真菌感染症: 病因、抗真菌薬、代替治療アプローチ

微生物科学における最新の研究
第3巻 2022年 100137号
真菌感染症: 病因、抗真菌薬、代替治療アプローチ
https://www.sciencedirect.com/science/article/pii/S2666517422000347






著者リンク オーバーレイパネルを開くG.Kiran Kumar Reddy a b 1, Alwar Ramanujam Padmavathi a 1, Y.V. Nancharaiah a b
もっと見る
概要
共有
引用
https://doi.org/10.1016/j.crmicr.2022.100137Get 権利と内容
クリエイティブ・コモンズ・ライセンス
オープンアクセス
要旨
近年、真菌感染症が増加しており、早急な対応が必要である。真菌感染症は初期段階で診断されることが少ないため、感染症の重症度が増し、治療手順が複雑になる。真菌病原体は宿主の免疫系を回避し、感染症を重症化させるために様々なメカニズムを用いている。多様な表在性・全身性感染症の治療には、利用可能なレパートリーの中から抗真菌薬が投与される。しかし、ほとんどの抗真菌薬に対する真菌の耐性が証明されているため、疾患の制御が困難であり、抗真菌療法に課題を投げかけている。真菌の耐性化は、抗真菌剤の存在下でのいくつかの生理的適応と遺伝的変異、それに続く淘汰によって引き起こされる。限られた抗真菌薬しか利用できないこと、耐性菌の出現やバイオフィルムがもたらす耐性によって、真菌症に対するより良い治療効果を得るためには、新規薬剤の開発や代替アプローチの必要性が高まっている。この総説では、様々な真菌感染症やその原因菌、病態、様々な抗真菌薬、そして宿主の免疫反応や回避戦略を含む耐性メカニズムについて明らかにする。ここでは、真菌感染症に対する新規抗真菌薬やその他の代替的アプローチに関する最近の開発に焦点を当てた。
図解抄録
ダウンロード 高解像度画像のダウンロード (164KB)
ダウンロード フルサイズ画像のダウンロード
前の記事
次の記事
キーワード
抗真菌剤耐性
代替抗真菌療法
バイオフィルム
真菌病原体
宿主免疫反応
略語
ABC
ATP結合カセット
APC
抗原提示細胞
BBB
血液脳関門
BAD1
ブラストマイセスアドヘシン-1
CDC
疾病管理予防センター
中枢神経系
中枢神経系
DC
樹状細胞
ERG
ETS関連遺伝子
ETS
赤芽球形質転換特異的
FCY
フルオロシトシンデアミナーゼ
FDA
食品医薬品局
G-CSF
顆粒球コロニー刺激因子
GM-CSF
顆粒球マクロファージコロニー刺激因子
HAI
医療関連感染
HIV
ヒト免疫不全ウイルス
Hsp
ヒートショックプロテイン
IFN
インターフェロン
MFS
主要ファシリテーター・スーパーファミリー
M-CSF
マクロファージ-コロニー刺激因子
NK
ナチュラルキラー
PRR
パターン認識受容体
PAMP
病原体関連分子パターン
活性酸素
活性酸素種
RNS
活性窒素種
TLR
Toll-Like受容体
はじめに
真菌感染症は公衆衛生上重大な問題である。Covid-19を含む他の疾患の患者における真菌感染症の発生率は、生命を脅かす真菌症と死亡率に関連している。真菌感染症には、表面感染、皮膚感染、皮下感染、粘膜感染、全身感染などがあり、その重症度はさまざまである。カンジダ属のような生物はヒトの微生物叢の一部であり、個人における日和見感染症や、HIV患者、化学療法を受けている癌患者、免疫抑制剤投与を受けている患者のような免疫不全患者における生命を脅かす感染症(侵襲性カンジダ症)を引き起こす可能性がある。日和見感染や全身感染に加えて、カンジダ、アスペルギルス、フザリウム、ムコール、カビなどの真菌病原体は、基礎疾患を有する患者において医療関連感染(HAI)を引き起こす可能性がある(Perlroth et al.) 特定の地域では、真菌病原体が、ブラストミセス症、コクシジオドミセス症、ヒストプラスマ症、タラロミセス症、パラコクシジオドミセス症、スポロトリコーシスなど、生命を脅かす流行性真菌症を引き起こしている(Lee and Lau, 2017)。
