megateriumによるビタミンB12生産のための2段階培養戦略の開発。Microb Cell Fact. 2014;13(1):102. doi:10.1186/s12934-014-0102-7. [Crossref] [PubMed], [Google Scholar].
Yao L, Wang Y, Qin S, Zhu S, Wu L. 抗糖尿病薬メトホルミンは、細菌がRcdAを介して環境からビタミンB12をハイジャックするのを助ける。また、メトホルミンは、ビタミンB12を環境からRcdAを介してハイジャックする。
Nilaweera TD, Nyenhuis DA, Cafiso DS. 無傷の大腸菌でのみ観察された構造中間体は、TonB依存性輸送のメカニズムを示している。Elife. 2021;10:e68548. doi:10.7554/eLife.68548. [この論文では、TonB依存的な輸送機構を明らかにした。
Thiankhaw K, Chattipakorn N, Chattipakorn SC. PM2.5曝露とAD関連神経病理および認知アウトカムとの関連。Environ Pollut. 2022;292:118320. doi:10.1016/j.envpol.2021.118320. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
ベイリーMJ、ホルツハウゼンEA、モーガンZEM、ナイクN、シェイファーJP、リャンD、チャンHH、サルナットJ、サンS、バーガーPK、ら。 周囲大気汚染物質への出生後の暴露は、生後6ヶ月の乳児の腸内細菌叢の組成と関連している。Gut Microbes. 2022;14(1):2105096. doi:10.1080/19490976.2022.2105096. [テーラー&フランシスオンライン] [PubMed] [Web of Science ®], [Google Scholar].
また、このような環境下での生物学的・生物学的研究は、生物学的・生物学的研究のみならず、生物学的・生物学的研究にも応用可能である。Chemosphere. 2016;144:2392–2400. doi:10.1016/j.chemosphere.2015.11.028. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Yilmaz LS, Walhout AJM. Worms, bacteria, and micronutrients: an elegant model of our diet. Trends Genet. 2014;30(11):496–503. doi:10.1016/j.tig.2014.07.010. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
線虫Caenorhabditis elegansのGABA作動性神経系に対する重金属曝露の神経毒性効果. Environ Toxicol Pharmacol. 2009;27(3):314–320. doi:10.1016/j.etap.2008.11.011. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
線虫におけるコリン作動性伝達:ACh活性化イオンチャネルの機能、多様性、成熟。J Neurochem. 2021;158(6):1274–1291. doi:10.1111/jnc.15164. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
虚血性脳卒中の病態生理におけるビタミンB12と腸脳恒常性。EBioMedicine. 2021;73:103676. doi:10.1016/j.ebiom.2021.103676. [クロスリファレンス] [PubMed] [Web of Science ®], [Google Scholar].
シアノコバラミンとメチルコバラミンが炎症性腸疾患と腸内細菌叢組成に及ぼす影響。J Agric Food Chem. 2019;67(3):916–926. doi:10.1021/acs.jafc.8b05730. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
コバラミン(ビタミンB12)はin vitro大腸シミュレーションにおいて微生物組成と代謝活性の変化を誘発した。Front Microbiol. 2018;9:2780. doi:10.3389/fmicb.2018.02780. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
Yan F, Xiang S, Shi L, Zhu X. Synthesis of queuine by colonic gut microbiome via crossfeeding. Food Front. 2023. doi:10.1002/fft2.307. [Crossref], [Google Scholar].
O'Riordan KJ, Collins MK, Moloney GM, Knox EG, Aburto MR, Fülling C, Morley SJ, Clarke G, Schellekens H, Cryan JF, et al. 短鎖脂肪酸:腸脳軸シグナル伝達のための微生物代謝産物。Mol Cell Endocrinol. 2022;546:111572. doi:10.1016/j.mce.2022.111572. [Crossref][PubMed][ウェブオブサイエンス®], [Google Scholar].
PM2.5慢性暴露によるマウスの認知機能低下:肺炎から神経細胞炎症まで。また、PM2.5による認知機能低下は、肺炎から神経細胞炎症に至る。 [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
フモニシンB1による線虫のGABAとセロトニンの神経伝達異常。Environ Pollut. 2022;304:119141. doi:10.1016/j.envpol.2022.119141. [Crossref] [PubMed] [Web of Science ®], [Google Scholar].
PDFダウンロード
関連研究
おすすめ記事
こんな記事も読まれています
引用元
情報提供者
著者
研究開発関係者
編集者
図書館員
学会
オープンアクセス
概要
オープンジャーナル
オープンセレクト
Dove Medical Press
F1000Research
機会
別刷りとe-prints
広告ソリューション
早期出版
企業アクセスソリューション
ヘルプと情報
ヘルプ&コンタクト
ニュースルーム
すべてのジャーナル
書籍
最新情報
ご登録いただくと、個別のリサーチやリソースをEメールでお届けします。
登録する
テイラー・アンド・フランシス・グループのFacebookページテイラー・アンド・フランシス・グループのTwitterページテイラー・アンド・フランシス・グループのLinkedinページ
テイラー・アンド・フランシス・グループYoutubeページテイラー・アンド・フランシス・グループWeiboページ
著作権 © 2023 Informa UK Limited
プライバシーポリシー
クッキー
利用規約
アクセシビリティ
イングランド&ウェールズ登録第3099067号
5 ハウィック・プレイス|ロンドン|SW1P 1WG
テイラー・アンド・フランシス・グループ
AcceptCookieポリシー
当社は、お客様のウェブサイト体験を向上させるためにクッキーを使用しています。当社のクッキーの使用およびクッキーの設定を管理する方法については、当社のクッキーポリシーをご覧ください。このメッセージを閉じることで、クッキーの使用に同意したことになります。
×
この記事が気に入ったらサポートをしてみませんか?