西洋近代と日本語人(仮題) その8

3. 暴力について ――人類学風の考察―― (続き)
暴力とヒトの社会性(続き)
3.148. さて、ボウルズ&ギンタス(2017)* の複数レベル淘汰の素描を続けます。今回は…………と書き始めて、あれこれ考えているうちに、暗礁に乗り上げた。困った。私は、ボウルズとギンタスの議論の根底にある直観を受け入れていないらしい。それにあらためて気づいてしまった。かれらの議論は、統計学やゲーム理論の知識を前提としているから、なかなか細部まで理解しきれない。そういう意味で、ちゃんと受け入れることができていないということはもちろんある。しかし、多少とも理解できた部分について、それにいささか同意できないことに気づいた。
注*: ボウルズ&ギンタス(2017)『協力する種 制度と心の共進化』竹澤正哲[監訳]、大槻久、高橋伸幸、稲葉美里、波多野礼佳[訳]、NTT出版。(原著、Bowles, S. and Gintis, H. (2011). A Cooperative Species: Human Reciprocity and Its Evolution. Princeton University Press.)
3.149. 理解できたが同意できないというのは、数学的定式化の出発点に置かれた基本的な仮定のことです。ボウルズとギンタスは、ホモ属における協力の進化を数学的に処理するために、淘汰の単位である個体の振る舞いを単純化する。端的にいうと、ホモ属の個体は、協力するかしないか相手の出方で態度を変えたりしないし、まわりの個体たちの影響で協力したりしなかったり態度を変えることもない、という仮定を導入している。私はそう読解した。
3.150. ボウルズとギンタスは自分たちの仮定が「非現実的(unrealistic)」であると認めていますが、数学的なモデル化にはこの仮定は不可欠です。そして、私は、ホモ属の個体をこういう原子的な個体と見なすのは妥当ではないように感じる。というのも、ヒトは家族のなかに生れ、周りの人々と親密な関係をもつことを通じてヒト社会の一員となるからです。他者との協力(cooperation)や協調(coordination)は、この親密な関係性のなかで始まって、徐々にその範囲を広げていく。それなのに、原子のような個体をまず仮定してモデルを作るのでは、出発点からヒトではない何か別のモノの話をすることになりはしないか。これが、多少とも理解できた部分について、私が全面的には同意できないと感じる理由です。*
注*: 別の言い方をすると、相手の気持ちや周囲の期待と独立の〝利他性〟や〝協力〟というのは、概念的に成り立つのだろうか、という疑問が生まれる。そういう関係は、生物学的には、むしろたんなる〝共生(symbiosis)〟であって、〝協力(cooperation)〟とは言えないのじゃないか。
3.151. しかし、この種の仮定は、力学の初歩で摩擦も空気抵抗もない状態を仮定するのと同じだから、いちいち引っかかるのは数学的定式化に対する〝筋の悪い〟疑問の抱き方というものかもしれない。とはいえ疑問は疑問なので、なんらかの対処が必要になります。どうしたらよいか。
3.152. 対処の方法は二つある。一つは、ボウルズとギンタスの議論を細部まで理解すること。非現実的な原子的個体の仮定から出発しても、より現実的なホモ属の個体の行動様式を再現できるよう、あとから条件を追加することはできる。そういう追加の条件を見つけて、その有効性を吟味すればよい。遺伝子と文化の共進化とか、戦争による〝偏狭な利他性(parochial altruism)〟の助長とか、あるいは、お互いの評判の伝播とか、それらしき条件は散見される。だから、それらが十分にホモ属の個体を再現できているかどうか吟味すればよい。
もう一つの対処の方法は、原子的個体の仮定を排して、ホモ属の個体は相手によって態度を変えるし、まわりの影響でも態度を変えるという、もっと現実的な設定で協力の進化を記述すること。これは、数学的定式化を排除はしないものの、おそらく考古学、人類学、心理学、歴史社会学などを主とする記述になる。
どちらも一朝一夕にはできないので、暗礁に乗り上げた感じがするわけです。
3.153. 今回は、そういう次第で、上述のような原子的個体の仮定(というのは、私の命名ですが)をボウルズとギンタスは本当に導入しているのかどうかを論じていきます。原子的個体を仮定する、とかれらが明言しているわけではないので、そんなもの導入していないということになったら、私の疑問はまるっきり見当はずれになる。だから、導入しているかどうか確かめる必要がある。この議論をするなかで、協力の進化の数学的定式化を紹介し、検討していきます。