全身性真菌症は、最近死亡率が増加していると診断されることが多い。米国疾病予防管理センター(CDC)は、2021年9月20日から24日までを真菌症啓発週間と定め、真菌感染症の早期診断の重要性を啓蒙し、その衰弱を緩和することを強調している(CDCウェブサイト)。本稿では、ヒトにおける真菌感染症のスペクトル、病因、免疫回避メカニズム、抗真菌薬とその作用機序、耐性メカニズム、真菌感染症と闘うための代替抗真菌アプローチについて概説する。真菌病原体と感染経路
子嚢菌門と担子菌門からなる真菌亜界Dikaryaは、ヒトにおけるすべての真菌病原体および感染症の主要な原因である。子のう菌類は、口腔咽頭、耳咽頭、皮膚、眼科、神経、泌尿生殖器、心臓、肺、および全身性の感染症を引き起こすことで知られている(図1)。クリプトコッカス(Cryptococcus)やマラセチア(Malassezia)といった担子菌門の菌は、それぞれ侵襲性髄膜炎や表在性皮膚感染症でよく知られている。
ダウンロード 高解像度画像ダウンロード(816KB)
ダウンロード フルサイズ画像のダウンロード
図1. ヒトにおける真菌感染症とその原因菌のスペクトル。
真菌の感染経路は主に直接接触や吸入である。Microsporum属、Epidermophyton属、Trichophyton属、Sporothrix属、Malassezia属に属する皮膚糸状菌は、損傷した皮膚に直接接触することによって感染する(図2A)。その他の主な感染経路は、胞子や分生子の吸入による肺感染である。ブラストミセス・デルマティディス(ブラストミセス症)、パラコクシジオイデス・ブラジリエンシスおよびパラコクシジオイデス・ルツィイ(パラコクシジオイデス症)、ヒストプラズマ・カプスラタム(ヒストプラズマ症)、ニューモシスチス・ジロベシイ(ニューモシスチス・プヌエモニア症)、アスペルギルス・フミガータスおよびA. flavus(アスペルギルス症)、Coccidioides immitisおよびC. posadasii(コクシジオイデス症)、C. neoformansおよびC. gattii(クリプトコッカス症)は主に吸入によって感染する。一方、Talaromyces marneffei(タラロマイセス・マルネフェイ)(タラロマイセス症)は、直接接触と吸入の両方の経路で感染する。
ダウンロード 高解像度画像のダウンロード (808KB)
ダウンロード フルサイズ画像のダウンロード
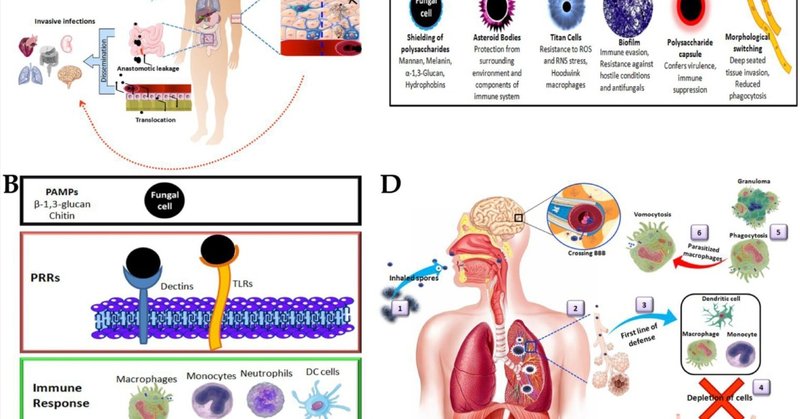
図2. A. 真菌病原体の侵入経路。B. 宿主免疫系における病原体の認識と反応。C. 免疫回避のための真菌細胞の形態変化。D. 侵入性真菌感染症の肺感染と病態。1. 1.胞子または分生子の吸入、2.肺胞への侵入、3.第一次防御ラインの誘導、4.貪食細胞の枯渇による肺結節や肺炎の発症、5.マクロファージによる真菌細胞の貪食、またはカプセル化、肉芽腫の形成、6.真菌細胞のマクロファージへの寄生による無傷の真菌の嘔吐、血流への循環、血液脳関門の通過による全身感染。宿主免疫系と真菌の免疫回避の武器
宿主免疫系にはデクチンやToll様受容体(TLR)のようなパターン認識受容体(PRR)が存在し、真菌細胞壁のキチンやβ-1,3-グルカンのような病原体関連分子パターン(PAMP)を認識する。このPRRによるPAMPの認識は、貪食、活性酸素種(ROS)、活性窒素種(RNS)の呼吸バースト、サイトカインやケモカインの産生を含む免疫応答の生成につながるシグナル伝達経路を引き起こし、真菌病原体を中和するための強い炎症反応を引き起こす(図2B)。単球、マクロファージ、樹状細胞(DC)、好中球は、自然免疫反応における防御の第一線として働く。真菌細胞はファゴライソソームで排除され、マクロファージに囲まれた肉芽腫として封じ込められる。単球はケモカインやサイトカインを分泌し、さらにTリンパ球への抗原提示細胞(APC)として働き、真菌細胞の除去に役立つ適応免疫応答を引き起こす。