
考察 Origin of the life

Paul M. Nurse, Ph.D (UK)の著書 "WHAT IS LIFE?" 竹内薫訳「生命とは何か?」より、不必要な修飾語を省略して示す。以下がkey pointsと考える。
---ここから引用、重要な語を(*!)で示す。(*!#)は後でその実態を検討する。
無生物の無秩序な化学物質が、秩序だった形態に自分を配置した。
自らを永続(*!)させ、自らをコピー(*!)し、最終的に自然淘汰(*!)によって進化する、能力を獲得した。
(中略)
生物には、他に依存するウイルスから、自給自足の生活を送る細菌や植物まで、、、、異形態がある。
全形態は、他生物に依存(*!)、自然淘汰で進化し、自らを律する物理的存在である。
(中略)
相互のつながり(*!)こそが、生命の中核だ。
(中略)
最古の化石は、現在の細菌のいくつかに似ている。その時点で生命がすでに、膜(*!1)に包まれた細胞、DNAに基づく遺伝(*!2)システム、タンパク質に基づく代謝(*!3)作用などを備え、、
どれが最初だったのだろう?
DNAに基づく遺伝子の複製、タンパク質をベースにした代謝作用、包み込む膜組織だろうか?
---ここまで引用
2022/8/18 追記:
竹内薫訳「生命とは何か, ダイアモンド社, ISBN978-4-478-11107-9, C0045
を通読した。この本は文系学科の教養課程生に向くように思われる。
興味深い点: p.186, チューリングの、相互作用している化学物質のふるまいと、それらが構造に拡散するときに起きる特定の化学反応とを予測する、一組の数学の方程式を編み出した。
予想外なことに、彼が反応拡散モデルと呼んだこの方程式は、
化学物質を精薇で時に美しくすらある空間パターンに配列する。。。
ここであった。ナースは具体的な方程式形を記載していないが、拡散方程式に発展項を追加すれば、反応拡散方程式になる。本ノートの後半で、それを計算する。
長沼毅「死なないやつら」ブルーバックスB-1844、講談社、2013.12.20,
ISBN978-4-06-257844-8, C0245 も興味深い本で、最初は極限状態で生存できる生物の紹介かと思って手に取ったが、内容は「生命とは何か」に言及している。著者はシュレーディンガーの、
「生命とは負のエントロピーを食って、構造と情報の秩序を保つシステム」
を紹介して、「生命とは自己増殖するロバストな散逸構造*」と言明する。
生命~散逸構造という見解は多くの人が指摘している。
私も、この解釈である。そして散逸構造ならしめる数式が反応拡散方程式と考えている。
*) さんいつこうぞう dissipative structure 熱力学的に平衡でない状態にある開放系構造。エネルギーが拡散する流れに自己組織化が発生すること。
この本で興味深かった点:
<1> p.46~なぜ高温でも平気なのか?の段のp.48、1種類のアミノ酸を複数のコドンが指定する。。。
のところで、アミノ酸を指定する情報的な意味は等価でも、水素結合の数により、高温でのDNA安定化に相違がある、ことの指摘である。
<2> p.119細胞の寿命の始まり。多細胞生物の遺伝子にはテロメアという細胞分裂を規定する部分があるが、こういう仕掛けが何故成立してきたのか?という疑問に、ある程度は答えている。
<3> p.157チューブワームの代謝機構。
< 2022.9.24 追加 >
T. NAKAMURA M. MATSUMOTOK. AMANOY. ENOKIDOM. E. ZOLENSKYT. MIKOUCHIH. GENDAS. TANAKAM. Y. ZOLOTOV[...]Y. TSUDA
"Formation and evolution of carbonaceous asteroid Ryugu: Direct evidence from returned samples", SCIENCE, 22 Sep 2022; DOI: 10.1126/science.abn8671
https://www.science.org/doi/10.1126/science.abn8671
「炭素質小惑星リュウグウの形成と進化:返還サンプルからの直接証拠」
Samples of the carbonaceous asteroid Ryugu were brought to Earth by the Hayabusa2 spacecraft. We analyzed seventeen Ryugu samples measuring 1-8 mm. CO2-bearing water inclusions are present within a pyrrhotite crystal, indicating that Ryugu’s parent asteroid formed in the outer Solar System. The samples contain low abundances of materials that formed at high temperatures, such as chondrules and Ca, Al-rich inclusions. The samples are rich in phyllosilicates and carbonates, which formed by aqueous alteration reactions at low temperature, high pH, and water/rock ratios < 1 (by mass). Less altered fragments contain olivine, pyroxene, amorphous silicates, calcite, and phosphide. Numerical simulations, based on the mineralogical and physical properties of the samples, indicate Ryugu’s parent body formed ~ 2 million years after the beginning of Solar System formation.
炭素質小惑星リュウグウのサンプルは、探査機はやぶさ 2 によって地球に運ばれました。1-8 mm のリュウグウのサンプル 17 個を分析しました。
(!) リュウグウの親小惑星が太陽系外縁部で形成されたことを示す、磁硫鉄鉱の結晶内に CO2 を含む水の含有物が存在します。
サンプルには、コンドリュールや Ca、Al が豊富な介在物など、高温で形成された材料が少量含まれています。
サンプルはフィロケイ酸塩と炭酸塩が豊富で、低温、高 pH、および水/岩石比 < 1 (質量) での水性変質反応によって形成されます。
変化の少ない破片には、かんらん石、輝石、非晶質ケイ酸塩、方解石、リン化物が含まれています。
サンプルの鉱物学的および物理的特性に基づく数値シミュレーションは、リュウグウの母体が太陽系形成の開始から約200万年後に形成されたことを示しています。
---- こういう発表を目にすると、以下の仮説が信憑性を帯びてくる。
長沼毅「隕石・彗星内ハビタブルゾーン(パンスペルミアの方舟)
Biological Sciences in Space, Vol.19(1), p8-24, 2005.
パンスペルミア(panspermia)仮説:
spermia=胚種、地球圏外で発生した圏外生物の胞子
宇宙にはpansperimiaが漂い、惑星に降下して生命の起源となる、ことを主張する仮説
事実:(1)ハレー彗星の核にはCHON粒子が全質量の10%以上存在する。Humber 1990.
(2)マーチソン隕石に、地球生物が蛋白質に使用しないアミノ酸(L型過剰)を含有する。Chrnin 1997.
この事実を重要と思うので、 2022.9.28 捕捉する。アミノ酸の光学活性について;
大島泰郎「生命の起源と不整性の起源」日本結晶学誌vol.19,p.177(1977)という、生命科学者の談話室という論文調の文がある。一部を要約して紹介する。氏の意見等は省略した。
ーーーここからーーー
生命とは情報化された分子機械と表現できる.分子レベルの情報システムが整然と動作する基礎は分子そのものの構造上の規則性である.
そこで生体物質には,化学構造上たくさんの“禁制律”が存在する.
タンパク質を作るアミノ酸は二十種,核酸の塩基は五種,等々である.
しかし生化学的禁制律のうちで最も代表的 なものは光学活性に関するものである。生体に利用されるアミノ酸はL型,糖の多くはD型である。
(!) 生体内では酵素とよばれるタンパク質でできた触媒の表面で化学反応が進行する.アミノ酸の合成も同様に固相表面に固定し,非対称な環境のもとでL型のみを生みだす.
その非対称な環境を作りだしているのは酵素タンパ ク質がL型アミノ酸とい う非対称性の構成単位からできているからである。光学活性の起源は二つの仮説{必然説、偶然説}に分けられる。
生命の起源が一回,確率の低い事象がおこったとして説明するなら,
非対称性の起源も簡単に説明がつく.
低い確率だが,すべてL型アミノ酸で自己増殖能をもったタンパク質が原始地球上で偶然合成され ,自己増殖系が突然に成立したとすれば生物界の不整性の起源も解決される.
あるいは,外からたった一つの細胞や胞子が飛来してきて原始地球上にすみついたのが生命の始まりである、とする説をとれば、地球上の不整性の起源は解決される。
どこか干潟で分別結晶が起こった。 風がL型の結晶1ケを,濃縮しつつあったとなりの池へ運び、これが結晶化の種になった。結晶は底に沈み,溶液は川へ洗い流された.こうして非対称の世界が成立する. そしてここが生命誕生の舞台となった.
ーー引用 終わり
(3)BIOPAN-1/2計画でアミノ酸を地球周回軌道で暴露した。アミノ酸は光分解した。ラセミ化はしなかった。Barbier 1998.
(4) NASAのLDEFによる5年の長期暴露試験、枯草菌胞子の生残率、O(-2)~O(-4)。ブドウ糖が保護剤となった。Shimizu 1999.
これはカッシーニが測定したエンケラドゥスの水蒸気スペクトルからリン酸イオンが見つかり、Simで生成過程を確認した、という報告: https://phys.org/news/2022-09-evidence-habitability-ocean-saturn-moon.html.
Home/Astronomy & Space/Astrobiology, SEPTEMBER 19, 2022
"Researcher helps identify new evidence for habitability in ocean of Saturn's moon Enceladus"
SwRI Lead Scientist Dr. Christopher Glein contributed to new findings that phosphorus in the form of orthophosphate (e.g., HPO42-) is likely abundant in the subsurface ocean of Saturn's moon Enceladus.
オルトリン酸塩 (例: H(PO4)2-)が土星の衛星エンケラドゥスの地下海に存在する、可能性が高い。
A soda or alkaline ocean (containing NaHCO3 and/or Na2CO3) inside of Enceladus interacts geochemically with a rocky core.
Modeling indicates that this interaction promotes the dissolution of phosphate minerals, making orthophosphate readily available to possible life in the ocean.
Because phosphorus is an essential ingredient for life, this finding bolsters mounting evidence for habitability within this small Saturnian moon.
エンケラドス内部のアルカリ性の海 (NaHCO3, Na2CO3 含有) は、岩石コアと化学的に相互作用する。
この相互作用でリン酸塩の溶解を促進し、オルトリン酸塩を海洋生物が利用できるようにする。
リンは生命にとって不可欠な成分である、この知見は、土星の衛星の生命可能性を補強する。
< 2022, 10/10 追記 > 書籍の紹介
中沢弘基「生命誕生」講談社現代新書2262、講談社、2014.5.20,
ISBN978-4-06-288262-0.
生命誕生の考察にimpactを与えるものは、事実である。
simulationはprogrammerの思考した仮想の現実である。
本書には、その事実が書かれている。一部を紹介する。
第1の注目すべき点はp.85の第3章、化石の証拠、であろう。
p.91 シアノバクテリアが27.2億年前に存在したこと、
p.93 30億年前の球菌の化石らしいものに一部無機起源と思われる組織があること、
それで球菌らしく見えても「化石」とは言えない、ことを紹介する。