3.154. 先に見た(3.139)式4.2への注釈で、ボウルズとギンタスは、問題の仮定を導入している。この原子的個体の仮定は、その後の数学的定式化の決定的なところで重要な役割を果たします(3.175、3.178)。式4.2とは、以下の式でした。
Wij = βo+Pj·βg+Pij·βi (式4.2)
それぞれの項の意味を、もう一度確認します。
① Wijは、〈グループ〉j の個体 i の適応度の期待値。したがって、式4.2は、ある〈グループ〉の任意の個体の適応度の計算方式を表している。
② βoは、周囲に利他的個体のいるいないかかわらず成立する基礎的な適応度。おそらく、自然環境その他もろもろの条件から決まっている。この値は今後とくに問題にならない。
③ Pj は、〈グループ〉j 内の利他性の遺伝子A の頻度。同じことだが、〈グループ〉j 内の利他的個体A の割合。また、〈グループ〉j 内の任意の個体が利他的である確率といってもよいはず。
④ βg は、訳者の補足説明によれば、事実上、利他行動がもたらす利益 bと見なしてよい(3.142)。そう見なしてよい理由のかなめの部分に、原子的個体の仮定がかかわっている(3.178~3.181)。しかし、ボウルズとギンタス自身の正式な説明では、βg は「Pj(〈グループ〉j内の遺伝子A の頻度)の変化が Wij へ及ぼす効果(effect)」(ボウルズ&ギンタス2017, 99)となっている。以下でまず扱うのは、正式な説明の方です。
⑤ 式4.2の右辺第2項「+Pj·βg」は、個体iには、自分の属す〈グループ〉jに利他的個体がいる割合に応じて βg なる適応度上の利益が得られることを示している。
⑥ Pij は、個体 i が利他的個体 A ならば 1、非利他的個体 N ならば 0 の値をとる変項。
⑦ βi は、これも訳者の補足説明によれば、利他行動のコスト-c と見なしてよい。そして、そう見なしてよい理由のかなめの部分に、やはり原子的個体の仮定がかかわる(3.178~3.181)。しかし、ボウルズとギンタス自身の正式な説明では、βiは「Pij(個体 i が遺伝子 A を持つかどうか)の変化が Wij へ及ぼす効果(effect)」(ボウルズ&ギンタス2017, 99)となっている。これについても、以下でまず扱うのは正式な説明の方です。
⑧ 式4.2の右辺第3項「+Pij·βi」は、個体iは自分が利他的個体なら βi なる利他行動のコストの分だけ適応度上の不利益を被るが、非利他的個体ならばコストは発生しない、ということを示している。
⑨ 式4.2の趣旨は、以下のとおり。
〈グループ〉j の個体 i の適応度の期待値=基礎的な適応度+自分の属す〈グループ〉に利他的個体がいる割合に応じた適応度上の利益+(利他行動のコスト分の不利益または 0)
3.155. 以上の考え方は、理に適っているように見えます。個体が生涯に育てることができる子供の数(適応度)は、自然環境やその動物種の繁殖の類型で決まる基礎的な数に加えて、周囲が協力してくれれば、それだけ繁殖に有利になるから数が増え、かつ、自分がコストをかけて他個体に協力すれば、そのコストの分だけ不利になって数は減る。だから、基礎的な数値と増減分の数値を合計すれば、個体の適応度は予測できる。私が同意できないと感じるのは、協力が起こるかどうかが、相手や周囲の状況とまったく無関係に、個体の初期設定だけで決定される、という一点です。
3.156. 問題の、原子的個体の仮定は、ボウルズとギンタスが式4.2について与えた説明のなかに現れます。
3.157. ボウルズとギンタスは、式4.2、Wij = βo+Pj·βg+Pij·βi について、以下のように述べています。この一節は、式4.2に関するなにげない注釈にしか見えない。けれども、私には、原子的個体の仮定を導入する決定的な言葉のように見える。なお、〔…〕内は、訳者が親切心から補足してくれた説明。読解の手がかりとして役に立ちます。
「式4.2では〈グループ〉間および〈グループ〉内淘汰の2つの効果(effects)が分離可能であるので、〈グループ〉が個体に及ぼす影響の大きさとその個体が利他的であるかどうかは独立であり〔βg と Pij は関係がないという意〕、利他的個体であることが及ぼす個人への効果は〈グループ〉中の利他的個体の頻度と独立である〔βi と Pj は関係がないという意〕。これら2つの効果が分離可能であるという仮定は現実的ではない(unrealistic)が、このことで明確な単純化が図れる。」(ボウルズ&ギンタス2017, 99)