真菌病原体は、宿主の免疫系を欺いて増殖するために、様々な生存メカニズムに従っている(図2C)。真菌細胞は、PRRによる認識を防ぐために、高度に保存されたPAMPsを遮蔽するように表面層を変化させる(Campuzano et al.) P. jiroveciiは、表面のβ-1,3-グルカンを覆う主要な表面糖タンパク質を持ち、細胞壁にはキチンが欠如している(Ma et al.) 真菌のマンナン、メラニン、α-1,3-グルカン、ハイドロフォビン(Aspergillus RodA)層は、PAMPsとして働く表面多糖を遮蔽し、それによって宿主免疫系を回避する(Hernández-Chávez et al.) H.capsulatumの表面に露出したβ-1,3-グルカンセグメントは、エンド型β-1,3-グルカナーゼによってトリミングされ、病原体認識の低下と炎症性サイトカインの刺激を引き起こした(Garfoot et al.) C. neoformansの莢膜多糖体は、表面の遮蔽以外にも病原性を付与し、T細胞の活性化や好中球の動員にも悪影響を及ぼす。真菌病原体は、免疫認識から逃れるために形態学的に異なる構造を形成する。例えば、C. neoformansはチタン細胞を形成し(Zaragoza, 2011)、S. schenckiiは小惑星を形成する(Rosa et al.) 真菌病原体の中には、病原性と免疫回避のために二型成長を適応させるものがある。例えば、カンジダ属は酵母の形態で増殖し、病気の発症と進行に役立つ糸状菌糸を形成するように変化する。この形態転換は、貪食細胞の膜を破裂させるだけでなく、組織への侵入を増加させ、貪食作用を低下させるという利点がある(Marcos et al.) C. immitisとC. posadasiiは、関節子実体から球状体への形態変化を起こすことがあり、この形態変化は外細胞壁の糖タンパク質とアルギナーゼやウレアーゼなどの酵素の産生に関連している(Diep and Hoyer, 2020)。球状体は宿主における真菌の生殖形態であり、貪食やRNSに対する抵抗性を示す。
一部の真菌はマクロファージに寄生し、貪食から逃れることができる(Zhi et al.) 例えば、C. neoformansは肺に感染し、肺胞マクロファージを枯渇させる(図2D)。マクロファージに寄生して嘔吐を引き起こし、無傷の真菌細胞を放出して血液脳関門(BBB)を通過し、脳、脳脊髄液、中枢神経系(CNS)に影響を及ぼし、クリプトコッカス髄膜炎を発症する(Elsegeiny et al.) B. dermatitidisの芽胞も同様の方法で免疫系を回避し、リンパ系や血液中を循環する。ブラストマイセス・アドヘシン-1(BAD1)タンパク質は補体カスケードを不活性化し、全身感染を拡大させる。全体的な免疫回避は、抗貪食作用、T細胞増殖の抑制、炎症性サイトカインの抑制、酸化的ストレスやニトロソ化ストレスに対する抵抗性といった利点をもたらす。
免疫回避に加え、真菌病原体(例えば、C. albicans、C. neoformans、H. casulatum、P. brasiliensis、A. fumigatus)におけるバイオフィルム形成は、過酷な環境下で増殖し、抗真菌治療を回避するという重大な利点をもたらす。抗真菌剤とその作用機序
現在、アゾール系、ポリエン系、エキノカンディン系、アリルアミン系、ピリミジンアナログ系など5つの一般的なクラスの抗真菌薬が、表在性および全身性の抗真菌療法に使用可能である(Hokken et al.) これらの薬剤の作用機序と潜在的な細胞標的を図3に示す。これらのうち、イミダゾール系(ミコナゾール、ケトコナゾール)およびトリアゾール系(フルコナゾール、ボリコナゾール)を含むアゾール系薬剤は、多数の抗真菌化合物を臨床使用可能にする上で最も成功したバックボーンである。これらの薬剤は、カンジダ属やその他の真菌病原体に対して有効であり、さまざまな経路から投与できる柔軟性が魅力である(Nett and Andes 2015)。アゾール系抗真菌薬は、ステロール生合成に不可欠な酵素であるステロール14α-デメチラーゼを阻害し、ラノステロールを真菌細胞膜の安定性と流動性の維持に不可欠な成分であるエルゴステロールに変換する。