これらの事実は考察に多大な影響を与える。
中沢先生も述べておられるが、球菌と見間違える小胞組織が非生物的に生成する、事実は生命の起源に迫る重要な発見と考えられる。
p.97 2011年硫黄を代謝に利用する34億年前のバクテリアの化石が発見されている。
球胞内部に1nmのFeS2の結晶粒があり、細胞周囲に1~10μmのFeS2がある。
生物進化の順番として、無酸素で生きる硫酸還元/硫黄を使った代謝のバクテリアがいた筈という報告があったが、本化石はその証拠である。重大である。
この部分はp.281のFe-Sワールド説も参照すること。p.285にはFe-S膜の小胞の記載まである。
本事実の指摘は他書になかなか見いだせない。
>> Biological Science in Space, vol.20, No.1 (2006), pp.3-9,
小林憲正「アミノ酸の前生物的合成と触媒機能の起源」には記されている。
一種の代謝系を形成するというアイデアと書かれている。FeS表面が化学進化を触媒する、ことは調べられている。
光学異性体の起源についても、マーチソン隕石のことが詳しく書かれている。ここは必読。
この総説には「ゴミ袋ワールド」まで記されているが、具体的な化学反応例を提示しないと、単なるお話しに終わってしまう。
ーーー
Feイオンが複素環化合物を錯体化して、触媒機能を高めたことが、書かれている。あり得ることである。
ここで、軟体動物の酸素輸送系のヘモシアニンについて書いておく。
理由は:: Cu原子は太陽では核反応で合成できない原子である。そういう原子を使った生物が地球上に存在する、ということは、生命材料の原子起源の一部は太陽系外なのである。
ヘモシアニン (hemocyanin, Hc) : 呼吸色素のひとつ。
エビ・カニ・昆虫の一部 等の節足動物、貝の一部やイカ・タコ等の軟体動物に見られる。
軟体動物の赤貝や環形動物のゴカイはヘモグロビンと似た鉄由来のエリトロクルオリンという呼吸色素を持つ。
ヘモシアニンは無色透明で、酸素と結合し銅イオンの青色となる。
3つのヒスチジン残基のもつイミダゾール環の窒素3つに囲まれた銅原子を2つ使い、酸素分子を挟むようにして結合する。
この構造中の銅原子により自然光から~600nmの橙色が吸収され、補色の青色になる。
この構造をサブユニットとして円柱状に配列した多量体をつくり、分子量は数百万に達する。
機能: 酸素を運搬する呼吸色素の役割を持つ。血球中に含まれるのではなく、血リンパ液に溶存する。
酸素の結合力は、ポルフィリン環内に鉄が配位されたヘモグロビンよりも弱い。
ヘモシアニンは、2つの銅イオンに蛋白質が結合して、普通10個が結合して中空の円柱構造を形成する。
ヘモグロビンの分子量は64,500であるが、ヘモシアニンは2,500,000以上の分子量。
ヘモグロビンに比べヘモシアニンは酸素親和性は低いので、肺のような酸素分圧が高いところでも酸素で飽和することはなく、運搬効率は悪い。酸素分圧の低い末梢では効率よく酸素を放出する。
カブトガニなどの祖先が誕生した頃は、水中の酸素濃度が低かったので、ヘモシアニンの方が酸素運搬能力が高かったのではないか?と言われている。
National Geographic, News:
https://natgeo.nikkeibp.co.jp/nng/article/20150316/439302/
南極のタコの血は青くて濃い、予想を裏切る新発見、赤青緑、透明…生きものの血の色はなぜ異なるのか 2015.03.17 という記事がある。紹介する。
南極海に生息するマダコ科の一種(Pareledone charcoti)、血液には銅を含むヘモシアニンという酸素運搬タンパク質が存在するため、青色を帯びている。
しかし、Pareledone charcotiにとっては、そう簡単な話ではない。水温が氷点下になる南極海では、ヘモシアニンと酸素の結合が強すぎて、切り離すのが困難である。ふつうに考えれば、体に酸素が行き渡らず、タコは死んでしまうはずなのだ。
3月11日に"Frontiers in Zoology"誌に発表された研究によると、南極海のタコはヘモシアニンを40%過剰に作り出すことで、この問題を解決している。
完全に透明な血を持つ魚: ジャノメコオリウオ(Chionodraco rastrospinosus)は、南極のタコと同じく冷たい海に生息しているが、その血液の色は透明だ。ヘモグロビンもヘモシアニンもない。
低温の水は、温かい水よりはるかに多くの酸素をたくわえることができ、同魚の生息する深度の海水には十分な量の酸素が溶け込んでいるので、酸素運搬タンパク質を利用する必要がない。
同魚にはウロコがない。ウロコがないことで、皮膚から直接酸素を取り入れることが可能になる。異常に大きい心臓が血漿中の酸素を全身に行き渡らせているのではないか?と考えられている。
ーーーNews 終わり
筆者コメント:: Fe-S worldは地球でなくとも可能な筈で、彗星、太陽系以外から飛来してくる天体、オウムアムア (Oumuamua)**、そういう天体の成分分析が切に欲しい、と思う。推測を検証する事実が、ここは必要である。
**) 軌道要素は判明している。表面は太陽系外縁天体に似た赤色である。彗星コマが確認されない。Astrophysical Journal Letters, 2017 に次の論文が掲載されている。主成分はH2分子であるが、含まれている微成分が知りたい。同Lettersの一部を引用する。
https://arxiv.org/abs/2005.12932,
Evidence that 1I/2017 U1 (`Oumuamua) was composed of molecular hydrogen
1I/2017 U1 (`オウムアムア) が水素分子の氷で構成されていた証拠
authors: Darryl Seligman, Gregory Laughlin
`Oumuamua (I1 2017) was the first macroscopic (L∼100m) body observed to traverse the inner solar system on an unbound hyperbolic orbit.
Its light curve displayed strong periodic variation, and it showed no hint of a coma or emission from molecular outgassing.
Astrometric measurements indicate that 'Oumuamua experienced non-gravitational acceleration on its outbound trajectory, but energy balance arguments indicate this acceleration is inconsistent with a water ice sublimation-driven jet of the type exhibited by solar system comets.
オウムアムア (I1 2017) は、非束縛・双曲軌道で太陽系を横断する、最初の観測 (L~100m) 天体です。光度曲線は周期的変動を示し、コマや分子ガス放出による発光の兆候は見られない。アストロメトリー測定によると、オウムアムアは外向軌道で非重力加速度をした。しかし、エネルギー収支の議論では、この加速が太陽系の彗星によって示されるタイプの水の氷の昇華駆動ジェットと矛盾している。
We show that all of `Oumaumua's observed properties can be explained if it contained a significant fraction of molecular hydrogen (H2) ice. H2 sublimation at a rate proportional to the incident solar flux generates a surface-covering jet that reproduces the observed acceleration.
オウマウムアの観測された全特性は、水素分子 (H2) が氷のかなりの部分を含む場合に説明できることを示している。 入射太陽フラックスに比例する速度での H2 昇華は、観測された加速度を再現する、表面を覆う ジェットを生成する。
Mass wasting from sublimation leads to monotonic increase in the body axis ratio, explaining `Oumuamua's shape.
Back-tracing `Oumuamua's trajectory through the Solar System permits calculation of its mass and aspect ratio prior to encountering the Sun.
昇華による質量の消耗は、体軸比の単調増加につながり、オウムアムアの形状を説明しています。
太陽系を通過するオウムアムアの軌道を逆追跡することで、太陽に遭遇する前にその質量と縦横比を計算することができます。
We show that H2-rich bodies plausibly form in the coldest dense cores of Giant Molecular Clouds, where number densities are of order n∼10**5, and temperatures approach the T=3K background.
Post-formation exposure to galactic cosmic rays implies a τ∼100 Myr age, explaining the kinematics of `Oumuamua's inbound trajectory.
密度がn〜10 ** 5のオーダーであり、温度がT = 3Kに近い巨大分子雲の最冷部の高密度中心で、H2に富む天体が形成されたことを示します。
形成後の銀河宇宙線への露出は、τ~100 Myr の年代を意味し、オウムアムアのインバウンド軌道の運動を説明しています。----引用終
ーーーコメントここまで
< 2022.10.28 追記 >
[1] Noble gases and nitrogen in samples of asteroid Ryugu record its volatile sources and recent surface evolution
Science, R. Okazaki, et.al., DOI: 10.1126/science.abo0431
生命の起源に重要な情報 図1:リュウグウの進化図。
1.リュウグウ母天体の形成と先太陽および始原的ガスの獲得。<-- (!) pre-solar and primordial gases
太陽系の惑星形成原子は先太陽の合成した重金属(Fe以上の原子量の)を継承している、ということ。ヘモシアニンを酸素輸送に使用する軟体動物の存在から予測した、太陽系外で合成した原子の存在が、ハヤブサ2の論文からも支持された。
2.リュウグウ母天体での水質変質(約45.6億年前)。
3.母天体破片の集積によるリュウグウ形成。
4.近地球軌道への移動(約500万年前)。
5.加熱による赤化(約100万年以上前)。6.現在のリュウグウ。
[2] https://www.jaxa.jp/press/2022/10/20221021-2_j.html
First asteroid gas sample delivered by the Hayabusa2 mission: A treasure box from Ryugu
Science Advances, R. Okazaki, et.al., DOI:10.1126/sciadv.abo7239
という論文も公表された。
ーーー小惑星リュウグウの分析 論文 引用終
中沢本に戻る。p.114 細胞内共生説: 細菌類の進化では遺伝子の変異によって生じる進化とは別の、
原核生物が他のバクテリアを呑み込んで内部に共生させ、小器官*として高度な機能を獲得して進化する、経路がある。現在、この説は多くの研究者にとって常識化している。
*) ミトコンドリア、葉緑体、宿主とは別のDNAを有す。
p.274 遺伝機構に引き継がれた「結晶成長の情報伝達機構」
この部分は興味深い。検証がさらに必要。現段階では仮説である。
以上、本書を推薦する。 ーーー中沢本、紹介終わり
柴谷篤弘、長野敬、養老孟司 編 「講座 進化5 生命の誕生」、東京大学出版会、1991.12.10; ISBN4-13-064215-4
古い本ですが、推薦できます。特に p.151生命の初期進化とRNA、柳川弘志
のヌクレオチドのリン酸化、ヌクレオチドの前生命的合成 に注目しました。
< 2022.11.6 追記 > 上記の本にはRNA worldのことが書かれているが、その世界の遺伝情報(ゲノム)を子孫に伝達する、その際の変異についての記載があまり見られないと考えるので、ここで少し考えてみたい。
RNA worldは失われた世界なので、その断片がRNA生物的なもの、たとえばvirusに残っている、と考える。