3.158. これだけ読んでも何が言いたいのかよく分からない。だが、問題の焦点が「分離可能」という点らしい、ということは一応分かります。主張の骨格を抜き書きしておきます。
⑩ 式4.2では、「2つの効果が分離可能」なので、
⑪ 「〈グループ〉が個体に及ぼす影響の大きさとその個体が利他的であるかどうかは独立であり」
⑫ 「利他的個体であることが及ぼす個人への効果は〈グループ〉中の利他的個体の頻度と独立である」
⑬ ただし、「2つの効果が分離可能であるという仮定は現実的ではない」のだが、これによって「単純化」を図ることができる。
こう言われている。
3.159. 明かにすべきは、第一に、⑩について、分離可能なのは何と何か。第二に、⑪と⑫で、独立であるのはいったい何と何であり、それらが独立であることから何がもたらされるのか。第三に、⑬について、分離可能であるという仮定が現実的でないのはどうしてか。第四に、同じく⑬について、何の単純化がどのように成立するのか。
3.160. 第一に、⑩について、分離可能なのは、もちろん「〈グループ〉間および〈グループ〉内淘汰の2つの効果(effects)」です。「効果」という言葉はすでに使われていた。3.148の④と⑦で引用したように、βg とβi がそれぞれ「効果」と呼ばれている。分離可能なのは、βg とβi というわけ。そして、βg は、〈グループ〉内のA遺伝子の頻度Pj が、個体 i の適応度の期待値Wij へ及ぼす「効果」である。βi は、個体 i が遺伝子A を持つかどうかの変化Pij が、同じくWij へ及ぼす「効果」である。この2つの効果が分離可能であるといわれている。
3.161. 依然として、分かりにくい。だが、この分かりにくさは、ある程度まで、“effect”を「効果」と訳したことに由来しています。 “effect”は、この文脈では、「効果」よりも「力」あるいは「影響」と訳した方が文意がよく通じる。
3.162. 日本語の「効果」は、多くの場合、なんらかの作用の〝結果〟を意味する。ところが、オックスフォード英語辞典(OED)の“effect”の項の説明を見ると、英語の“effect”は作用する力とその影響の繋がった〝状態〟を言い、〝作用の流れ〟を含意するように読める。「力」や「影響」を"effect"の訳語としてもさしつかえない。
3.163. それゆえ、βg は、「遺伝子 A の頻度 Pj の変化が個体 i の適応度の期待値に及ぼす〝力〟ないし〝影響〟」である。同じく、βiも、「個体が遺伝子 A を持つかどうかという Pij の変化が適応度の期待値に及ぼす〝力〟ないし〝影響〟」である。
3.164. というわけで、分離可能というのは、(ア)〈グループ〉内に利他性の遺伝子 A の持ち主がどのくらいの割合でいるかということが個体の適応度に及ぼす影響と、(イ)ある個体が利他的 A であるか非利他的 N であるかの違いが個体の適応度に及ぼす影響が、分離可能だということを言っている。
3.165. 具体的に言うと、(ア)は、利他性の遺伝子 A の持ち主が〈グループ〉内に多いと、個体の適応度は高くなり、少ないと低くなるということを言う。これは〈グループ〉間の違いによって、個体の適応度に変化が生じるということを言っている。よって、たしかに、これは〈グループ〉間淘汰の影響(「効果」)である。
3.166. (イ)は、個体が利他的であると利他行動のコストの分だけ適応度が低くなり、非利他的であるとコストを支払わない分だけ高くなるということを言う。これは〈グループ〉内の個体間の相違によって、それぞれの個体の適応度に変化が生じるということを言っている。よって、たしかに、これが〈グループ〉内淘汰の影響(「効果」)である。
3.167. 結局、ボウルズとギンタスは、〈グループ〉内の遺伝子 A の頻度 Pj の影響力と、〈グループ〉の各個体が利他的 A であるか非利他的 N であるかという Pij の影響力は、式4.2では、分離できるようになっている、と言っているわけである。
3.168. このよう分離できるとどうなるのか。⑪と⑫がもたらされる。まず⑪について。⑪の「〈グループ〉が個体に及ぼす影響の大きさとその個体が利他的であるかどうかは独立であり」とは、βg と Pij が関係がないということだ、と訳者が補足説明してくれています。βgすなわち「Pj(〈グループ〉j内の遺伝子 A の頻度)の変化が Wij へ及ぼす力ないし影響」(3.151④)と、Pij すなわち、その個体 i が利他的 A なのか非利他的 N なのか(3.151⑥)は、関係ないということ。これはいったいどういうことか。