一方、アムホテリシンBやナイスタチンなどのポリエン系抗真菌剤は、疎水性相互作用によって真菌細胞膜に作用し、膜ステロールを封鎖することで膜孔を形成し、細胞死を引き起こす。半合成リポペプチドであるエキノカンディン系薬剤(カスポファンギン、ミカファンギン、アニデュラファンギン)は真菌の細胞壁合成を阻害し、カンジダ属やアスペルギルス属に活性を示す。 これらの薬剤は、様々な真菌の重要な細胞壁成分(1,3-β-d-グルカン)の合成に不可欠なFKSファミリー遺伝子がコードする1,3-β-d-グルカン合成酵素を阻害し、殺菌活性を示す。アリルアミン系薬剤は、エルゴステロールの生合成を阻害することで真菌の増殖を抑制し、表在性皮膚糸状菌症の治療に一般的に推奨されている(Newland and Abdel-Rahman 2009)。これらの薬剤には、スクアレンをラノステロールに変換するスクアレンエポキシダーゼを阻害するテルビナフィンやナフチフィンが含まれる。ピリミジンアナログ(5-フルオロシトシン、5-FC)などの薬剤は、カンジダ属やクリプトコッカス属に対して活性がある。5-FCはシトシンパーミアーゼを介して細胞内に入り、核酸合成(DNAとRNAの両方)に影響を与え、最終的にはタンパク質合成を阻害する5-フルオロウラシルに脱アミノ化される。これらの主要なメカニズムとは別に、アムホテリシンBとミコナゾールは酸化ストレスを誘導し、抗真菌活性を増強することが報告されている。さらに、ソルダリンによるタンパク質合成の阻害、グリセオフルビンによる微小管集合の阻害、トリフェニルエチレンによるカルシニューリンシグナル伝達の阻害(Scorzoni et al.
ダウンロード 高解像度画像のダウンロード(870KB)
ダウンロード フルサイズ画像のダウンロード
図3. 抗真菌薬の作用機序を含む既存および新規の細胞標的。抗真菌剤耐性:抗真菌療法における障壁
使用可能な薬剤に対する真菌病原体の耐性化は、抗真菌療法における新たな臨床問題である(Berman and Krysen 2020)。耐性菌の発生には、宿主、真菌、環境要因のいくつかが影響している。適応的な表現型の可塑性、選択に続く標的遺伝子の変異、染色体異数性、有性生殖、水平遺伝子移動が抗真菌剤耐性の出現の原動力である(Hokken et al.) 真菌がもたらす主な耐性メカニズムを図4にまとめた。医療用インプラントや組織上でのカンジダ属菌によるバイオフィルム形成は、抗真菌薬に対してより高い耐性を付与する(Silva et al.) 真菌のバイオフィルムは、細胞外高分子物質マトリックス、遅い増殖速度、パーシスター細胞の存在、遺伝物質の交換、バイオフィルム細胞内の相乗的相互作用などいくつかの要因により、薬剤に対して高い耐性を示す(Silva et al.) アゾール系薬剤の使用は、耐性発現の多さから魅力的ではなくなりつつある。耐性メカニズムには、ABC(ATP結合カセット)トランスポーター、MFS(major facilitator superfamily)クラスの排出ポンプの過剰発現、ERG11またはCyp51遺伝子の変異によるアゾール系薬剤に対する親和性が低い/ない標的酵素の変化、ERG11酵素の過剰発現などがある(Berman and Krysen 2020)。ミトコンドリアの機能不全とストレスシグナルの活性化も、アゾール系薬剤に対する耐性の発現に寄与する可能性がある(Scorzoni et al.) ポリエン系抗真菌薬は数十年にわたり使用されているが、他の抗真菌薬に比べ耐性化は非常に少ない。ERG3の機能喪失につながる変異は、エルゴステロール含量の低下をもたらす。エルゴステロール含量の低下と、アムホテリシンBとの親和性が低い変化型ステロールの取り込みが、カンジダ属およびアスペルギルス属におけるポリエン系薬剤耐性の原因である(Scorzoni et al.) 5-FCに対する真菌耐性は、シトシンパーミアーゼ(Fcy2)および/またはシトシンデアミナーゼ(Fcy1)、それぞれ5-FCの細胞内への侵入と脱アミノ化を制限するFur1酵素の変異によって引き起こされる(Hokken et al.) スクアレンエポキシダーゼをコードするErg1遺伝子の変異はポリエンとの結合を阻害し、耐性化を引き起こす。FKS酵素(グルカン合成酵素)のようなエキノカンジン薬物標的の変異は、コンフォメーションを変化させ、それによって結合親和性を低下させ、耐性を増強する(Scorzoniら、2017)。遺伝子レベルでの耐性メカニズムの大部分は、抗真菌剤の標的抗真菌酵素/タンパク質をコードする遺伝子の点変異または過剰発現に由来する。図5