インフルエンザ・ウイルスの遺伝の研究で最近、興味深い発表があった。
https://www.amed.go.jp/news/release_20180105.html
国立大学法人東京大学、国立研究開発法人日本医療研究開発機構、プレスリリース、「インフルエンザウイルスの遺伝の仕組みを解明」
河岡義裕、野田岳志
1. 研究の背景・先行研究における問題点
インフルエンザウイルスは8本に分かれたRNAをゲノムとして持ちます。
感染細胞から新たに作られるウイルスは、その粒子の中に8本に分かれたRNAをもれなく取り込む必要があります。そのメカニズムの詳細はわかっておらず、8本のRNAを効率よく子孫ウイルスに取り込ませるような特別な仕組みがあるのかどうか、長年の論争となっていました。
2. 研究内容
次世代シークエンス解析と電子顕微鏡解析によって、子孫インフルエンザウイルス粒子の中に取り込まれているRNAを調べました。通常の子孫ウイルス粒子には8本のウイルスRNAが“1+7” という配置をとって取り込まれていました。ウイルスRNA以外のRNA(感染細胞のRNA)は、ほとんど取り込まれていませんでした。<7は@@@数>
次にリバースジェネティクス法を用いて、ウイルスRNAを1本欠き7本しかウイルスRNAを持たない変異ウイルスを人工合成し、その変異ウイルス粒子の中に取り込まれたRNAを解析しました。
予想に反して、7本しかウイルスRNAを持たない変異ウイルスにも、“1+7”に束ねられた8本のRNAが取り込まれていました。
この変異ウイルスに取り込まれた8本目のRNAは、インフルエンザウイルスのRNAではなく、感染細胞に存在するリボソームRNA(!)でした。
今回の発見から、インフルエンザウイルスが子孫ウイルスにゲノムを伝えるとき、8本のRNAを“1+7” に集合させるステップが重要であることがわかりました。
さらに、ウイルスのRNAが足りないときには、“1+7” に集合させるために細胞のリボソームRNAを奪い取る仕組みを持つことが明らかになりました。
ーーー
このプレスリリースを読んで、2022.7.25の東北大プレスリリースを思い出す。
https://www.tohoku.ac.jp/japanese/2022/07/press20220725-01-sars.html
コロナウイルスのゲノムに蓄積した進化の痕跡を発見 コロナウイルスがもたらすパンデミックの機序解明へ期待
2022年7月25日 11:00
概要:
SARS関連コロナウイルスの変異において、長い塩基配列の挿入・欠失が見られること、この変異がウイルスの進化やパンデミック発生に関与している可能性を明らかにした。
新たに挿入された塩基配列と類似の配列はコロナウイルスゲノム中に存在せず、宿主動物など外界に由来するもの(!)と考えられる。
本研究は、未知の部分の多いSARS関連コロナウイルスの進化やパンデミック発生に関与している可能性がある。
自然界における不連続変異や宿主ジャンプをしばしば起こす同ウイルスの更なる有害変異を阻止する手段として。。。
ーー引用ここまで、PDFありDL可能
以上。
失われたRNA worldにおける「遺伝」とは、今のDNA worldの遺伝情報変異(random mutations in genomes、中立変異の蓄積 etc)とは相違する、RNA機能単位の奪取(上記に宿主ジャンプと書かれていることに同じ)、の様相を示していた、ように感ずる。
< 2022.10.17 追記 > Nature Astronomy, https://www.nature.com/articles/s41550-022-01786-w
Letter Published: 10 October 2022
Early Mars habitability and global cooling by H2-based methanogens
初期火星、居住可能性とH2ベース・メタン生成菌による全球規模冷却
Boris Sauterey, Benjamin Charnay, Antonin Affholder, Stéphane Mazevet & Régis Ferrière
Nature Astronomy (2022)Cite this article
Abstract
During the Noachian, Mars’ crust may have provided a favourable environment for microbial life1,2.
The porous brine-saturated regolith3,4,5 would have created a physical space sheltered from ultraviolet and cosmic radiation and provided a solvent, whereas the below-ground temperature2 and diffusion6,7 of a dense, reduced atmosphere8,9 may have supported simple microbial organisms that consumed H2 and CO2 as energy and carbon sources and produced methane as a waste.
ノアキア時代*、火星の地殻は微生物にとって好ましい環境を提供していた可能性があります 1,2。<この#は元論文のref.引用#です>
ーーー
*) 先ノアキス代、先ノアキア代(Pre-Noachian): 約45億年前の惑星への降着と分化から、ヘラス平原の形成までの間隔を表す。
火星に豊富な水をもたらす前の初期の気候だと考えられている。
しかし、この時代の地質学的記録のほとんどは、その後の侵食及び後期重爆撃期や火山作用によってなくなっている。
火星は、先ノアキア代の最初の4億年間で大量の水を宇宙空間への流出で失った。
アルギュレ平原やイシディス盆地はこの時代に形成されたと考えられている。
多くの隕石が衝突した時代で、多くのクレーターが作られた。
火星は約45億年前から地磁気が存在していたとされる。
当時の火星の磁場は現在の地球とほぼ同じ磁力だったとされる。
火星の地磁気は、約42億年前まで存在していたとされる。
ただし、約39億年前に火星の地磁気は消失したという説もある。
ーーー
多孔質の塩水で飽和したレゴリス3,4,5 は、紫外線や宇宙線から保護された物理的空間を作り出し、
溶媒を提供したと考えられますが、地下温度 2 と高密度で還元された大気 8,9 の拡散 6,7 は、微生物を支援する可能性があります。
エネルギーと炭素源として H2 と CO2 を消費し、廃棄物としてメタンを生成する生物に。
On Earth, hydrogenotrophic methanogenesis was among the earliest metabolisms10,11, but its viability on early Mars has never been quantitatively evaluated.
Here we present a probabilistic assessment of Mars’ Noachian habitability to H2-based methanogens and quantify their biological feedback on Mars’ atmosphere and climate.
地球上では、水素栄養メタン生成は最も初期の代謝の 10,11 でしたが、
初期の火星でのその実現可能性は定量評価されたことはありません。
ここでは、H2ベースのメタン生成菌に対する、火星のノアキア期の居住可能性の確率論的評価を提示し、
火星の大気と気候に対する生物学的フィードバックを定量化します。
We find that subsurface habitability was very likely, and limited mainly by the extent of surface ice coverage.
Biomass productivity could have been as high as in the early Earth’s ocean.
However, the predicted atmospheric composition shift caused by methanogenesis would have triggered a global cooling event, ending potential early warm conditions, compromising surface habitability and forcing the biosphere deep into the Martian crust.
Spatial projections of our predictions point to lowland sites at low-to-medium latitudes as good candidates to uncover traces of this early life at or near the surface.
地下の居住可能性はあり得、表面の氷の被覆範囲によって制限されていることがわかりました。
バイオマスの生産性は、初期の地球の海洋と同じくらい高かった可能性があります。
しかし、メタン生成によって引き起こされる、大気組成の変化は、全球の寒冷化を引き起こし、
初期の温暖状態を終わらせ、表面の居住性を損ない、生物圏を火星の地殻の奥深くに追いやったでしょう.
私たちの予測は、低緯度から中緯度の低地域を、地表または地表近くで、
初期の生命の痕跡を明らかにするための、適切な候補地として示しました。
ーーー追記終ーーー
以上、様々な研究の概要を紹介しました。が、、重要な事柄が言語により示されている。本記事では、言語の内容をcomputer simulationで検討するapproachを採用して、各語の内容を考察したい。
---前提: 内容を理解するために必要な事:
(1) Fortran言語のcodeが読める、(2) 偏微分方程式、移流拡散方程式、反応拡散方程式の知識、(3) 時系列超行列・画像化の知識。
まず小生のblog記事https://scienceimg140e.fc2.net/
(1) 2022/3/5: Difference equation for proliferating particles
(2) 2022/1/17: Reaction-Diffusion equation
をお読み下さい。
上記(1)では外界と区別される粒子に、膜(*!1)の存在を導入しています。
この膜は細胞膜でなくとも良く、異なる性質の媒体境界の化学物質の面です。
さらに親子の粒子に、遺伝(*!2)情報の存在を導入しています。
この遺伝情報を担う分子はDNA/RNAでなくとも良く、粒子の寿命を有限にする紫外線などによる確率的な分子分解過程でも良いです。
親から子を生成するのは、粒子を分割する触媒分子でも良いです。以下の論文では「触媒」でなくとも可能です。
---公表論文による補足---
ここまで、(*!1,*!2)という過程を天下りに導入しています。それは「あり得る過程」なのか?ということで、調べてみると以下の論文が発表されています。
https://www.nature.com/articles/nphys3984.epd,
Nature Physics; David Zwicker, Rabea Seyboldt, Christoph A. Weber, Anthony A. Hyman and Frank Jüliche, “Growth and division of active droplets provides a model for protocells”.
Abstract: It has been proposed that during the early steps in the origin of life, small droplets could have formed via the segregation of molecules from complex mixtures by phase separation.
These droplets could have provided chemical reaction centres. However, whether these droplets could divide and propagate is unclear.
Here we examine the behaviour of droplets in systems that are maintained away from thermodynamic equilibrium by an external supply of energy.
In these systems, droplets grow by the addition of droplet material generated by chemical reactions.