3.169. 〈グループ〉内に利他的個体が多いと、当然、その〈グループ〉内の個体の適応度上の利得は多くなる。利他的個体が少ないと、利得は少なくなる。そして、この利得の増減は、その個体が利他的 A であっても非利他的 N であっても、それとは独立に、つまり無関係に、成り立つ。こう言っているように読める。もっと具体的に言うと、〈グループ〉内のどの個体も〈グループ〉で利他的個体が占める割合に応じた適応度を得る、と言っていると解釈できる。
ということは、裏を返せば〈グループ〉内の利他的個体は、自分の遭遇した相手が利他的個体であってもなくても、態度を変えることなく、つねに利他的に振る舞って相手方の適応度に貢献していることになる。もちろん、非利他的個体も相手の出方に応じて態度を変えたりはしない。というわけで、この⑪の述べていることが、原子的個体の仮定の前半部、それぞれの個体は「協力するかしないか相手の出方で態度を変えたりしない」に該当します。
3.170. ⑫の「利他的個体であることが及ぼす個人への効果は〈グループ〉中の利他的個体の頻度と独立である」とは、βi と Pj は関係がないということだ、とこれまた訳者が補足説明してくれている。βi すなわち「Pij(個体 i が遺伝子 A を持つかどうか)の変化が Wij へ及ぼす力ないし影響」(3.151⑦)と、Pj すなわち、〈グループ〉j 内の利他性の遺伝子 A の頻度(3.151③)とが独立ということ。いったいどういうことか。
3.171. 個体 i が利他的 A であると、利他行動のコストを引き受けなければならない。個体 i が利他的 A であるとき、Pij = 1であるから、Pij·βi = βiであって、つまり βi だけのコストがかかる。利他的でない N の場合は、Pij = 0であるから、利他行動のコストは発生せず、Pij·βi = 0である。この βi または 0 という値が、Pij が個体 i の適応度の期待値に及ぼす影響である。この影響が、Pj すなわち〈グループ〉j 内の利他性の遺伝子 A の頻度(3.151③)と関係がないとは、〈グループ〉内に利他的個体が多かろうが少なかろうが、個体 i が A か N かに応じて βi または 0 という値のコストが発生する、ということだと解釈できる。
3.172. わかりやすく言えば、〈グループ〉内に利他的個体が多いか少ないかということと、それぞれの個体が利他的か非利他的かは連動しない、ということ。具体的に言うと、利他的個体が多いと、各個体が利他的になる(大勢順応)ということはないし、逆に非利他的になる(あまのじゃく)ということもない。利他的個体が少ない場合も、個体が非利他的になる(大勢順応)ということはないし、逆に利他的になる(あまのじゃく)ということもない。〈グループ〉内の利他遺伝子 A の頻度は、個体が利他的であるか否かに影響を及ぼさない。独立とはこのことだと解釈できる。
というわけで、この⑫の述べていることが、原子的個体の仮定の後半部、それぞれの個体は「まわりの個体たちの影響で協力したりしなかったり態度を変えることもない」に該当します。
3.173. こうして、⑩のとおり分離可能であることから、⑪と⑫が帰結する。つまり、原子的個体の仮定が帰結するということになります。
3.174. 以上のように解釈してみると、⑬の「2つの効果が分離可能であるという仮定は現実的ではない」の趣旨は、容易にわかる。現実の人間は、相手が協力的か非協力的かに応じて態度を変えるし、まわりが利他行動を頻繁に行なっている環境では、影響されて自分も利他的に振る舞うようになったり、あるいは逆にあまのじゃくに徹したりするだろう。だから、相手がどうあれ、また、周囲がどうあれ、利他的素質を持つものは利他的に振る舞い、非利他的素質を持つものは非利他的に振る舞う、という仮定は非現実的なのだ。
だが、これによって、大いに価値のある「単純化」を図ることができる。すなわち、この分離可能性の仮定の下で、プライス方程式という定式を使えるようになるのです。
「Price(1972)*は、この場合において〈集団〉における利他的個体の頻度の変化であるΔP≡P′-Pは、〈グループ〉間および〈グループ〉内効果に分割できることを示した。」(ボウルズ&ギンタス2017, 99)
注*: Price, G. R. (1972). Extension of Covariance Selection Mathematics. Annals of Human Genetics 35: 485-490.