ダウンロード 高解像度画像ダウンロード(2MB)
ダウンロード フルサイズ画像のダウンロード
図4. 一般的な抗真菌薬に対する主な耐性メカニズム。
ダウンロード 高解像度画像のダウンロード(818KB)
ダウンロード フルサイズ画像のダウンロード
図5. 真菌感染症を効果的に治療するための代替戦略。効果的な抗真菌療法のための代替戦略
抗真菌薬には限りがあり、また使用可能な抗真菌薬に対する耐性がよく報告されていることから、真菌感染症に対抗するための効果的な代替戦略の開発が必要とされている。真菌は真核生物であるため、抗真菌薬は宿主に対して毒性を示すことがある。従って、抗真菌剤の哺乳動物細胞毒性は薬剤開発における重要な基準である。効果的な抗真菌療法を開発するために、いくつかの戦略が提案されている。その最たるものが真菌細胞に特異的な新規薬剤の開発である。オロロフィム、VT-1129、VT-1161、VT-1598、CD101、APX-001、T-2307、VL-2397などの潜在的な抗真菌剤は、さまざまな臨床試験段階にあり、将来の抗真菌剤の候補となりうる(Wiederhold 2018)。イサブコナゾールやイブレキサファンガープなどの抗真菌薬は、真菌感染症の治療薬として、それぞれ2015年と2021年に米国FDAの認可を受けている(Van Matreら、2019;Schwebkeら、2021)。
単剤ベースの単独療法と比較して、相乗的併用療法は薬剤の有効性を高め、薬剤耐性の出現を抑制する。いくつかのin vitroおよびin vivo研究では、真菌感染症の治療におけるコンビナトリアルアプローチの有効性が報告されている(Spitzerら、2017;Vitale 2021)。コンビナトリアル・アプローチにおける相乗作用は、異なる細胞標的を有する薬剤の組み合わせ、または生物学的利用能の向上によって達成される。併用療法に使用される薬剤は、2種類の既知の抗真菌薬を組み合わせる方法と、抗真菌薬と非抗真菌薬を組み合わせる方法がある。併用療法における非抗真菌薬は、抗真菌薬の有効性を向上させることを目的としている。例えば、Hsp90(例:17-AAG)、カルシニューリン(例:シクロスポリンAおよびFK506)、リジン脱アセチル化酵素およびリジンアセチルトランスフェラーゼ(例:トリコスタチンA)の阻害剤などの分子は、抗真菌薬の活性を増強することが示されている(Spitzerら、2017)。
一般に、新しい抗真菌薬の開発には数年にわたる研究開発が必要であり、長期間のスケジュールを伴う。この問題は、承認された薬剤のリポジトリから抗真菌薬以外の薬剤を再利用することで克服できる。そのためには、計算モデリングやドッキング・アプローチにより、抗真菌活性を持つ可能性のある分子をスクリーニング/探索し、実験的に検証する必要がある(Kim et al.) このアプローチに基づく最近の研究では、抗がん剤(タモキシフェン)、抗リウマチ剤(オーラノフィン)、カルシウム拮抗剤(ニソルジピン、ニフェジピン、フェロジピン)、抗炎症剤(アスプリン、イブプロフェン、タクロリムス)、心血管系薬剤(アトルバスタチン)などの既知の薬剤と抗真菌活性の可能性が同定されている(Dasら、2021)。アリピプラゾール、ミルテフォシン、ベンズイミダゾール、キナクリン、ロベニジン、ラルテグラビル、シスプラチン、ピタバスタチンなどの他の再利用薬剤は、菌糸誘導、バイオフィルム形成を阻害し、感染動物モデルにおいて治療効果を示すことが示された(Kim et al.)