Surprisingly, we find that chemically driven droplet growth can lead to shape instabilities that trigger the division of droplets into two smaller daughters. ←(!)
Therefore, chemically active droplets can exhibit cycles of growth and division that resemble the proliferation of living cells.
Dividing active droplets could serve as a model for prebiotic protocells, where chemical reactions in the droplet play the role of a prebiotic metabolism.
訳: 「活性液滴の成長・分裂は原始生命体のモデルを提供する」
生命の起源の初期段階では、相分離による複雑な混合物からの分子の分離により小液滴が形成された可能性があると提案されています。これらの液滴は、化学反応の中心を提供する可能性があります。
ただし、これらの液滴が分裂して伝播できるかどうかは不明です。
ここでは、エネルギーの外部供給によって熱力学的平衡から離れて維持されているシステム内の液滴の挙動を調べます。
これらのシステムでは、化学反応によって生成された液滴材料の添加によって液滴が成長します。
驚くべきことに、化学的に駆動される液滴の成長は、液滴の2つの小さな娘への分割を引き起こす形状の不安定性につながる可能性があることがわかりました。←(!)
したがって、化学的に活性な液滴は、生細胞の増殖に似た成長と分裂のサイクルを示す可能性があります。
アクティブな液滴を分割することは、液滴内の化学反応がプレバイオティクス代謝の役割を果たすプレバイオティクス原始細胞のモデルとして役立つ可能性があります。
---ここまで論文を引用
暗に粒子の時間方向の継続に、代謝作用(*!3)を前提としています。
Paul M. Nurseが代謝と書いたので、そうしましたが、本当はenergy & compound resource ではないかと考えます。生命現象を発生・継続させるための元エネルギー・元物質、元化合物のことです。虚無から有意な構造体(生命)を構成できません。
微粒子系が数百step時間存続するために、有限寿命の粒子が失活したときに、構成成分をresourceの戻す過程を前提としています。この過程は別種の粒子(生命体)でなくとも良く、物理化学の分解反応であっても可能です。
<3/16追加分>
反応拡散方程式の「反応項」は微分方程式形で定義するが、その形では天下りに増殖粒子の挙動をmodelerが式で与える。論文査読ではその式の正当性を議論する。正当性の検証には対象の増殖粒子の実験が必要である。推論ではだめである。
小林憲正(地球宇宙化学/横浜国立大学)教授が、「物質が生命となった瞬間」https://www.ddbj.nig.ac.jp/activities/gene-genome.html; において、次のように主張されている。---ここから内容を引用---
生命とは,自己複製と物質交代をする物質である。
物質というからには,生命を持たない物質と本質的に違うわけではない。
生命の本質である「自己複製」と「物質交代」の中心に遺伝子がある。
--- ここまで主張を引用
大古の現生命系とは異なるメカニズムの可能性がありうる。
粒子系の挙動simulationではどのようにすべきか?
本考のsimulation algorithmを以下とした。
増殖粒子拡散: 一様乱数による移流(物理学の乱流拡散)として扱う。
物質拡散:物理学の拡散方程式の数値解法で計算する。増殖粒子拡散と「近傍」の扱いが異なるので、物質拡散は1 stepあたり10回以上必要である。
メッシュ幅:粒子の近傍=中心位置から±10 mesh正方形領域、物質の近傍=±1 mesh
直径などの物性が異なる粒子なので、大きさのorderを変える。
Information of a gene (one particle): finite vector,
Gene(1)=age 粒子は物理化学的な周囲の擾乱により有限stepで分解する。
Gene(2)=x-coordinate, real# in [0,400.5]、位置座標は生物学の遺伝子には無いが、simulationを実行する上で必須である。移流拡散simulationの要請である。
Gene(3)=y-coordinate, real# in [0,400.5] ← until Gene2D.f90
--- ここまでの性質は生命特有の現象でない。以下、遺伝子の特徴的な性質である。
Gene(4)=stack area, real#
Gene()の具体的な物質表現(塩基配列等)を議論しない。機能のみを考える。
Functions of each Gene-information: Coding
遺伝子発現:Gene(4)領域の値とcodingの両方の効果
<3/25 補足>
Syozo OSAWA, “Evolution of the Genetic Code”, Oxford University Press, 1995.
大澤省三「遺伝暗号の起源と進化」訳:渡辺公剛、上田卓也、大濱武、
共立出版、1997, ISBN4-329-05475-X.
以下、訳本のページで生命の起源についての要点をしめす。
p.213 図11.1 RNA world以前の「前生物的進化」この部分を、本noteで言及する。
幾多の生命起源論がRNA world以後のDNA worldの中の遺伝暗号に言及する。同暗号codeは細胞内の蛋白質合成にアミノ酸配列を介して関与する。暗号codeはアミノ酸に関連づけられる。その関連は全生物共通である、と初歩的な本に書かれることが多いが、その「関連」も時間が経つと変化する。
大澤教授の「遺伝暗号の起源と進化」には機構が詳細に記述されている。
それは「分子の他分子集団に及ぼす機能の時間変化」現象である。RNA/DNAでなくともあり得る、化学分子の集団的反応現象の時間変化である。本noteはこの立場で「考察」する。
<3/29 補足>
生理機能を有す分子は蛋白、糖蛋白、脂質蛋白質である。蛋白質を生成する機構の元はtransfer RNA (t-RNA)である。tRNAの構造はp.17の図1.1(核酸塩基76個)にある。すでにanti-codon (3塩基の組み合わせ)が存在している。受容、T-、D-、anti-codon stemsの副構造が見られる。このような複雑な有機分子は無生物的な反応の組み合わせでは容易に生成できない。
p.40 t-RNAとアミノ酸の「アミノアシル化反応」にはaminoacyl t-RNA synthetase (ARS)群が必要である。それで特異的に各種アミノ酸との結合がなされる。
生物(シアノ・バクテリア)の痕跡がある化石は35億年前である*(地球は46億年前)。35億年前には細胞の体制が既に出来ている、ことになる。
*) 宮田隆「分子から見た生物変化」講談社ブルーバックス B-1849, 講談社, 2014.1.20. ISBN978-4-06-257849-3, p.288に記載あり。
本書p.182~184、免疫系の所に「DNAは細胞分化の過程で変化」とあり、かつ「DNA断片化の組み合わせにより遺伝子の多様化が進化過程で起きた可能性」とある。機能の要素化、その組み合わせの反応、という考え方は従来の物理化学では、ほとんど無い。
p.165~200にt-RNA, ARSの(分子)進化についての諸説を紹介した9章がある。重要な点を列記する。
(1) p. 167 表9.7 Murchison meteorite (マーチソン隕石)のアミノ酸構成比
p.187のアミノ酸の遺伝子codeへの参加順、表9.3
(2) p.171 原始蛋白質のアミノ酸候補, Ser, Asp, Gly, Lys.
ARS保存domainとt-RNA amino acid accepter dmainの相互作用は「アミノ酸に対する機能的RNA暗号」を生じさせる。
(3) p.183 t-RNAの出現は38±5億年前。遺伝子codeは地球と同程度に古い。
< t-RNAのような分子が無生物的な化学反応では容易に出来ない。なので暗に「生命の根幹部は地球起源でない」と言っているようである>
<4/7追加> 下記資料が公開されている。一部を抜粋して記す。
https://www.rnaj.org/component/k2/item/759-suzuki-3 「RNAと生命の起源: 鈴木勉(日本RNA学会長)」2022.4.7 net上に存在する。
略
RNAは..分子を認識する能力を備え、リボザイムとして酵素の働きを担う。
40億年前の地球において、RNAが生命誕生の鍵を握る...RNAワールド仮説です (Gilbert, 1986)。
略
東北大学の古川善博先生を中心とし、北海道大学とNASAとの共同研究チームから、隕石からリボースを発見したという、大変興味深い発表がありました (Furukawa et al., 2019)。
隕石にはアミノ酸や核酸塩基が含まれることが知られていました。
炭素の同位体分析から、このリボースは地上で生じたものではなく、宇宙空間で形成されたことが明らかになっています。
ホルモース反応 (formose reaction) と呼ばれる化学反応で、ホルムアルデヒドから糖が合成できることは知られていましたが、今回の成果は宇宙空間において、非生物的にリボースが生成したことを示した画期的な成果であると言えるでしょう。
生命誕生前の地球上でも同様の反応でリボースが生成した可能性はありますが、今回の成果は宇宙で生成したリボースが地球に降り注ぎRNAを生成した可能性を示しています。
---引用ここまで
<4/24,28追記>
Astrobiologyという学問分野が既に存在する(NASAにHP: https://astrobiology.nasa.gov/まである。
”Our work centers around life's origin, evolution, & distribution in the stars. And still, everything we know to be alive resides on this pale ... と記載されている。)、と聞き、時代の進歩には驚かされます。それで天体物理の啓蒙書を読みました。
鳴沢真也著「連星からみた宇宙」講談社ブルーバックスB-2150, 2020.12.20; ISBN978-4-06-521354-4.
生命現象に関係するのは元素の歴史です。p.228の表11-1元素合成のまとめ、が重要と思いました。
{H, He, Li} by big-bang,
{He~Fe; ex. C, N} by nuclear fusion in stars,
{O, Na, Cl, C~Rb} by supernova (gravitational collapse),
{Ca, Ni, Cu, S} by supernova ( Ia-type),
{Sr ~ Bi} by long-time process in the giant stars,
{Pu, Au, U} by neutron star collision,
{Rb, Sr, Mo} by X-ray burst.
これで見ると、C~Oの周期律表の第1段の元素を主成分として生命が生まれるしかあり得ません。
生命の起源とは関係ないですが、現代天文学は興味深いです。特に以下の点:
7章には「謎のエネルギー」とあり、p.147には宇宙が現在、膨張しており、しかもその速度が加速している、ことが書かれています。ということは明らかになっていない反発力を生じさせる何かが存在します。”dark energy"と呼ばれています。
p.153には重力波の性質が書かれています。振幅が距離に反比例(1/r)と書かれています。p.163にはGW150914のブラックホール連星の衝突で3太陽質量が物質からエネルギーに変換されて、重力波として地球に伝わったことが書かれています。
ブラックホールからは物質、光子のような素粒子でも出てこない、とされています。しかし空間のゆがみの波、重力波は出てきます。そしてエネルギーを持っている。
ブラックホール単独では何もエネルギー、物質は出しません。量子力学を導入しなければ、確かにそうですが。
---感想文ここまで。
<5/4, 追記, 重要> https://www.nature.com/articles/s41467-022-29612-x, Nature Communications;
"Identifying the wide diversity of extraterrestrial purine and pyrimidine nucleobases in carbonaceous meteorites", 炭素質隕石中の地球外プリンおよびピリミジン核酸塩基の幅広い多様性の特定,
Yasuhiro OBA, et.al.; https://doi.org/10.1038/s41467-022-29612-x;