3.175. 「この場合において(in this case)」というのは、〝〈グループ〉間淘汰の効果と〈グループ〉内淘汰の効果が分離可能な場合において〟ということ。だから、これが「この2つの効果が分離可能であるという仮定」(ボウルズ&ギンタス2017, 99)が決定的な役を果たす第一の局面です。そして、この分離可能性の仮定は、ここまでの解釈に即していえば、原子的個体の仮定と同じことです。
3.176. プライス方程式は、個別の〈グループ〉ではなく〈集団〉全体を視野に入れて利他的個体の頻度の変化を扱うために導入される。ボウルズとギンタスは、プライス方程式の導出を付録のA10節で行なっています。その結果、得られるのが次の式です。
WΔP = βG·var(Pj)+βi·var(Pij) (式4.3)(ボウルズ&ギンタス2017, 99)
まず、記号の説明をします。
⑭ Wは、〈集団〉全体の平均適応度。なお、ボウルズとギンタスは、便宜的にこれをW = 1と規約して、計算過程から外します。
⑮ ΔP は、〈集団〉のある世代と次の世代の利他遺伝子 A の割合の差。ある世代の開始時の〈集団〉の利他遺伝子 A の割合を P とし、次の世代の開始時における〈集団〉のAの割合を P′ で表わすとき、ΔP は P′-P のこと。ΔP = P′-P > 0 ならば、利他遺伝子 A はその〈集団〉で割合を増している。元々、この値が正になる条件を求めることが、利他遺伝子 A の進化を数学的に定式化することの関心事でした(3.138参照)。
⑯ βGは、β×Gではなく、βGで一個の記号です。Gは本当は下付き文字ですが、仕様上、下付きにできないので仕方がない。βG は、Pj の変化が〈グループ〉の個体の平均適応度 Wj に及ぼす影響(邦訳では「効果」)を表す(ボウルズ&ギンタス2017, 104)。Pj は、再三言うように、〈グループ〉j の利他遺伝子 A の頻度です。〈グループ〉における利他的個体 A の割合が違うのに応じて、その〈グループ〉の個体の平均適応度は違ってくる。その Pj の違いが平均適応度 Wj に及ぼす影響、ないし力を、βG が表わしている。
⑰ var(Pj)は、Pj の分散。ある量 X の分散とは、X と X の平均 Xとの差を2乗した Y = (X-X)^2 という新しい値 Y に関する平均値。ここでは Pj が〈グループ〉j の遺伝子 A の頻度だから、まず Pj の平均値を考える。するとこれは要するに、〈集団〉全体の遺伝子 A の頻度 P のことなので、Pと Pj の差の2乗を〈グループ〉の大きさで重み付けした平均値が var(Pj) ということになります(下の式参照)。なお下の式の fj は、〈グループ〉の大きさ、つまり〈集団〉の全個体数に対する〈グループ〉j の個体数の割合を表します。これが重み付けの係数になる。
この var(Pj) は、各〈グループ〉の遺伝子A の頻度の、〈集団〉全体におけるばらつき具合を表わす統計量です。
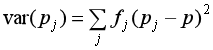
⑱ βi は、⑦で既出。くり返しておけば、「Pij(個体 i が遺伝子 A を持つかどうか)の変化が Wj へ及ぼす力ないし影響(effect)」
⑲ var(Pij) は、まず、var(Pij) が Pij の分散だから、var(Pij)は、 var(Pij) の重み付き平均。下の式で与えられる。なお、下の式の fj は、⑰と同じく〈集団〉の全個体数に対する〈グループ〉jの個体数の割合を表す。 var(Pij) については、3.186の説明を参照のこと。
![]()
3.177. ⑯の βG は新しい値なので、どういう値なのか、計算しておかないといけない。βG は、Pj の変化が〈グループ〉の個体の平均適応度 Wj に及ぼす影響なので、まず Wj を求める必要がある。Wj は、〈グループ〉j の各個体の期待される適応度 Wij の平均値なので、Wij から求めることができる。Wij を全部足して〈グループ〉j の個体数で割ればよい。Wij は式4.2で表わされているので、これを全部足して個体数で割って整理すると、次のようになる。
Wj = βo+Pj(βg+βi)(式4.5) (ボウルズ&ギンタス2017, 104)
そして、βGとは、Pj の変化に応じて Wj が変化する度合いということなので、式4.5を Pj について微分すればよい。すると βG = βg+βi となる。
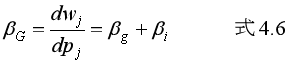
3.178. 式4.6を使って、式4.3を見通しのよい定式に書き換えることが試みられるわけです。ここで、分離可能性の仮定、つまりは原子的個体の仮定が、再び決定的な役割を果たす。ボウルズとギンタスは次のように言います。
「分離可能性の仮定によって式4.3を利得の言葉で表現する事が可能になる。それは βi = -c と βg = bである。したがってこれと式4.6を用い、W=1であることを思いだせば、式4.3 は
ΔP = (b-c) var(Pj)-c·var(Pij) (式4.7)
と書くことができる。」(ボウルズ&ギンタス2017, 104)