侵襲性真菌感染症に対抗するための宿主免疫の調節は、抗真菌療法の有望な戦略となりうる。感染動物モデルにおいて、顆粒球コロニー刺激因子(G-CSF)や顆粒球マクロファージコロニー刺激因子(GM-CSF)のようなアジュバントの使用は、カンジダ、アスペルギルス、クリプトコッカス感染症の治療における抗真菌薬の有効性を改善した(Scrivenら、2017;Samら、2018)。侵襲性アスペルギルス症動物モデルにおいて、IFN-γは抗真菌薬に対する反応を増強し、生存率を改善した。カンジダ、アスペルギルス、クリプトコッカス感染動物モデルでは、真菌細胞表面特異的モノクローナル抗体の投与により生存率が向上した(Scriven et al.) 侵襲性真菌感染モデルにおいて、顆粒球、T細胞、ナチュラルキラー(NK)細胞の輸血により生存率が向上したことが報告されている(Sam et al.) ワクチン戦略としての生きた弱毒真菌細胞または細胞壁成分は、動物モデルにおいて適応免疫応答を誘発し、真菌感染から保護することが示されている(Scrivenら、2017)。さらに、プロバイオティクス、ビタミン、マイクロバイオームによる免疫調節は、将来の抗真菌療法の有望な戦略である。
これらの戦略に加えて、いくつかの抗真菌ペプチド、抗真菌酵素、ナノ粒子、合成化学物質、低分子阻害剤、天然物、精油が、代替抗真菌剤として有望な可能性を示している(Padmavathi et al.) しかし、これらの薬剤を単独またはアジュバント抗真菌剤として使用する場合は、適切なアッセイ系と哺乳類の細胞毒性データによって検証する必要がある。
CRediT著者貢献声明
G.Kiran Kumar Reddy: 構想、原案執筆。Alwar Ramanujam Padmavathi:概念化、原案執筆。Y.V.ナンチャライヤ: 執筆-校閲・編集、監修。
利益相反宣言
著者らは、本論文で報告された研究に影響を及ぼすと思われる競合する金銭的利益や個人的関係はないことを宣言する。
推薦論文
参考文献
バーマンとクリサン, 2020
J. バーマン、D.J.クライサン
真菌における薬剤耐性と薬剤耐性
Nat. Rev. Microbiol., 18 (6) (2020), pp.
論文を見る
CrossRefView in ScopusGoogle Scholar
カンプザーノとワームリー、2018
A. カンプザーノ、F.L.ワームリー
クリプトコッカスに対する自然免疫、認識から排除まで
J. Fungi (Basel)., 4 (2018), p. 33
論文を見る
CrossRefView in ScopusGoogle Scholar
ダ・ローザら、2008
D.W. Da Rosa、E. Gezuele、L. Calegari、F. Goñi
胞子形成におけるアステロイド体。宿主免疫反応における酵母の生存率と生物学的意義
Med. Mycol, 46 (5) (2008), 443-448頁
PDFで見る
スコープで見るGoogle Scholar
Dasら、2021
R. Das, K. Kotra, P. Singh, B. Loh, S. Leptihn, U. Bajpai
COVID-19に伴う二次性細菌・真菌感染に対する代替治療戦略
Infect. Dis. Ther., 22 (2021), pp.
PDFを見る記事を見るGoogle Scholar
ディエップおよびホイヤー、2020年
A.L.ディープ、K.K.ホイヤー
コクシジオイデス感染に対する宿主応答:真菌免疫
Front. Cell. Infect. Microbiol., 10 (2020), Article 581101
スコープで見るGoogle Scholar
Elsegeiny et al.
W. エルゼゲイニー、K.A.マー、P.R.ウィリアムソン
クリプトコッカス感染症の免疫学:患者治療への合理的アプローチの開発
Front. Immunol., 9 (2018), p. 651
ScopusGoogle Scholarで見る
ガーフットら、2016年
A.L.ガーフート、Q.シェン、M.ヴートリッヒ、B.S.クライン、C.A.ラップリー
Eng1βグルカナーゼはβグルカンの露出を減少させることによりヒストプラスマの病原性を高める
MBio, 7 (2) (2016), pp.
グーグル・スカラー
エルナンデス-チャベスら、2017年
M.J.エルナンデス-チャベス、L.A.ペレス-ガルシア、G.A.ニーニョ-ベガ、H.M.モラ-モンテス
宿主の免疫認識を回避する真菌の戦略
J. Fungi (Basel)., 3 (2017), p. 51
記事を見る
CrossRefView in ScopusGoogle Scholar
ホッケンら、2019
M.W.J.ホッケン、B.J.ズワン、W.J.G.メルチャーズ、P.E.フェルヴァイ
臨床関連真菌における適応と抗真菌耐性メカニズムの促進因子
Fungal Genet. Biol., 132 (2019), Article 103254
https://www.cdc.gov/fungal/awareness-week.html
2021年12月31日アクセス
PDFを見る記事を見るScopusGoogle Scholarで見る
キムら、2020
Kim, J.H., Cheng, L.W., Chan, K.L., Tam, C.C., Mahoney, N., Friedman. M., Shilman, M.M., Land, K.M., 2020. 抗真菌薬の再利用。Antibiotics (Basel). 9(11), 812.