The lack of pyrimidine diversity in meteorites remains a mystery since prebiotic chemical models and laboratory experiments have predicted that these compounds can also be produced from chemical precursors found in meteorites.
Here we report the detection of nucleobases in three carbonaceous meteorites using state-of-the-art analytical techniques optimized for small-scale quantification of nucleobases down to the range of parts per trillion (ppt).
プレバイオティクスの化学モデルと実験室での実験により、これらの化合物は隕石に含まれる化学前駆体からも生成できると予測されているため、隕石にピリミジンの多様性がないことは謎のままです。
ここでは、トリリオンあたりの部分(ppt)の範囲までの核酸塩基の小規模な定量化に最適化された最先端の分析技術を使用して、3つの炭素質隕石中の核酸塩基の検出を報告します。
In addition to previously detected purine nucleobases in meteorites such as guanine and adenine, we identify various pyrimidine nucleobases such as cytosine, uracil, and thymine, and their structural isomers such as isocytosine, imidazole-4-carboxylic acid, and 6-methyluracil, respectively.
Given the similarity in the molecular distribution of pyrimidines in meteorites and those in photon-processed interstellar ice analogues, some of these derivatives could have been generated by photochemical reactions prevailing in the interstellar medium and later incorporated into asteroids during solar system formation.
This study demonstrates that a diversity of meteoritic nucleobases could serve as building blocks of DNA and RNA on the early Earth.
グアニンやアデニンなどの隕石で以前に検出されたプリン核酸塩基に加えて、シトシン、ウラシル、チミンなどのさまざまなピリミジン核酸塩基と、それぞれイソシトシン、イミダゾール-4-カルボン酸、6-メチルウラシルなどの構造異性体を特定します。
隕石中のピリミジンの分子分布と光子処理された星間氷類似体中のピリミジンの分子分布の類似性を考えると、これらの誘導体のいくつかは、星間物質で優勢な光化学反応によって生成され、後に太陽系形成中に小惑星に組み込まれた可能性があります。
この研究は、多様な隕石核酸塩基が初期の地球上のDNAとRNAの構成要素として役立つ可能性があることを示しています。 ----- Nature Comm. 引用ここまで。
< 9/3 追記 >宇宙のプラズマ衝撃波は宇宙線を生成すると考えられている。
実験室でプラズマ衝撃波を生成させる論文があったので報告する。
九大の解説 https://www.kyushu-u.ac.jp/f/49387/22_0830_01.pdf
以下にRhys.Rev.Eの論文(abstract)と訳を示す。
High-power laser experiment forming a supercritical collisionless shock in a magnetized uniform plasma at rest
R. Yamazaki et al.
Phys. Rev. E 105, 025203 – Published 11 February 2022
We present an experimental method to generate quasiperpendicular supercritical magnetized collisionless shocks. In our experiment, ambient nitrogen (N) plasma is at rest and well magnetized, and it has uniform mass density. The plasma is pushed by laser-driven ablation aluminum (Al) plasma. Streaked optical pyrometry and spatially resolved laser collective Thomson scattering clarify structures of plasma density and temperatures, which are compared with one-dimensional particle-in-cell simulations. It is indicated that just after the laser irradiation, the Al plasma is magnetized by a self-generated Biermann battery field, and the plasma slaps the incident N plasma. The compressed external field in the N plasma reflects N ions, leading to counterstreaming magnetized N flows. Namely, we identify the edge of the reflected N ions. Such interacting plasmas form a magnetized collisionless shock.
Received 23 September 2020Revised 30 November 2021Accepted 19 January 2022
DOI:https://doi.org/10.1103/PhysRevE.105.025203
準垂直超臨界磁化無衝突衝撃を生成する実験的方法を提示します。 私たちの実験では、周囲の窒素 (N) プラズマは静止しており、十分に磁化されており、均一な質量密度を持っています。
プラズマは、レーザー駆動のアブレーション アルミニウム (Al) プラズマによって押し出されます。
ストリーク光高温測定と空間分解レーザー集団トムソン散乱により、プラズマ密度と温度の構造が明らかになり、1 次元の細胞内粒子シミュレーションと比較されます。
レーザー照射の直後に、Al プラズマが自己生成された Biermann バッテリー フィールドによって磁化され、プラズマが入射 N プラズマを平手打ちすることが示されています。
N プラズマ内の圧縮された外部場は N イオンを反射し、反対方向の磁化された N の流れを引き起こします。
つまり、反射した N イオンのエッジを識別します。 このような相互作用するプラズマは、磁化された無衝突衝撃を形成します。
High-power laser experiment on developing supercritical shock propagating in homogeneously magnetized plasma of ambient gas origin
S. Matsukiyo et al.
Phys. Rev. E 106, 025205 – Published 26 August 2022
A developing supercritical collisionless shock propagating in a homogeneously magnetized plasma of ambient gas origin having higher uniformity than the previous experiments is formed by using high-power laser experiment. The ambient plasma is not contaminated by the plasma produced in the early time after the laser shot. While the observed developing shock does not have stationary downstream structure, it possesses some characteristics of a magnetized supercritical shock, which are supported by a one-dimensional full particle-in-cell simulation taking the effect of finite time of laser-target interaction into account.
Received 25 September 2020Accepted 24 July 2022
DOI:https://doi.org/10.1103/PhysRevE.106.025205
高出力レーザー実験を用いて、前の実験よりも高い均一性を有する周囲ガス起源の均一磁化プラズマ内を伝播する発生中の超臨界無衝突衝撃を形成する。
周囲のプラズマは、レーザー ショット後の初期に生成されたプラズマによって汚染されません。 観測された発生中の衝撃波は、定常的な下流構造を持っていませんが、磁化された超臨界衝撃波のいくつかの特徴を持っています。これは、レーザーとターゲットの相互作用の有限時間の影響を考慮した、1 次元の完全な粒子内セル シミュレーションによってサポートされています。
--- Rhys.Rev. 引用ここまで
=== 以下 blog の続き ===
4. Code, Gene2D.f90:
4.1 Simulation #11
初期状態:(1)1粒子、年齢1[step]
粒子特性:(1)有限寿命5[step]、(2)1step毎に1[mass]のリソースを消費する。
(3)近傍メッシュ内の総消費リソースが1に満たない場合は死す。
(4)増殖可能期間2~4[step]
(5)Gene上にstep時間の経過と共に蓄積する一様乱数正値[0, 0.01]情報を置く。世代間の遺伝情報スタックである。この情報は価が1.0になるまで発現しない。
発現すると、粒子が自分を中心にして近傍メッシュ区間をI~IV象限に分け、最もresource massの大きい象限に一様乱数[0, 0.2]だけ移動する。粒子が周囲を認知するように見えるが、ここでは浸透圧のような非生命的な現象を考えている。
(6)粒子は、世代間の遺伝情報スタック値によらず、現在のx,y座標を中心に[-1,+1]区間の一様乱数で移動する。項番(5,6)の粒子移動は加算される。Simulation #10の実効的移動量:1.4/2=0.7, expectation=0; #11の移動量:1/2+0.1=0.6, expectation=0+0.1=0.1 このexpectation valueが粒子分布図で描かれる。
(7)2D-map外に出る場合は、0~400.5区間に反射させる。
増殖:(1)1粒子から別の1粒子、年齢1を生成する。
(2)生成時10[mass]のリソースを消費する(近傍メッシュ内)。
(3)10[mass]のリソースが無い場合、増殖しない。粒子数が9990以上は増殖できない。Simulation stepを粒子数が9990に達する前に止める。
(4) 世代間の遺伝情報スタック値を子粒子に継承させる。これは世代間を跨って存在するtimer領域である。そこに蓄積された変異が閾値を超えると機能を発現する。
結果:
1. 粒子移動機能の発現による粒子数の変化
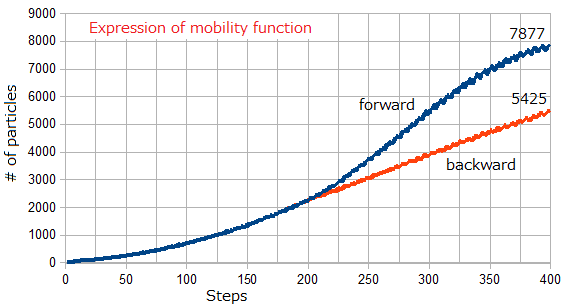
Blue line: uniform random number +[0,0.2]:=forward,
Red line: uniform random number -[0,0.2]:=backward
The time step of the expression is about 205.
粒子数変化曲線の微小な揺れの原因はSimulation#2のロジスティック曲線と同じである。
Forward方向に移動すると400*400の境界面に接近するので、増加数が頭打ちになる。
常微分方程式dP/dt=α(1-P)Pの解に漸近する。
Forward/Backwardの 2D空間分布図。