3.179. 分離可能性の仮定は、ここでは、βi を-c、つまり利他行動のコストと見なし、βg を b、すなわち利他行動の利益と見なすことを可能にしている。βi と βg は、何度でも言いますが、それぞれ、利他的個体であるか否かが個体の適応度に及ぼす影響と、〈グループ〉内の利他的個体の頻度が個体の適応度に及ぼす影響、でした。この2つの影響が分離できるので、利他的個体であるか否かの及ぼす影響 βi と利他的個体の頻度 Pj は独立になる。また、利他的個体の頻度の及ぼす影響 βg と利他的個体であるか否か Pij は独立になる。上述(3.169~3.173)の解釈によれば、これは、ある個体が利他的か非利他的かは〈グループ〉内に利他的個体がどれくらいいるかに左右されず、ある個体が利他的に振る舞うか否かは相手が利他的か非利他的かに左右されない、という原子的個体の仮定と同等である。
3.180. まだ分からないのは、この分離可能性の仮定(原子的個体の仮定)から、どうして βi = -c、βg = b という値が出てくるのか、という点です。上の引用でボウルズとギンタスは、それは自明であるかのように述べているけれど、私には自明ではない。訳者による「コラム2:プライス方程式とは」(ボウルズ&ギンタス2017, 100-103)を参照すると、以下の手続きで βi = -c と βg = b が導き出されている。この過程のどこかに、分離可能性の仮定がかかわっているはずです。
3.181. まず、以下のようにモデルを設定する。これは、ボウルズとギンタスが式 4.2を定式化する際の記述(3.135~3.144参照)に沿ったモデルです。
「自分を含む〈グループ〉全体に利益を与える協力行動を考える。利他遺伝子 A の保持者はコスト c を支払い、……〈グループ〉中に n 個体がいるならば、自分を含めた〈グループ〉全員にそれぞれ b/n の利益を与える協力をする。(期待値を考えるので、「自分を含む〈グループ〉の誰か1人をランダムに選び、その人だけに集中的に利益 b を与える」としても結果は変わらない)」(ボウルズ&ギンタス2017, 101)
〈グループ〉j には利他遺伝子 A を保持する個体が k 個体いるとして、このモデルで、〈グループ〉j にいる i さんの適応度を考える。
i さんが A を持っていない場合、基礎的適応度 βo に加えて、b/n の利益を k 個体から得られ、かつ、自分は利他行動のコスト c を負担しないわけだから、i さんの適応度は次のように表わすことができる。
Wij = βo+(kb/n)
i さんが A を持っている場合、利他行動のコスト c を負担するから、i さんの適応度は次の通りとなる。
Wij = βo+(kb/n)-c
上の2つの式をひとまとめに記せば、
Wij = βo+(kb/n)-(cもしくは0)
これを整理すると、
Wij = βo+b(k/n)-c(iさんがAを持てば1、そうでなければ0)
ここで、k/n とは、〈グループ〉j 内のA遺伝子の頻度だから、これは Pj であり、「i さんが A を持てば1、そうでなければ 0」とは、Pij にほかならない。そこで、上の式を Pj と Pij を使って書き換えれば、
Wij = βo+b·Pj-c·Pij
一方、式4.2は、Wij = βo+Pj·βg+Pij·βi だった。
したがって、ボウルズ&ギンタス(2017)に則したモデルにおいては、βg = b であり、βi = -c であることになる。
3.182. 式の変形過程は、暗黙裡に式4.2を下敷きにしているから、すでに分離可能性の仮定の下での操作になっています。だから、式変形に先立つ引用部分が分離可能性の仮定の実質を与えている。実質は、〝利他遺伝子 A の保持者がコスト c を支払って、〈グループ〉内の各個体に b/n の利益を与える〟という部分にある。そして、この設定が、原子的個体の仮定を暗黙に前提していることは明らかです。
この設定が告げているのは以下のことです。すなわち、利他遺伝子 A の保持者は、相手が利他的 A であれ非利他的 N であれ、〈グループ〉の全員に無差別に b/n の利益を与える。また、任意の個体が協力行動をするか否かは、〈グループ〉内の Aを 持つ個体の頻度 k/n と無関係である。これは、原始的個体の仮定そのものです。
この仮定を、既知の記号で言い表すと、Pij の如何に関わらず βg は影響を及ぼすのであり、βi の影響は Pj と無関係である。これは同一の設定を、分離可能性の言葉で表現したものになっている。
こういう次第で、「分離可能性の仮定によって式4.3を利得の言葉で表現する事が可能になる。それは βi = -c と βg = b である」(ボウルズ&ギンタス2017, 104)と主張できるわけです。
3.183. 以上から、ボウルズとギンタスは、原子的個体の仮定をたしかに導入しており、それは数学的定式化において、プライス方程式を導入する局面(3.175)と、プライス方程式を利得の言葉で表現する局面(3.178)で、不可欠の役割を果たしている、と言い得るでしょう。
3.184. では、そもそもプライス方程式とは何を告げているのか。式4.7の各項を読み解いてみます。
3.185. 式4.7、ΔP = (b-c) var(Pj)-c·var(Pij) において、右辺第1項の (b-c) var(Pj) は、利他行動の利得とコストの差 (b-c) に、各〈グループ〉の利他遺伝子 A の頻度の〈集団〉全体でのばらつき var(Pj) を掛け合わせた値になっている。b-c > 0と置いたから(3.136)、この第2項は正の値をとる(分散は負にはならない)。