Google Scholar
リーとラウ、2017
P.P.リー、Y.-.L.ラウ
侵襲性真菌感染症の根底にある細胞および分子の欠陥-風土病真菌症からの解明
Front. Immunol., 8 (2017), p. 735
ScopusGoogle Scholarで見る
李ら、2019
Z. Li, G. Lu, G. Meng
肺における病原性真菌感染症
Front. Immunol., 10 (2019), p. 1524
ScopusGoogle Scholarで見る
マーら、2016
L. Ma, Z. Chen, D.W. Huang, G. Kutty, M. Ishihara, H. Wang, A. Abouelleil, L. Bishop, E. Davey, R. Deng, et al.
3種のニューモシスチスのゲノム解析から、哺乳類宿主のみで生活するための適応機構が明らかになった。
Nat. Commun., 7 (2016), p. 10740
PDFで見る
スコープで見るGoogle Scholar
マルコスら、2016
Marcos, C.M., de Oliveira, H.C., de Melo, W.C., da Silva, J.F., Assato, P.A., Scorzoni, L., Rossi, S.A., de Paula, E. Silva.A.C., Mendes-Giannini, M.J., Fusco-Almeida, A.M., 2016. 病原性真菌の抗免疫戦略。Front. Cell. Infect. Microbiol. 6, 142.
Google Scholar
ネットとアンデス, 2016
J.E.ネット、D.R.アンデス
抗真菌薬:活性スペクトル、薬理学、臨床適応症
Infect. Dis. Clin. North Am., 30 (1) (2016), 51-83頁
PDFを見る記事を見るScopusGoogle Scholarで見る
ニューランドとアブデルラーマン, 2009
J.G.ニューランド、S.M.アブデルラーマン
皮膚糸状菌症を中心としたテルビナフィンの最新情報
Clin. Cosmet. Investig. Dermatol., 2 (2009), 49-63ページ
スコープで見るGoogle Scholar
パドマバティら、2020年
A.R. Padmavathi、S.M. Murthy、A. Das、A. Priya、T.J. Sushmitha、S.K. Pandian
銅酸化物ナノ粒子によるカンジダ・アルビカンスの増殖阻害と酵母から菌糸体への移行
バイオファウリング, 36 (2020), pp.
論文を見る
CrossRefView in ScopusGoogle Scholar
Perlrothら、2007年
J. パーロス、チョイ、スペルバーグ
院内真菌症:疫学、診断、治療
Med. Mycol., 45 (4) (2007), pp.
PDFで見る
CrossRefView in ScopusGoogle Scholar
レディおよびナンチャライヤ、2020年
G.K.K.レッディ, Y.V.ナンチャライヤ
抗真菌剤としてのアルキルイミダゾリウムイオン液体:カンジダ・アルビカンスに対する抗バイオフィルム活性とその作用機序
Front. Microbiol., 11 (2020), p. 730
スコープで見るGoogle Scholar
サムら、2018
Q.H.サム、W.S.ユー、C.J.セネビラトネ、M.W.チャン、L.Y.A.チャイ
真菌感染症の治療としての免疫調節:我々は近づいているか?