Particle distribution on Backward
点描法:Timer領域発現前 R=(240, 180,120,60,0), G=(0,60,120,180,240), for Age=(1,2,3,4,5).
Timer領域発現後 R=(240, 180,120,60,0), B=(0,60,120,180,240), for Age=(1,2,3,4,5).
5. Code, Gene2E.f90, Gene2S.f90
5.1 Simulation #12
初期状態:(1)1粒子(位置:矩形空間の中央)、年齢1[step]
粒子特性:(1)有限寿命5[step]、(2)1step毎に1[mass]のリソースを消費する。
(3)近傍メッシュ内の総消費リソースが1に満たない場合は死す。
(4)増殖可能期間2~4[step]
(5)Gene上にstep時間の経過と共に蓄積する一様乱数正値[0, 0.01]情報を置く。世代間の遺伝情報スタックである。世代間の情報を子粒子に継承できる確率は1/2である。
継承できないときは、その情報は0にリセットされる。継承・非継承は乱数で決定する。
この情報は価が0.04になるまで発現しない。
発現するとSimulation #11のForward機能が働く。
発現しない場合、その粒子はresourceとは異なる二種類の「物質」を粒子位置に残す。
物質1特性:Step毎に累積 し、分解しない。位置がsimulation時間により変化しない。
物質2特性:位置がsimulation時間により変化しないが、半減期5 stepsで分解する。
これらの物質は粒子群の構造を生成する。
増殖:(1)1粒子から別の1粒子、年齢1を生成する。
(2)生成時10[mass]のリソースを消費する(近傍メッシュ内)。
(3)10[mass]のリソースが無い場合、増殖しない。粒子数が9990以上は増殖できない。Simulation stepを粒子数が9990に達する前に止める。
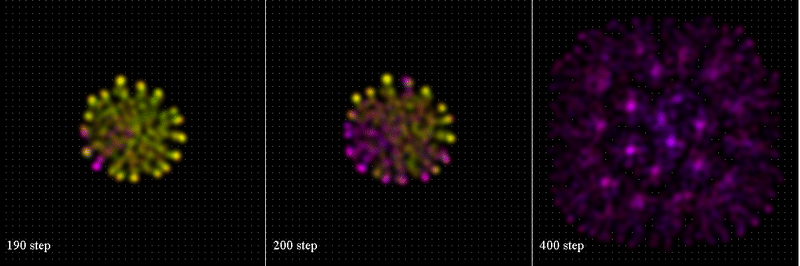
結果: Sim12A.png: 1. 粒子数、拡散回数の変化, 2. 粒子の平均年齢、蓄積エリアの平均値