したがって、〈集団〉を見わたした時に各〈グループ〉の利他遺伝子の頻度のばらつき var(Pj) の値が大きいと、右辺第1項の値は大きくなる。〈グループ〉ごとの利他遺伝子の頻度の、〈集団〉全体でのばらつきが大きいとは、〈グループ〉のあいだで成員に占める利他的個体の割合が大きく異なっている状態、具体的に言えば、いくつかの〈グループ〉に利他的個体が偏って所属している状態を意味する。要は、ある〈グループ〉には利他的個体がたくさんいるが、別の〈グループ〉にはあまりいない状態です。右辺第1項は、したがって、〈グループ〉間の違い(ばらつき)の影響力(〈グループ〉間淘汰の効果)を表現している。
3.186. 式4.7の右辺第2項の-c·var(Pij) は、利他行動のコスト-c に、各個体が利他的 A か非利他的 N か(即ち、Pij)に関する〈グループ〉内の分散の〈集団〉全体での平均値 var(Pij) を掛けた値になっている。
まず、Pij の〈グループ〉内の分散 var(Pij) とは、Pij 自体と Pij の平均との差の2乗という新しい値に関する〈グループ〉内の平均のこと。Pijは1または 0 の値をとる。そして、Pij の平均とは、〈グループ〉j 内の任意の個体 i が 1(個体iは A をもつ)なのか 0(個体iは A をもたない)なのかの期待値だから、これは Pj である。したがって、Pij の〈グループ〉内の分散var(Pij)とは、(Pij-Pj)^2を i についてすべて足し合わせて〈グループ〉の個体数で割った値になる。
var(Pij) は、上の var(Pij) の、〈集団〉を見わたした時の平均です(3.176⑲)。〈グループ〉内におけるA個体とN個体のばらつき具合の〈集団〉全体での平均値ということ。
分散は負にならないから、分散の平均値も負にならない。すると、-c·var(Pij) は常に負になる。そして、〈グループ〉内における A 個体と N 個体の分散の平均値が大きいほど、-c·var(Pij) は大きな負の値になる。A 個体と N 個体の分散の平均値が大きいというのは、それぞれの〈グループ〉内が A 個体にも N 個体にも偏っていない(ばらついている)ということを意味している。そういう場合に、全体としての適応度は減る方向の力を受けるわけである。
というわけで、式4.7の右辺第3項の -c·var(Pij) は、〈グループ〉内の個体が A か N かという違い(ばらつき)の影響(〈グループ〉内淘汰の効果)を表わしている。
3.187. したがって、プライス方程式、即ち ΔP = (b-c) var(Pj)-c·var(Pij)(式4.7)の告げていることは、全体としては、以下のとおりになる。
〈集団〉の利他遺伝子の頻度の世代間での差 ΔP は、利他的個体が各〈グループ〉内に占める割合の〈グループ〉間でのばらつきが大きいと――いくつかの〈グループ〉に利他的個体がかたまって所属していると――右辺第1項の効果で増大する方向に動き、〈グループ〉内で利他的個体と非利他的個体が混ざって――どちらかに偏らずばらついて――いれば、右辺第2項の効果で減少する方向に動く。これは、先に述べた「ややこしい状態」(3.133)の、数式による表現になっている。
3.188. プライス方程式を導入し、それを利得の言葉で表現することによって、複数レベル淘汰における利他性の進化の条件が表現できるようになる。その手順を、簡単にまとめておきます。といっても、私の理解には大きな限界があって、その手順のカナメの部分を〝いったいどうして〟ボウルズとギンタスが発見できたのかが分からない。とにかく、ボウルズとギンタスは、まずもって、次のような比を定義する。
「分散比 Fst を、利他的個体の頻度の〈グループ〉間分散の、〈集団〉の全分散、すなわち利他的個体の頻度の〈グループ〉内分散と〈グループ〉間分散の和、に対する比として定義しよう。言い換えれば、
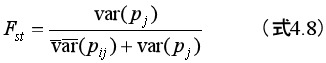
である。」(ボウルズ&ギンタス2017, 105)
3.189. 私に分からないのは、まさにこの分散比を組み立てることを、どうしてボウルズとギンタスが思いついたのか、という〝ことの次第〟です。こういう分散比を作って分析するということは、統計に関わる常識的で基本的な手順の一つなのか? それとも、なにか天才的なひらめきなのか? たぶん、前者だろうが、統計処理にくらい私には、なぜこの形式の比が重要な意味をもつと分かったのか、ということが分からない。というのも、この分散比 Fst はこれだけ見ても意味不明ですが、下の式4.9のように書き表すことができて、その重要性がわかるからです。
![]()
式4.8と式4.9は同じことを言っている*。なぜ、こんなかけ離れた表現の式が同じ値をもつと、ボウルズとギンタスは洞察できたのだろうか。それが私には分からない。
注*: ボウルズとギンタスは、式4.9を証明抜きで天下り的に提示しているので、訳者が「コラム3」(ボウルズ&ギンタス2017, 107-111)で、式4.8と式4.9が同値であることを丁寧に説明している。それぞれの式を変形すると同じ値に帰着する。