Front. Microbiol., 9 (2018), p. 1612
スコープで見るGoogle Scholar
シュウェブケら、2021年
J.R. Schwebke、R. Sobel、J.K. Gersten、S.A. Sussman、S.N. Lederman、M.A. Jacobs、B.T. Chappell、D.L. Weinstein、A.H. Moffett、N.E. Azie、D.A. Angulo、I.A. Harriott、K. Borroto-Esoda、M.A. Ghannoum、P. Nyirjesy、J.D. Sobel
外陰膣カンジダ症治療におけるイブレキサファンガープとプラセボの比較:第3相無作為化比較優越性試験(VANISH 303試験)
Clin. Infect. Dis. (2021), p. ciab750
Google Scholar
スコルツォーニら、2017
L. スコルゾーニ、A.C.デ・パウラ・エ・シルバ、C.M.マルコス、P.A.アサト、W.C.デ・メロ、H.C.デ・オリベイラ、C.B.コスタ-オルランディ、M.J.S.メンデス-ジャンニーニ、A.M.フスコ-アルメイダ
抗真菌療法:真菌症の理解と治療における新たな進歩
Front. Microbiol., 8 (2017), p. 36
ScopusGoogle Scholarで見る
スクリブンら、2017年
J.E. Scriven、M.W. Tenforde、S.M. Levitz、J.N. Jarvis
侵襲性真菌感染症と闘うための宿主免疫応答の調節
Curr. Opin. Microbiol., 40 (2017), 95-103頁
PDFで記事を見るScopusで記事を見るGoogle Scholar
シルバら、2017
S. シルバ、C.F.ロドリゲス、D.アラウージョ、M.E.ロドリゲス、M.ヘンリケス
カンジダ種バイオフィルムの抗真菌抵抗性
J. Fungi (Basel)., 3 (1) (2017), p. 8
論文を見る
CrossRefView in ScopusGoogle Scholar
スピッツァーら、2017
M. スピッツァー、N.ロビンス、G.D.ライト
侵襲性真菌感染症に対抗するためのコンビナトリアル戦略
Virulence, 8 (2) (2017), pp.
PDFで見る
CrossRefView in ScopusGoogle Scholar
ヴァン・マートルら、2019
E.T. Van Matre、S.L. Evans、S.W. Mueller、R. MacLaren、D.N. Fish、T.H. Kiser
学術医療センターにおける侵襲性真菌感染症管理に対する硫酸イサブコナゾニウム、ボリコナゾール、ポサコナゾールの比較評価
Ann. Clin. Microbiol. Antimicrob., 18 (1) (2019), p. 13
PDFで見る
スコープで見るGoogle Scholar
ヴィターレ、2021
R.G.ヴィターレ
治療困難なカンジダ感染症における抗真菌薬併用の役割
J. Fungi (Basel)., 7 (9) (2021), p. 731
論文を見る
CrossRefView in ScopusGoogle Scholar
ヴィーダーホールド, 2018
N.P.ヴィーダーホールド
抗真菌薬:代替薬と将来のターゲット
Int. J. Antimicrob. Agents., 51 (3) (2018), 333-339頁
PDFで記事を見るScopusで記事を見るGoogle Scholar
サラゴサ、2011
O. サラゴサ
クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)における形態形成と表現型変異の概念。
Front. Microbiol., 2 (2011), pp.
論文を見る
CrossRefGoogle Scholar
引用者: (7)
新規抗カンジダ薬としてのイサチン-ベンゾトリアゾールハイブリッドの設計、合成および生物学的評価
2023, 分子構造学会誌
引用抜粋:
真菌感染症の中でもカンジダ感染症は最も蔓延しており、微生物感染症による死亡原因の第4位を占めている。それとは別に、カンジダ感染症は、中心静脈ライン関連血流感染症の3番目に多い感染源であり、その多くは小児、高齢者、免疫不全患者を標的としている[3,4]。口腔咽頭カンジダ症、外陰膣カンジダ症、粘膜皮膚カンジダ症、播種性カンジダ症は、最も一般的であり、医療システムにとって新たな課題となっている[5-7]。
抄録を表示
カルコン・シッフ塩基はSaccharomyces cerevisiaeおよびCandida albicans細胞の細胞膜の完全性を破壊する
2023年、微生物学アーカイブ
臨床非皮膚糸状菌カビのレパートリー
2023年、真菌学雑誌
抗菌剤としてのラムノピラノシドベースの脂肪酸エステル: 抗菌剤としてのラムノピラノシド系脂肪酸エステル:合成、スペクトル特性、PASS、抗菌、分子ドッキング研究
2023、分子
酸化亜鉛ナノ粒子のカンジダ・アルビカンスに対する抗真菌特性
2022, コーティング
抗真菌デリバリーを強化するためのシクロデキストリンベースのナノキャリアの現在の概要
2022, 医薬
Scopusの全ての引用論文を見る
1
これらの著者は同等に貢献した。
© 2022 The Authors. 発行:エルゼビアB.V.
サイエンスダイレクトについて
リモートアクセス
ショッピングカート
広告掲載
お問い合わせ
利用規約
プライバシーポリシー
当社は、サービスの提供・向上、コンテンツや広告のカスタマイズのためにクッキーを使用しています。続行することで、クッキーの使用に同意したことになります。
Copyright © 2023 Elsevier B.V. or its licensors or contributors. ScienceDirect® は Elsevier B.V. の登録商標です。
この記事が気に入ったらサポートをしてみませんか?