dirSex.png: Step=190,200,400の時の粒子分布図。粒子の色はSim#11と同様

Protein.png: 粒子の残存物質1,2の分布, Step=(190,200,400)
色彩:物質1: G:1/2, B:1 brightness, mass*2.5
物質2: G:1/2, R:1 brightness, mass*30 (!) Gene2S.f90で描くこと。

<3/21追加>
6. Code, Gene2{SA,SB,SC}.f90
6.1 Simulation #13
粒子が増殖、自発的に移動、双極子モーメントを有す代謝物を排出する場合に、該代謝物の構造がどうなるか?を調べる。-OH基は水素結合により双極子であり、周囲に電場がある。新たな代謝物は、その電場の影響を受け、最もエネルギーの低い配向になる。それにより規則性のある構造物が生成されるのではないかと考えて本simulationを行う。
初期状態:(1A)1粒子(位置:矩形空間の中央)、年齢1[step]
(1B) 3粒子(矩形空間の中央と±20 mesh x,y方向にズレた位置)、年齢 1 step
粒子特性:(1)有限寿命5[step]、(2)1step毎に1[mass]のリソースを消費する。
(3)近傍メッシュ内の総消費リソースが1に満たない場合は死す。
(4)増殖可能期間2~4[step]
(5)Gene上にstep時間の経過と共に蓄積する一様乱数正値[0, 0.01]情報を置く。世代間の遺伝情報スタックである。世代間の情報を子粒子に継承できる確率は1/2である。
継承できないときは、その情報は0にリセットされる。継承・非継承は乱数で決定する。
この情報は価が0.02になるまで発現しない。
発現するとSimulation #11のForward機能が働く。ただし移動量は乱数*0.001(ほとんど乱流拡散しないで、resource最大の方向へ粒子が自発的に移動する)。
発現しない場合、その粒子はresourceとは異なる、双極子モーメントのある「物質」を粒子位置に残す。配向は粒子位置の電場により、最小エネルギーの配向となる。
物質特性:Step毎に累積 し、分解しない。位置がsimulation時間により変化しない。
物質は正負のcharge構造を生成する。
増殖:(1)1粒子から別の1粒子、年齢1を生成する。
(2)生成時10[mass]のリソースを消費する(近傍メッシュ内)。
(3)10[mass]のリソースが無い場合、増殖しない。粒子数が9990以上は増殖できない。Simulation stepを粒子数が9990に達する前に止める。
結果: DPorig1.png: 初期粒子1の場合の、正負の電荷分布図。
正電荷は白、負電荷は黒で表示される。灰色はcharge 0部分。"nk"は"n"回simulation stepを計算したことを示す。
DPorig3.png: 初期粒子3の場合の、正負の電荷分布図。
正負電荷が交互に連続する「ひも状」の構造体が生成される、傾向がある。これは重要な結果ではないか、と考える。


<4/2,4追加>
移流項*の効果をsimulationで調べる。そのために矩形領域の境界条件を変える。
*) 増殖粒子の拡散とは、物理的に見れば、粒子位置の変化である。移流拡散方程式の移流項部に相当する。「移流」が乱数的に不定方向に変化するから、巨視的にみれば拡散のように観察されるので、今まで「粒子・拡散」と言った。ゆえに粒子に対する∇演算子はない。
一方、resourceは粒子を構成する分子であるから、物理的な拡散である。移流方程式の拡散項が存在する。
*補足)粒子とresourceの移流も別個に考え得る。水中の粒子の挙動ならば同一であるが、体内ではウイルスのような粒子とresourceの移流は異なる。一度に様々なsimulationは出来ないので、ここでは「体内」を想定して、粒子とresourceの移流を別のもの、とする。
<4/7補足>
Robart M. HAZEN, "The Story of Earth", 2012.
訳:円城寺守、渡会圭子「地球進化46億年の物語」講談社、ブルーバックスB-1865,2014, ISBN978-4-06-257865-3.
p.186, 自己複製できる分子の疑似生命組織がどのようにしてできたかを説明するモデルが3つ...
いくつかの小分子の回路....クエン酸回路...
クエン酸が自然に2つの小分子に分かれて酢酸と...になると、この回路が自己複製を始め、
.... 地球上の細胞すべてにクエン酸回路が組み込まれている.....
最初の生命体から継承されている化学的化石...と書かれている。
クエン酸回路は「好気的代謝に関する反応回路」なので、地球誕生直後の無酸素時代に、この回路が在ったとは考えられない。調べると、
還元的クエン酸回路(reductive TCA cycle):クエン酸回路と逆向きの反応による代謝経路... がある。
どちらも回路が一巡して元の分子に戻ってしまうのだから自己複製と同じである。そして回路外からクエン酸への生成経路もある。
回路の中の1種の分子、たとえば「クエン酸」に注目すると、これが本稿の増殖粒子である。他の分子は全部をまとめてresourceである。
<4/8 追記>
本書p.215に、35億年前の微生物の化石(シアノバクテリアとされる)への疑問点が書かれている。
記載はp.220まで、最古の分子化石の議論が続いている。2014年代の暫定見解では、27億年の化石が最古のものらしい。ここにホパン*という有機化合物名が記されている。細胞膜を安定化し、細胞以外ではめったに存在しない、と書かれている。約20億年前に存在していた原始微生物には核がない、とも書かれている。
*) ホパン( hopane, Hopanoids)についてnet上を検索した。https://www.geo.kyushu-u.ac.jp/img/graduate_school/fields/orge.pdf, 奈良岡、山内、北島、「宇宙・地球環境における有機物の存在と役割」というpdfが公開されている。
ーーーここまで"The Story of Earth"の引用、補足
旧:境界で粒子を反射させる。
新:境界外の粒子位置を「剰余の位置」に換える。これにより有限系から無限系に近くなる。粒子近傍のmeshも同様である。
粒子の初期条件: 1粒子(年齢1)、位置は左端中央から20 mesh位置。
粒子のproperty: 初期値0のスカラー遺伝情報1個、毎step +0.01増加する。
移流量: x軸方向:uniform random*0.2 +0.1, x軸正方向に乱流拡散しながら毎step 0.1 mesh移動する。y軸方向: uniform random*0.2, y軸方向は乱流拡散のみ。
スカラー遺伝情報値が0.04を超えると粒子の運動性が変わる。
0.04以下の上記移動量に加えて、その粒子位置を中心に1~4象限の近傍meshに在るresource量を加算し、最大の象限に向かってabs(random)*0.2移動する。自発的粒子移動の1モデルである。
自発移動した粒子はスカラー量のtracerを残さない。
0.04以下の非自発移動粒子は、自己位置にtracer mass 1を残す。Tracerには3種あり、それぞれの半減期は5, 15, 45 stepである。Tracer図により粒子群の移動状況が分かる。同図では半減期の短い順にRGB色で表示する。
粒子の増殖:Age 2~4のsimulation step毎に子粒子を1個生成する。ただし親の粒子の近傍mesh内にresource mass >10の条件が必要である。条件が満足されなければ、その近傍内の最大age粒子が分解される(粒子間の淘汰戦)。粒子分解でresource mass 10が捻出され、それを使って新粒子を生成する。他の粒子がない場合は、子生成は行われない。
子粒子のスカラー遺伝情報値は、親粒子のそれを継続する場合とリセットされる2種類があって、その選択は一様乱数で決定される。それぞれの確率は1/2である。
粒子の性質をsimulationに取り入れようと考え、最初は雄雌のような形を考えたが、もっと根源的なものとして、遺伝の特徴である「情報の継続性」に焦点を当てた。
どの粒子も1 step存続するためにresource mass 1が必要である。近傍mesh範囲にその量が無ければ粒子は死滅し、その近傍に均等に(resource mass 10)/(21*21)のmass値を増加させる。
粒子のageが5を超えると死滅し、同様に近傍mesh内均等にmass値を増加させる。
Resource massの拡散:物質の物理拡散を想定している。近傍meshが±10 meshであるので、10回の物理的拡散を無条件に行う。その拡散プロセスでも全空間内にmass値0以下の点が存在する場合、最大99回の物理的拡散処理を行う。その回数を記録に残し図示する。それは粒子数の変化と共に、それを成立させる背景条件の空間内の時系列変化である。生命存続モデルである増殖粒子群の存在の背景条件の変化は重要と考えている。
7. Code, Gene2Saa.f90
7.1 Simulation #14
粒子群が移動(X軸方向に全空間でrandom*0.2+0.1)しているか否かを調べる。
粒子の分布をStep={600, 900, 1800)で表示する。Age=1~5は赤~青で表示される。下段の図はTracerの分布である。青い影が左方向(-X軸)に尾を引いているので確かに+X軸方向に移動している。
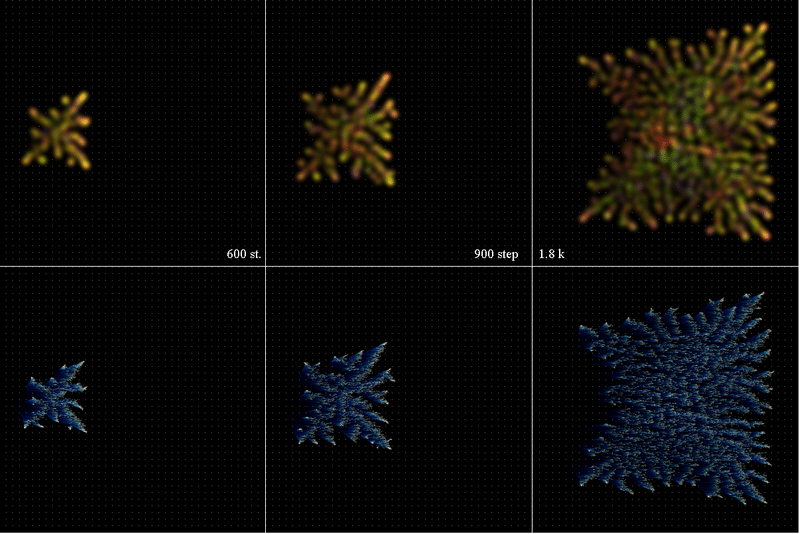
7.2 Simulation#15 空間内に粒子を移動させるbandを置く(Y軸値150~200)。Band内では粒子はrandom*0.2+0.1移動する。Band外では粒子移動はX,Y軸共にrandom*0.2である。剰余無限系で粒子の飽和状態を調べるために、粒子の分布をStep={2k, 3k, 4k)で表示する。Age=1~5は赤~青で表示される。
下段の図はTracerの分布である。青い影が左方向(-X軸)に尾を引いているので確かに+X軸方向に移動している。<Gene2Sab.png>