3.190. 式4.9の趣旨を説明する。まず Prob(X|Y) は、「Yを条件としたXの確率」と読む。条件付き確率(conditional probability)の記号は、通常 P(X|Y) だが、本稿では、大文字「P」をすでに遺伝子Aの頻度を表す記号 P、Pj、Pij などとして使用したので、混乱を避けるため Prob(X|Y) とした。AとNはこれまで通り、ある個体が利他的であること(A)と非利他的であること(N)を表わす。すると、式4.9は、とりあえず記号だけ読めば、以下のことを述べている。
分散比 Fst は、利他的個体 A を条件としたときの利他的個体 A の確率から、非利他的個体 N を条件としたときの利他的個体 A の確率を、差し引いた値である。
3.191. ボウルズとギンタスは、言葉を補って、これを分かりやすく説明している。
「この比〔Fst〕は、自らが利他的個体である場合に利他的個体とペアを組む条件付き確率 Prob(A|A) と、自らが利他的個体でない場合にそのようになる〔利他的個体とペアを組む〕条件付き確率 Prob(A|N) の差を測って〔いる〕」(ボウルズ&ギンタス2017, 105. ただし〔…〕内は筆者の補足)
式4.9は、Fst なる分散比を、利他的個体 A が利他的個体 A と出会う確率から、非利他的個体 N が利他的個体 A と出会う確率を引いた値、として提示しているということ。利他的個体 A 同士が出会う現象は、
「自身の〈グループ〉に多くのAがいる時により起こりやすい。もし、Aである個体が他にもたくさんAがいる〈グループ〉に所属しやすいならば、AはNよりも有利である」(ボウルズ&ギンタス2017, 105)
そして、Fst が「この有利度の指標」(同上)となっているのである。
3.192. AとAが出会う確率が、NがAと出会う確率よりも、かなり高い場合、Aは沢山かたまって幾つかの〈グループ〉に属しており、Nはそれらとは別のAが稀な〈グループ〉に属しているはずである。〈集団〉がそういう構造をもっているとき、ΔP を表わす式4.7は、右辺の第1項が大きな値となり、第2項は小さな値となる(3.186)。つまり、ΔPは正になりうる。ΔPが正の値をとるということは、〈集団〉内で利他遺伝子Aの頻度が増しているということにほかならない。協力が進化しているのである。分散比 Fst はその指標になっている。
3.193. ΔP = 0 のとき、利他性の遺伝子 A は、世代をまたいで増えても減ってもいない。そこで、式4.7で、ΔP = (b-c) var(Pj)-c·var(Pij) = 0 と置き、式4.8の定義を使って式4.7の項を書き換えて整理する。すると、次の式が得られる。
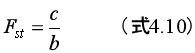
すなわち、Fst = c/b のとき ΔP = 0 であり、〈集団〉において利他的個体Aは増えても減ってもいない。そして、Fst > c/b ならば、利他的個体の割合は増加し、Fst < c/b ならば割合は減少するだろう(ボウルズ&ギンタス2017, 105)。ここから、次のことが分かる。
「式4.10は Fst を〈集団〉構造の指標とした単なるハミルトン則である」(ボウルズ&ギンタス2017, 106)
ここは感動するところ。なんと、瓢箪から駒のように、ハミルトン則が飛び出した。
3.194. 利他性の進化に関するハミルトン則は、協力者と受益者の血縁の平均的な近さ r と、利他行動の利得 b と、利他行動のコスト c の間に、rb > c という関係が成り立っているとき、その〈集団〉において利他的な特性が進化する、という法則である(3.123)。式4.10はもちろん Fst·b > c のとき、利他性が進化することを示しているから、Fst は、一般化された意味の〝血縁度〟ないし近縁度、つまり正の同類性(positive assortment)の指標になっている(3.128)。
3.195. しかし、私には、上に述べた通り、Fst という分散比が、利他的個体同士がペアになる確率と非利他的個体が利他的個体とペアになる確率の差であることを、〝いったいどうして〟ボウルズとギンタスは洞察できたのか、という〝ことの次第〟が分からない。だから、その Fst が正の同類性を示す近縁度 r なのだと告げられても、うまく行ったもんだなあと思うものの、狐につままれたような感じが残る。基本的なモデルの設定(3.135~3.139)を終えたあとは、プライス方程式を導入して数式の変形をしているだけなので、その意味では、得られるべき結果が得られたにすぎない。何がどうしてこうなったのだろう。わからない。いまのところ、わからないったらわからないので、私としては、ボウルズとギンタスはハミルトン則から話を開始して(3.124)、たしかにハミルトン則に沿って利他性が進化する条件を見いだすことができました(3.193)、という確認で話を終えるしかありません。
3.196. これにてとりあえず、その7の冒頭(3.117)で述べた素描の試みを済ませることができたので、次回は、以前から予告(3.88)していた民族誌や考古学のデータにもとづくボウルズ&ギンタス(2017)への異論を紹介したいと思います。
この記事が気に入ったらサポートをしてみませんか?