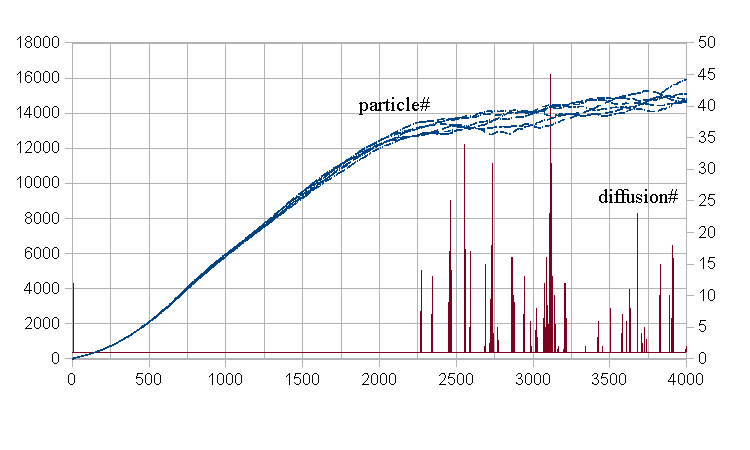
The property<G2Saa1:3,png>

Step>2kでは系は飽和に近くなっている。Diffusion#も多くなっており粒子間の淘汰戦が行われている。
閉じた系での微生物の増殖はsigmoid functionになる。上図のparticle# plotは概ねその関数形を再現する。粒子間の淘汰戦が、生成粒子の±10 mesh内の局所空間となるため、粒子群の内部に局所構造が生成して、粒子数が振動する。淘汰戦がstep 2000以降に多くなるため、分解された粒子のresourceの拡散に多くの操作が成される。これが微粒子増殖と微粒子数飽和の実相である。
<G2Saa2.png>

上図は1 mesh あたりのresource mass量の時間変化である。本simulationは初め全空間均一に"1”のresourceである。粒子は増殖して、そのmassを消費する。その状況をstep毎に展開して示したのが上図である。{max, average, min}は1 meshあたりのmass量である。初期値1に対する相対値である。3曲線とも細かく振動している。特に系が粒子で飽和に近い状態になるとmax resource mass曲線の振動が顕著になる。粒子間の淘汰戦の結果である「粒子→resource」の拡散過程の激しさを反映している。
<G2Saa3.png>

上図は粒子の年齢と遺伝子に蓄積されるスカラー値の平均値を時系列で示した。
粒子はage 1~5であるから、平均するとage=3となる。値が振動するのは粒子間の淘汰戦の結果である。蓄積値は毎step+0.01、子生成の時に継承する、しないは1/2の確率で決まるから、蓄積値の平均は0.025になる。しかし1k step以降にわずかに低下して0.02に近づく。蓄積値>0.04で、粒子は乱数的な移動の他に、自発的にresource量の多い象限へ移動する。そうすると、飽和状態では他粒子との淘汰戦になる。蓄積量>0.04はage~4なので、負ける確率が高い。それで蓄積値が低下する。
--
粒子集団では「環境により遺伝情報が変化している(情報の発現が変化)」ように見える現象である。
本simulationでは「自発的粒子運動」発現条件、あるいは子生成時の「蓄積値継承」比率へのフィードバック処理がcodingされていないので、遺伝子機能・変更圧が働かない。
現生命体のDNA淘汰メカニズムをsimulationしていない、この単純化simulationでも「遺伝情報変更」のきざしが現れる。
7.3 Simulation#16 2-流れ場(+/- 0.3 to X-axis)
空間内に粒子を移動させる2 bandsを置く(Y軸値150~200, 50~100)。Bands内では粒子はrandom*0.2±0.3(3倍)移動する。Bands外では粒子移動はX,Y軸共にrandom*0.2である。剰余無限系で粒子の飽和状態を調べるために、粒子の分布をStep={2k, 3k, 4k)で表示する。Age=1~5は赤~青で表示される。
下段の図はTracerの分布である。青い影が左方向(±X軸)に尾を引いているので確かに±X軸方向に移動している。
<Gene2Sac.png>,

The property<G2Sac1:3,png>
Simulation#15と違い、particle#の上昇曲線に小さな波うち現象がある。Bandが粒子拡散運動の微かな障壁となっている。

Max resource in a meshの挙動がsimulation #15との違いである。小stepsから粒子間の淘汰戦が起こっている。遺伝情報蓄積値によらない2Bands内の速い粒子移動が、このような現象を起こす。

Simulation#15と同じ現象がacc_value曲線に見いだせる。
8. Code, Gene2path.f90
8.1 Simulation #17
毛細血管のような複雑な経路を粒子が増殖しながら拡散、移動すると、どのように粒子数が変化するか?を調べた。<@>初期値:5粒子、位置:@@@
<Gene2path.png>
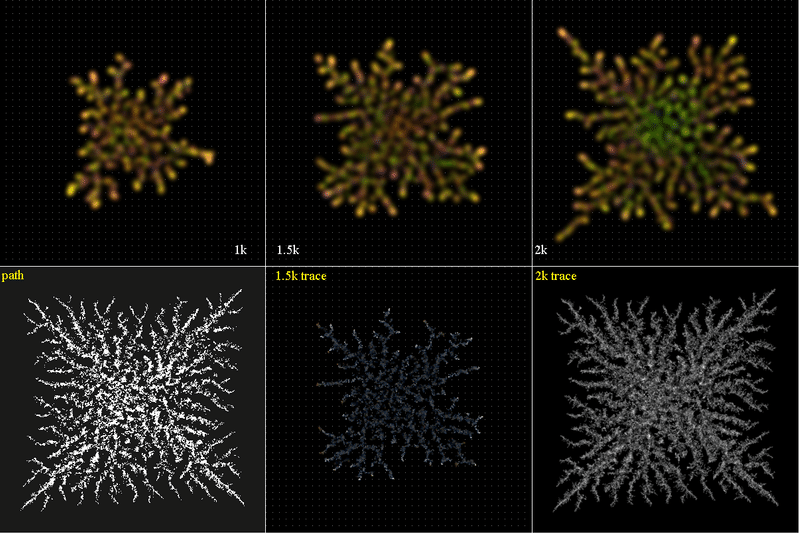
下段、左端の”path”が粒子の移動経路である。白い部分ではrandom in [-1,1] mesh, 1 step毎に移動する。黒い部分ではrandom in [-0.1, 0.1] mesh移動する。
上段はstep={1k, 1.5k, 2k}の時の粒子分布図である。赤~紫で粒子群のageを示した。
下段にstep={1.5k, 2k}の時のtracer分布図を示した。粒子は一定方向に移流していないので、青く尾を引いて流れの方向を示す、ことはない。
Tracerの分布は粒子の移動経路を示し、それはほぼpathに一致する。このpath経路はGene2Saa.f90の移流項を0としたsimulationのtracer分布を2値化したものである。
ゆえに「別の粒子群の移動記憶を継承した」と解釈できる。生命体の記憶にはDNA codon codeばかりではなく、このような記憶も存在すると考えている。
The property:

細血管のような経路の終端までしかsimulationしていないので、粒子の飽和状態にはならない。
粒子数は直線的に増加している。Acc_value (遺伝情報の蓄積)値も0.025で一定である。淘汰戦が少なかったことを示している。


<@>感染・浸透simulation: 外界、細胞膜、細胞、体腔、構造を増殖粒子が浸透する、1モデル




この記事が気に入ったらサポートをしてみませんか?